
Cysteine Biosynthesis in Campylobacter jejuni: Substrate Specificity of CysM and the Dualism of Sulfide

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Heterologous Production and Purification of C. jejuni CysM
2.3. Spectral Analysis of CysM First Half Reaction
2.4. Kinetic Measurements of Cysteine Synthase Activity
2.5. Construction of C. jejuni Mutant Strains
2.6. Construction of C. jejuni Complement Strains
2.7. C. jejuni Growth Assays on Sulfur Limited Media
2.8. Oxidase Activity Measurements by Oxygen Electrode
3. Results
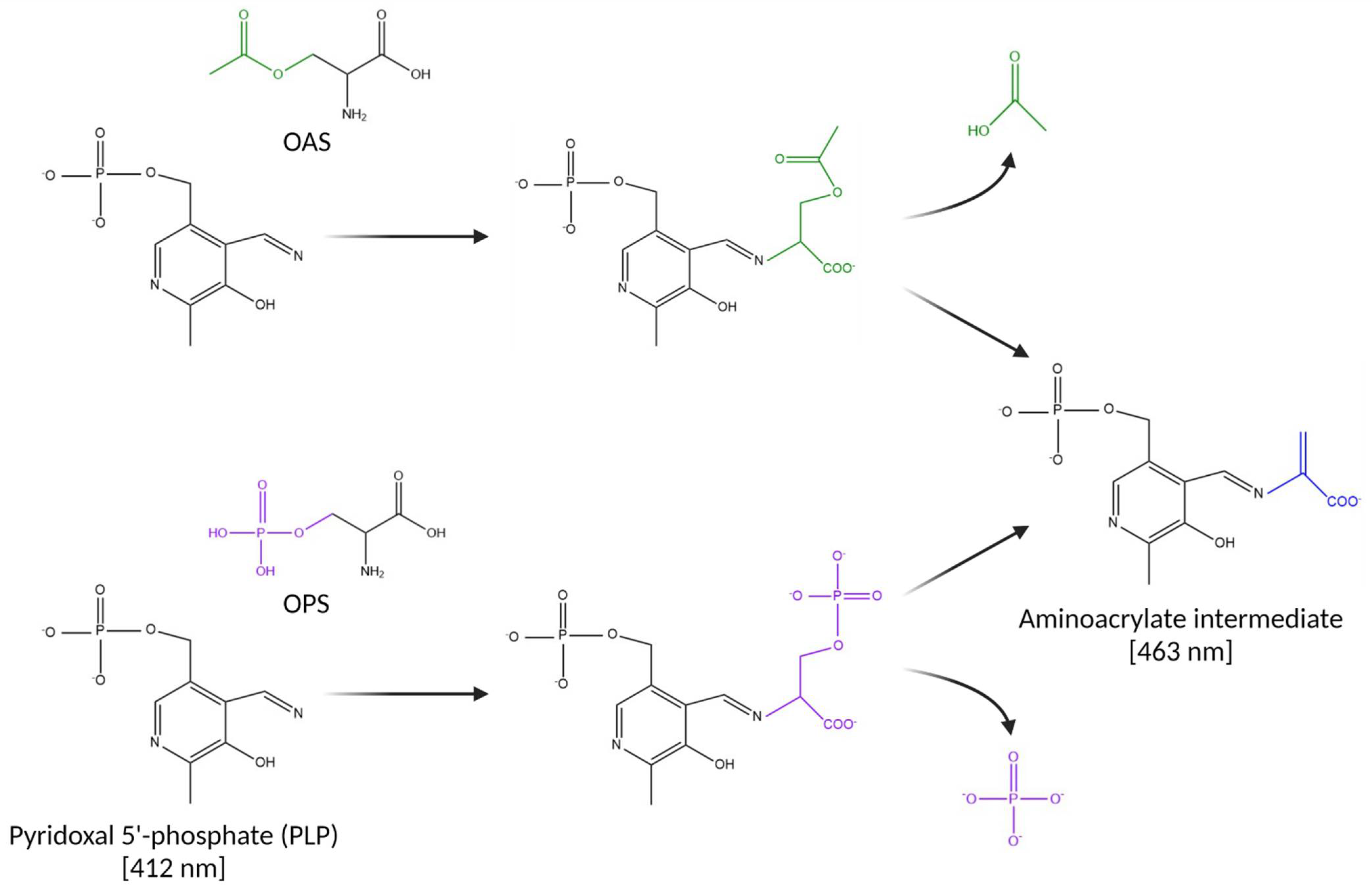
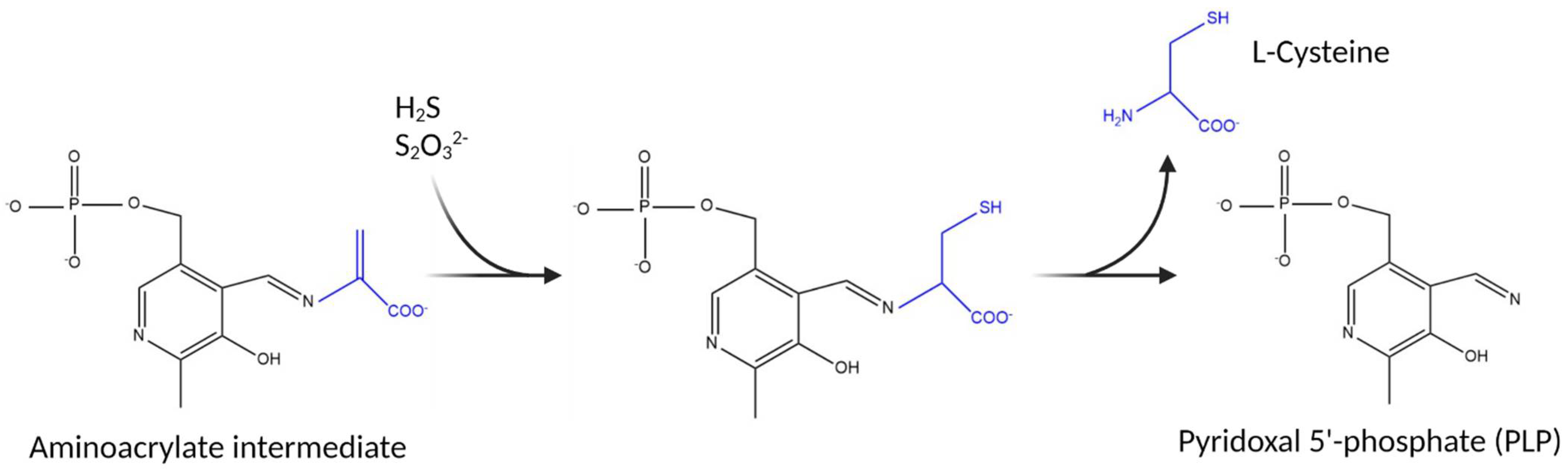
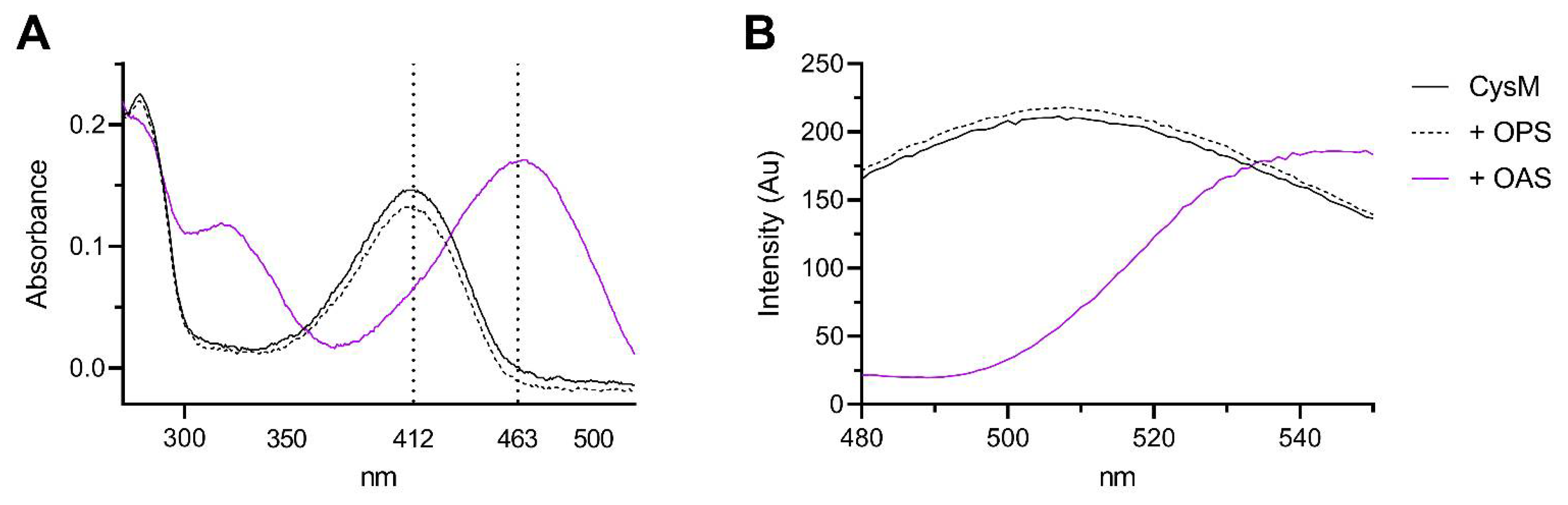
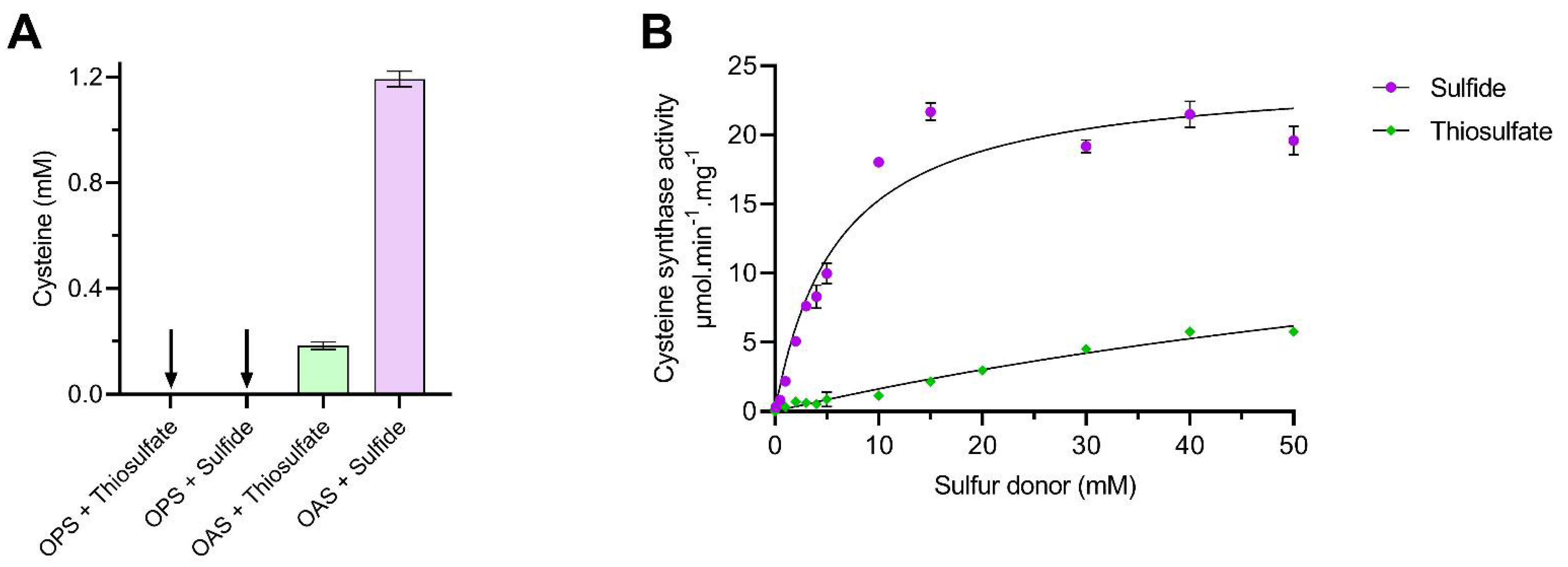
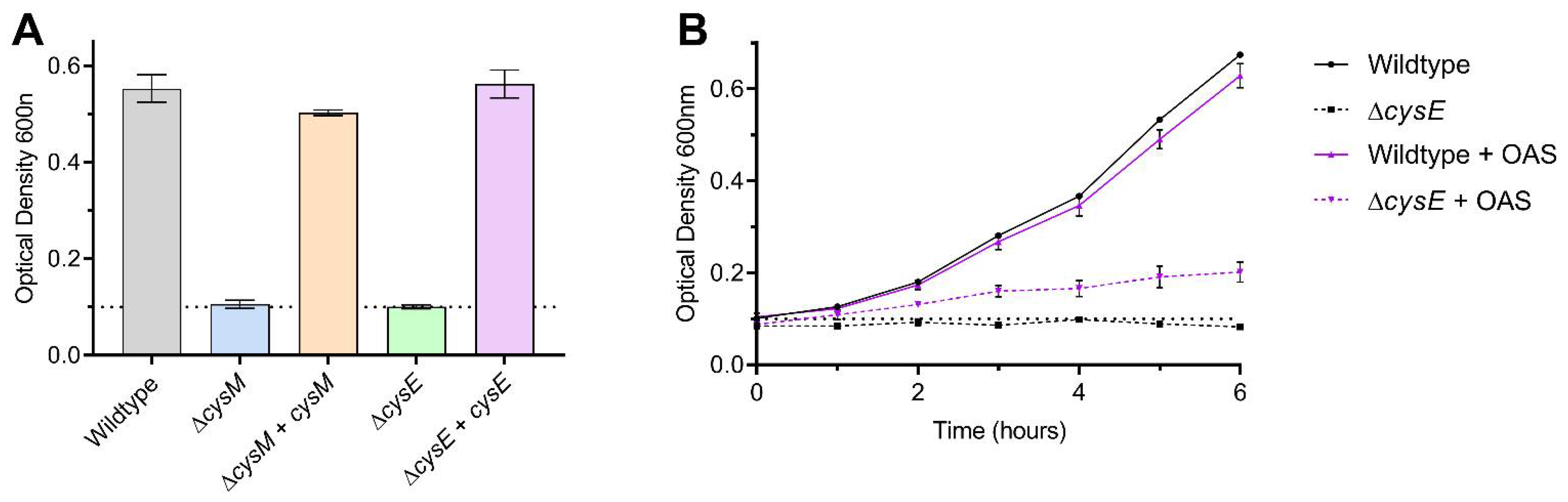
3.1. Substrate Specificity of the C. jejuni Cysteine Synthase, Cj0912
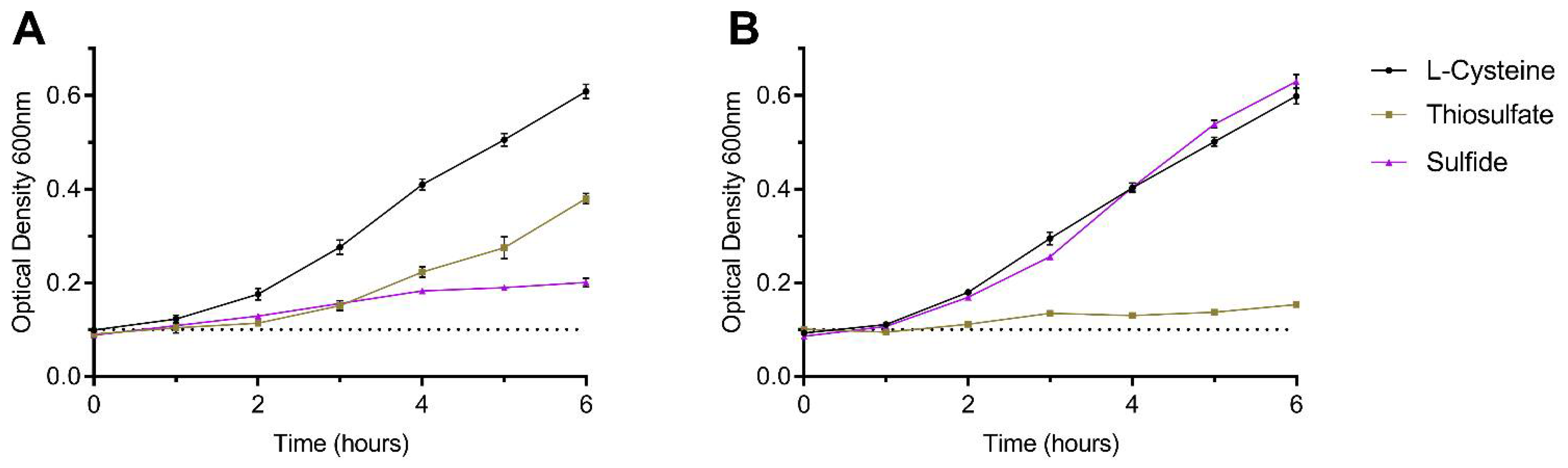
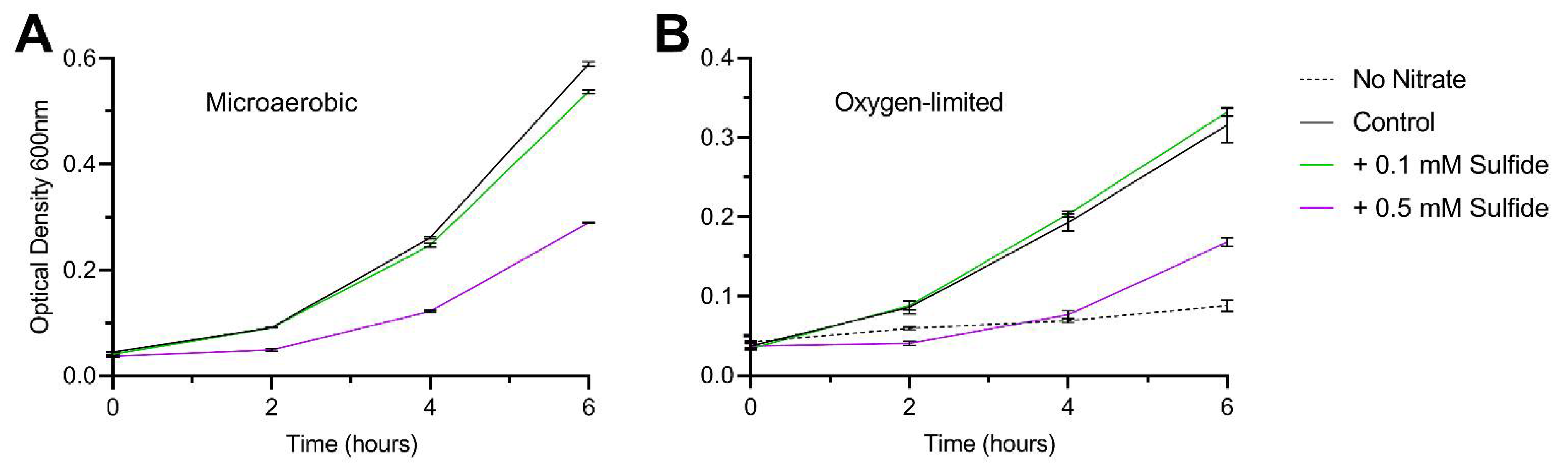
3.2. Sulfide as an Effective Sole Sulfur Source for C. jejuni
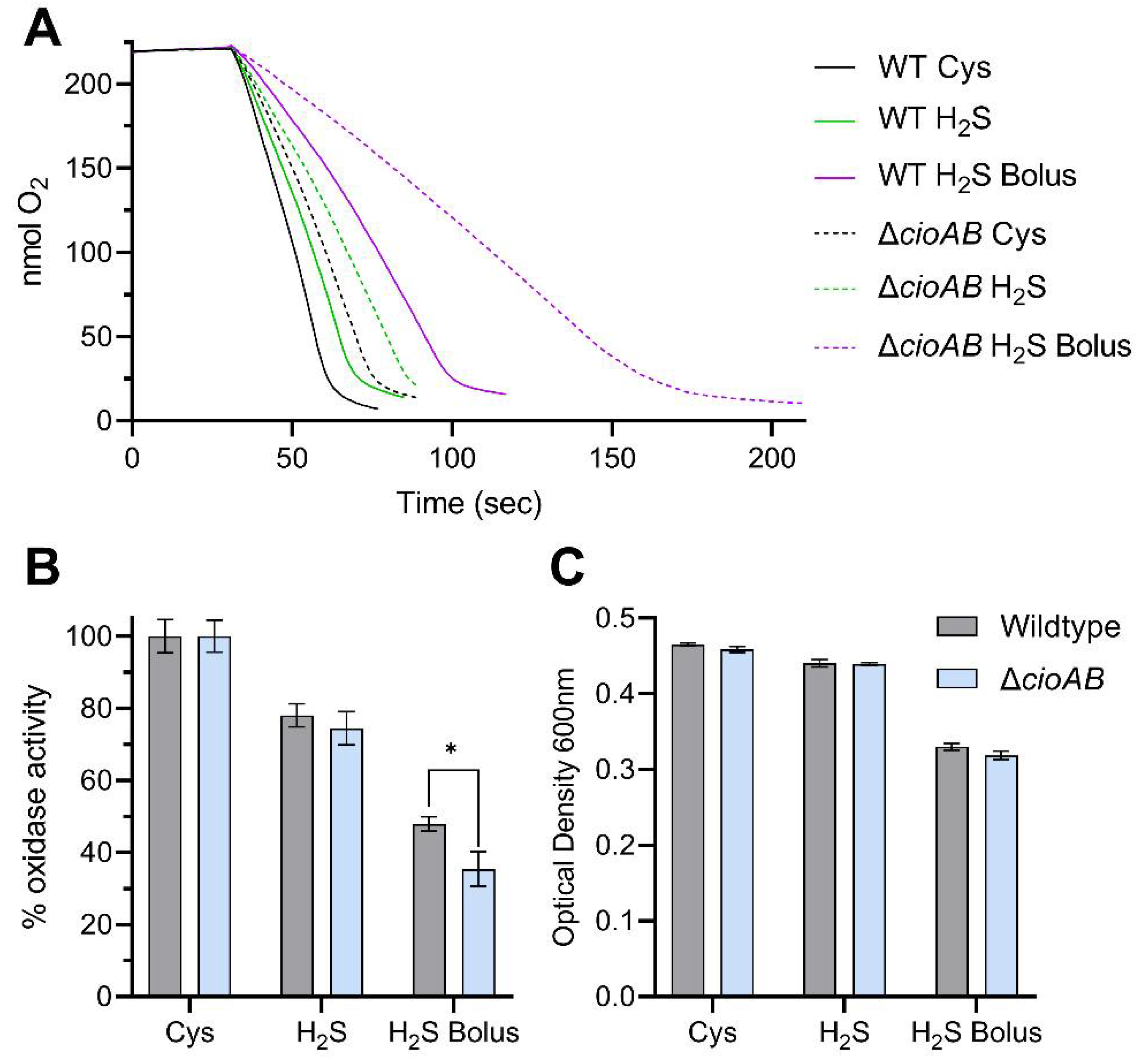
3.3. Sulfide Toxicity in C. jejuni Is Not Solely Attributed to Inactivation of the Terminal Cytochrome C Oxidase
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Igwaran, A.; Okoh, A.I. Human campylobacteriosis: A public health concern of global importance. Heliyon 2019, 5, e02814. [Google Scholar] [CrossRef] [PubMed]
- Galanis, E. Campylobacter and bacterial gastroenteritis. Can. Med. Assoc. J. 2007, 177, 570–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altekruse, S.F.; Stern, N.J.; Fields, P.I.; Swerdlow, D.L. Campylobacter jejuni—An emerging foodborne pathogen. Emerg. Infect. Dis. 1999, 5, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Man, L.; Dale, A.L.; Klare, W.P.; Cain, J.A.; Sumer-Bayraktar, Z.; Niewold, P.; Solis, N.; Cordwell, S.J. Proteomics of Campylobacter jejuni growth in deoxycholate reveals Cj0025c as a cystine transport protein required for wild-type human infection phenotypes. Mol. Cell. Proteom. 2020, 19, 1263–1280. [Google Scholar] [CrossRef] [PubMed]
- Toledano, M.B.; Kumar, C.; le Moan, N.; Spector, D.; Tacnet, F. The system biology of thiol redox system in Escherichia coli and yeast: Differential functions in oxidative stress, iron metabolism and DNA synthesis. FEBS Lett. 2007, 581, 3598–3607. [Google Scholar] [CrossRef] [Green Version]
- Kredich, N.M. The molecular basis for positive regulation of cys promoters in Salmonella typhimurium and Escherichia coli. Mol. Microbiol. 1992, 6, 2747–2753. [Google Scholar] [CrossRef]
- Garvis, S.G.; Tipton, S.L.; Konkel, M.E. Identification of a functional homolog of the Escherichia coli and Salmonella typhimurium cysM gene encoding O-acetylserine sulfhydrylase B in Campylobacter jejuni. Gene 1997, 185, 63–67. [Google Scholar] [CrossRef]
- Vorwerk, H.; Mohr, J.; Huber, C.; Wensel, O.; Schmidt-Hohagen, K.; Gripp, E.; Josenhans, C.; Schomburg, D.; Eisenreich, W.; Hofreuter, D. Utilization of host-derived cysteine-containing peptides overcomes the restricted sulphur metabolism of Campylobacter jejuni. Mol. Microbiol. 2014, 93, 1224–1245. [Google Scholar] [CrossRef]
- Ågren, D.; Schnell, R.; Oehlmann, W.; Singh, M.; Schneider, G. Cysteine synthase (CysM) of Mycobacterium tuberculosis is an O-phosphoserine sulfhydrylase: Evidence for an alternative cysteine biosynthesis pathway in Mycobacteria. J. Biol. Chem. 2008, 283, 31567–31574. [Google Scholar] [CrossRef] [Green Version]
- Takeda, E.; Kunimoto, K.; Kawai, Y.; Kataoka, M.; Ishikawa, K.; Nakamura, T. Role of F225 in O-phosphoserine sulfhydrylase from Aeropyrum pernix K1. Extremophiles 2016, 20, 733–745. [Google Scholar] [CrossRef]
- Burns, K.E.; Baumgart, S.; Dorrestein, P.C.; Zhai, H.; McLafferty, F.W.; Begley, T.P. Reconstitution of a new cysteine biosynthetic pathway in mycobacterium tuberculosis. J. Am. Chem. Soc. 2005, 127, 11602–11603. [Google Scholar] [CrossRef] [Green Version]
- Maier, T.H.P. Semisynthetic production of unnatural l-α-amino acids by metabolic engineering of the cysteine-biosynthetic pathway. Nat. Biotechnol. 2003, 21, 422–427. [Google Scholar] [CrossRef]
- Nakamura, T.; Iwahashi, H.; Eguchi, Y. Enzymatic Proof for the Identity of the S-Sulfocysteine Synthase and Cysteine Synthase B of Salmonella typhimurium. J. Bacteriol. 1984, 158, 1122–1127. [Google Scholar] [CrossRef] [Green Version]
- Rabeh, W.M.; Alguindigue, S.S.; Cook, P.F. Mechanism of the addition half of the O-acetylserine sulfhydrylase—A reaction. Biochemistry 2005, 44, 5541–5550. [Google Scholar] [CrossRef]
- Rabeh, W.M.; Cook, P.F. Structure and mechanism of O-acetylserine sulfhydrylase. J. Biol. Chem. 2004, 279, 26803–26806. [Google Scholar] [CrossRef] [Green Version]
- Rege, V.D.; Kredich, N.M.; Tai, C.H.; Karsten, W.E.; Schnackerz, K.D.; Cook, P.F. A change in the internal aldimine lysine (K42) in O-acetylserine sulfhydrylase to alanine indicates its importance in transimination and as a general base catalyst. Biochemistry 1996, 35, 13485–13493. [Google Scholar] [CrossRef]
- Cameron, A.; Gaynor, E.C. Hygromycin B and apramycin antibiotic resistance cassettes for use in Campylobacter jejuni. PLoS ONE 2014, 9, e95084. [Google Scholar] [CrossRef]
- Garg, N.; Taylor, A.J.; Pastorelli, F.; Flannery, S.E.; Jackson, P.J.; Johnson, M.P.; Kelly, D.J. Genes Linking Copper Trafficking and Homeostasis to the Biogenesis and Activity of the cbb3-Type Cytochrome c Oxidase in the Enteric Pathogen Campylobacter jejuni. Front. Microbiol. 2021, 12, 1620. [Google Scholar] [CrossRef]
- Leskova, A.; Pardue, S.; Glawe, J.D.; Kevil, C.G.; Shen, X. Role of thiosulfate in hydrogen sulfide-dependent redox signaling in endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H256–H264. [Google Scholar] [CrossRef] [Green Version]
- Sirko, A.; Zatyka, M.; Sadowy, E.; Hulanicka, D. Sulfate and thiosulfate transport in Escherichia coli K-12: Evidence for a functional overlapping of sulfate- and thiosulfate-binding proteins. J. Bacteriol. 1995, 177, 4134–4136. [Google Scholar] [CrossRef]
- Liu, Y.W.; Denkmann, K.; Kosciow, K.; Dahl, C.; Kelly, D.J. Tetrathionate stimulated growth of Campylobacter jejuni identifies a new type of bi-functional tetrathionate reductase (TsdA) that is widely distributed in bacteria. Mol. Microbiol. 2013, 88, 173–188. [Google Scholar] [CrossRef]
- Irons, J.; Sacher, J.C.; Szymanski, C.M.; Downs, D.M. Cj1388 Is a RidA Homolog and Is Required for Flagella Biosynthesis and/or Function in Campylobacter jejuni. Front. Microbiol. 2019, 10, 2058. [Google Scholar] [CrossRef]
- Rajaram, V.; Bhavani, B.S.; Kaul, P.; Prakash, V.; Appaji Rao, N.; Savithri, H.S.; Murthy, M.R.N. Structure determination and biochemical studies on Bacillus stearothermophilus E53Q serine hydroxymethyltransferase and its complexes provide insights on function and enzyme memory. FEBS J. 2007, 274, 4148–4160. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Vorwerk, H.; Huber, C.; Lara-Tejero, M.; Mohr, J.; Goodman, A.L.; Eisenreich, W.; Galán, J.E.; Hofreuter, D. Metabolic and fitness determinants for in vitro growth and intestinal colonization of the bacterial pathogen Campylobacter jejuni. PLoS Biology 2017, 15, e2001390. [Google Scholar] [CrossRef] [PubMed]
- Tomasova, L.; Konopelski, P.; Ufnal, M. Gut Bacteria and Hydrogen Sulfide: The New Old Players in Circulatory System Homeostasis. Molecules 2016, 21, 1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dordević, D.; Jančíková, S.; Vítězová, M.; Kushkevych, I. Hydrogen sulfide toxicity in the gut environment: Meta-analysis of sulfate-reducing and lactic acid bacteria in inflammatory processes. J. Adv. Res. 2020, 27, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.H.; Wei, Z.Z.; Hu, K.D.; Hu, L.Y.; Li, Y.H.; Chen, X.Y.; Han, Z.; Yao, G.F.; Zhang, H. Hydrogen sulfide inhibits the growth of Escherichia coli through oxidative damage. J. Microbiol. 2018, 56, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Ríos-González, B.B.; Román-Morales, E.M.; Pietri, R.; López-Garriga, J. Hydrogen sulfide activation in hemeproteins: The sulfheme scenario. J. Inorg. Biochem. 2014, 133, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Blachier, F.; Davila, A.M.; Mimoun, S.; Benetti, P.H.; Atanasiu, C.; Andriamihaja, M.; Benamouzig, R.; Bouillaud, F.; Tomé, D. Luminal sulfide and large intestine mucosa: Friend or foe? Amino Acids 2010, 39, 335–347. [Google Scholar] [CrossRef]
- Blachier, F.; Andriamihaja, M.; Larraufie, P.; Ahn, E.; Lan, A.; Kim, E. Production of hydrogen sulfide by the intestinal microbiota and epithelial cells and consequences for the colonic and rectal mucosa. Am. J. Physiology. Gastrointest. Liver Physiol. 2021, 320, G125–G135. [Google Scholar] [CrossRef]
- Forte, E.; Borisov, V.B.; Falabella, M.; Colaço, H.G.; Tinajero-Trejo, M.; Poole, R.K.; Vicente, J.B.; Sarti, P.; Giuffre, A. The Terminal Oxidase Cytochrome bd Promotes Sulfide-resistant Bacterial Respiration and Growth. Sci. Rep. 2016, 6, 23788. [Google Scholar] [CrossRef] [Green Version]
- Korshunov, S.; Imlay, K.R.C.; Imlay, J.A. The cytochrome bd oxidase of Escherichia coli prevents respiratory inhibition by endogenous and exogenous hydrogen sulfide. Mol. Microbiol. 2016, 101, 62–77. [Google Scholar] [CrossRef] [Green Version]
- Boubeta, F.M.; Bieza, S.A.; Bringas, M.; Palermo, J.C.; Boechi, L.; Estrin, D.A.; Bari, S.E. Hemeproteins as Targets for Sulfide Species. Antioxid. Redox Signal. 2020, 32, 247–257. [Google Scholar] [CrossRef]
- Pietri, R.; Román-Morales, E.; López-Garriga, J. Hydrogen sulfide and hemeproteins: Knowledge and mysteries. Antioxid. Redox Signal. 2011, 15, 393–404. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hitchcock, N.; Kelly, D.J.; Hitchcock, A.; Taylor, A.J. Cysteine Biosynthesis in Campylobacter jejuni: Substrate Specificity of CysM and the Dualism of Sulfide. Biomolecules 2023, 13, 86. https://doi.org/10.3390/biom13010086
Hitchcock N, Kelly DJ, Hitchcock A, Taylor AJ. Cysteine Biosynthesis in Campylobacter jejuni: Substrate Specificity of CysM and the Dualism of Sulfide. Biomolecules. 2023; 13(1):86. https://doi.org/10.3390/biom13010086
Chicago/Turabian StyleHitchcock, Noah, David J. Kelly, Andrew Hitchcock, and Aidan J. Taylor. 2023. "Cysteine Biosynthesis in Campylobacter jejuni: Substrate Specificity of CysM and the Dualism of Sulfide" Biomolecules 13, no. 1: 86. https://doi.org/10.3390/biom13010086