
Iron Deprivation by Oral Deferoxamine Application Alleviates Acute Campylobacteriosis in a Clinical Murine Campylobacter jejuni Infection Model
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbiota-Depleted IL-10−/− Mice
2.2. Campylobacter jejuni Infection and Desferoxamine Pretreatment
2.3. Gastrointestinal Pathogen Concentrations
2.4. Clinical Aspects
2.5. Sampling Procedures and Measurement of Inflammatory Mediators
2.6. Histopathological Analysis and Immunohistochemistry
2.7. Statistics
3. Results
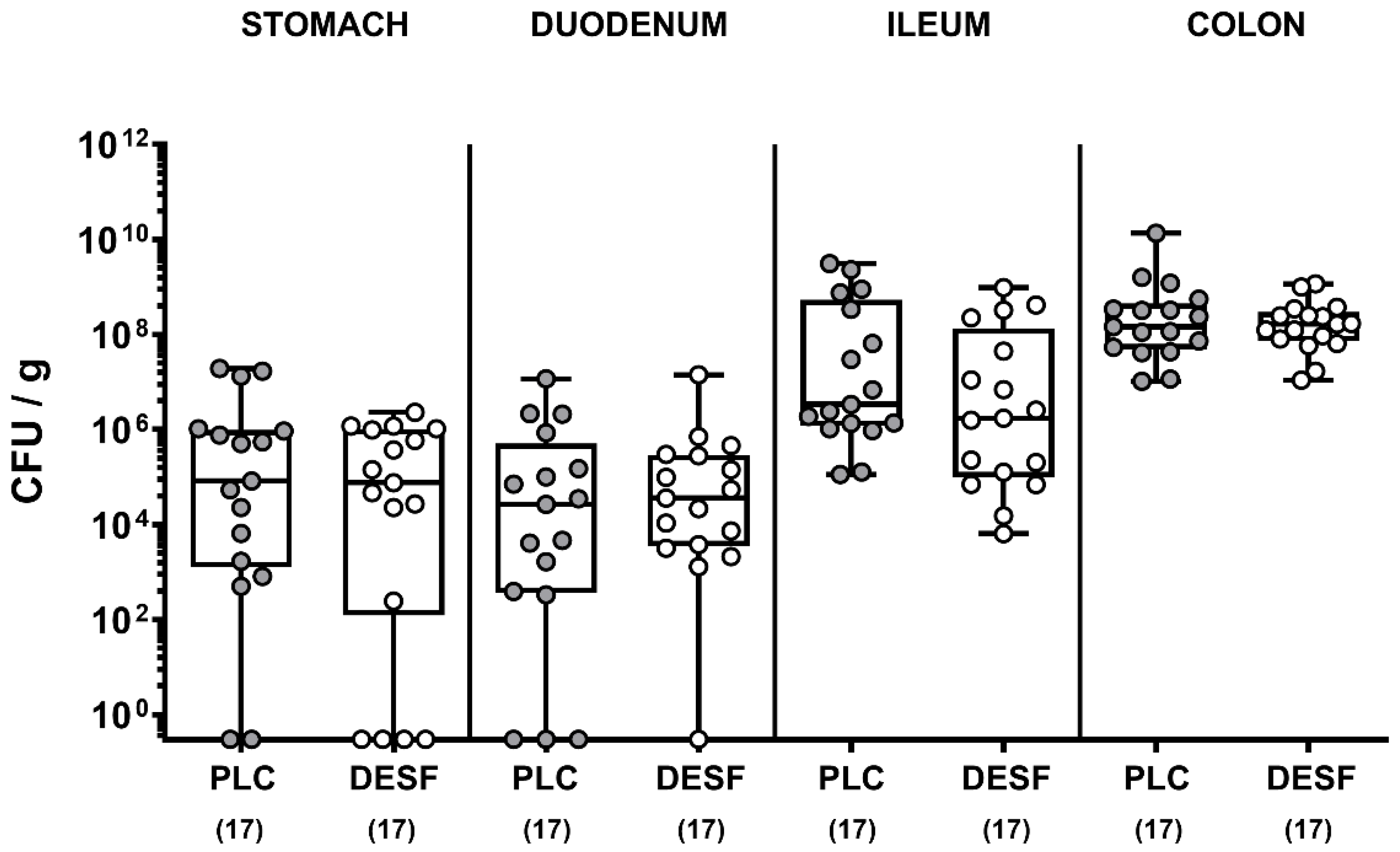
3.1. Pathogenic Loads in the Gastrointestinal Tract Following C. jejuni Infection of Microbiota-Depleted IL-10−/− Mice That Had Been Pretreated with Deferoxamine
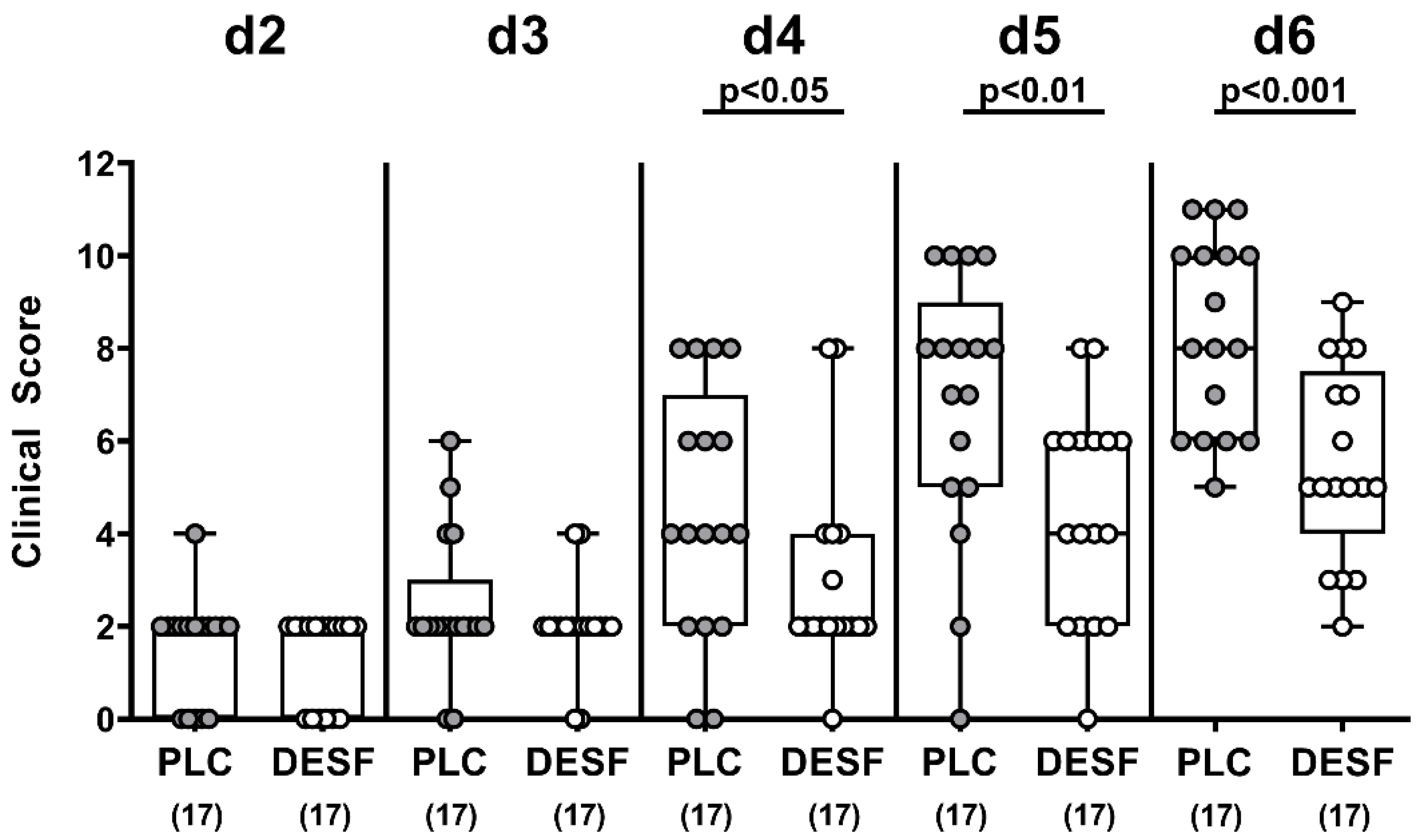
3.2. Kinetic Survey of Clinical Conditions Following C. jejuni Infection of Microbiota-Depleted IL-10−/− Mice That Had Been Pretreated with Deferoxamine
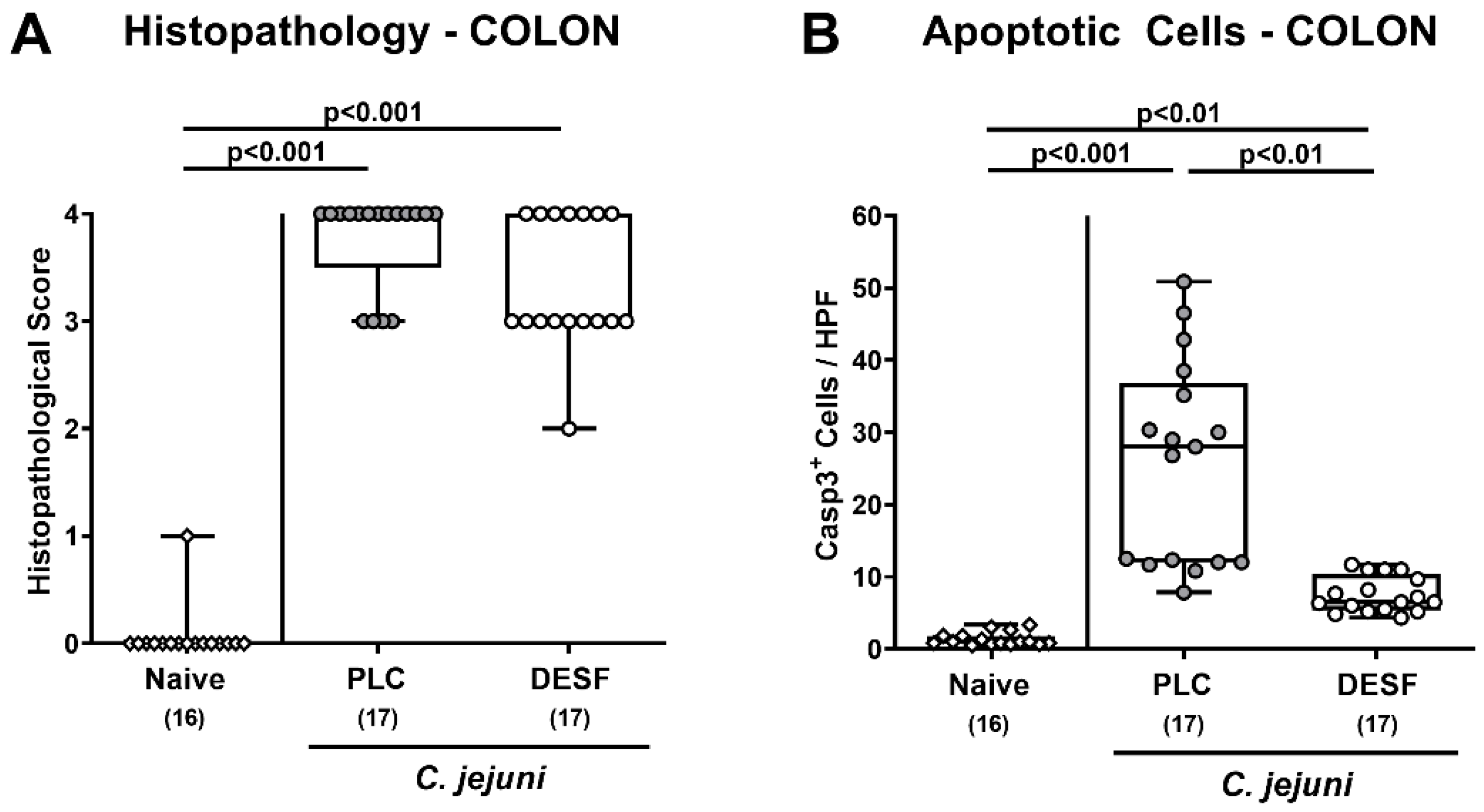
3.3. Microscopic Inflammatory Responses in the Colon Following C. jejuni Infection of Microbiota-Depleted IL-10−/− Mice That Had Been Pretreated with Deferoxamine
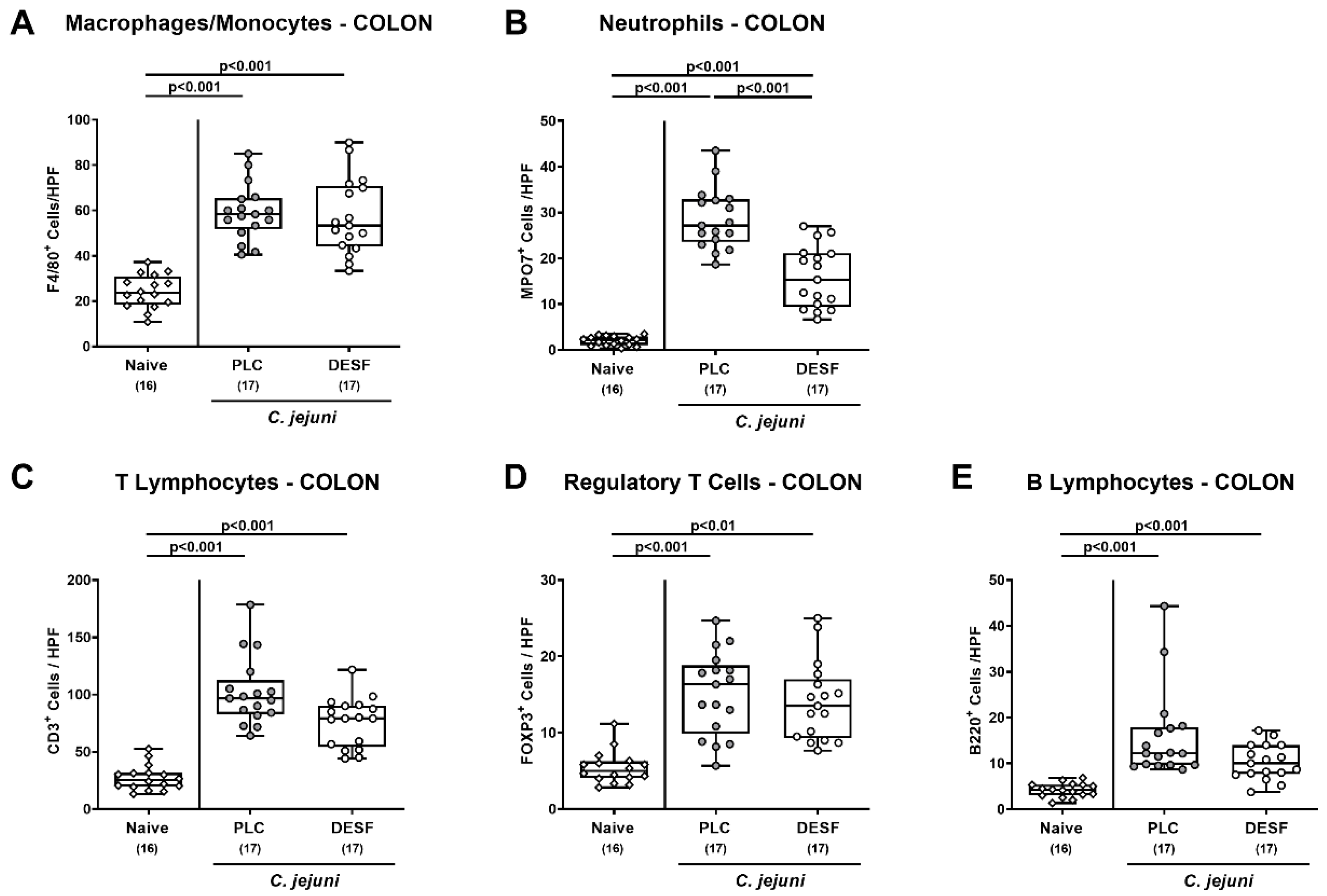
3.4. Colonic Immune Cells Responses Following C. jejuni Infection of Microbiota-Depleted IL-10−/− Mice That Had Been Pretreated with Deferoxamine
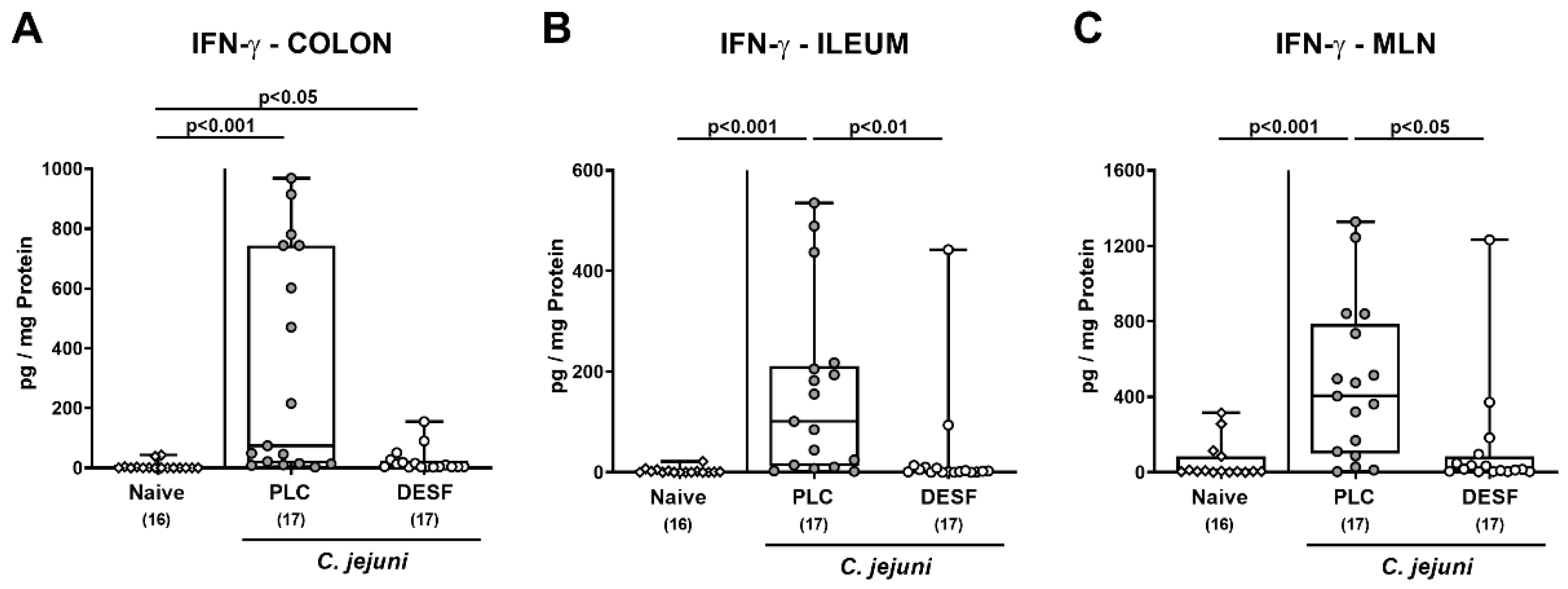
3.5. Intestinal IFN-γ Secretion Following C. jejuni Infection of Microbiota-Depleted IL-10−/− Mice That Had Been Pretreated with Deferoxamine
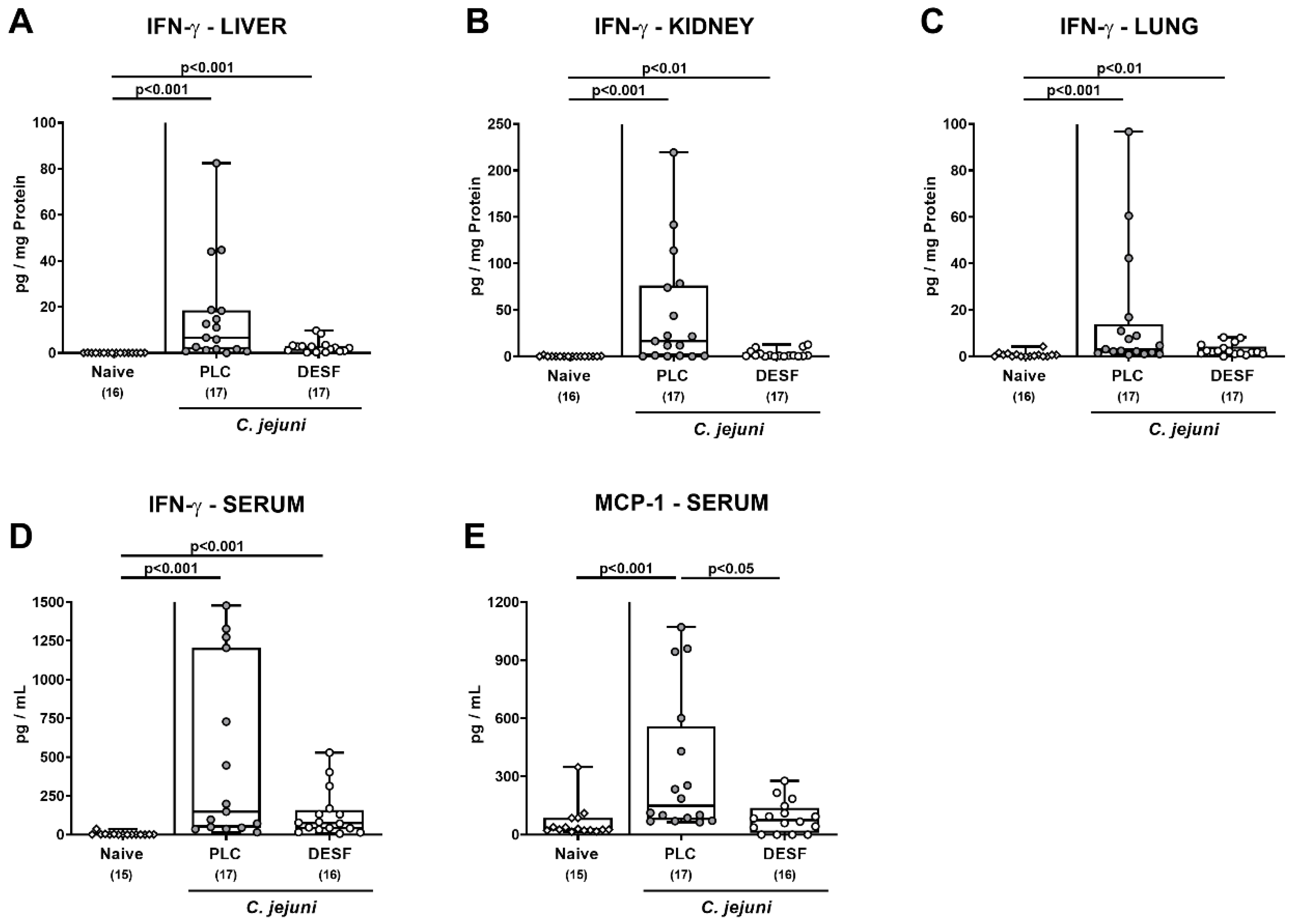
3.6. Pro-Inflammatory Mediator Responses in Extra-Intestinal Compartments Following C. jejuni Infection of Microbiota-Depleted IL-10−/− Mice That Had Been Pretreated with Deferoxamine
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Backert, S.; Tegtmeyer, N.; Cróinín, T.Ó.; Boehm, M.; Heimesaat, M.M. Human campylobacteriosis. In Campylobacter; Klein, G., Ed.; Academic Press: Cambridge, MA, USA, 2017; Chapter 1; pp. 1–25. [Google Scholar] [CrossRef]
- Blaser, M.J. Epidemiologic and clinical features of Campylobacter jejuni infections. J. Infect. Dis. 1997, 176, S103–S105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortensen, N.; Jonasson, S.A.; Lavesson, I.V.; Emberland, K.E.; Litleskare, S.; Wensaas, K.A.; Rortveit, G.; Langeland, N.; Hanevik, K. Characteristics of hospitalized patients during a large waterborne outbreak of Campylobacter jejuni in Norway. PLoS ONE 2021, 16, e0248464. [Google Scholar] [CrossRef] [PubMed]
- Alter, T.; Reich, F. Management Strategies for Prevention of Campylobacter Infections Through the Poultry Food Chain: A European Perspective. Curr. Top. Microbiol. Immunol. 2021, 431, 79–102. [Google Scholar] [CrossRef] [PubMed]
- Heimesaat, M.M.; Backert, S.; Alter, T.; Bereswill, S. Human Campylobacteriosis-A Serious Infectious Threat in a One Health Perspective. Curr. Top. Microbiol. Immunol. 2021, 431, 1–23. [Google Scholar] [PubMed]
- Tegtmeyer, N.; Sharafutdinov, I.; Harrer, A.; Esmaeili, D.S.; Linz, B.; Backert, S. Campylobacter Virulence Factors and Molecular Host–Pathogen Interactions. Curr. Top. Microbiol. Immunol. 2021, 431, 169–202. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Mitchell, H.M.; Man, S.M. Campylobacter. In Molecular Medical Microbiology; Elsevier: Amsterdam, The Netherlands, 2015; pp. 1187–1236. [Google Scholar]
- Price, A.; Jewkes, J.; Sanderson, P. Acute diarrhoea: Campylobacter colitis and the role of rectal biopsy. J. Clin. Pathol. 1979, 32, 990–997. [Google Scholar] [CrossRef] [Green Version]
- de Sá, F.L.; Schulzke, J.-D.; Bücker, R. Diarrheal mechanisms and the role of intestinal barrier dysfunction in Campylobacter infections. Curr. Top. Microbiol. Immunol. 2021, 431, 203–231. [Google Scholar]
- Bücker, R.; Krug, S.M.; Moos, V.; Bojarski, C.; Schweiger, M.R.; Kerick, M.; Fromm, A.; Janssen, S.; Fromm, M.; Hering, N.A.; et al. Campylobacter jejuni impairs sodium transport and epithelial barrier function via cytokine release in human colon. Mucosal. Immunol. 2018, 11, 575–577. [Google Scholar] [CrossRef] [Green Version]
- Kaakoush, N.O.; Castano-Rodriguez, N.; Mitchell, H.M.; Man, S.M. Global Epidemiology of Campylobacter Infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, N.P.; Kuijf, M.L.; Ang, C.W.; Schiellerup, P.; Krogfelt, K.A.; Jacobs, B.C.; van Belkum, A.; Endtz, H.P.; Bergman, M.P. Sialylation of Campylobacter jejuni lipo-oligosaccharides is associated with severe gastro-enteritis and reactive arthritis. Microbes Infect. 2009, 11, 988–994. [Google Scholar] [CrossRef]
- Sun, X.; Threadgill, D.; Jobin, C. Campylobacter jejuni induces colitis through activation of mammalian target of rapamycin signaling. Gastroenterology 2012, 142, 86–95.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mousavi, S.; Bereswill, S.; Heimesaat, M.M. Murine Models for the Investigation of Colonization Resistance and Innate Immune Responses in Campylobacter Jejuni Infections. Curr. Top. Microbiol. Immunol. 2021, 431, 233–263. [Google Scholar] [PubMed]
- Bereswill, S.; Fischer, A.; Plickert, R.; Haag, L.M.; Otto, B.; Kuhl, A.A.; Dasti, J.I.; Zautner, A.E.; Munoz, M.; Loddenkemper, C.; et al. Novel murine infection models provide deep insights into the menage a trois of Campylobacter jejuni, microbiota and host innate immunity. PLoS ONE 2011, 6, e20953. [Google Scholar] [CrossRef]
- Mousavi, S.; Bereswill, S.; Heimesaat, M.M. Novel Clinical Campylobacter jejuni Infection Models Based on Sensitization of Mice to Lipooligosaccharide, a Major Bacterial Factor Triggering Innate Immune Responses in Human Campylobacteriosis. Microorganisms 2020, 8, 482. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, S.; Escher, U.; Thunhorst, E.; Kittler, S.; Kehrenberg, C.; Bereswill, S.; Heimesaat, M.M. Vitamin C alleviates acute enterocolitis in Campylobacter jejuni infected mice. Sci. Rep. 2020, 10, 2921. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, S.; Lobo de Sa, F.D.; Schulzke, J.D.; Bucker, R.; Bereswill, S.; Heimesaat, M.M. Vitamin D in Acute Campylobacteriosis-Results From an Intervention Study Applying a Clinical Campylobacter jejuni Induced Enterocolitis Model. Front. Immunol. 2019, 10, 2094. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, S.; Schmidt, A.-M.; Escher, U.; Kittler, S.; Kehrenberg, C.; Thunhorst, E.; Bereswill, S.; Heimesaat, M.M. Carvacrol ameliorates acute campylobacteriosis in a clinical murine infection model. Gut Pathog. 2020, 12, 2. [Google Scholar] [CrossRef] [Green Version]
- Heimesaat, M.M.; Mousavi, S.; Escher, U.; de Sá, F.D.L.; Peh, E.; Schulzke, J.-D.; Kittler, S.; Bücker, R.; Bereswill, S. Resveratrol Alleviates Acute Campylobacter jejuni Induced Enterocolitis in a Preclinical Murine Intervention Study. Microorganisms 2020, 8, 1858. [Google Scholar] [CrossRef]
- De Sá, F.D.L.; Heimesaat, M.M.; Bereswill, S.; Nattramilarasu, P.K.; Schulzke, J.-D.; Bücker, R. Resveratrol prevents Campylobacter jejuni-induced leaky gut by restoring occludin and claudin-5 in the paracellular leak pathway. Front. Pharmacol. 2021, 12, 454. [Google Scholar]
- Mousavi, S.; Weschka, D.; Bereswill, S.; Heimesaat, M.M. Preclinical Evaluation of Oral Urolithin-A for the Treatment of Acute Campylobacteriosis in Campylobacter jejuni Infected Microbiota-Depleted IL-10−/−Mice. Pathogens 2021, 10, 7. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Mousavi, S.; Weschka, D.; Bereswill, S. Anti-Pathogenic and Immune-Modulatory Effects of Peroral Treatment with Cardamom Essential Oil in Acute Murine Campylobacteriosis. Microorganisms 2021, 9, 169. [Google Scholar] [CrossRef] [PubMed]
- Bereswill, S.; Mousavi, S.; Weschka, D.; Buczkowski, A.; Schmidt, S.; Heimesaat, M.M. Peroral Clove Essential Oil Treatment Ameliorates Acute Campylobacteriosis-Results from a Preclinical Murine Intervention Study. Microorganisms 2021, 9, 735. [Google Scholar] [CrossRef] [PubMed]
- Heimesaat, M.M.; Mousavi, S.; Weschka, D.; Bereswill, S. Garlic Essential Oil as Promising Option for the Treatment of Acute Campylobacteriosis—Results from a Preclinical Placebo-Controlled Intervention Study. Microorganisms 2021, 9, 1140. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, S.; Weschka, D.; Bereswill, S.; Heimesaat, M.M. Immune-Modulatory Effects upon Oral Application of Cumin-Essential-Oil to Mice Suffering from Acute Campylobacteriosis. Pathogens 2021, 10, 818. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Mousavi, S.; Kløve, S.; Genger, C.; Weschka, D.; Giladi, E.; Bereswill, S.; Gozes, I. Immune-modulatory Properties of the Octapeptide NAP in Campylobacter jejuni Infected Mice Suffering from Acute Enterocolitis. Microorganisms 2020, 8, 802. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Mousavi, S.; Kløve, S.; Genger, C.; Weschka, D.; Tamas, A.; Reglodi, D.; Bereswill, S. Pituitary Adenylate Cyclase-Activating Polypeptide Alleviates Intestinal, Extra-Intestinal and Systemic Inflammatory Responses during Acute Campylobacter jejuni-induced Enterocolitis in Mice. Pathogens 2020, 9, 805. [Google Scholar] [CrossRef]
- Ekmekciu, I.; Fiebiger, U.; Stingl, K.; Bereswill, S.; Heimesaat, M.M. Amelioration of intestinal and systemic sequelae of murine Campylobacter jejuni infection by probiotic VSL#3 treatment. Gut Pathog. 2017, 9, 17. [Google Scholar] [CrossRef] [Green Version]
- Heimesaat, M.M.; Mrazek, K.; Bereswill, S. Murine Fecal Microbiota Transplantation Alleviates Intestinal and Systemic Immune Responses in Campylobacter jejuni Infected Mice Harboring a Human Gut Microbiota. Front. Immunol. 2019, 10, 2272. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Mrazek, K.; Bereswill, S. Murine fecal microbiota transplantation lowers gastrointestinal pathogen loads and dampens pro-inflammatory immune responses in Campylobacter jejuni infected secondary abiotic mice. Sci. Rep. 2019, 9, 19797. [Google Scholar] [CrossRef] [Green Version]
- Bereswill, S.; Mousavi, S.; Weschka, D.; Heimesaat, M.M. Disease-Alleviating Effects of Peroral Activated Charcoal Treatment in Acute Murine Campylobacteriosis. Microorganisms 2021, 9, 1424. [Google Scholar] [CrossRef]
- Bellotti, D.; Remelli, M. Deferoxamine B: A natural, excellent and versatile metal chelator. Molecules 2021, 26, 3255. [Google Scholar] [CrossRef] [PubMed]
- Jurado, R.L. Iron, infections, and anemia of inflammation. Clin. Infect. Dis. 1997, 25, 888–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hershko, C.; Peto, T.E.; Weatherall, D.J. Iron and infection. Br. Med. J. (Clin. Res. Ed.) 1988, 296, 660–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perricone, C.; Bartoloni, E.; Bursi, R.; Cafaro, G.; Guidelli, G.M.; Shoenfeld, Y.; Gerli, R. COVID-19 as part of the hyperferritinemic syndromes: The role of iron depletion therapy. Immunol. Res. 2020, 68, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Palyada, K.; Threadgill, D.; Stintzi, A. Iron acquisition and regulation in Campylobacter jejuni. J. Bacteriol. 2004, 186, 4714–4729. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.E.; Williams, P.H.; Ketley, J.M. Pumping iron: Mechanisms for iron uptake by Campylobacter. Microbiol. 2009, 155, 3157–3165. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.M.; Boinett, C.J.; Chan, A.C.; Parkhill, J.; Murphy, M.E.; Gaynor, E.C. Investigating the Campylobacter jejuni transcriptional response to host intestinal extracts reveals the involvement of a widely conserved iron uptake system. MBio 2018, 9, e01347-18. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Lin, J. Characterization of high affinity iron acquisition systems in Campylobacter jejuni. In Campylobacter jejuni; Springer: Berlin/Heidelberg, Germany, 2017; pp. 65–78. [Google Scholar]
- Butcher, J.; Stintzi, A. The transcriptional landscape of Campylobacter jejuni under iron replete and iron limited growth conditions. PLoS ONE 2013, 8, e79475. [Google Scholar] [CrossRef] [Green Version]
- Barollo, M.; D’Incà, R.; Scarpa, M.; Medici, V.; Cardin, R.; Fries, W.; Angriman, I.; Sturniolo, G. Effects of iron deprivation or chelation on DNA damage in experimental colitis. Int. J. Color. Dis. 2004, 19, 461–466. [Google Scholar] [CrossRef]
- Minaiyan, M.; Mostaghel, E.; Mahzouni, P. Preventive therapy of experimental colitis with selected iron chelators and anti-oxidants. Int. J. Prev. Med. 2012, 3, S162–S169. [Google Scholar]
- Millar, A.; Rampton, D.; Blake, D. Effects of iron and iron chelation in vitro on mucosal oxidant activity in ulcerative colitis. Aliment. Pharmacol. Ther. 2000, 14, 1163–1168. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, N.F.; Brittenham, G.M. Iron-chelating therapy and the treatment of thalassemia. Blood 1997, 89, 739–761. [Google Scholar] [CrossRef] [PubMed]
- Heimesaat, M.M.; Mousavi, S.; Bandick, R.; Bereswill, S. Campylobacter jejuni infection induces acute enterocolitis in IL-10−/−mice pretreated with ampicillin plus sulbactam. Eur. J. Microbiol. Immunol. 2022, 12, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Heimesaat, M.M.; Alutis, M.; Grundmann, U.; Fischer, A.; Tegtmeyer, N.; Bohm, M.; Kuhl, A.A.; Gobel, U.B.; Backert, S.; Bereswill, S. The role of serine protease HtrA in acute ulcerative enterocolitis and extra-intestinal immune responses during Campylobacter jejuni infection of gnotobiotic IL-10 deficient mice. Front. Cell. Infect. Microbiol. 2014, 4, 77. [Google Scholar] [CrossRef] [Green Version]
- Erben, U.; Loddenkemper, C.; Doerfel, K.; Spieckermann, S.; Haller, D.; Heimesaat, M.M.; Zeitz, M.; Siegmund, B.; Kühl, A.A. A guide to histomorphological evaluation of intestinal inflammation in mouse models. Int. J. Clin. Exp. Pathol. 2014, 7, 4557–4576. [Google Scholar]
- Heimesaat, M.M.; Giladi, E.; Kuhl, A.A.; Bereswill, S.; Gozes, I. The octapetide NAP alleviates intestinal and extra-intestinal anti-inflammatory sequelae of acute experimental colitis. Peptides 2018, 101, 1–9. [Google Scholar] [CrossRef]
- Schmidt, A.-M.; Escher, U.; Mousavi, S.; Tegtmeyer, N.; Boehm, M.; Backert, S.; Bereswill, S.; Heimesaat, M.M. Immunopathological properties of the Campylobacter jejuni flagellins and the adhesin CadF as assessed in a clinical murine infection model. Gut Pathog. 2019, 11, 24. [Google Scholar] [CrossRef] [Green Version]
- Fisher, A.L.; Sangkhae, V.; Balušíková, K.; Palaskas, N.J.; Ganz, T.; Nemeth, E. Iron-dependent apoptosis causes embryotoxicity in inflamed and obese pregnancy. Nat. Commun. 2021, 12, 4026. [Google Scholar] [CrossRef]
- El-Sheikh, A.A.; Ameen, S.H.; AbdEl-Fatah, S.S. Ameliorating iron overload in intestinal tissue of adult male rats: Quercetin vs deferoxamine. J. Toxicol. 2018, 2018, 8023840. [Google Scholar] [CrossRef]
- Xu, M.; Tao, J.; Yang, Y.; Tan, S.; Liu, H.; Jiang, J.; Zheng, F.; Wu, B. Ferroptosis involves in intestinal epithelial cell death in ulcerative colitis. Cell Death Dis. 2020, 11, 86. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.; Meyer, D. Desferrioxamine as immunomodulatory agent during microorganism infection. Curr. Pharm. Des. 2009, 15, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, C.; Pan, S.; Miao, Q.; Xue, J.; Xun, J.; Zhang, Y.; Gao, Y.; Duan, X.; Fan, Y. Deferoxamine attenuates lipopolysaccharide-induced inflammatory responses and protects against endotoxic shock in mice. Biochem. Biophys. Res. Commun. 2015, 465, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Cao, J.B.; Zhang, L.M.; Li, Y.F.; Mi, W.D. Deferoxamine attenuates lipopolysaccharide-induced neuroinflammation and memory impairment in mice. J. Neuroinflamm. 2015, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-J.; Wei, H.; Frei, B. The iron chelator, desferrioxamine, reduces inflammation and atherosclerotic lesion development in experimental mice. Exp. Biol. Med. 2010, 235, 633–641. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.J.; Frei, B. Intracellular metal ion chelators inhibit TNFalpha-induced SP-1 activation and adhesion molecule expression in human aortic endothelial cells. Free Radic. Biol. Med. 2003, 34, 674–682. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bereswill, S.; Mousavi, S.; Weschka, D.; Buczkowski, A.; Schmidt, S.; Heimesaat, M.M. Iron Deprivation by Oral Deferoxamine Application Alleviates Acute Campylobacteriosis in a Clinical Murine Campylobacter jejuni Infection Model. Biomolecules 2023, 13, 71. https://doi.org/10.3390/biom13010071
Bereswill S, Mousavi S, Weschka D, Buczkowski A, Schmidt S, Heimesaat MM. Iron Deprivation by Oral Deferoxamine Application Alleviates Acute Campylobacteriosis in a Clinical Murine Campylobacter jejuni Infection Model. Biomolecules. 2023; 13(1):71. https://doi.org/10.3390/biom13010071
Chicago/Turabian StyleBereswill, Stefan, Soraya Mousavi, Dennis Weschka, Agnes Buczkowski, Sebastian Schmidt, and Markus M. Heimesaat. 2023. "Iron Deprivation by Oral Deferoxamine Application Alleviates Acute Campylobacteriosis in a Clinical Murine Campylobacter jejuni Infection Model" Biomolecules 13, no. 1: 71. https://doi.org/10.3390/biom13010071