
The Yin and Yang of Mesenchymal Cells in the Corneal Stromal Fibrosis Response to Injury: The Cornea as a Model of Fibrosis in Other Organs


Abstract
:1. Introduction
2. Mesenchymal Cells or Mesenchymal Progenitors
2.1. The Keratocyte
2.2. The Corneal Fibroblast
- (1)
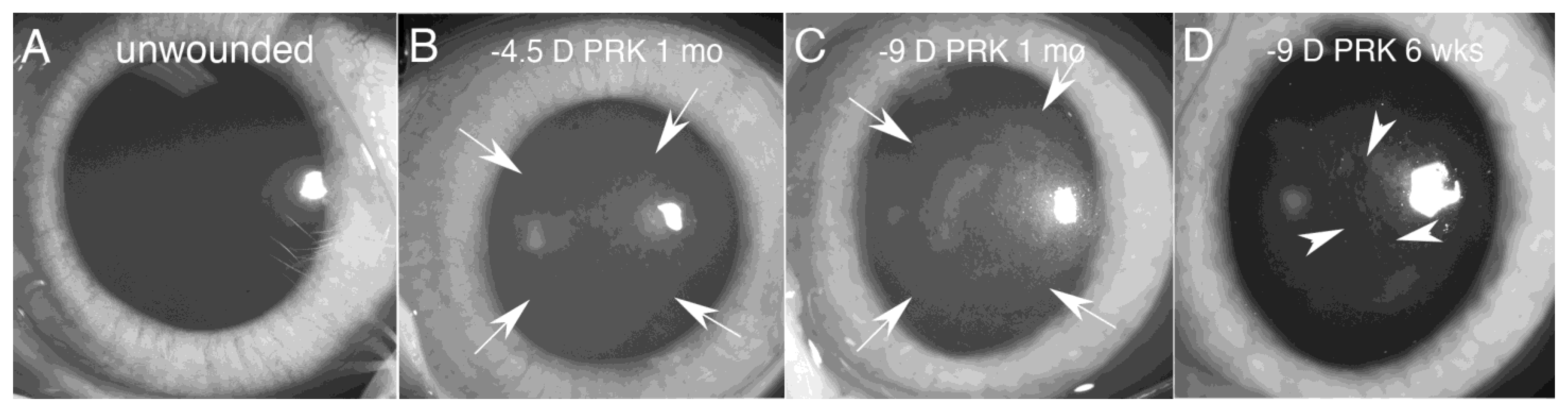
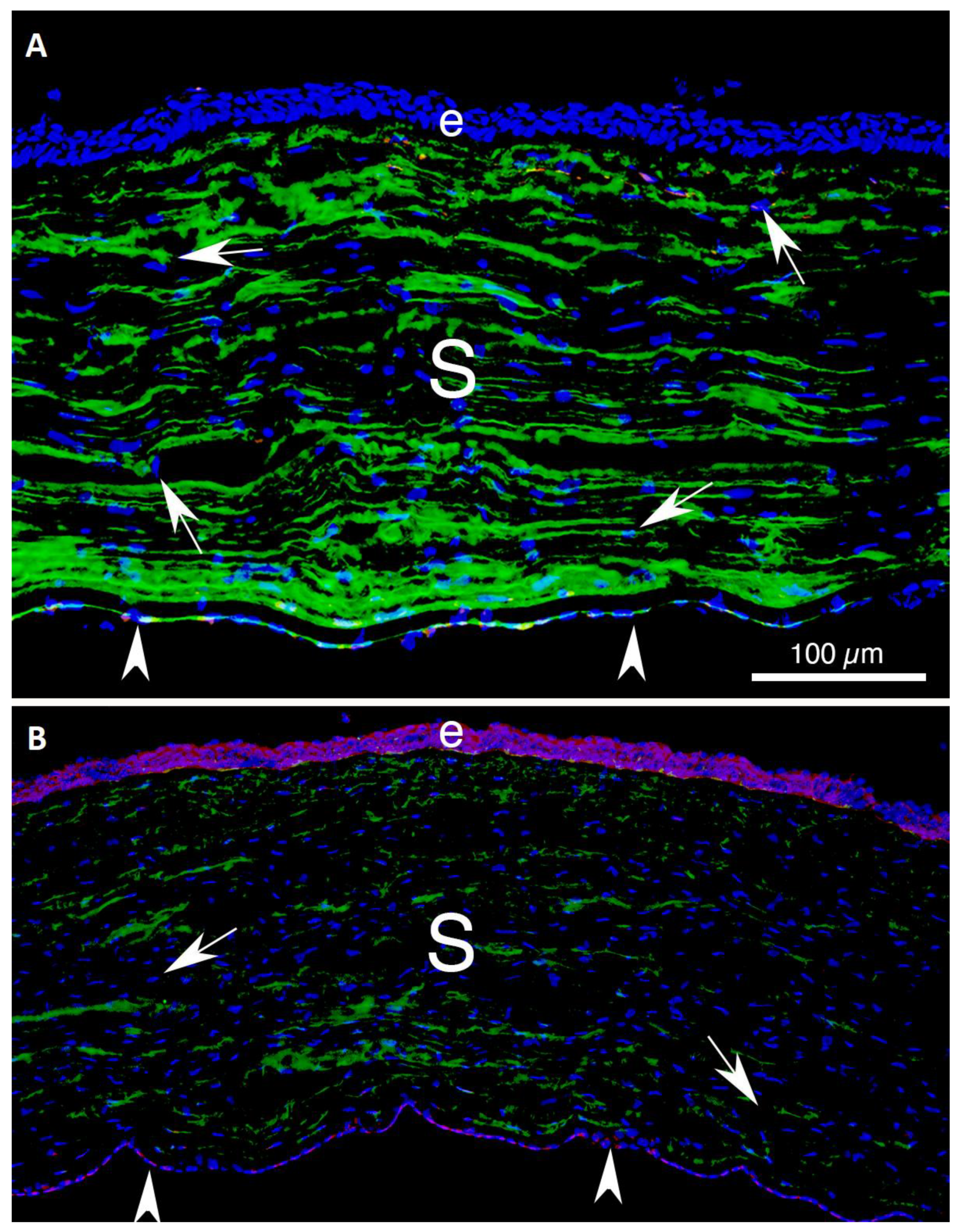
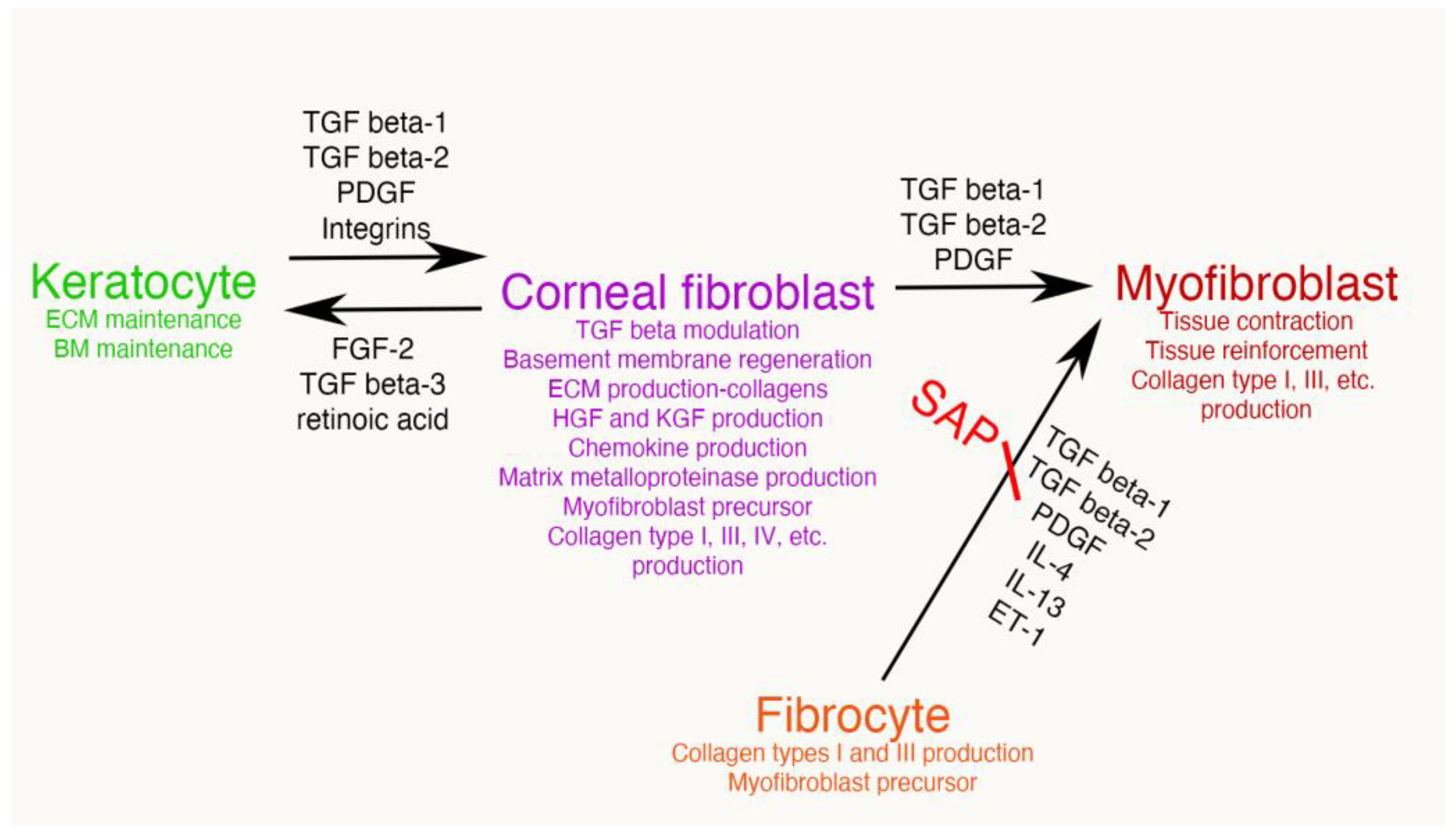
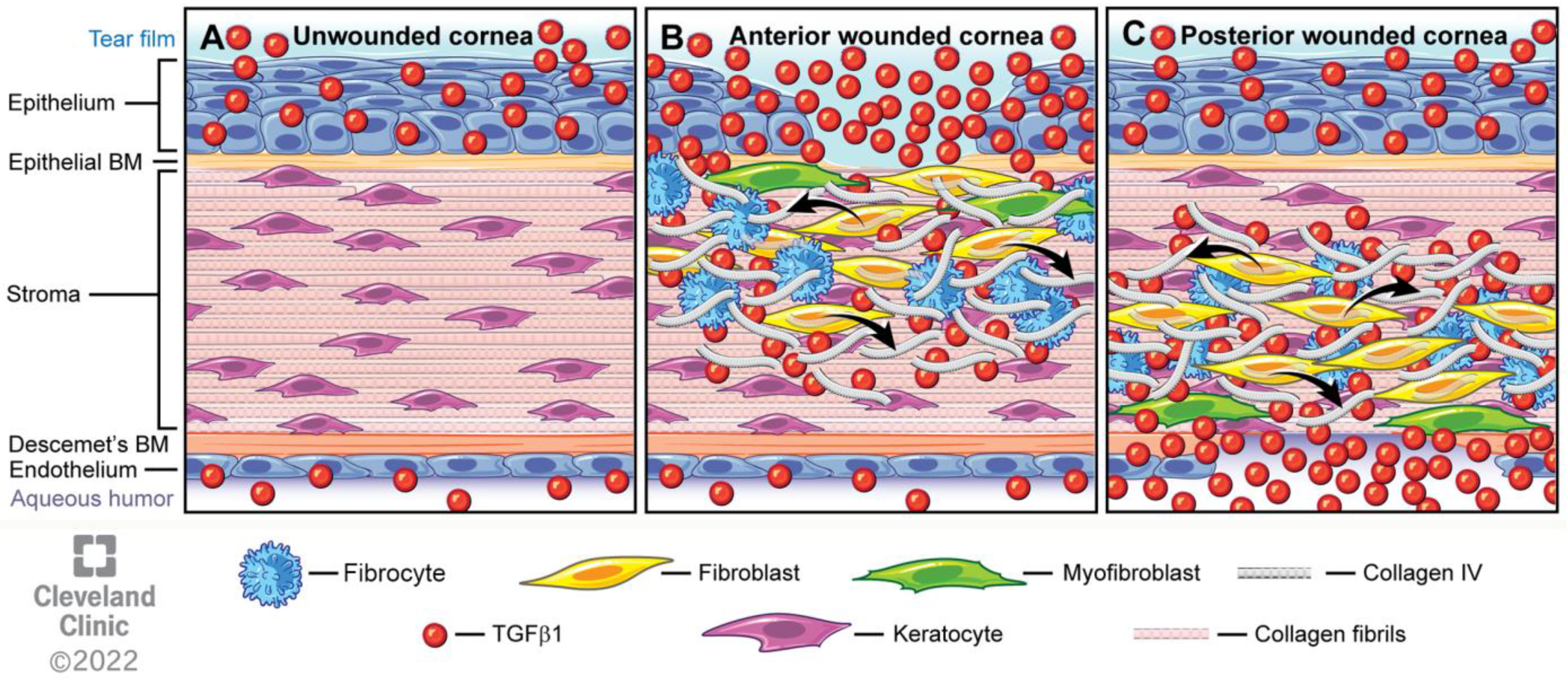
- First, the TGF beta stimulation that drives corneal fibroblast development, also upregulates collagen type IV production by these cells [9,15,16]. This collagen type IV produced in large quantities by corneal fibroblasts serves two major functions. First, collagen type IV binds TGF beta-1 or TGF beta-2 and prevents these growth factors from binding their cognate TGF beta receptors [17,18]. Therefore, the large amounts of collagen type IV produced by the corneal fibroblasts, much of which is not associated with the corneal basement membranes (BM) [9,15,16], down-regulates the effects of TGF beta-1 and TGF beta-2 that enter the stroma from the tears, epithelium, corneal endothelium and/or aqueous humor, and are activated after injury. A major impact of that non-BM collagen type IV, therefore, is to moderate the pro-fibrotic effects of TGF beta by regulating (decreasing) myofibroblast development from the corneal fibroblasts themselves and bone marrow-derived fibrocytes [19,20], the two best-characterized precursors to myofibroblasts in the corneal stroma [1,4,8,9,12,13,14,19,20]. This non-BM collagen type IV production by corneal fibroblasts may be a major determinant of why many relatively low-level corneal injuries, such as abrasions or lower correction photorefractive keratectomy (PRK) surgeries, generate corneal fibroblasts but not myofibroblasts in the corneal stroma [8,21]. With more significant injuries, such as high correction PRKs, alkali burns, or Descemetorhexis, this modulatory system is overwhelmed and large numbers of myofibroblasts develop in the stroma [8,9,14,15,21]. The second function of the collagen type IV produced by corneal fibroblasts is to contribute to EBM and/or Descemet’s membrane regeneration after injury to these important adhesion and regulatory structures in the cornea [9,14]. This regeneration of the BMs is critical to reestablish normal regulation of TGF beta localization to the stroma, eventually trigger myofibroblast apoptosis, and return of the cornea to its normal transparent structure and function [8,9,14].Figure 4. Mesenchymal cells of the injured corneal stroma. The unwounded corneal stroma is populated mostly with keratocytes, with a few nerve cells and resident dendritic cells. At the site of injury, driven by TGF beta-1 and TGF beta-2, along with PDGF and integrin signaling, keratocytes transition into corneal fibroblasts. Corneal fibroblasts that receive sustained and adequate levels of TGF beta-1 and/or TGF beta-2 develop into mature α-SMA-positive myofibroblasts. Another established source of corneal myofibroblasts is fibrocytes which differentiate from bone marrow-derived progenitors that migrate into the cornea from the limbal blood vessels after injury. Fibrocytes that also receive sustained and adequate levels of activated TGF beta-1 and/or TGF beta-2 develop into mature α-SMA-positive myofibroblasts. It remains controversial whether myofibroblasts, once generated, can return to corneal fibroblasts and/or fibrocytes, or rather are always eliminated via apoptosis. Some of the functions of each corneal stromal mesenchymal or mesenchymal progenitor cell type are listed. SAP is serum amyloid protein. SAP most likely functions to inhibit fibrocyte development from precursors but may also directly inhibit fibrocyte differentiation into myofibroblasts [22].Figure 4. Mesenchymal cells of the injured corneal stroma. The unwounded corneal stroma is populated mostly with keratocytes, with a few nerve cells and resident dendritic cells. At the site of injury, driven by TGF beta-1 and TGF beta-2, along with PDGF and integrin signaling, keratocytes transition into corneal fibroblasts. Corneal fibroblasts that receive sustained and adequate levels of TGF beta-1 and/or TGF beta-2 develop into mature α-SMA-positive myofibroblasts. Another established source of corneal myofibroblasts is fibrocytes which differentiate from bone marrow-derived progenitors that migrate into the cornea from the limbal blood vessels after injury. Fibrocytes that also receive sustained and adequate levels of activated TGF beta-1 and/or TGF beta-2 develop into mature α-SMA-positive myofibroblasts. It remains controversial whether myofibroblasts, once generated, can return to corneal fibroblasts and/or fibrocytes, or rather are always eliminated via apoptosis. Some of the functions of each corneal stromal mesenchymal or mesenchymal progenitor cell type are listed. SAP is serum amyloid protein. SAP most likely functions to inhibit fibrocyte development from precursors but may also directly inhibit fibrocyte differentiation into myofibroblasts [22].
![Biomolecules 13 00087 g004]()

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
- (2)
- Secondly, corneal fibroblasts produce other BM components that contribute to regeneration of the EBM and/or Descemet’s membrane, such as perlecan, to facilitate regeneration of nascent EBM and Descemet’s membrane after injury [8,14,23]. After the epithelium or endothelium, lays down the earliest components of the EBM or Descemet’s membrane, respectively, including self-polymerizing laminins, then corneal fibroblasts contribute other components [8,9,14]. Defective perlecan insertion into the nascent EBM after injury has been demonstrated in corneas that develop fibrosis compared to those that do not [23].
- (3)
- A third major function performed by corneal fibroblasts is the production of growth factors that control the proliferation, migration, differentiation, and apoptosis of the corneal epithelium, including hepatocyte growth factor (HGF) and keratinocyte growth factor (KGF or fibroblast growth factor (FGF)-7) [24,25]. HGF and KGF production by keratocytes is nearly undetectable, but after injury these growth factors are produced at high levels by corneal fibroblasts [25]. Corneal fibroblasts, therefore, are important contributors to the healing and restoration of normal epithelium following injuries.
- (4)
- Immediately after corneal injury, fibrocytes, monocytes, polymorphonuclear cells, and other bone marrow-derived cells stream into the corneal stroma from the limbal blood vessels. These cells are attracted by interleukin (IL)-1 alpha, IL-1 beta and other cytokines produced by corneal epithelial and endothelial cells [26]. A fourth important function of corneal fibroblast is the production of chemokines, such as granulocyte-colony stimulating factor (G-CSF), neutrophil-activating peptide (ENA-78), monocyte-derived neutrophil chemotactic factor (MDNCF), and monocyte chemotactic and activating factor (MCAF), after corneal fibroblasts are stimulated by IL-1 or tumor necrosis activating factor alpha (TNFa) [27]. The reference title refers to keratocytes, but the cells studied in vitro were corneal fibroblasts. Therefore, corneal fibroblasts contribute to attracting fibrocytes and immune cells to the injured cornea.
- (5)
- A fifth major function of corneal fibroblasts is that they serve as major precursors to stromal myofibroblasts when driven by ongoing and adequate levels of TGF beta-1 and/or TGF beta-2 [1,12,26,28,29,30]. What determines whether a particular corneal fibroblast persists and continues to perform its numerous functions or develops into a myofibroblast in a cornea? That is an area of considerable research interest. Likely, it is the overall milieu that corneal fibroblast finds itself in with regard to the concentrations of activated TGF beta, fibroblast growth factor-2, PDGF, IL-1, and other growth factors and cytokines that determines the fate of that cell at a particular moment during the corneal wound healing response. This is the Yin and Yang of corneal fibroblasts. Although they have an essential role in downregulating myofibroblast development and fibrosis, and non-fibrotic healing of the cornea, they are a primary precursor for myofibroblasts in the cornea.
2.3. The Fibrocyte, a Bone Marrow-Derived Mesenchymal Progenitor
2.4. Myofibroblasts
3. Immune Cells
4. Mesenchymal Cellular Interactions in the Responses to Corneal Injury
5. Conclusions
6. Patents/Conflicts of Interest
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Yam, G.H.F.; Riau, A.K.; Funderburgh, M.L.; Mehta, J.S.; Jhanji, V. Kratocyte biology. Exp. Eye. Res. 2020, 196, 108062. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.A.; Hertsenberg, A.J.; Funderburgh, M.L.; Mann, M.M.; Du, Y.; Davoli, K.A.; Mich-Basso, J.D.; Yang, L.; Funderburgh, J.L. Differentiation of human embryonic stem cells into cells with corneal keratocyte phenotype. PLoS One 2013, 8, e56831. [Google Scholar] [CrossRef] [Green Version]
- Jester, J.V.; Barry, P.A.; Lind, G.J.; Petroll, W.M.; Garana, R.; Cavanagh, H.D. Corneal keratocytes: In situ and in vitro organization of cytoskeletal contractile proteins. Invest. Ophthalmol. Vis. Sci. 1994, 35, 730–743. [Google Scholar] [PubMed]
- Funderburgh, M.L.; Mann, M.M.; Funderburgh, J.L. Keratocyte phenotype is enhanced in the absence of attachment to the substratum. Mol. Vis. 2008, 14, 308–317. [Google Scholar] [PubMed]
- Hassell, J.R.; Birk, D.E. The molecular basis of corneal transparency. Exp. Eye. Res. 2010, 91, 326–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medeiros, C.S.; Marino, G.K.; Santhiago, M.R.; Wilson, S.E. The corneal basement membranes and stromal fibrosis. Invest. Ophthalmol. Vis. Sci. 2018, 59, 4044–4053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saikia, P.; Medeiros, C.S.; Thangavadivel, S.; Wilson, S.E. Basement membranes in the cornea and other organs that commonly develop fibrosis. Cell Tissue. Res. 2018, 374, 439–453. [Google Scholar] [CrossRef]
- de Oliveira, R.C.; Tye, G.; Sampaio, L.P.; Shiju, T.M.; Dedreu, J.; Menko, A.S.; Santhiago, M.R.; Wilson, S.E. TGFβ1 and TGFβ2 proteins in corneas with and without stromal fibrosis: Delayed regeneration of epithelial barrier function and the epithelial basement membrane in corneas with stromal fibrosis. Exp. Eye Res. 2021, 202, 108325. [Google Scholar] [CrossRef]
- Sampaio, L.P.; Shiju, T.M.; Hilgert, G.S.L.; de Oliveira, R.C.; DeDreu, J.; Menko, A.S.; Santhiago, M.R.; Wilson, S.E. Descemet’s membrane injury and regeneration, and posterior corneal fibrosis in rabbits. Exp. Eye Res. 2021, 213, 108803. [Google Scholar] [CrossRef]
- Jester, J.V.; Huang, J.; Petroll, W.M.; Cavanagh, H.D. TGFbeta induced myofibroblast differentiation of rabbit keratocytes requires synergistic TGFbeta, PDGF and integrin signaling. Exp. Eye Res. 2002, 75, 645–657. [Google Scholar] [CrossRef]
- Sidney, L.E.; Hopkinson, A. Corneal keratocyte transition to mesenchymal stem cell phenotype and reversal using serum-free medium supplemented with fibroblast growth factor-2, transforming growth factor-β3 and retinoic acid. J. Tissue. Eng. Regen. Med. 2018, 12, e203–e215. [Google Scholar] [CrossRef] [PubMed]
- Fini, M.E. Keratocyte and fibroblast phenotypes in the repairing cornea. Prog. Retin. Eye. Res. 1999, 18, 529–551. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.E.; Sampaio, L.P.; Shiju, T.M.; Hilgert, G.S.L.; de Oliveira, R.C. Corneal opacity: Cell biological determinants of the transition from transparency to transient haze to scarring fibrosis, and resolution, after injury. Invest. Ophthalmol. Vis. Sci. 2022, 63, 22. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, R.C.; Sampaio, L.P.; Shiju, T.M.; Santhiago, M.R.; Wilson, S.E. Epithelial basement membrane regeneration after PRK-induced epithelial-stromal injury in rabbits: Fibrotic vs. non-fibrotic corneal healing. J. Ref. Surg. 2022, 38, 50–60. [Google Scholar] [CrossRef]
- Sampaio, L.P.; Hilgert, G.S.L.; Shiju, T.M.; Murillo, S.E.; Santhiago, M.R.; Wilson, S.E. Topical losartan inhibits corneal scarring fibrosis and collagen type IV deposition after Descemet’s membrane-endothelial excision in rabbits. Exp. Eye Res. 2022, 216, 108940. [Google Scholar] [CrossRef]
- Wilson, S.E.; Shiju, T.M.; Sampaio, L.P.; Hilgert, G.S.L. Corneal fibroblast collagen type IV negative feedback loop modulation of TGF beta: A likely fibrosis modulating system in other organs. Matrix. Biol. 2022, 109, 162–172. [Google Scholar] [CrossRef]
- Paralkar, V.M.; Vukicevic, S.; Reddi, A.H. Transforming growth factor beta type 1 binds to collagen IV of basement membrane matrix: Implications for development. Dev. Biol. 1991, 143, 303–308. [Google Scholar] [CrossRef]
- Shibuya, H.; Okamoto, O.; Fujiwara, S. The bioactivity of transforming growth factor-beta1 can be regulated via binding to dermal collagens in mink lung epithelial cells. J. Dermatol. Sci. 2006, 41, 187–195. [Google Scholar] [CrossRef]
- Lassance, L.; Marino, G.K.; Medeiros, C.S.; Thangavadivel, S.; Wilson, S.E. Fibrocyte migration, differentiation, and apoptosis apoptosis during the corneal wound healing response to injury. Exp. Eye Res. 2018, 170, 177–187. [Google Scholar] [CrossRef]
- de Oliveira, R.C.; Wilson, S.E. Fibrocytes, wound healing and corneal fibrosis. Invest. Ophthalmol. Vis. Sci. 2020, 61, 28–35. [Google Scholar] [CrossRef]
- Sampaio, L.P.; Hilgert, G.S.L.; Shiju, T.M.; Santhiago, M.R.; Wilson, S.E. Topical losartan and corticosteroid additively inhibit corneal stromal myofibroblast generation and scarring fibrosis after alkali burn injury. Trans. Vis. Sci. Tech. 2022, 11, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Naik-Mathuria, B.; Pilling, D.; Crawford, J.R.; Gay, A.N.; Smith, C.W.; Gomer, R.H.; Olutoye, O.O. Serum amyloid P inhibits dermal wound healing. Wound Repair Regen. 2008, 16, 266–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, S.E. Defective perlecan-associated basement membrane regeneration and altered modulation of transforming growth factor beta in fibrosis. Cell Molec. Life Sci 2022, 79, 144. [Google Scholar] [CrossRef]
- Wilson, S.E.; Walker, J.W.; Chwang, E. L; He, Y-G. Hepatocyte growth factor (HGF), keratinocyte growth factor (KGF), their receptors, FGF receptor-2, and the cells of the cornea. Invest. Oph thalmol. Vis. Sci 1993, 34, 2544–2561. [Google Scholar]
- Wilson, S.E.; He, Y.-G.; Weng, J.; Zieske, J.D.; Jester, J.V.; Schultz, G.S. Effect of epidermal growth factor, hepatocyte growth factor, and keratinocyte growth factor, on proliferation, motility, and differentiation of human corneal epithelial cells. Exp. Eye Res. 1994, 59, 665–678. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.E. Biology of keratorefractive surgery—PRK, PTK, LASIK, SMILE, inlays and other refractive surgeries. Exp. Eye Res. 2020, 198, 108136. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.-W.; Liu, J.; Lee, J.-S.; Mohan, R.R.; Mohan, R.R.; Woods, D.J.; He, Y.-G.; Wilson, S.E. Pro–inflammatory chemokine induction in keratocytes and inflammatory cell infiltration into the cornea. Invest. Ophthalmol. Vis. Sci. 2001, 42, 2795–2803. [Google Scholar]
- Singh, V.; Agrawal, V.; Santhiago, M.R.; Wilson, S.E. Stromal fibroblast-bone marrow-derived cell interactions: Implications for myofibroblast development in the cornea. Exp. Eye Res. 2012, 98, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.; Barbosa, F.L.; Torricelli, A.A.M.; Santhiago, M.R.; Wilson, S.E. Transforming growth factor β and platelet-derived growth factor modulation of myofibroblast development from corneal fibroblasts in vitro. Exp. Eye Res. 2014, 120, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Wilson, S.E. Corneal myofibroblasts and fibrosis. Exp. Eye Res. 2020, 201, 108272. [Google Scholar] [CrossRef]
- Jester, J.V.; Moller-Pedersen, T.; Huang, J.; Sax, C.M.; Kays, W.T.; Cavangh, H.D.; Petroll, W.M.; Piatigorsky, J.J. The cellular basis of corneal transparency: Evidence for ‘corneal crystallins’. Cell Sci. 1999, 112 Pt 5, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Bucala, R.; Spiegel, L.A.; Chesney, J.; Hogan, M.; Cerami, A. Circulating fibrocytes define a new leukocyte subpopulation that mediates tissue repair. Mol. Med. 1994, 1, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Abe, R.; Donnelly, S.C.; Peng, T.; Bucala, R.; Metz, C.N. Peripheral blood fibrocytes: Differentiation pathway and migration to wound sites. J. Immunol. 2001, 166, 7556–7562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellini, A.; Mattoli, S. The role of the fibrocyte, a bone marrow-derived mesenchymal progenitor, in reactive and reparative fibroses. Lab. Invest. 2007, 87, 858–870. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, F.L.; Chaurasia, S.; Cutler, A.; Asosingh, K.; Kaur, H.; de Medeiros, F.W.; Agrawal, V.; Wilson, S.E. Corneal myofibroblast generation from bone marrow-derived cells. Exp. Eye Res. 2010, 91, 92–96. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Scott, P.G.; Giuffre, J.; Shankowsky, H.A.; Ghahary, A.; Tredget, E.E. Peripheral blood fibrocytes from burn patients: Identification and quantification of fibrocytes in adherent cells cultured from peripheral blood mononuclear cells. Lab. Invest. 2002, 82, 1183–1192. [Google Scholar] [CrossRef] [Green Version]
- Pilling, D.; Buckley, C.D.; Salmon, M.; Gomer, R.H. Inhibition of fibrocyte differentiation by serum amyloid P. J. Immunol. 2003, 171, 5537–5546. [Google Scholar] [CrossRef] [Green Version]
- Pilling, D.; Tucker, N.M.; Gomer, R.H. Aggregated IgG inhibits the differentiation of human fibrocytes. J. Leukoc. Biol. 2006, 79, 1242–1251. [Google Scholar] [CrossRef] [Green Version]
- Varcoe, R.L.; Mikhail, M.; Guiffre, A.K.; Pennings, G.; Vicaretti, M.; Hawthorne, W.J.; Fletcher, J.P.; Medbury, H.J. The role of the fibrocyte in intimal hyperplasia. J. Thromb. Haemost. 2006, 4, 1125–1133. [Google Scholar] [CrossRef]
- Sunderkotter, C.; Nikolic, T.; Dillon, M.J.; Van Rooijen, N.; Stehling, M.; Drevets, D.A.; Leenen, P.J. Subpopulations of mouse blood monocytes differ in maturation stage and inflammatory response. J. Immunol. 2004, 172, 4410–4417. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S.; Taylor, P.R. Monocyte and macrophage heterogeneity. Nat. Rev. Immunol. 2005, 5, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Tacke, F.; Randolph, G.J. Migratory fate and differentiation of blood monocyte subsets. Immunobiology 2006, 211, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Glesne, D.; Huberman, E. A human peripheral blood monocyte-derived subset acts as pluripotent stem cells. Proc. Natl. Acad. Sci. USA 2003, 100, 2426–2431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuwana, M.; Okazaki, Y.; Kodama, H.; Izumi, K.; Yasuoka, H.; Ogawa, Y.; Kawakami, Y.; Ikeda, Y. Human circulating CD14+ monocytes as a source of progenitors that exhibit mesenchymal cell differentiation. J. Leukoc. Biol. 2003, 74, 833–845. [Google Scholar] [CrossRef]
- Pilling, D.; Gomer, R.H. Regulatory pathways for fibrocyte differentiation. In Fibrocytes: New Insights Into Tissue Repair and Systemic Fibroses; Bucala, R., Ed.; World Scientific Publishing Co. Pte. Ltd.: Singapore, 2007; pp. 37–60. [Google Scholar]
- Schmidt, M.; Sun, G.; Stacey, M.A.; Mori, L.; Mattoli, S. Identification of circulating fibrocytes as precursors of bronchial myofibroblasts in asthma. J. Immunol. 2003, 171, 380–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saikia, P.; Crabb, J.S.; Dibbin, L.L.; Juszczak, M.J.; Willard, B.; Jang, G.-F.; Shiju, M.T.; Crabb, J.W.; Wilson, S.E. Quantitative proteomic comparison of myofibroblasts derived from bone marrow or locally from the cornea. Sci. Rep. 2020, 10, 16717. [Google Scholar] [CrossRef]
- Gressner, A.M. Transdifferentiation of hepatic stellate cells (Ito cells) to myofibroblasts: A key event in hepatic fibrogenesis. Kidney Int. Suppl. 1996, 54, S39–S45. [Google Scholar]
- Rojas, A.; Chang, F.C.; Lin, S.L.; Duffield, J.S. The role played by perivascular cells in kidney interstitial injury. Clin. Nephrol. 2012, 77, 400–408. [Google Scholar] [CrossRef] [Green Version]
- Bargagna-Mohan, P.; Ishii, A.; Lei, L.; Sheehy, D.; Pandit, S.; Chan, G.; Bansal, R.; Mohan, R. Sustained activation of ERK1/2 MAPK in Schwann cells causes corneal neurofibroma. J. Neurosci. Res. 2017, 95, 1712–1729. [Google Scholar] [CrossRef] [Green Version]
- Jester, J.V.; Rodrigues, M.M.; Herman, I.M. Characterization of avascular corneal wound healing fibroblasts. New insights into the myofibroblast. Am. J. Pathol. 1987, 127, 140–148. [Google Scholar]
- Jester, J.V.; Petroll, W.M.; Barry, P.A.; Cavanagh, H.D. Expression of alpha-smooth muscle (alpha-SM) actin during corneal stromal wound healing. Invest. Ophthalmol. Vis. Sci. 1995, 36, 809–819. [Google Scholar] [PubMed]
- Ishizaki, M.; Wakamatsu, K.; Matsunami, T.; Yamanaka, N.; Saiga, T.; Shimizu, Y.; Zhu, G.; Kao, W.W. Dynamics of the expression of cytoskeleton components and adherens molecules by fibroblastic cells in alkali-burned and lacerated corneas. Exp. Eye Res. 1994, 59, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Masur, S.K.; Dewal, H.S.; Dinh, T.T.; Erenburg, I.; Petridou, S. Myofibroblasts differentiate from fibroblasts when plated at low density. Proc. Natl. Acad. Sci. USA 1996, 93, 4219–4223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffield, J.S.; Lupher, M.; Thannickal, V.J.; Wynn, T.A. Host responses in tissue repair and fibrosis. Annu. Rev. Pathol. 2013, 8, 241–276. [Google Scholar] [CrossRef]
- Šalingová, B.; Červenák, Z.; Adamičková, A.; Chomanicová, N.; Valášková, S.; Gažová, A.; Kyselovič, J. Endothelial-Mesenchymal Transition or Functional Tissue Regeneration-Two Outcomes of Heart Remodeling. Physiol. Res. 2021, 70, S13–S20. [Google Scholar] [CrossRef]
- Chaurasia, S.; Kaur, H.; Medeiros, F.W.; Smith, S.D.; Wilson, S.E. Dynamics of the expression of intermediate filaments vimentin and desmin during myofibroblast differentiation after corneal injury. Exp. Eye Res. 2009, 89, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Marino, G.K.; Santhiago, M.R.; Santhanam, A.; Lassance, L.; Thangavadivel, S.; Medeiros, C.S.; Torricelli, A.A.M.; Wilson, S.E. Regeneration of defective epithelial basement membrane and restoration of corneal transparency. J. Ref. Surg. 2017, 33, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Marino, G.K.; Santhiago, M.R.; Santhanam, A.; Lassance, L.; Thangavadivel, S.; Medeiros, C.S.; Bose, K.; Tam, K.P.; Wilson, S.E. Epithelial basement membrane injury and regeneration modulates corneal fibrosis after pseudomonas corneal ulcers in rabbits. Exp. Eye Res. 2017, 161, 101–105. [Google Scholar] [CrossRef]
- Wilson, S.E.; He, Y.-G.; Weng, J.; Li, Q.; McDowall, A.W.; Vital, M.; Chwang, E.L. Epithelial injury induces keratocyte apoptosis: Hypothesized role for the interleukin-1 system in the modulation of corneal tissue organization and wound healing. Exp. Eye Res. 1996, 62, 325–328. [Google Scholar] [CrossRef]
- Medeiros, C.S.; Lassance, L.; Saikia, P.; Wilson, S.E. Posterior stromal keratocyte apoptosis triggered by mechanical endothelial injury and nidogen-1 production in the cornea. Exp. Eye Res. 2018, 172, 30–35. [Google Scholar] [CrossRef]
- Wilson, S.E.; Li, Q.; Weng, J.; Barry-Lane, P.A.; Jester, J.V.; Liang, Q.; Wordinger, R.J. The Fas/Fas ligand system and other modulators of apoptosis in the cornea. Invest. Ophthalmol. Vis. Sci. 1996, 37, 1582–1592. [Google Scholar] [PubMed]
- Mohan, R.R.; Liang, Q.; Kim, W.-J.; Helena, M.C.; Baerveldt, F.; Wilson, S.E. Apoptosis in the cornea: Further characterization of Fas-Fas ligand system. Exp. Eye Res. 1997, 65, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, C.S.; Saikia, P.; de Oliveira, R.C.; Lassance, L.; Santhiago, M.R.; Wilson, S.E. Descemet’s membrane modulation of posterior corneal fibrosis. Invest. Ophth. Vis. Sci. 2019, 60, 1010–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zieske, J.D.; Guimarães, S.R.; Hutcheon, A.E. Kinetics of keratocyte proliferation in response to epithelial debridement. Exp. Eye Res. 2001, 72, 33–39. [Google Scholar] [CrossRef]
- Mohan, R.R.; Hutcheon, A.E.; Choi, R.; Hong, J.; Lee, J.; Mohan, R.R.; Ambrósio, R., Jr.; Zieske, J.D.; Wilson, S.E. Apoptosis, necrosis, proliferation, and myofibroblast generation in the stroma following LASIK and PRK. Exp. Eye Res. 2003, 76, 71–87. [Google Scholar] [CrossRef]
- Sampaio, L.P.; Hilgert, G.S.L.; Shiju, T.M.; Santhiago, M.R.; Wilson, S.E. Losartan inhibition of myofibroblast generation and late haze (scarring fibrosis) after PRK in rabbits. J. Ref. Surg. 2022, 38, 820–829. [Google Scholar] [CrossRef]
- Lipshitz, I.; Loewenstein, A.; Varssano, D.; Lazar, M. Late onset corneal haze after photorefractive keratectomy for moderate and high myopia. Ophthalmology. 1997, 104, 369–373. [Google Scholar] [CrossRef]
- Souza, A.L.P.; Ambrosio, R., Jr.; Bandeira, F.; Salomão, M.Q.; Lima, A.S.; Wilson, S.E. Topical losartan for treating corneal fibrosis (haze): First clinical experience. J. Ref. Surg. 2022, 38, 741–746. [Google Scholar] [CrossRef]
- Wilson, S.E. Topical losartan: Practical guidance for clinical trials in the prevention and treatment of corneal scarring fibrosis and other eye diseases and disorders. J. Ocul. Pharmacol. Ther. 2022; in press. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilson, S.E. The Yin and Yang of Mesenchymal Cells in the Corneal Stromal Fibrosis Response to Injury: The Cornea as a Model of Fibrosis in Other Organs. Biomolecules 2023, 13, 87. https://doi.org/10.3390/biom13010087
Wilson SE. The Yin and Yang of Mesenchymal Cells in the Corneal Stromal Fibrosis Response to Injury: The Cornea as a Model of Fibrosis in Other Organs. Biomolecules. 2023; 13(1):87. https://doi.org/10.3390/biom13010087
Chicago/Turabian StyleWilson, Steven E. 2023. "The Yin and Yang of Mesenchymal Cells in the Corneal Stromal Fibrosis Response to Injury: The Cornea as a Model of Fibrosis in Other Organs" Biomolecules 13, no. 1: 87. https://doi.org/10.3390/biom13010087