
First Evidence of the Protective Effects of 2-Pentadecyl-2-Oxazoline (PEA-OXA) in In Vitro Models of Acute Lung Injury
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Poly-(I:C)-Induced Inflammatory Response in A549 Cells
2.3. RNA Extraction and Quantitative PCR (qPCR)
2.4. Quantification by Liquid Chromatography–Atmospheric Pressure Chemical Ionization–Mass Spectrometry (LC-APCI-MS) of the Endogenous AEA, 2-AG and PEA Levels in A549 Cells
2.5. TGF-β-Induced Epithelial–Mesenchymal Transition in A549 Cells
2.6. Data Analysis
3. Results
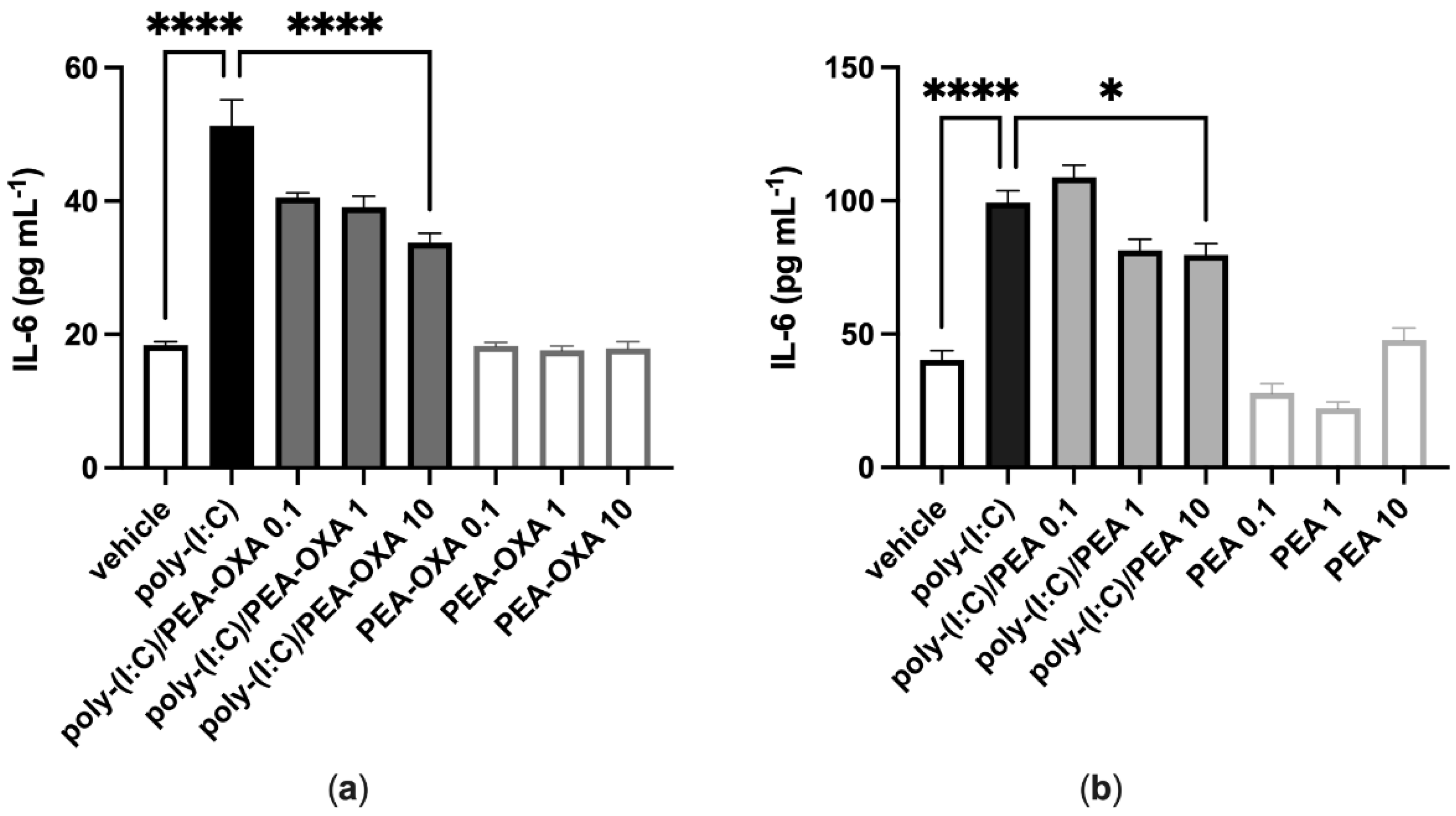
3.1. PEA-OXA and PEA Reduce Poly-(I:C)-Induced Release of Il-6 in Lung Epithelial Cells
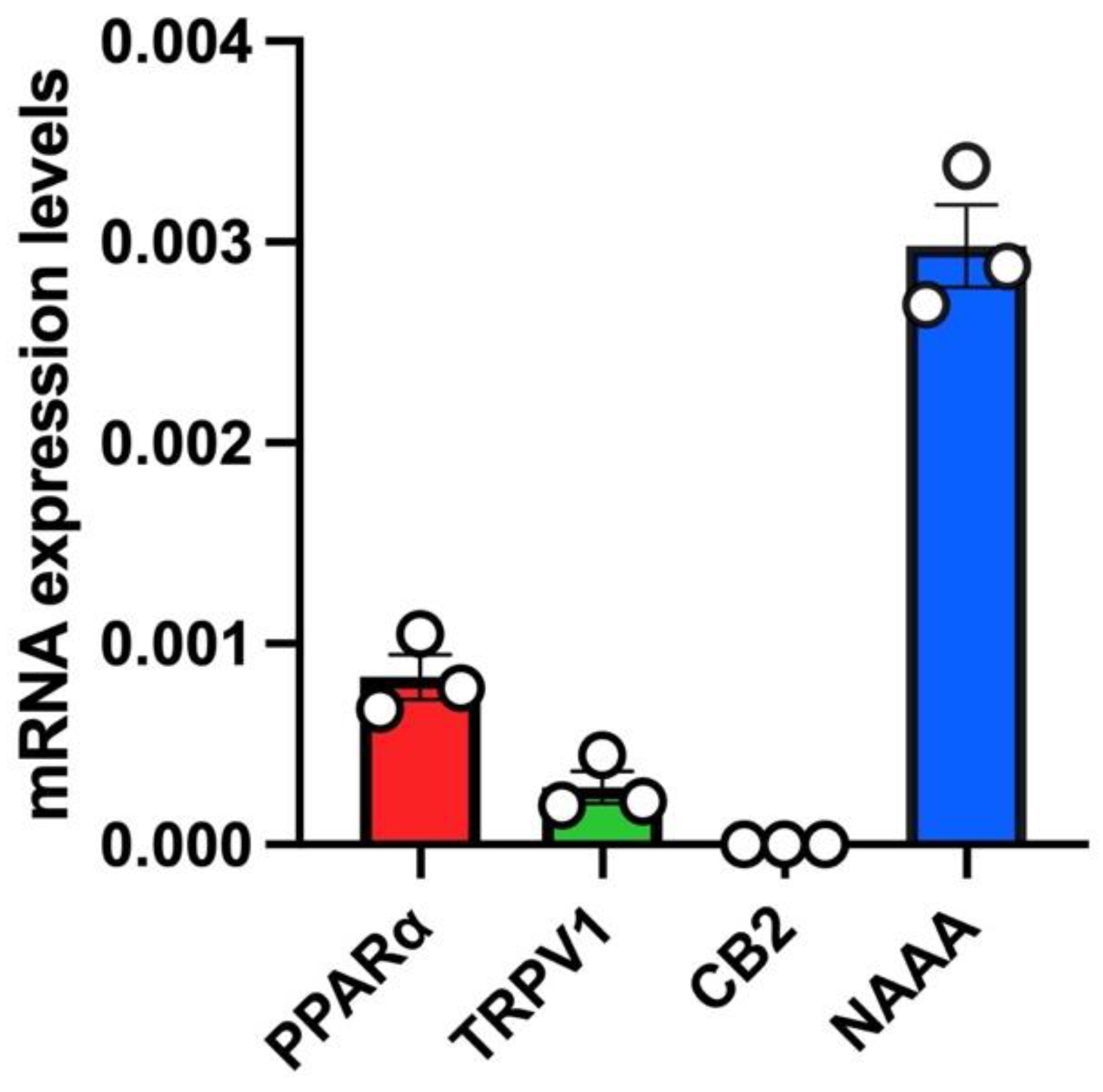
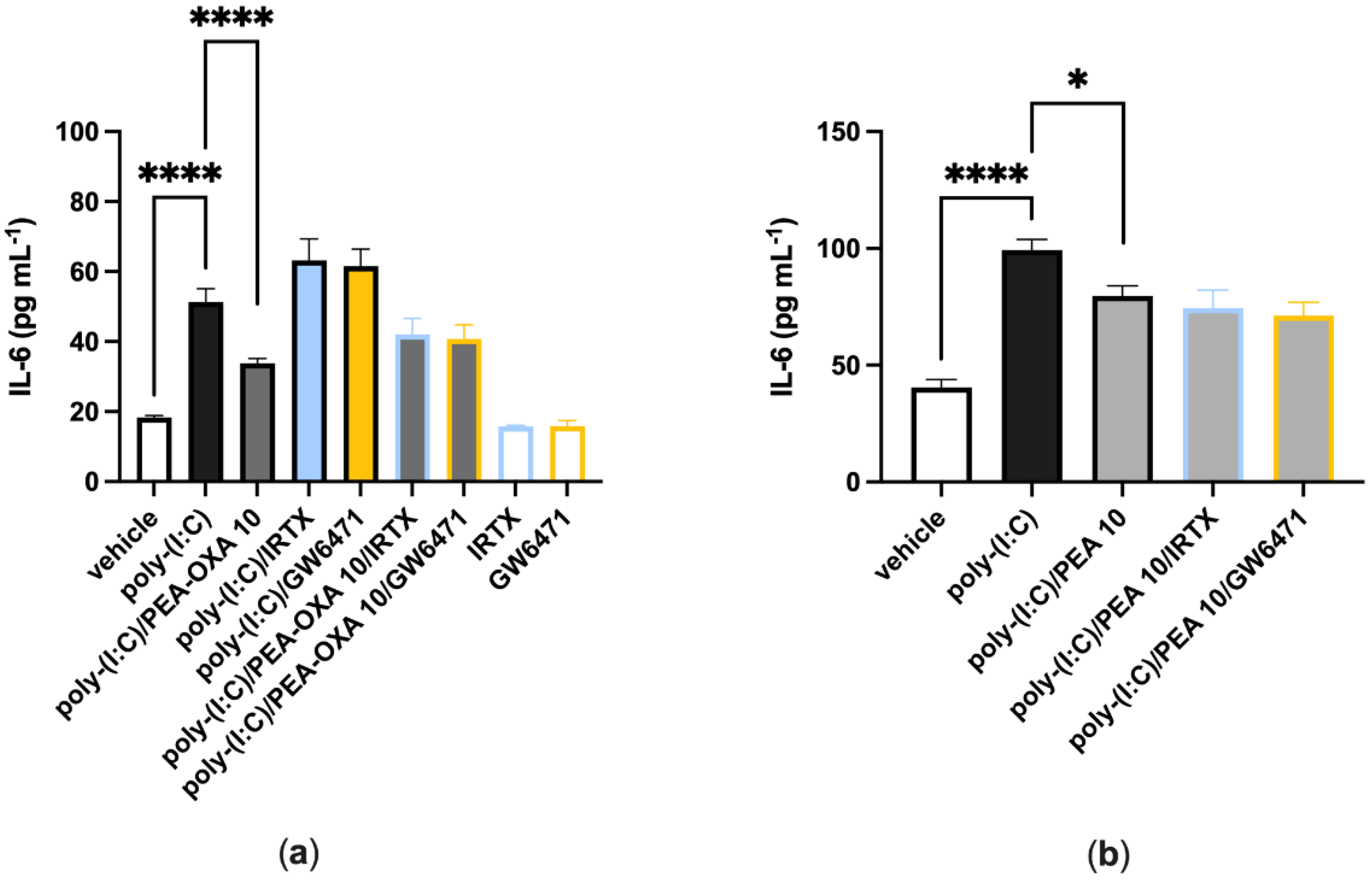
3.2. The Anti-Inflammatory Effect of PEA-OXA and PEA Is Not Reverted by Antagonism at the TRPV1 or PPAR-α Receptors in Lung Epithelial Cells
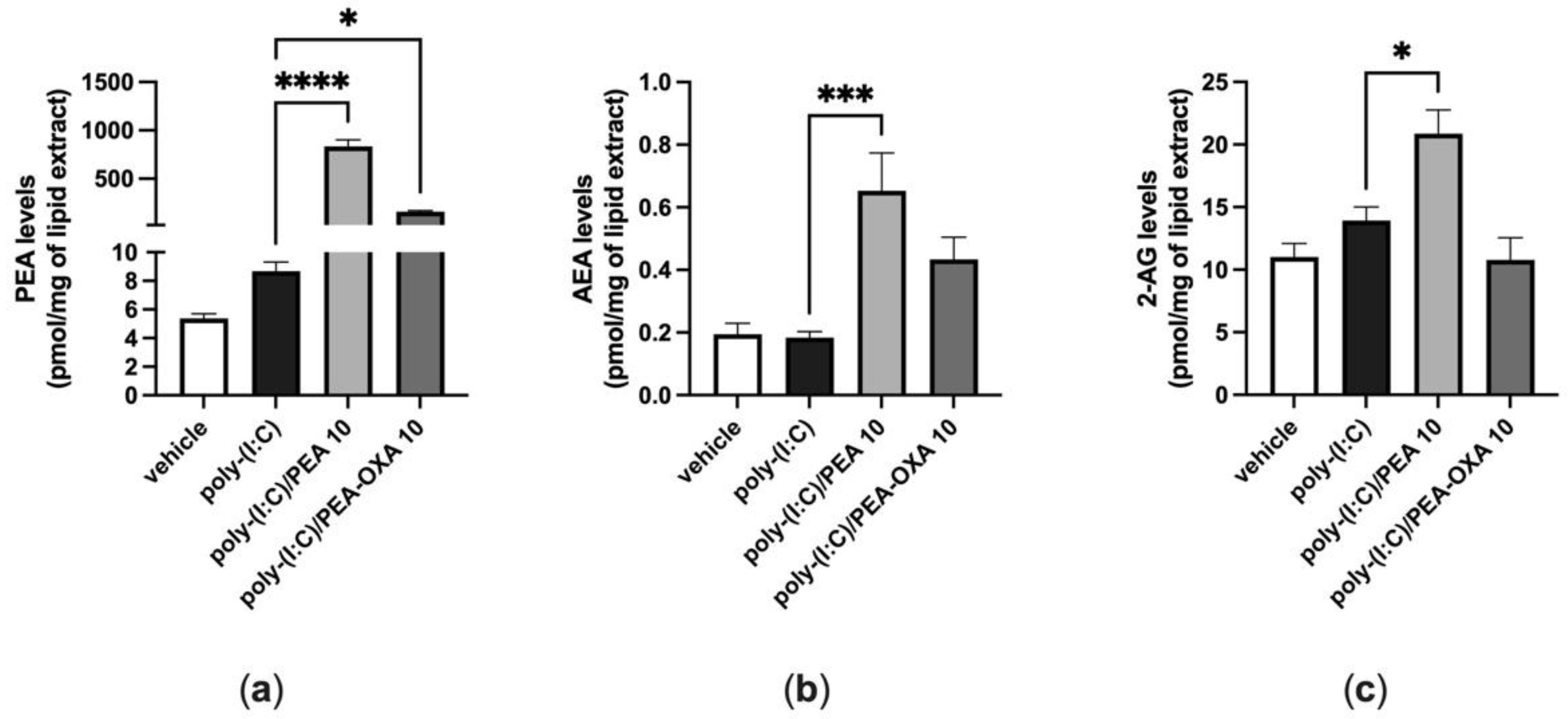
3.3. Effect of PEA and PEA-OXA Treatment on AEA, 2-AG and PEA Endogenous Levels, in Poly-(I:C)-Stimulated A549 Cells
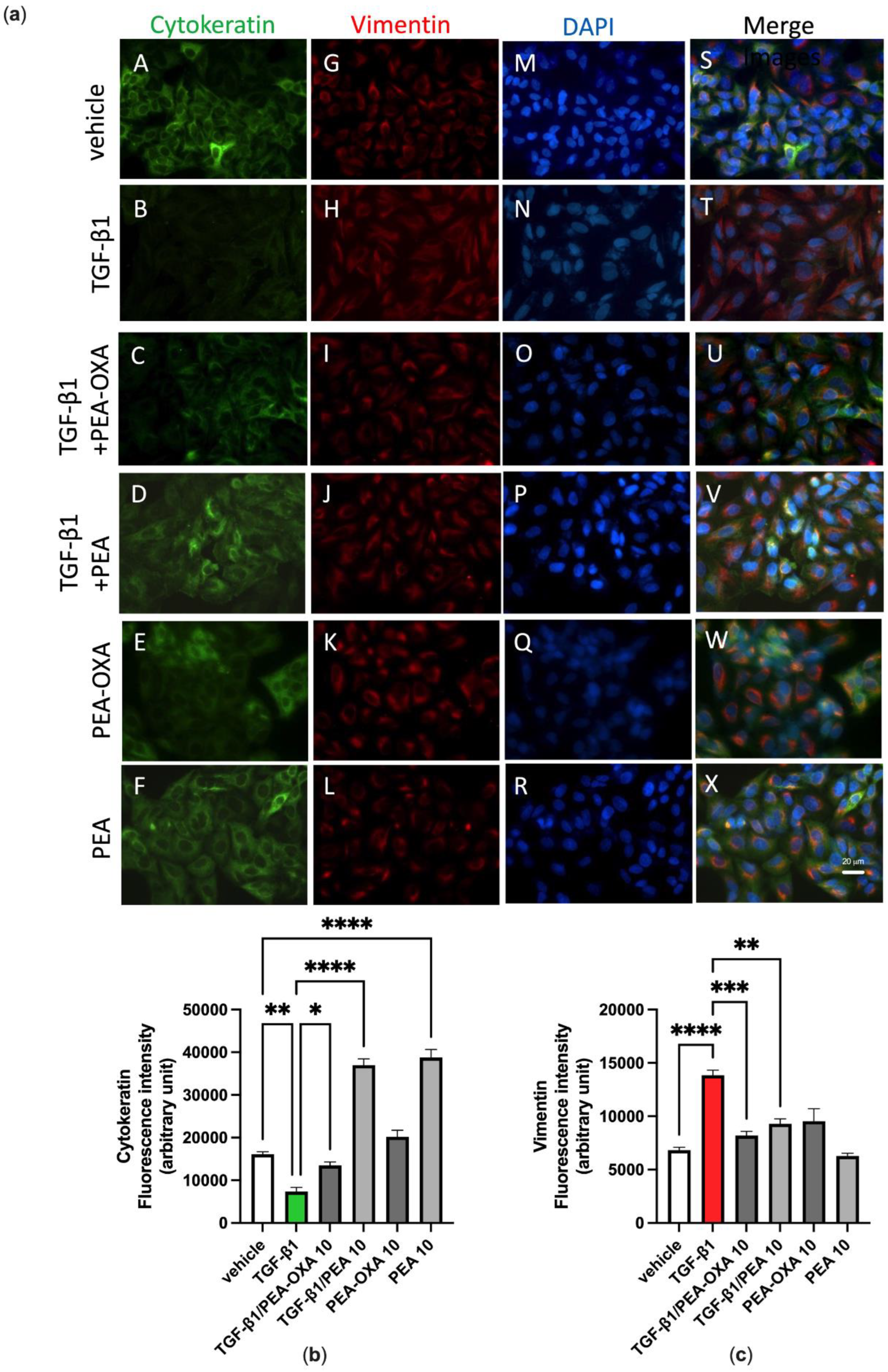
3.4. PEA-OXA and PEA Block TGF-β-Induced Epithelial–Mesenchymal Transition in Lung Epithelial Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Su, C.-F.; Kao, S.J.; Chen, H.I. Acute Respiratory Distress Syndrome and Lung Injury: Pathogenetic Mechanism and Therapeutic Implication. World J. Crit. Care Med. 2012, 1, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Rezoagli, E.; Fumagalli, R.; Bellani, G. Definition and Epidemiology of Acute Respiratory Distress Syndrome. Ann. Transl. Med. 2017, 5, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gragossian, A.; Siuba, M.T. Acute Respiratory Distress Syndrome. Emerg. Med. Clin. North Am. 2022, 40, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Khodadadi, H.; Salles, É.L.; Jarrahi, A.; Chibane, F.; Costigliola, V.; Yu, J.C.; Vaibhav, K.; Hess, D.C.; Dhandapani, K.M.; Baban, B. Cannabidiol Modulates Cytokine Storm in Acute Respiratory Distress Syndrome Induced by Simulated Viral Infection Using Synthetic RNA. Cannabi. annabinoid. Res. 2020, 5, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.H.; Kim, J.; Hong, S.P.; Choi, S.Y.; Yang, M.J.; Ju, Y.S.; Kim, Y.T.; Kim, H.M.; Rahman, M.D.T.; Chung, M.K.; et al. Nasal Ciliated Cells Are Primary Targets for SARS-CoV-2 Replication in the Early Stage of COVID-19. J. Clin. Invest. 2021, 131, 148517. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.J.; Okuda, K.; Edwards, C.E.; Martinez, D.R.; Asakura, T.; Dinnon, K.H.; Kato, T.; Lee, R.E.; Yount, B.L.; Mascenik, T.M.; et al. SARS-CoV-2 Reverse Genetics Reveals a Variable Infection Gradient in the Respiratory Tract. Cell 2020, 182, 429–446.e14. [Google Scholar] [CrossRef]
- Justet, A.; Zhao, A.Y.; Kaminski, N. From COVID to Fibrosis: Lessons from Single-Cell Analyses of the Human Lung. Hum. Genomics 2022, 16, 20. [Google Scholar] [CrossRef]
- Lamers, M.M.; Haagmans, B.L. SARS-CoV-2 Pathogenesis. Nat. Rev. Microbiol. 2022, 20, 270–284. [Google Scholar] [CrossRef]
- Petrosino, S.; Schiano Moriello, A. Palmitoylethanolamide: A Nutritional Approach to Keep Neuroinflammation within Physiological Boundaries-A Systematic Review. Int. J. Mol. Sci. 2020, 21, 9526. [Google Scholar] [CrossRef]
- Petrosino, S.; Di Marzo, V. The Pharmacology of Palmitoylethanolamide and First Data on the Therapeutic Efficacy of Some of Its New Formulations. Br. J. Pharmacol. 2017, 174, 1349–1365. [Google Scholar] [CrossRef]
- Aloe, L.; Leon, A.; Levi-Montalcini, R. A Proposed Autacoid Mechanism Controlling Mastocyte Behaviour. Agents Actions 1993, 39 Spec. No., C145–C147. [Google Scholar] [CrossRef]
- Lo Verme, J.; Fu, J.; Astarita, G.; La Rana, G.; Russo, R.; Calignano, A.; Piomelli, D. The Nuclear Receptor Peroxisome Proliferator-Activated Receptor-Alpha Mediates the Anti-Inflammatory Actions of Palmitoylethanolamide. Mol. Pharmacol. 2005, 67, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Ryberg, E.; Larsson, N.; Sjögren, S.; Hjorth, S.; Hermansson, N.-O.; Leonova, J.; Elebring, T.; Nilsson, K.; Drmota, T.; Greasley, P.J. The Orphan Receptor GPR55 Is a Novel Cannabinoid Receptor. Br. J. Pharmacol. 2007, 152, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V.; Melck, D.; Orlando, P.; Bisogno, T.; Zagoory, O.; Bifulco, M.; Vogel, Z.; De Petrocellis, L. Palmitoylethanolamide Inhibits the Expression of Fatty Acid Amide Hydrolase and Enhances the Anti-Proliferative Effect of Anandamide in Human Breast Cancer Cells. Biochem. J. 2001, 358, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Petrosino, S.; Schiano Moriello, A.; Verde, R.; Allarà, M.; Imperatore, R.; Ligresti, A.; Mahmoud, A.M.; Peritore, A.F.; Iannotti, F.A.; Di Marzo, V. Palmitoylethanolamide Counteracts Substance P-Induced Mast Cell Activation in Vitro by Stimulating Diacylglycerol Lipase Activity. J. Neuroinflamm. 2019, 16, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, W.-S.V.; Barrett, D.A.; Randall, M.D. “Entourage” Effects of N-Palmitoylethanolamide and N-Oleoylethanolamide on Vasorelaxation to Anandamide Occur through TRPV1 Receptors. Br. J. Pharmacol. 2008, 155, 837–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Petrocellis, L.; Davis, J.B.; Di Marzo, V. Palmitoylethanolamide Enhances Anandamide Stimulation of Human Vanilloid VR1 Receptors. FEBS Lett. 2001, 506, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Petrosino, S.; Schiano Moriello, A.; Cerrato, S.; Fusco, M.; Puigdemont, A.; De Petrocellis, L.; Di Marzo, V. The Anti-Inflammatory Mediator Palmitoylethanolamide Enhances the Levels of 2-Arachidonoyl-Glycerol and Potentiates Its Actions at TRPV1 Cation Channels. Br. J. Pharmacol. 2016, 173, 1154–1162. [Google Scholar] [CrossRef] [Green Version]
- Iannotti, F.A.; Di Marzo, V.; Petrosino, S. Endocannabinoids and Endocannabinoid-Related Mediators: Targets, Metabolism and Role in Neurological Disorders. Prog. Lipid Res. 2016, 62, 107–128. [Google Scholar] [CrossRef]
- Skaper, S.D.; Di Marzo, V. Endocannabinoids in Nervous System Health and Disease: The Big Picture in a Nutshell. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2012, 367, 3193–3200. [Google Scholar] [CrossRef]
- Skaper, S.D.; Facci, L. Mast Cell-Glia Axis in Neuroinflammation and Therapeutic Potential of the Anandamide Congener Palmitoylethanolamide. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 3312–3325. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, D.; Cordaro, M.; Bruschetta, G.; Crupi, R.; Pascali, J.; Alfonsi, D.; Marcolongo, G.; Cuzzocrea, S. 2-Pentadecyl-2-Oxazoline: Identification in Coffee, Synthesis and Activity in a Rat Model of Carrageenan-Induced Hindpaw Inflammation. Pharmacol. Res. 2016, 108, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Petrosino, S.; Campolo, M.; Impellizzeri, D.; Paterniti, I.; Allarà, M.; Gugliandolo, E.; D’Amico, R.; Siracusa, R.; Cordaro, M.; Esposito, E.; et al. 2-Pentadecyl-2-Oxazoline, the Oxazoline of Pea, Modulates Carrageenan-Induced Acute Inflammation. Front. Pharmacol. 2017, 8, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordaro, M.; Siracusa, R.; Crupi, R.; Impellizzeri, D.; Peritore, A.F.; D’Amico, R.; Gugliandolo, E.; Di Paola, R.; Cuzzocrea, S. 2-Pentadecyl-2-Oxazoline Reduces Neuroinflammatory Environment in the MPTP Model of Parkinson Disease. Mol. Neurobiol. 2018, 55, 9251–9266. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Cordaro, M.; Bruschetta, G.; Siracusa, R.; Crupi, R.; Esposito, E.; Cuzzocrea, S. N-Palmitoylethanolamine-Oxazoline as a New Therapeutic Strategy to Control Neuroinflammation: Neuroprotective Effects in Experimental Models of Spinal Cord and Brain Injury. J. Neurotrauma 2017, 34, 2609–2623. [Google Scholar] [CrossRef]
- Aliyu, M.; Zohora, F.T.; Anka, A.U.; Ali, K.; Maleknia, S.; Saffarioun, M.; Azizi, G. Interleukin-6 Cytokine: An Overview of the Immune Regulation, Immune Dysregulation, and Therapeutic Approach. Int. Immunopharmacol. 2022, 111, 109130. [Google Scholar] [CrossRef]
- Rezaei Tolzali, M.M.; Noori, M.; Shokri, P.; Rahmani, S.; Khanzadeh, S.; Nejadghaderi, S.A.; Fazlollahi, A.; Sullman, M.J.M.; Singh, K.; Kolahi, A.-A.; et al. Efficacy of Tocilizumab in the Treatment of COVID-19: An Umbrella Review. Rev. Med. Virol. 2022, e2388. [Google Scholar] [CrossRef]
- Del Re, A.; Corpetti, C.; Pesce, M.; Seguella, L.; Steardo, L.; Palenca, I.; Rurgo, S.; De Conno, B.; Sarnelli, G.; Esposito, G. Ultramicronized Palmitoylethanolamide Inhibits NLRP3 Inflammasome Expression and Pro-Inflammatory Response Activated by SARS-CoV-2 Spike Protein in Cultured Murine Alveolar Macrophages. Metabolites 2021, 11, 592. [Google Scholar] [CrossRef]
- Albanese, M.; Marrone, G.; Paolino, A.; Di Lauro, M.; Di Daniele, F.; Chiaramonte, C.; D’Agostini, C.; Romani, A.; Cavaliere, A.; Guerriero, C.; et al. Effects of Ultramicronized Palmitoylethanolamide (Um-PEA) in COVID-19 Early Stages: A Case-Control Study. Pharmaceuticals 2022, 15, 253. [Google Scholar] [CrossRef]
- Di Marzo, V.; Goparaju, S.K.; Wang, L.; Liu, J.; Bátkai, S.; Járai, Z.; Fezza, F.; Miura, G.I.; Palmiter, R.D.; Sugiura, T.; et al. Leptin-Regulated Endocannabinoids Are Involved in Maintaining Food Intake. Nature 2001, 410, 822–825. [Google Scholar] [CrossRef]
- Marsicano, G.; Wotjak, C.T.; Azad, S.C.; Bisogno, T.; Rammes, G.; Cascio, M.G.; Hermann, H.; Tang, J.; Hofmann, C.; Zieglgänsberger, W.; et al. The Endogenous Cannabinoid System Controls Extinction of Aversive Memories. Nature 2002, 418, 530–534. [Google Scholar] [CrossRef] [PubMed]
- Riemma, M.A.; Cerqua, I.; Romano, B.; Irollo, E.; Bertolino, A.; Camerlingo, R.; Granato, E.; Rea, G.; Scala, S.; Terlizzi, M.; et al. Sphingosine-1-Phosphate/TGF-β Axis Drives Epithelial Mesenchymal Transition in Asthma-like Disease. Br. J. Pharmacol. 2022, 179, 1753–1768. [Google Scholar] [CrossRef] [PubMed]
- Peritore, A.F.; D’Amico, R.; Siracusa, R.; Cordaro, M.; Fusco, R.; Gugliandolo, E.; Genovese, T.; Crupi, R.; Di Paola, R.; Cuzzocrea, S.; et al. Management of Acute Lung Injury: Palmitoylethanolamide as a New Approach. Int. J. Mol. Sci. 2021, 22, 5533. [Google Scholar] [CrossRef] [PubMed]
- Stowell, N.C.; Seideman, J.; Raymond, H.A.; Smalley, K.A.; Lamb, R.J.; Egenolf, D.D.; Bugelski, P.J.; Murray, L.A.; Marsters, P.A.; Bunting, R.A.; et al. Long-Term Activation of TLR3 by Poly(I:C) Induces Inflammation and Impairs Lung Function in Mice. Respir. Res. 2009, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, K.; Takezaki, N.; Ueda, N. The N-Acylethanolamine-Hydrolyzing Acid Amidase (NAAA). Chem. Biodivers. 2007, 4, 1914–1925. [Google Scholar] [CrossRef]
- Di Paola, R.; Impellizzeri, D.; Fusco, R.; Cordaro, M.; Siracusa, R.; Crupi, R.; Esposito, E.; Cuzzocrea, S. Ultramicronized Palmitoylethanolamide (PEA-Um(®)) in the Treatment of Idiopathic Pulmonary Fibrosis. Pharmacol. Res. 2016, 111, 405–412. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| gene | Forward (5′–3′) | Reverse (5′–3′) |
|---|---|---|
| CNR2 | TAGTGCTGAGAGGACCCA | CGCTATCCACCTTCCTACAA |
| TRPV1 | CTGCCCGACCATCACAGTC | CTGCGATCATAGAGCCTGAGG |
| PPARα | TTCGCAATCCATCGGCGAG | CCACAGGATAAGTCACCGAGG |
| NAAA | TGACAGTGGATGTGCAATTCTT | GCCTTTATCTCGTTCATCACCAG |
| S16 | TCGGACGCAAGAAGACAGCGA | AGCGTGCGCGGCTCAATCAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiano Moriello, A.; Roviezzo, F.; Iannotti, F.A.; Rea, G.; Allarà, M.; Camerlingo, R.; Verde, R.; Di Marzo, V.; Petrosino, S. First Evidence of the Protective Effects of 2-Pentadecyl-2-Oxazoline (PEA-OXA) in In Vitro Models of Acute Lung Injury. Biomolecules 2023, 13, 33. https://doi.org/10.3390/biom13010033
Schiano Moriello A, Roviezzo F, Iannotti FA, Rea G, Allarà M, Camerlingo R, Verde R, Di Marzo V, Petrosino S. First Evidence of the Protective Effects of 2-Pentadecyl-2-Oxazoline (PEA-OXA) in In Vitro Models of Acute Lung Injury. Biomolecules. 2023; 13(1):33. https://doi.org/10.3390/biom13010033
Chicago/Turabian StyleSchiano Moriello, Aniello, Fiorentina Roviezzo, Fabio Arturo Iannotti, Giuseppina Rea, Marco Allarà, Rosa Camerlingo, Roberta Verde, Vincenzo Di Marzo, and Stefania Petrosino. 2023. "First Evidence of the Protective Effects of 2-Pentadecyl-2-Oxazoline (PEA-OXA) in In Vitro Models of Acute Lung Injury" Biomolecules 13, no. 1: 33. https://doi.org/10.3390/biom13010033