
Ultramicronized N-Palmitoylethanolamine Regulates Mast Cell-Astrocyte Crosstalk: A New Potential Mechanism Underlying the Inhibition of Morphine Tolerance
 , , , , , , , ,
, , , , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Cell Culture
2.4. Cell Viability Assay
2.5. Cell Count
2.6. β-hexosaminidase Assay
2.7. Histamine Assay
2.8. Collection of Mast Cell-Conditioned Medium
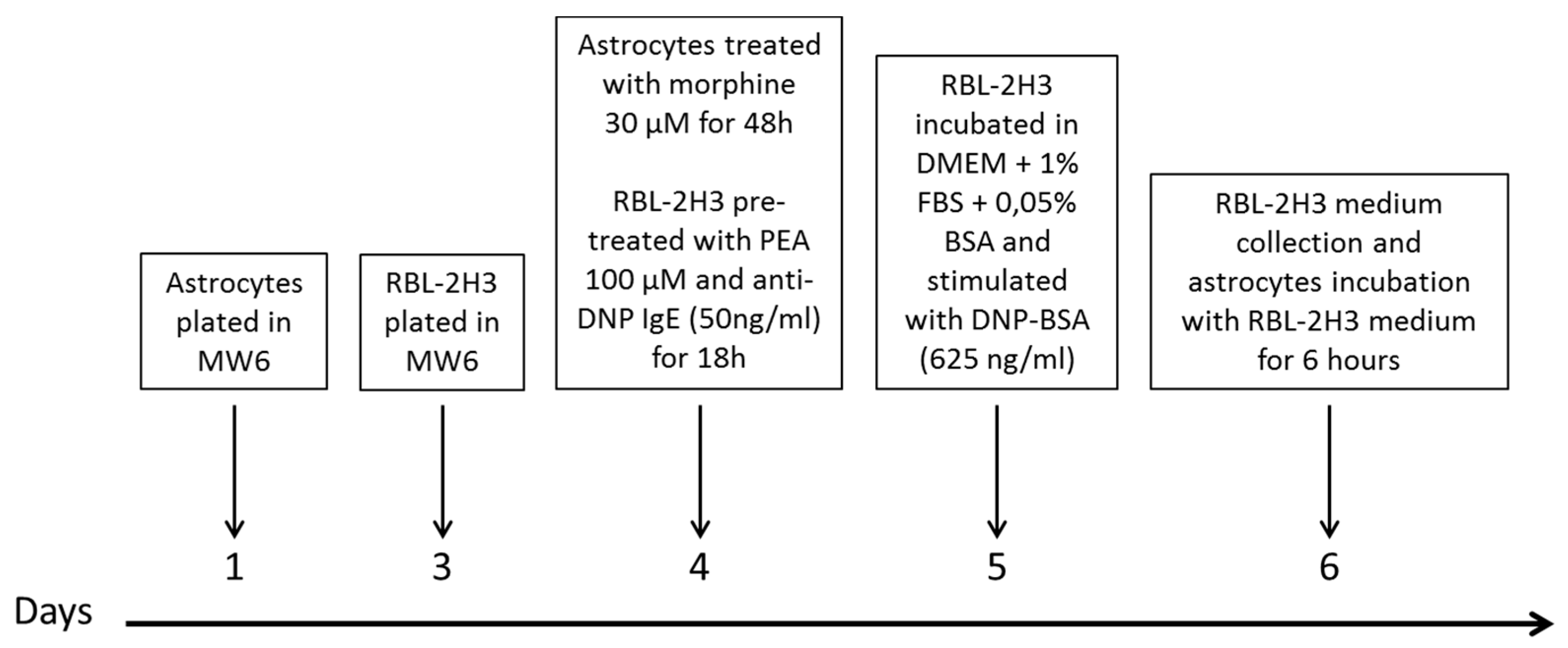
2.9. Timeline of Astrocytes Treatment with Mast Cell-Conditioned Medium
2.10. Protein Extraction, Electrophoresis, and Western Blot
2.11. Immunofluorescence
2.12. Real Time Polymerase Chain Reaction (RT-PCR)
2.13. Measurement of Intracellular Ca2+
2.14. Statistical Analysis
3. Results
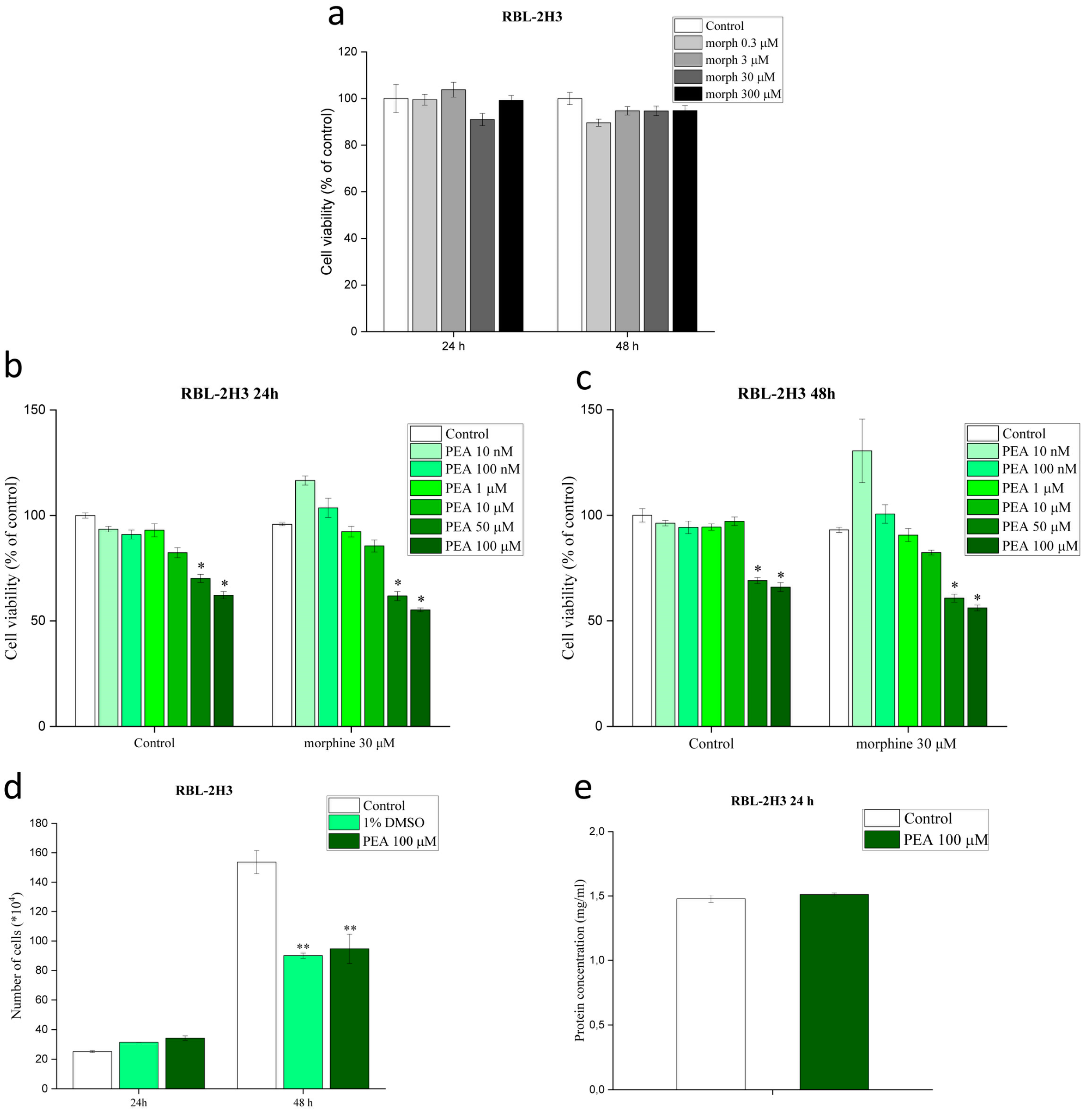
3.1. Morphine and PEA, Used Solely or in Combination, Do Not Affect Mast Cell Viability
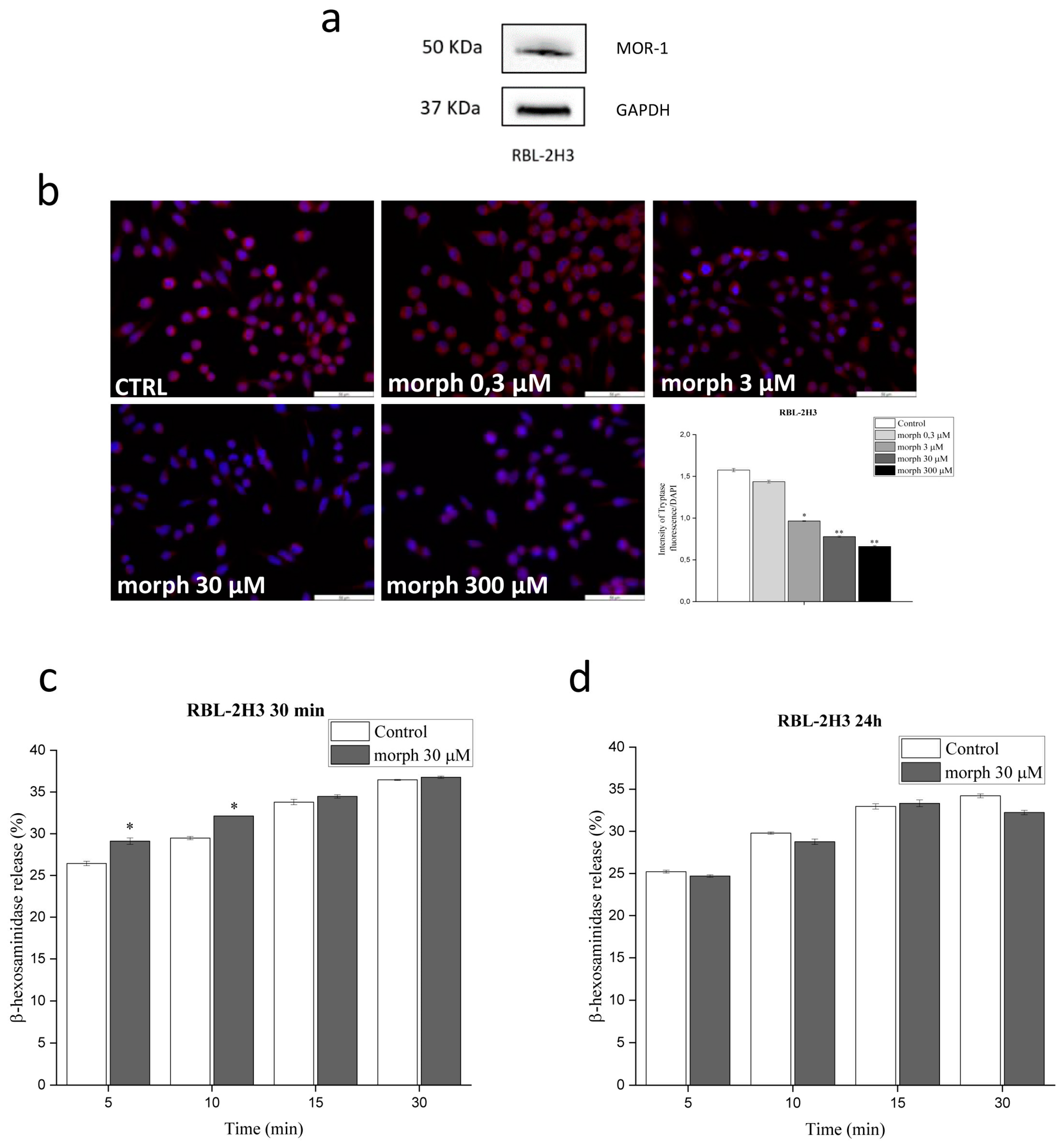
3.2. RBL-2H3 Cells Express the µ-Receptor and Morphine Augments Mast Cell Degranulation
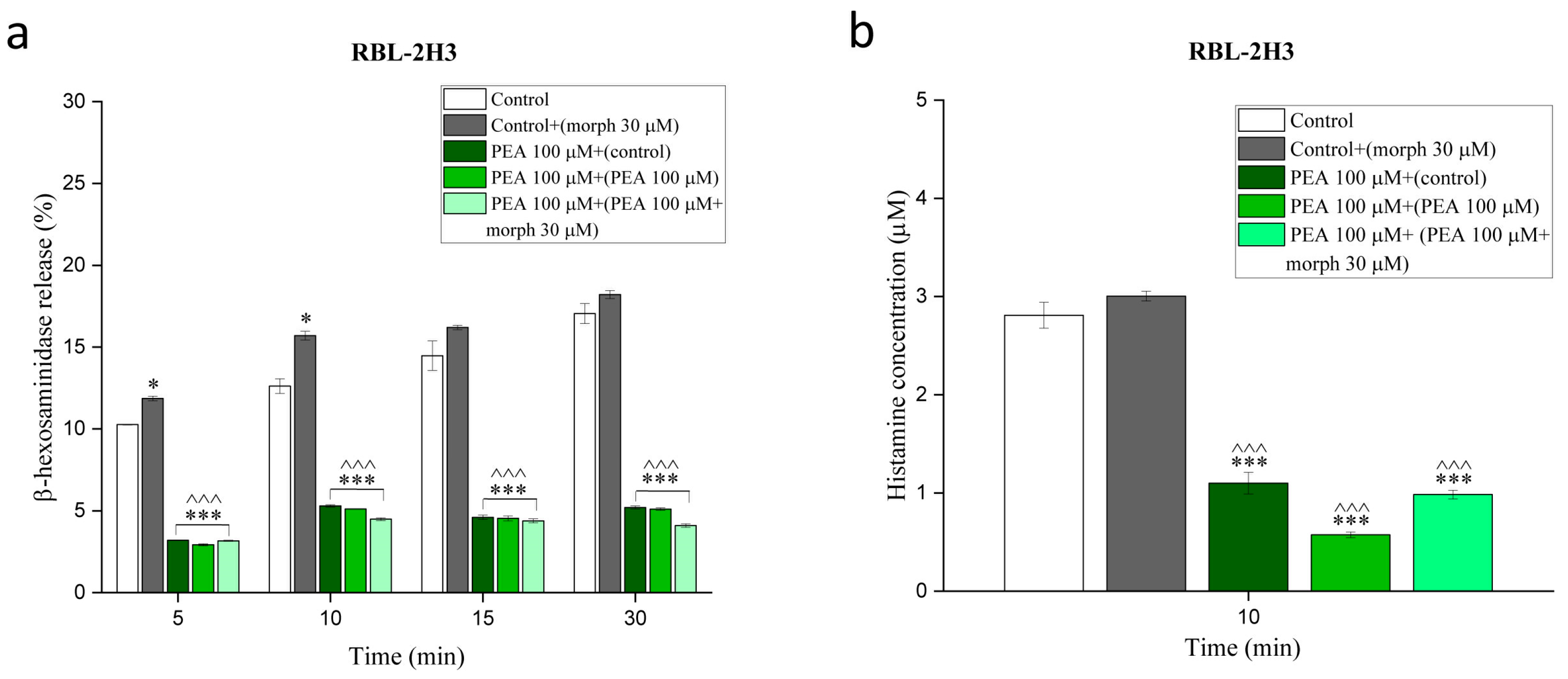
3.3. PEA Counteracts Morphine-Augmented Mast Cell Degranulation
3.4. PEA Down-Modulation of Mast Cell Degranulation Is Long-Lasting
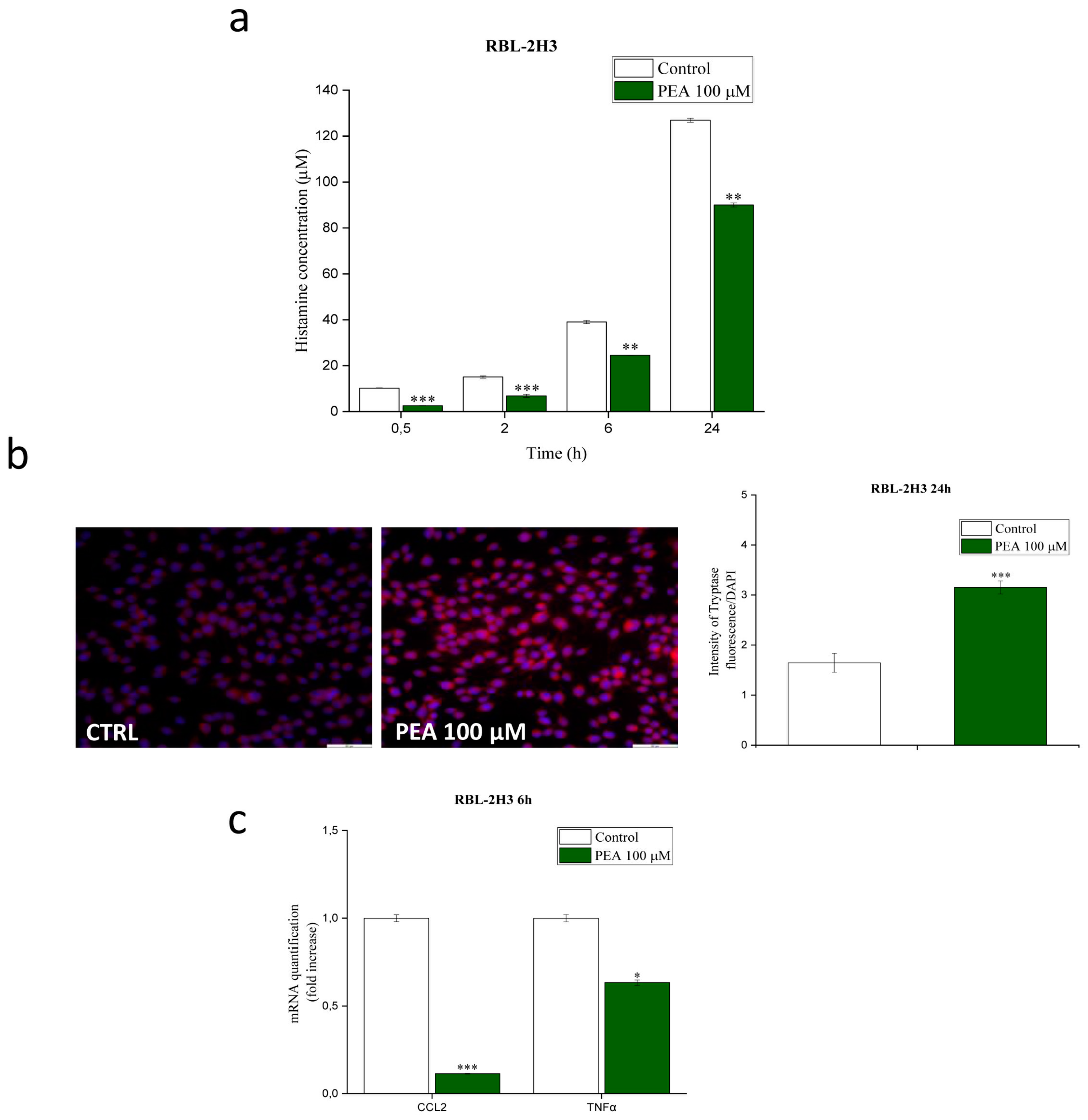
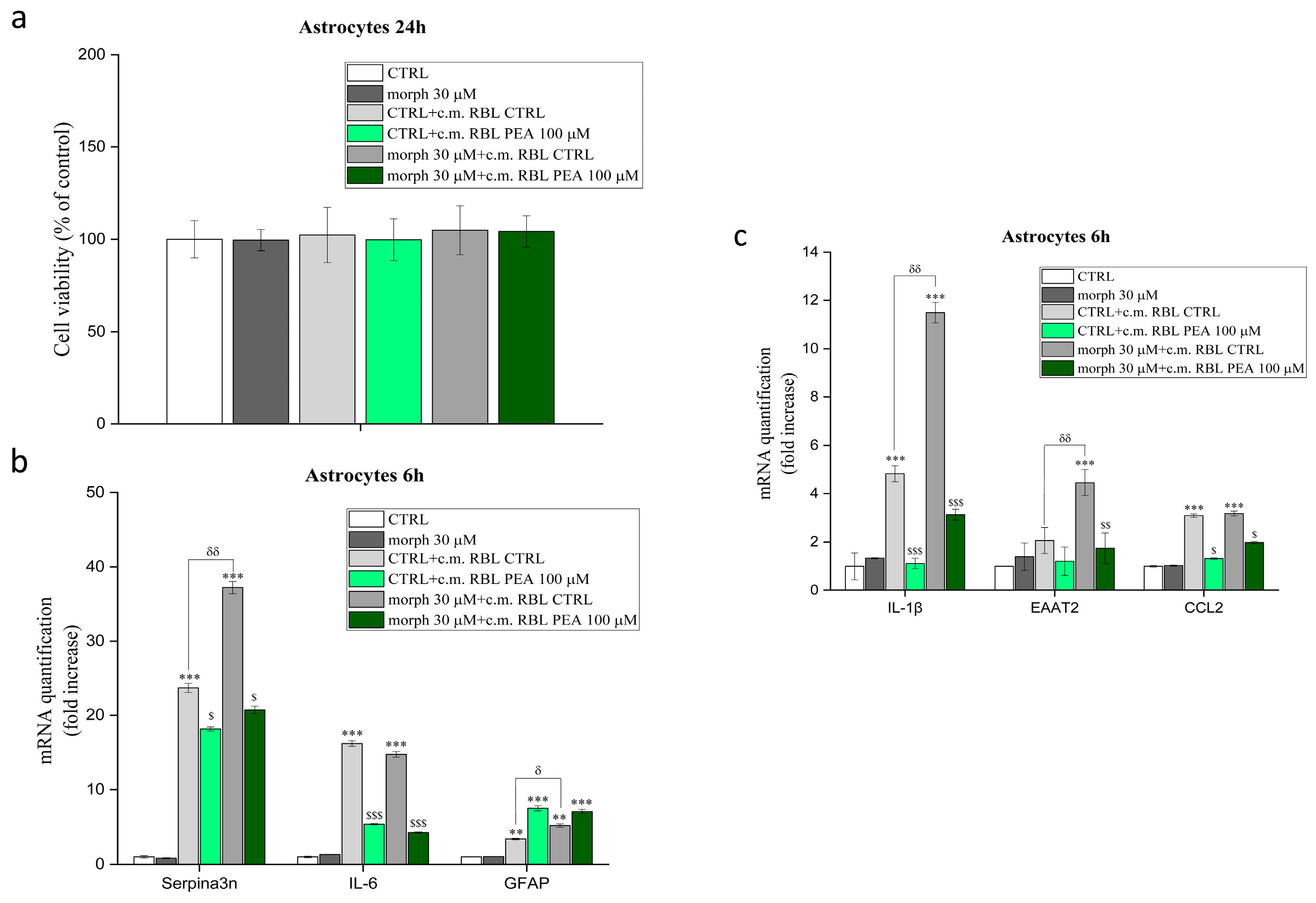
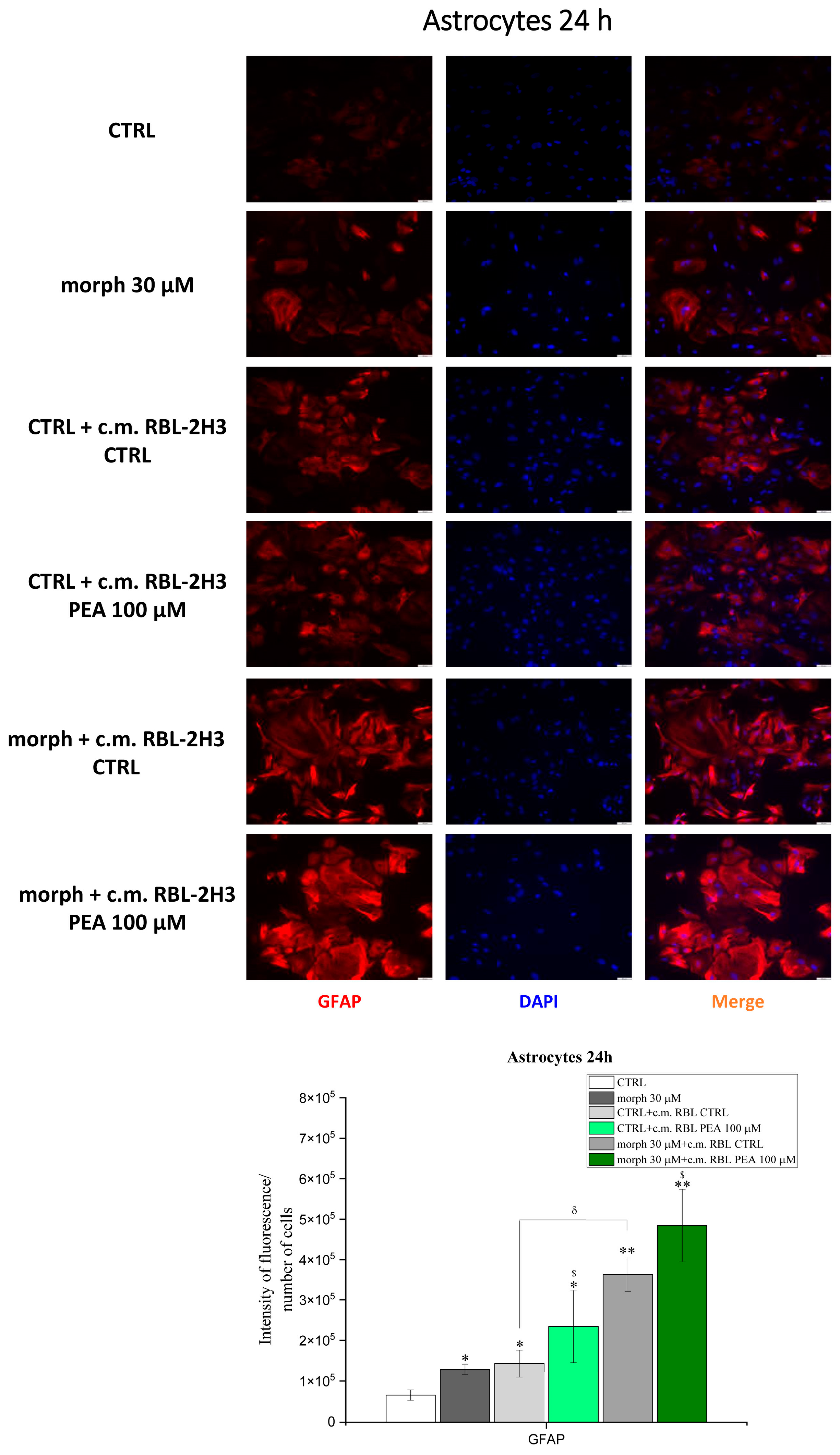
3.5. PEA Down-Modulates Mast Cell-Induced Astrocyte Expression of Genes Involved in Inflammation and Pain
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gatchel, R.J.; McGeary, D.D.; McGeary, C.A.; Lippe, B. Interdisciplinary Chronic Pain Management: Past, Present, and Future. Am. Psychol. 2014, 69, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Yaksh, T.L. Pain after Injury: Some Basic Mechanisms. J. Fla. Med. Assoc. 1997, 84, 16–19. [Google Scholar]
- White, P.F.; Kehlet, H. Improving Postoperative Pain Management: What Are the Unresolved Issues? Anesthesiology 2010, 112, 220–225. [Google Scholar] [CrossRef] [Green Version]
- Sierra-Silvestre, E.; Somerville, M.; Bisset, L.; Coppieters, M.W. Altered Pain Processing in Patients with Type 1 and 2 Diabetes: Systematic Review and Meta-Analysis of Pain Detection Thresholds and Pain Modulation Mechanisms. BMJ Open Diabetes Res. Care 2020, 8, e001566. [Google Scholar] [CrossRef] [PubMed]
- Dieppe, P.A.; Lohmander, L.S. Pathogenesis and Management of Pain in Osteoarthritis. Lancet 2005, 365, 965–973. [Google Scholar] [CrossRef]
- Portenoy, R.K.; Ahmed, E. Cancer Pain Syndromes. Hematol. Oncol. Clin. N. Am. 2018, 32, 371–386. [Google Scholar] [CrossRef]
- Farquhar-Smith, P. Chemotherapy-Induced Neuropathic Pain. Curr. Opin. Support. Palliat. Care 2011, 5, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Shaheed, C.A.; Machado, G.C.; Underwood, M. Drugs for Chronic Pain. Br. J. Gen. Pract. 2020, 70, 576–577. [Google Scholar] [CrossRef]
- Haghjooy-Javanmard, S.; Ghasemi, A.; Laher, I.; Zarrin, B.; Dana, N.; Vaseghi, G. Influence of Morphine on TLR4/NF-KB Signaling Pathway of MCF-7 Cells. Bratisl. Lek. Listy 2018, 119, 229–233. [Google Scholar] [CrossRef]
- Ghelardini, C.; Di Cesare Mannelli, L.; Bianchi, E. The Pharmacological Basis of Opioids. Clin. Cases Miner. Bone Metab. 2015, 12, 219–221. [Google Scholar] [CrossRef]
- Vanegas, G.; Ripamonti, C.; Sbanotto, A.; de Conno, F. Side Effects of Morphine Administration in Cancer Patients. Cancer Nurs. 1998, 21, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Dumas, E.O.; Pollack, G.M. Opioid Tolerance Development: A Pharmacokinetic/Pharmacodynamic Perspective. AAPS J. 2008, 10, 537–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mika, J.; Wawrzczak-Bargiela, A.; Osikowicz, M.; Makuch, W.; Przewlocka, B. Attenuation of Morphine Tolerance by Minocycline and Pentoxifylline in Naive and Neuropathic Mice. Brain Behav. Immun. 2009, 23, 75–84. [Google Scholar] [CrossRef]
- Davis, M. Modulation of Microglia Can Attenuate Neuropathic Pain Symptoms and Enhance Morphine Effectiveness. J. Pain Palliat. Care Pharmacother. 2009, 23. [Google Scholar]
- Mahmoud, S.; Gharagozloo, M.; Simard, C.; Gris, D. Astrocytes Maintain Glutamate Homeostasis in the CNS by Controlling the Balance between Glutamate Uptake and Release. Cells 2019, 8, 184. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Suárez, E.; Caldwell, A.L.M.; Allen, N.J. Role of Astrocyte-Synapse Interactions in CNS Disorders. J. Physiol. 2017, 595, 1903–1916. [Google Scholar] [CrossRef] [Green Version]
- Duran, J.; Gruart, A.; López-Ramos, J.C.; Delgado-García, J.M.; Guinovart, J.J. Glycogen in Astrocytes and Neurons: Physiological and Pathological Aspects. Adv. Neurobiol. 2019, 23, 311–329. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare Mannelli, L.; Corti, F.; Micheli, L.; Zanardelli, M.; Ghelardini, C. Delay of Morphine Tolerance by Palmitoylethanolamide. Biomed. Res. Int. 2015, 2015, 894732. [Google Scholar] [CrossRef] [Green Version]
- Song, P.; Zhao, Z.Q. The Involvement of Glial Cells in the Development of Morphine Tolerance. Neurosci. Res. 2001, 39, 281–286. [Google Scholar] [CrossRef]
- Mika, J.; Osikowicz, M.; Makuch, W.; Przewlocka, B. Minocycline and Pentoxifylline Attenuate Allodynia and Hyperalgesia and Potentiate the Effects of Morphine in Rat and Mouse Models of Neuropathic Pain. Eur. J. Pharmacol. 2007, 560, 142–149. [Google Scholar] [CrossRef]
- Cui, Y.; Liao, X.X.; Liu, W.; Guo, R.X.; Wu, Z.Z.; Zhao, C.M.; Chen, P.X.; Feng, J.Q. A Novel Role of Minocycline: Attenuating Morphine Antinociceptive Tolerance by Inhibition of P38 MAPK in the Activated Spinal Microglia. Brain Behav. Immun. 2008, 22, 114–123. [Google Scholar] [CrossRef]
- Raghavendra, V.; Tanga, F.Y.; DeLeo, J.A. Attenuation of Morphine Tolerance, Withdrawal-Induced Hyperalgesia, and Associated Spinal Inflammatory Immune Responses by Propentofylline in Rats. Neuropsychopharmacology 2003, 29, 327–334. [Google Scholar] [CrossRef] [Green Version]
- D’aloia, A.; Molteni, L.; Gullo, F.; Bresciani, E.; Artusa, V.; Rizzi, L.; Ceriani, M.; Meanti, R.; Lecchi, M.; Coco, S.; et al. Palmitoylethanolamide Modulation of Microglia Activation: Characterization of Mechanisms of Action and Implication for Its Neuroprotective Effects. Int. J. Mol. Sci. 2021, 22, 3054. [Google Scholar] [CrossRef] [PubMed]
- Re, G.; Barbero, R.; Miolo, A.; di Marzo, V. Palmitoylethanolamide, Endocannabinoids and Related Cannabimimetic Compounds in Protection against Tissue Inflammation and Pain: Potential Use in Companion Animals. Vet. J. 2007, 173, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Beggiato, S.; Tomasini, M.C.; Ferraro, L. Palmitoylethanolamide (PEA) as a Potential Therapeutic Agent in Alzheimer’s Disease. Front. Pharmacol. 2019, 10, 821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabrielsson, L.; Mattsson, S.; Fowler, C.J. Palmitoylethanolamide for the Treatment of Pain: Pharmacokinetics, Safety and Efficacy. Br. J. Clin. Pharmacol. 2016, 82, 932–942. [Google Scholar] [CrossRef] [Green Version]
- Artukoglu, B.B.; Beyer, C.; Zuloff-Shani, A.; Brener, E.; Bloch, M.H. Efficacy of Palmitoylethanolamide for Pain: A Meta-Analysis. Pain Physician 2017, 20, 353. [Google Scholar]
- Rankin, L.; Fowler, C.J. The Basal Pharmacology of Palmitoylethanolamide. Int. J. Mol. Sci. 2020, 21, 7942. [Google Scholar] [CrossRef]
- Gabrielsson, L.; Gouveia-Figueira, S.; Häggström, J.; Alhouayek, M.; Fowler, C.J. The Anti-Inflammatory Compound Palmitoylethanolamide Inhibits Prostaglandin and Hydroxyeicosatetraenoic Acid Production by a Macrophage Cell Line. Pharmacol. Res. Perspect. 2017, 5, e00300. [Google Scholar] [CrossRef]
- Di Cesare Mannelli, L.; D’Agostino, G.; Pacini, A.; Russo, R.; Zanardelli, M.; Ghelardini, C.; Calignano, A. Palmitoylethanolamide Is a Disease-Modifying Agent in Peripheral Neuropathy: Pain Relief and Neuroprotection Share a PPAR-Alpha-Mediated Mechanism. Mediat. Inflamm. 2013, 2013, 328797. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, J.; Luk, K.; Vang, D.; Soto, W.; Vincent, L.; Robiner, S.; Saavedra, R.; Li, Y.; Gupta, P.; Gupta, K. Morphine Stimulates Cancer Progression and Mast Cell Activation and Impairs Survival in Transgenic Mice with Breast Cancer. Br. J. Anaesth. 2014, 113 (Suppl. 1), i4–i13. [Google Scholar] [CrossRef] [Green Version]
- Grosman, N. Histamine Release from Isolated Rat Mast Cells: Effect of Morphine and Related Drugs and Their Interaction with Compound 48/80. Agents Actions 1981, 11, 196–203. [Google Scholar] [CrossRef]
- Vincent, L.; Vang, D.; Nguyen, J.; Gupta, M.; Luk, K.; Ericson, M.E.; Simone, D.A.; Gupta, K. Mast Cell Activation Contributes to Sickle Cell Pathobiology and Pain in Mice. Blood 2013, 122, 1853–1862. [Google Scholar] [CrossRef] [Green Version]
- Mazzari, S.; Canella, R.; Petrelli, L.; Marcolongo, G.; Leon, A. N-(2-Hydroxyethyl) Hexadecanamide Is Orally Active in Reducing Edema Formation and Inflammatory Hyperalgesia by down-Modulating Mast Cell Activation. Eur. J. Pharmacol. 1996, 300, 227–236. [Google Scholar] [CrossRef]
- Beggiato, S.; Borelli, A.C.; Ferraro, L.; Tanganelli, S.; Antonelli, T.; Tomasini, M.C. Palmitoylethanolamide Blunts Amyloid-Β42-Induced Astrocyte Activation and Improves Neuronal Survival in Primary Mouse Cortical Astrocyte-Neuron Co-Cultures. J. Alzheimer’s Dis. 2018, 61, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Blank, U.; Varin-Blank, N. La Lignée Mastocytaire RBL: Modèle Expérimental in Vitro et Application En Pharmacologie Clinique. Rev. Fr. Allergol. Immunol. Clin. 2004, 44, 51–56. [Google Scholar] [CrossRef]
- McCarthy, K.D.; de Vellis, J. Preparation of Separate Astroglial and Oligodendroglial Cell Cultures from Rat Cerebral Tissue. J. Cell Biol. 1980, 85, 890–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berridge, M.V.; Tan, A.S. Characterization of the Cellular Reduction of 3-(4,5-Dimethylthiazol-2-Yl)-2,5-Diphenyltetrazolium Bromide (MTT): Subcellular Localization, Substrate Dependence, and Involvement of Mitochondrial Electron Transport in MTT Reduction. Arch. Biochem. Biophys. 1993, 303, 474–482. [Google Scholar] [CrossRef]
- Jönsson, D.; Nilsson, B.O. The Antimicrobial Peptide LL-37 Is Anti-Inflammatory and Proapoptotic in Human Periodontal Ligament Cells. J. Periodontal Res. 2012, 47, 330–335. [Google Scholar] [CrossRef]
- Kawahara, T. Establishment and Characterization of Mouse Bone Marrow-Derived Mast Cell Hybridomas. Exp. Cell Res. 2012, 318, 2385–2396. [Google Scholar] [CrossRef] [Green Version]
- Micheli, L.; Lucarini, E.; Toti, A.; Ferrara, V.; Ciampi, C.; Parisio, C.; Bartolucci, G.; Mannelli, L.D.C.; Ghelardini, C. Effects of Ultramicronized N-Palmitoylethanolamine Supplementation on Tramadol and Oxycodone Analgesia and Tolerance Prevention. Pharmaceutics 2022, 14, 403. [Google Scholar] [CrossRef] [PubMed]
- Micheli, L.; Parisio, C.; Lucarini, E.; Vona, A.; Toti, A.; Pacini, A.; Mello, T.; Boccella, S.; Ricciardi, F.; Maione, S.; et al. VEGF-A/VEGFR-1 Signalling and Chemotherapy-Induced Neuropathic Pain: Therapeutic Potential of a Novel Anti-VEGFR-1 Monoclonal Antibody. J. Exp. Clin. Cancer Res. 2021, 40, 320. [Google Scholar] [CrossRef] [PubMed]
- Nistri, S.; Mannelli, L.D.C.; Ghelardini, C.; Zanardelli, M.; Bani, D.; Failli, P. Pretreatment with Relaxin Does Not Restore NO-Mediated Modulation of Calcium Signal in Coronary Endothelial Cells Isolated from Spontaneously Hypertensive Rats. Molecules 2015, 20, 9524–9535. [Google Scholar] [CrossRef] [Green Version]
- Passante, E.; Ehrhardt, C.; Sheridan, H.; Frankish, N. RBL-2H3 Cells Are an Imprecise Model for Mast Cell Mediator Release. Inflamm. Res. 2009, 58, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Passante, E.; Frankish, N. The RBL-2H3 Cell Line: Its Provenance and Suitability as a Model for the Mast Cell. Inflamm. Res. 2009, 58, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Qian, F.; Guo, G.; Li, Y.; Kulka, M. A Novel Eremophilane Lactone Inhibits FcεRI-Dependent Release of pro-Inflammatory Mediators: Structure-Dependent Bioactivity. Inflamm. Res. 2016, 65, 303–311. [Google Scholar] [CrossRef]
- Rujitharanawong, C.; Yoodee, S.; Sueksakit, K.; Peerapen, P.; Tuchinda, P.; Kulthanan, K.; Thongboonkerd, V. Systematic Comparisons of Various Markers for Mast Cell Activation in RBL-2H3 Cells. Cell Tissue Res. 2022, 390, 413–428. [Google Scholar] [CrossRef]
- Di Cesare Mannelli, L.; Micheli, L.; Lucarini, E.; Ghelardini, C. N-Palmitoylethanolamine Supplementation for Long-Lasting, Low-Dosed Morphine Antinociception. Front. Pharmacol. 2018, 9, 473. [Google Scholar] [CrossRef] [Green Version]
- Micheli, L.; Lucarini, E.; Nobili, S.; Bartolucci, G.; Pallecchi, A.; Toti, A.; Ferrara, V.; Ciampi, C.; Ghelardini, C.; Di Cesare Mannelli, L. Ultramicronized N-Palmitoylethanolamine Contributes to Morphine Efficacy against Neuropathic Pain: Implication of Mast Cells and Glia. Curr. Neuropharmacol. 2022, 21. [Google Scholar] [CrossRef]
- Petrosino, S.; Schiano Moriello, A.; Verde, R.; Allarà, M.; Imperatore, R.; Ligresti, A.; Mahmoud, A.M.; Peritore, A.F.; Iannotti, F.A.; di Marzo, V. Palmitoylethanolamide Counteracts Substance P-Induced Mast Cell Activation in Vitro by Stimulating Diacylglycerol Lipase Activity. J. Neuroinflamm. 2019, 16, 274. [Google Scholar] [CrossRef] [Green Version]
- Moon, T.C.; Dean Befus, A.; Kulka, M. Mast Cell Mediators: Their Differential Release and the Secretory Pathways Involved. Front. Immunol. 2014, 5, 569. [Google Scholar] [CrossRef] [Green Version]
- Payne, V.; Kam, P.C.A. Mast Cell Tryptase: A Review of Its Physiology and Clinical Significance. Anaesthesia 2004, 59, 695–703. [Google Scholar] [CrossRef]
- Mittal, A.; Sagi, V.; Gupta, M.; Gupta, K. Mast Cell Neural Interactions in Health and Disease. Front. Cell. Neurosci. 2019, 13, 110. [Google Scholar] [CrossRef] [Green Version]
- Sayed, B.A.; Christy, A.L.; Walker, M.E.; Brown, M.A. Meningeal Mast Cells Affect Early T Cell Central Nervous System Infiltration and Blood-Brain Barrier Integrity through TNF: A Role for Neutrophil Recruitment? J. Immunol. 2010, 184, 6891–6900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, G.; Singh, N.; Jaggi, A.S. Mast Cells in Neuropathic Pain: An Increasing Spectrum of Their Involvement in Pathophysiology. Rev. Neurosci. 2017, 28, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Koda, H.; Mizumura, K. Sensitization to Mechanical Stimulation by Inflammatory Mediators and by Mild Burn in Canine Visceral Nociceptors in Vitro. J. Neurophysiol. 2002, 87, 2043–2051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skaper, S.D.; Facci, L.; Giusti, P. Mast Cells, Glia and Neuroinflammation: Partners in Crime? Immunology 2014, 141, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Xanthos, D.N.; Gaderer, S.; Drdla, R.; Nuro, E.; Abramova, A.; Ellmeier, W.; Sandkühler, J. Central Nervous System Mast Cells in Peripheral Inflammatory Nociception. Mol. Pain 2011, 7, 1744–8069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, D.; Kainz, V.; Burstein, R.; Strassman, A.M. Mast Cell Degranulation Distinctly Activates Trigemino-Cervical and Lumbosacral Pain Pathways and Elicits Widespread Tactile Pain Hypersensitivity. Brain Behav. Immun. 2012, 26, 311–317. [Google Scholar] [CrossRef] [Green Version]
- Lian, Q.; Cheng, Y.; Zhong, C.; Wang, F. Inhibition of the IgE-Mediated Activation of RBL-2H3 Cells by TIPP, a Novel Thymic Immunosuppressive Pentapeptide. Int. J. Mol. Sci. 2015, 16, 2252. [Google Scholar] [CrossRef] [Green Version]
- Fu, H.; Cheng, H.; Cao, G.; Zhang, X.; Tu, J.; Sun, M.; Mou, X.; Shou, Q.; Ke, Y. The Inhibition of Mast Cell Activation of Radix Paeoniae Alba Extraction Identified by TCRP Based and Conventional Cell Function Assay Systems. PLoS ONE 2016, 11, e0155930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okahashi, N.; Nakata, M.; Hirose, Y.; Morisaki, H.; Kataoka, H.; Kuwata, H.; Kawabata, S. Streptococcal H2O2 Inhibits IgE-Triggered Degranulation of RBL-2H3 Mast Cell/Basophil Cell Line by Inducing Cell Death. PLoS ONE 2020, 15, e0231101. [Google Scholar] [CrossRef]
- Yaksh, T.L.; Allen, J.W.; Veesart, S.L.; Horais, K.A.; Malkmus, S.A.; Scadeng, M.; Steinauer, J.J.; Rossi, S.S. Role of Meningeal Mast Cells in Intrathecal Morphine-Evoked Granuloma Formation. Anesthesiology 2013, 118, 664–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dybendal, T.; Guttormsen, A.B.; Elsayed, S.; Askeland, B.; Harboe, T.; Florvaag, E. Screening for Mast Cell Tryptase and Serum IgE Antibodies in 18 Patients with Anaphylactic Shock during General Anaesthesia. Acta Anaesthesiol. Scand. 2003, 47, 1211–1218. [Google Scholar] [CrossRef]
- Sheen, C.H.; Schleimer, R.P.; Kulka, M. Codeine Induces Human Mast Cell Chemokine and Cytokine Production: Involvement of G-Protein Activation. Allergy 2007, 62, 532–538. [Google Scholar] [CrossRef] [Green Version]
- Fukuishi, N.; Murakami, S.; Ohno, A.; Yamanaka, N.; Matsui, N.; Fukutsuji, K.; Yamada, S.; Itoh, K.; Akagi, M. Does β-Hexosaminidase Function Only as a Degranulation Indicator in Mast Cells? The Primary Role of β-Hexosaminidase in Mast Cell Granules. J. Immunol. 2014, 193, 1886–1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alanazi, S.; Grujic, M.; Lampinen, M.; Rollman, O.; Sommerhoff, C.P.; Pejler, G.; Melo, F.R. Mast Cell β-Tryptase Is Enzymatically Stabilized by DNA. Int. J. Mol. Sci. 2020, 21, 5065. [Google Scholar] [CrossRef] [PubMed]
- Atiakshin, D.; Buchwalow, I.; Samoilova, V.; Tiemann, M. Tryptase as a Polyfunctional Component of Mast Cells. Histochem. Cell Biol. 2018, 149, 461–477. [Google Scholar] [CrossRef]
- Shim, J.K.; Kennedy, R.H.; Weatherly, L.M.; Abovian, A.V.; Hashmi, H.N.; Rajaei, A.; Gosse, J.A. Searching for Tryptase in the RBL-2H3 Mast Cell Model: Preparation for Comparative Mast Cell Toxicology Studies with Zebrafish. J. Appl. Toxicol. 2019, 39, 473–484. [Google Scholar] [CrossRef]
- Li, Y.; Sun, X.; Juan, Z.; Guan, X.; Wang, M.; Meng, Y.; Ma, R. Propofol Pretreatment Alleviates Mast Cell Degranulation by Inhibiting SOC to Protect the Myocardium from Ischemia–Reperfusion Injury. Biomed. Pharmacother. 2022, 150, 113014. [Google Scholar] [CrossRef]
- Yaksh, T.L.; Eddinger, K.A.; Kokubu, S.; Wang, Z.; Dinardo, A.; Ramachandran, R.; Zhu, Y.; He, Y.; Weren, F.; Quang, D.; et al. Mast Cell Degranulation and Fibroblast Activation in the Morphine-Induced Spinal Mass: Role of Mas-Related G Protein-Coupled Receptor Signaling. Anesthesiology 2019, 131, 132–147. [Google Scholar] [CrossRef] [PubMed]
- Thangam, E.B.; Jemima, E.A.; Singh, H.; Baig, M.S.; Khan, M.; Mathias, C.B.; Church, M.K.; Saluja, R. The Role of Histamine and Histamine Receptors in Mast Cell-Mediated Allergy and Inflammation: The Hunt for New Therapeutic Targets. Front. Immunol. 2018, 9, 1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlos, D.; Sá-Nunes, A.; de Paula, L.; Matias-Peres, C.; Jamur, M.C.; Oliver, C.; Serra, M.F.; Martins, M.A.; Faccioli, L.H. Histamine Modulates Mast Cell Degranulation through an Indirect Mechanism in a Model IgE-Mediated Reaction. Eur. J. Immunol. 2006, 36, 1494–1503. [Google Scholar] [CrossRef]
- Gu, Y.; Yang, D.K.; Spinas, E.; Kritas, S.K.; Saggini, A.; Caraffa, A.; Antinolfi, P.; Saggini, R.; Conti, P. Role of TNF in Mast Cell Neuroinflammation and Pain. J. Biol. Regul. Homeost. Agents 2015, 29, 787–791. [Google Scholar]
- Castellani, M.L.; Bhattacharya, K.; Tagen, M.; Kempuraj, D.; Perrella, A.; de Lutiis, M.; Boucher, W.; Conti, P.; Theoharides, T.C.; Cerulli, G.; et al. Anti-Chemokine Therapy for Inflammatory Diseases. Int. J. Immunopathol. Pharmacol. 2016, 20, 447–453. [Google Scholar] [CrossRef]
- Bischoff, S.C.; Krieger, M.; Brunner, T.; Rot, A.; Tscharner, V.V.; Baggiolini, M.; Dahinden, C.A. RANTES and Related Chemokines Activate Human Basophil Granulocytes through Different G Protein-coupled Receptors. Eur. J. Immunol. 1993, 23, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Facci, L.; Dal Toso, R.; Romanello, S.; Buriani, A.; Skaper, S.D.; Leon, A. Mast Cells Express a Peripheral Cannabinoid Receptor with Differential Sensitivity to Anandamide and Palmitoylethanolamide. Proc. Natl. Acad. Sci. USA 1995, 92, 3376–3380. [Google Scholar] [CrossRef] [Green Version]
- Cerrato, S.; Brazis, P.; della Valle, M.F.; Miolo, A.; Puigdemont, A. Effects of Palmitoylethanolamide on Immunologically Induced Histamine, PGD2 and TNFalpha Release from Canine Skin Mast Cells. Vet. Immunol. Immunopathol. 2010, 133, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Abramo, F.; Lazzarini, G.; Pirone, A.; Lenzi, C.; Albertini, S.; della Valle, M.F.; Schievano, C.; Vannozzi, I.; Miragliotta, V. Ultramicronized Palmitoylethanolamide Counteracts the Effects of Compound 48/80 in a Canine Skin Organ Culture Model. Vet. Dermatol. 2017, 28, 104–456. [Google Scholar] [CrossRef] [Green Version]
- Bettoni, I.; Comelli, F.; Colombo, A.; Bonfanti, P.; Costa, B. Non-Neuronal Cell Modulation Relieves Neuropathic Pain: Efficacy of the Endogenous Lipid Palmitoylethanolamide. CNS Neurol. Disord. Drug Targets 2013, 12, 34–44. [Google Scholar] [CrossRef]
- Scarampella, F.; Abramo, F.; Noli, C. Clinical and Histological Evaluation of an Analogue of Palmitoylethanolamide, PLR 120 (Comicronized Palmidrol INN) in Cats with Eosinophilic Granuloma and Eosinophilic Plaque: A Pilot Study. Vet. Dermatol. 2001, 12, 29–39. [Google Scholar] [CrossRef]
- Aloe, L.; Leon, A.; Levi-Montalcini, R. A Proposed Autacoid Mechanism Controlling Mastocyte Behaviour. Agents Actions 1993, 39, C145–C147. [Google Scholar] [CrossRef] [PubMed]
- Watkins, L.R.; Hutchinson, M.R.; Johnston, I.N.; Maier, S.F. Glia: Novel Counter-Regulators of Opioid Analgesia. Trends Neurosci. 2005, 28, 661–669. [Google Scholar] [CrossRef]
- Liu, D.-Q.; Zhou, Y.-Q.; Gao, F. Targeting Cytokines for Morphine Tolerance: A Narrative Review. Curr. Neuropharmacol. 2019, 17, 366–376. [Google Scholar] [CrossRef]
- Dozio, V.; Daali, Y.; Desmeules, J.; Sanchez, J.C. Deep Proteomics and Phosphoproteomics Reveal Novel Biological Pathways Perturbed by Morphine, Morphine-3-Glucuronide and Morphine-6-Glucuronide in Human Astrocytes. J. Neurosci. Res. 2022, 100, 220–236. [Google Scholar] [CrossRef]
- Ma, R.; Kutchy, N.A.; Hu, G. Astrocyte-Derived Extracellular Vesicle-Mediated Activation of Primary Ciliary Signaling Contributes to the Development of Morphine Tolerance. Biol. Psychiatry 2021, 90, 575–585. [Google Scholar] [CrossRef]
- Congiu, M.; Micheli, L.; Santoni, M.; Sagheddu, C.; Muntoni, A.L.; Makriyannis, A.; Malamas, M.S.; Ghelardini, C.; Di Cesare Mannelli, L.; Pistis, M. N-Acylethanolamine Acid Amidase Inhibition Potentiates Morphine Analgesia and Delays the Development of Tolerance. Neurotherapeutics 2021, 18, 2722–2736. [Google Scholar] [CrossRef] [PubMed]
- Ruzicka, B.B.; Fox, C.A.; Thompson, R.C.; Meng, F.; Watson, S.J.; Akil, H. Primary Astroglial Cultures Derived from Several Rat Brain Regions Differentially Express μ, δ and κ Opioid Receptor MRNA. Mol. Brain Res. 1995, 34, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Nam, M.H.; Han, K.S.; Lee, J.; Bae, J.Y.; An, H.; Park, S.; Oh, S.J.; Kim, E.; Hwang, E.; Bae, Y.C.; et al. Expression of Μ-Opioid Receptor in CA1 Hippocampal Astrocytes. Exp. Neurobiol. 2018, 27, 120. [Google Scholar] [CrossRef]
- Sil, S.; Periyasamy, P.; Guo, M.L.; Callen, S.; Buch, S. Morphine-Mediated Brain Region-Specific Astrocytosis Involves the ER Stress-Autophagy Axis. Mol. Neurobiol. 2018, 55, 6713. [Google Scholar] [CrossRef]
- Esposito, E.; Paterniti, I.; Mazzon, E.; Genovese, T.; di Paola, R.; Galuppo, M.; Cuzzocrea, S. Effects of Palmitoylethanolamide on Release of Mast Cell Peptidases and Neurotrophic Factors after Spinal Cord Injury. Brain Behav. Immun. 2011, 25, 1099–1112. [Google Scholar] [CrossRef]
- Kempuraj, D.; Thangavel, R.; Selvakumar, G.P.; Zaheer, S.; Ahmed, M.E.; Raikwar, S.P.; Zahoor, H.; Saeed, D.; Natteru, P.A.; Iyer, S.; et al. Brain and Peripheral Atypical Inflammatory Mediators Potentiate Neuroinflammation and Neurodegeneration. Front. Cell. Neurosci. 2017, 11, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shavit, Y.; Wolf, G.; Goshen, I.; Livshits, D.; Yirmiya, R. Interleukin-1 Antagonizes Morphine Analgesia and Underlies Morphine Tolerance. Pain 2005, 115, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.H.; Tsai, R.Y.; Shih, M.S.; Lin, S.L.; Tai, Y.H.; Chien, C.C.; Wong, C.S. Etanercept Restores the Antinociceptive Effect of Morphine and Suppresses Spinal Neuroinflammation in Morphine-Tolerant Rats. Anesth. Analg. 2011, 112, 454–459. [Google Scholar] [CrossRef]
- Raghavendra, V.; Rutkowski, M.D.; Deleo, J.A. The Role of Spinal Neuroimmune Activation in Morphine Tolerance/Hyperalgesia in Neuropathic and Sham-Operated Rats. J. Neurosci. 2002, 22, 9980–9989. [Google Scholar] [CrossRef] [Green Version]
- Niu, Z.J.; Ma, J.G.; Chu, H.C.; Zhao, Y.; Feng, W.; Cheng, Y.W. Melanocortin 4 Receptor Antagonists Attenuates Morphine Antinociceptive Tolerance, Astroglial Activation and Cytokines Expression in the Spinal Cord of Rat. Neurosci. Lett. 2012, 529, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowski, K.; Popiolek-Barczyk, K.; Piotrowska, A.; Rojewska, E.; Ciapała, K.; Makuch, W.; Mika, J. Chemokines CCL2 and CCL7, but Not CCL12, Play a Significant Role in the Development of Pain-Related Behavior and Opioid-Induced Analgesia. Cytokine 2019, 119, 202–213. [Google Scholar] [CrossRef]
- Zhang, Y.; He, J.; Zhao, J.; Xu, M.; Lou, D.; Tso, P.; Li, Z.; Li, X. Effect of ApoA4 on SERPINA3 Mediated by Nuclear Receptors NR4A1 and NR1D1 in Hepatocytes. Biochem. Biophys. Res. Commun. 2017, 487, 327–332. [Google Scholar] [CrossRef] [Green Version]
- Grek, C.L.; Townsend, D.M.; Uys, J.D.; Manevich, Y.; Coker, W.J.; Pazoles, C.J.; Tew, K.D. S-Glutathionylated Serine Proteinase Inhibitors as Plasma Biomarkers in Assessing Response to Redox-Modulating Drugs. Cancer Res. 2012, 72, 2383–2393. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Masaki, T.; Mezaki, Y.; Yokoyama, H.; Nakamura, M.; Maehashi, H.; Fujimi, T.J.; Gouraud, S.S.; Nagatsuma, K.; Nakagomi, M.; et al. Alpha-1 Antichymotrypsin Is Involved in Astrocyte Injury in Concert with Arginine-Vasopressin during the Development of Acute Hepatic Encephalopathy. PLoS ONE 2017, 12, e0189346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanemaru, K.; Meckelein, B.; Marshall, D.C.L.; Sipe, J.D.; Abraham, C.R. Synthesis and Secretion of Active Alpha 1-Antichymotrypsin by Murine Primary Astrocytes. Neurobiol. Aging 1996, 17, 767–771. [Google Scholar] [CrossRef] [PubMed]
- Kiss, D.L.; Xu, W.; Gopalan, S.; Buzanowska, K.; Wilczynska, K.M.; Rydel, R.E.; Kordula, T. Duration of A1-Antichymotrypsin Gene Activation by IL-1 Is Determined by Efficiency of IkBα Resynthesis in Primary Human Astrocytes. J. Neurochem. 2005, 92, 730. [Google Scholar] [CrossRef] [Green Version]
- Vicuña, L.; Strochlic, D.E.; Latremoliere, A.; Bali, K.K.; Simonetti, M.; Husainie, D.; Prokosch, S.; Riva, P.; Griffin, R.S.; Njoo, C.; et al. The Serine Protease Inhibitor SerpinA3N Attenuates Neuropathic Pain by Inhibiting T Cell–Derived Leukocyte Elastase. Nat. Med. 2015, 21, 518–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bannon, M.J.; Johnson, M.M.; Michelhaugh, S.K.; Hartley, Z.J.; Halter, S.D.; David, J.A.; Kapatos, G.; Schmidt, C.J. A Molecular Profile of Cocaine Abuse Includes the Differential Expression of Genes That Regulate Transcription, Chromatin, and Dopamine Cell Phenotype. Neuropsychopharmacology 2014, 39, 2191–2199. [Google Scholar] [CrossRef] [PubMed]
- Zamanian, J.L.; Xu, L.; Foo, L.C.; Nouri, N.; Zhou, L.; Giffard, R.G.; Barres, B.A. Genomic Analysis of Reactive Astrogliosis. J. Neurosci. 2012, 32, 6391–6410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Kou, D.; Yu, Z.; Yang, S.; Jiang, C.; Xiong, D.; Xiao, L.; Deng, Q.; Xie, H.; Hao, Y. A Transcriptomic Analysis of Neuropathic Pain in Rat Dorsal Root Ganglia Following Peripheral Nerve Injury. Neuromol. Med. 2020, 22, 250–263. [Google Scholar] [CrossRef]
- Maragakis, N.J.; Dykes-Hoberg, M.; Rothstein, J.D. Altered Expression of the Glutamate Transporter EAAT2b in Neurological Disease. Ann. Neurol. 2004, 55, 469–477. [Google Scholar] [CrossRef]
- Middeldorp, J.; Hol, E.M. GFAP in Health and Disease. Prog. Neurobiol. 2011, 93, 421–443. [Google Scholar] [CrossRef]
- Pekny, M.; Wilhelmsson, U.; Pekna, M. The Dual Role of Astrocyte Activation and Reactive Gliosis. Neurosci. Lett. 2014, 565, 30–38. [Google Scholar] [CrossRef]
- Linnerbauer, M.; Rothhammer, V. Protective Functions of Reactive Astrocytes Following Central Nervous System Insult. Front. Immunol. 2020, 11, 573256. [Google Scholar] [CrossRef]
- Beitner-Johnson, D.; Guitart, X.; Nestler, E.J. Glial Fibrillary Acidic Protein and the Mesolimbic Dopamine System: Regulation by Chronic Morphine and Lewis-Fischer Strain Differences in the Rat Ventral Tegmental Area. J. Neurochem. 1993, 61, 1766–1773. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency. Guideline Bioanalytical Method Validation. 2015. Available online: https://www.ema.europa.eu/en/documents/scientific-guideline/guideline-bioanalytical-method-validation_en.pdf (accessed on 12 January 2023).
- ICH Q2(R2) Validation of Analytical Procedures. Available online: https://www.ema.europa.eu/en/ich-q2r2-validation-analytical-procedures (accessed on 12 January 2022).
- Matuszewski, B.K.; Constanzer, M.L.; Chavez-Eng, C.M. Strategies for the Assessment of Matrix Effect in Quantitative Bioanalytical Methods Based on HPLC−MS/MS. Anal. Chem. 2003, 75, 3019–3030. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toti, A.; Micheli, L.; Lucarini, E.; Ferrara, V.; Ciampi, C.; Margiotta, F.; Failli, P.; Gomiero, C.; Pallecchi, M.; Bartolucci, G.; et al. Ultramicronized N-Palmitoylethanolamine Regulates Mast Cell-Astrocyte Crosstalk: A New Potential Mechanism Underlying the Inhibition of Morphine Tolerance. Biomolecules 2023, 13, 233. https://doi.org/10.3390/biom13020233
Toti A, Micheli L, Lucarini E, Ferrara V, Ciampi C, Margiotta F, Failli P, Gomiero C, Pallecchi M, Bartolucci G, et al. Ultramicronized N-Palmitoylethanolamine Regulates Mast Cell-Astrocyte Crosstalk: A New Potential Mechanism Underlying the Inhibition of Morphine Tolerance. Biomolecules. 2023; 13(2):233. https://doi.org/10.3390/biom13020233
Chicago/Turabian StyleToti, Alessandra, Laura Micheli, Elena Lucarini, Valentina Ferrara, Clara Ciampi, Francesco Margiotta, Paola Failli, Chiara Gomiero, Marco Pallecchi, Gianluca Bartolucci, and et al. 2023. "Ultramicronized N-Palmitoylethanolamine Regulates Mast Cell-Astrocyte Crosstalk: A New Potential Mechanism Underlying the Inhibition of Morphine Tolerance" Biomolecules 13, no. 2: 233. https://doi.org/10.3390/biom13020233