
Palmitoylethanolamide Mitigates Paclitaxel Toxicity in Primary Dorsal Root Ganglion Neurons

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Materials
2.3. Isolation and Preparation of DRG Neurons
2.4. Seeding and Growth of DRG Neurons
2.5. Cell Viability (MTT Assay)
2.6. Immunofluorescence Staining and Microscopy
2.7. Image Analysis and Determination of Neurite Lengths and Soma Sizes
2.8. Statistical Analysis
3. Results
3.1. Characterization of DRG Neuronal Cells
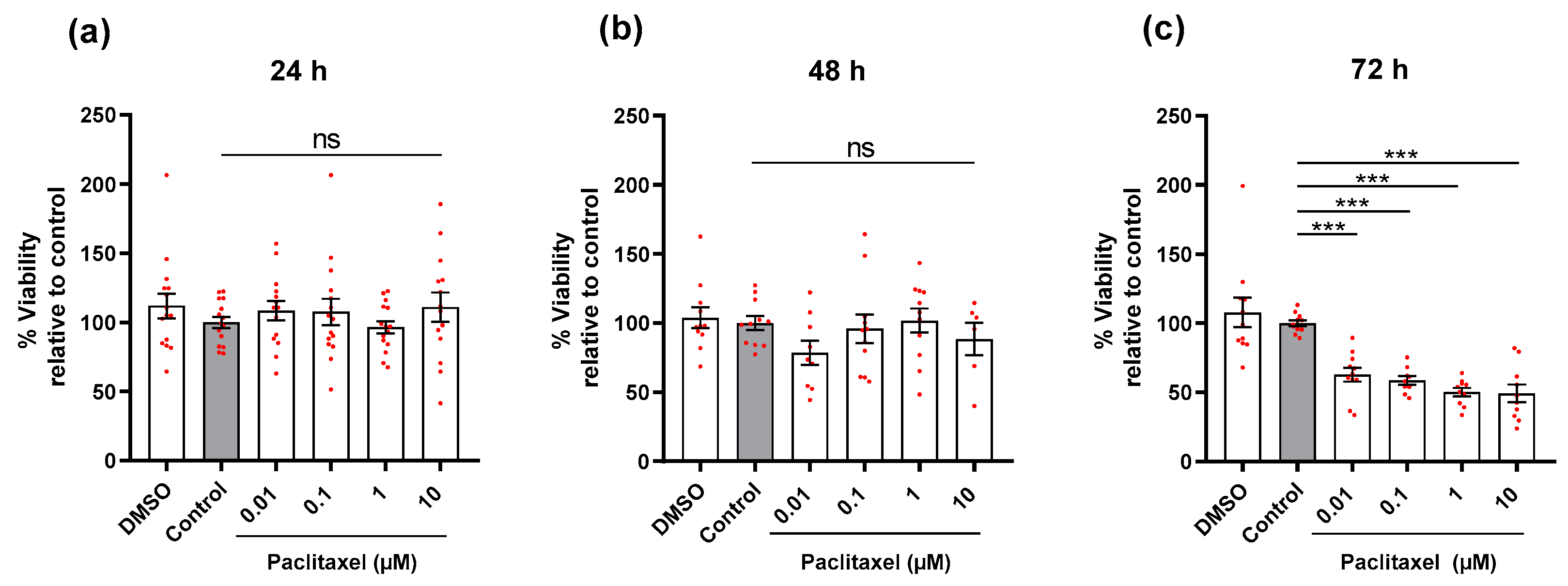
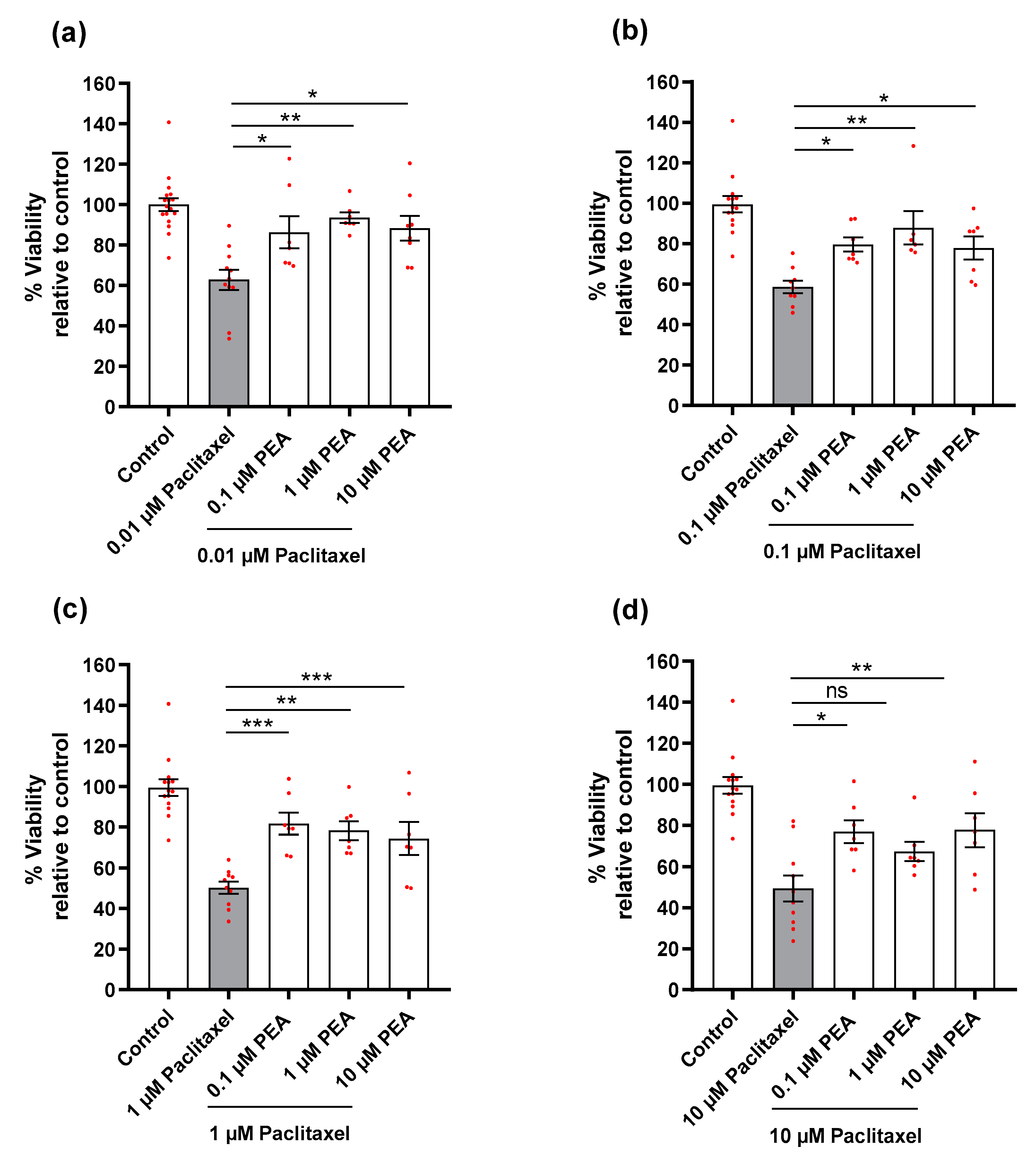
3.2. Effects of Paclitaxel or/and PEA on Cell Viability of DRG Neurons
3.3. Effects of Paclitaxel or/and PEA on Morphology of DRG Neurons
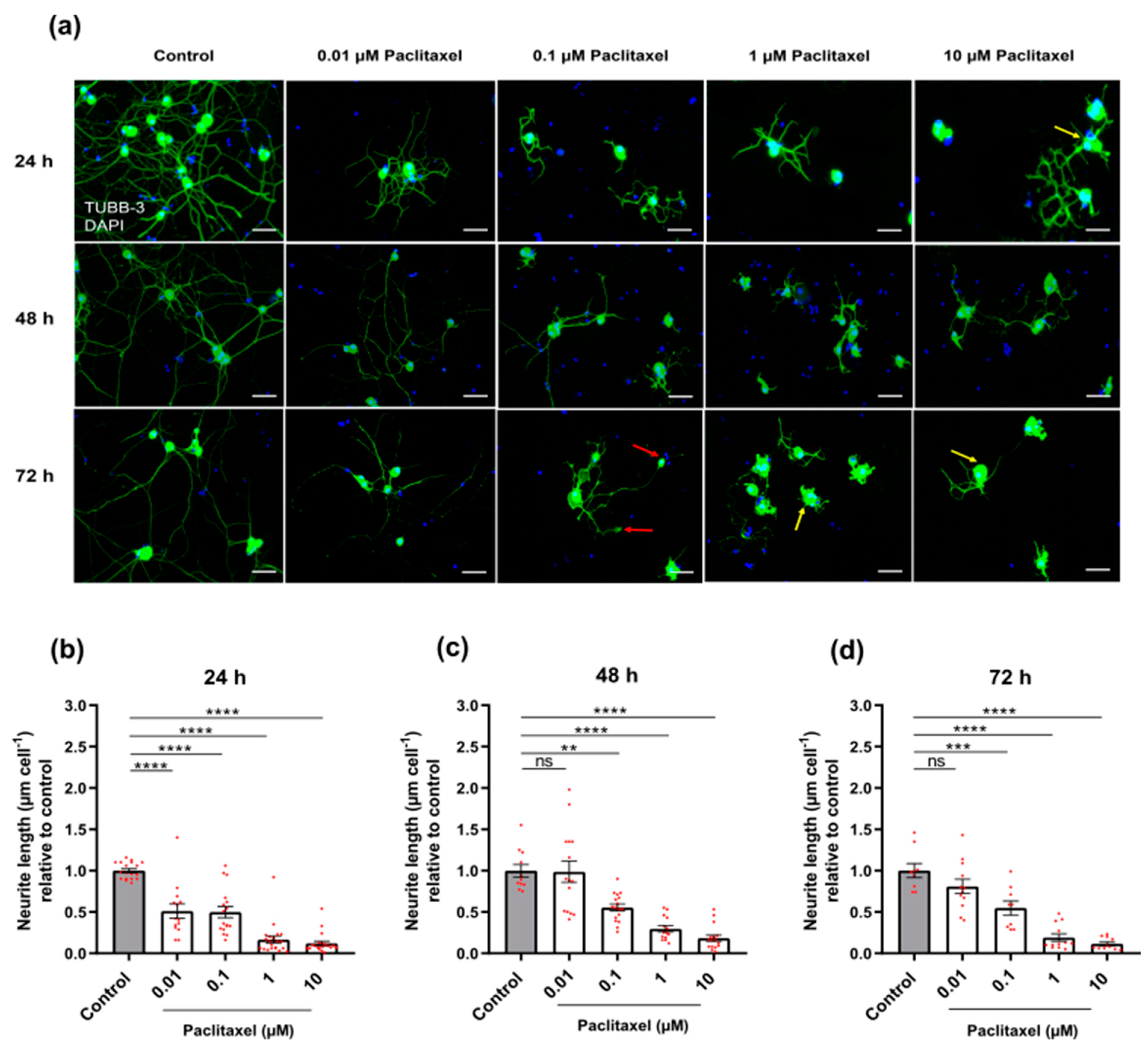
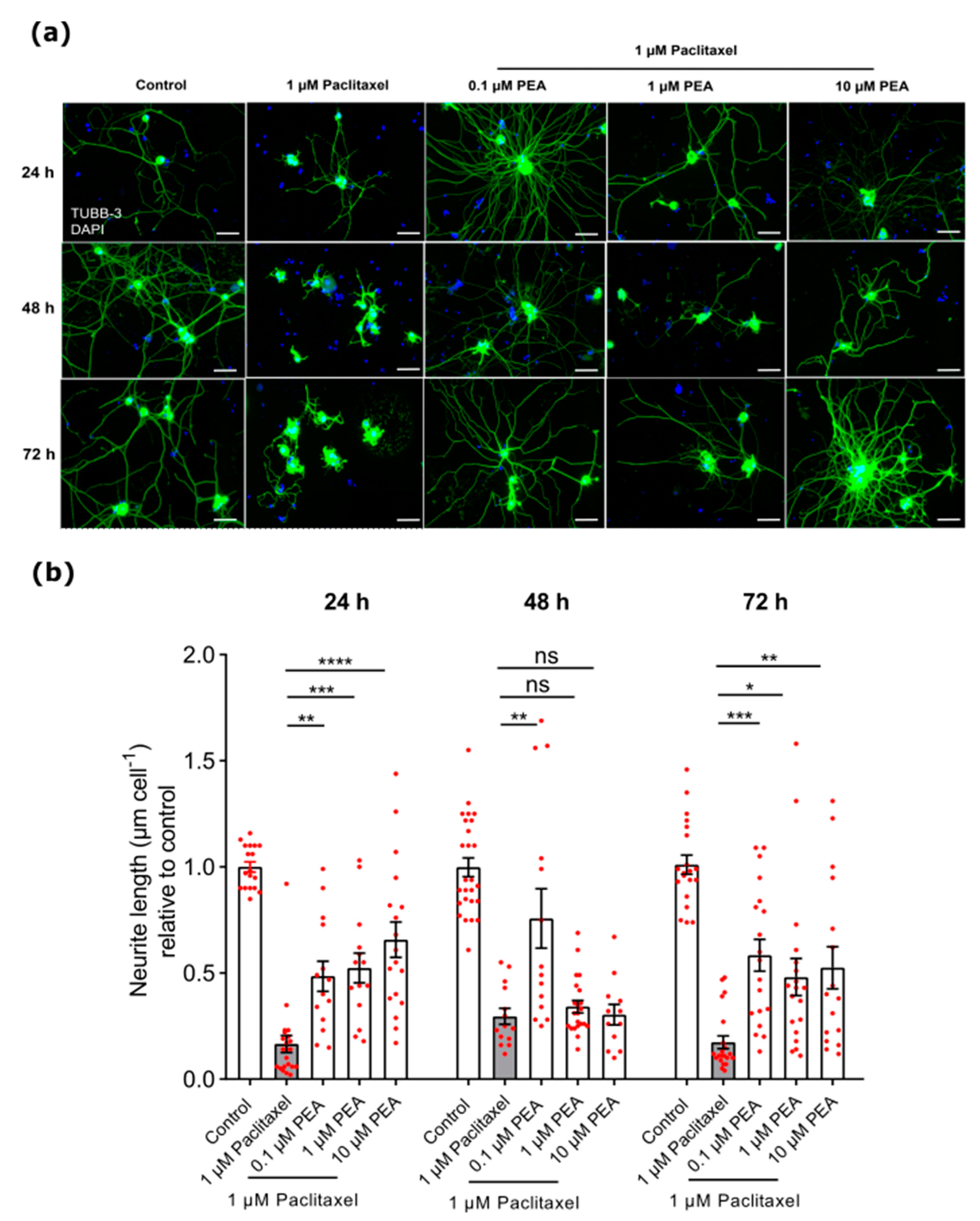
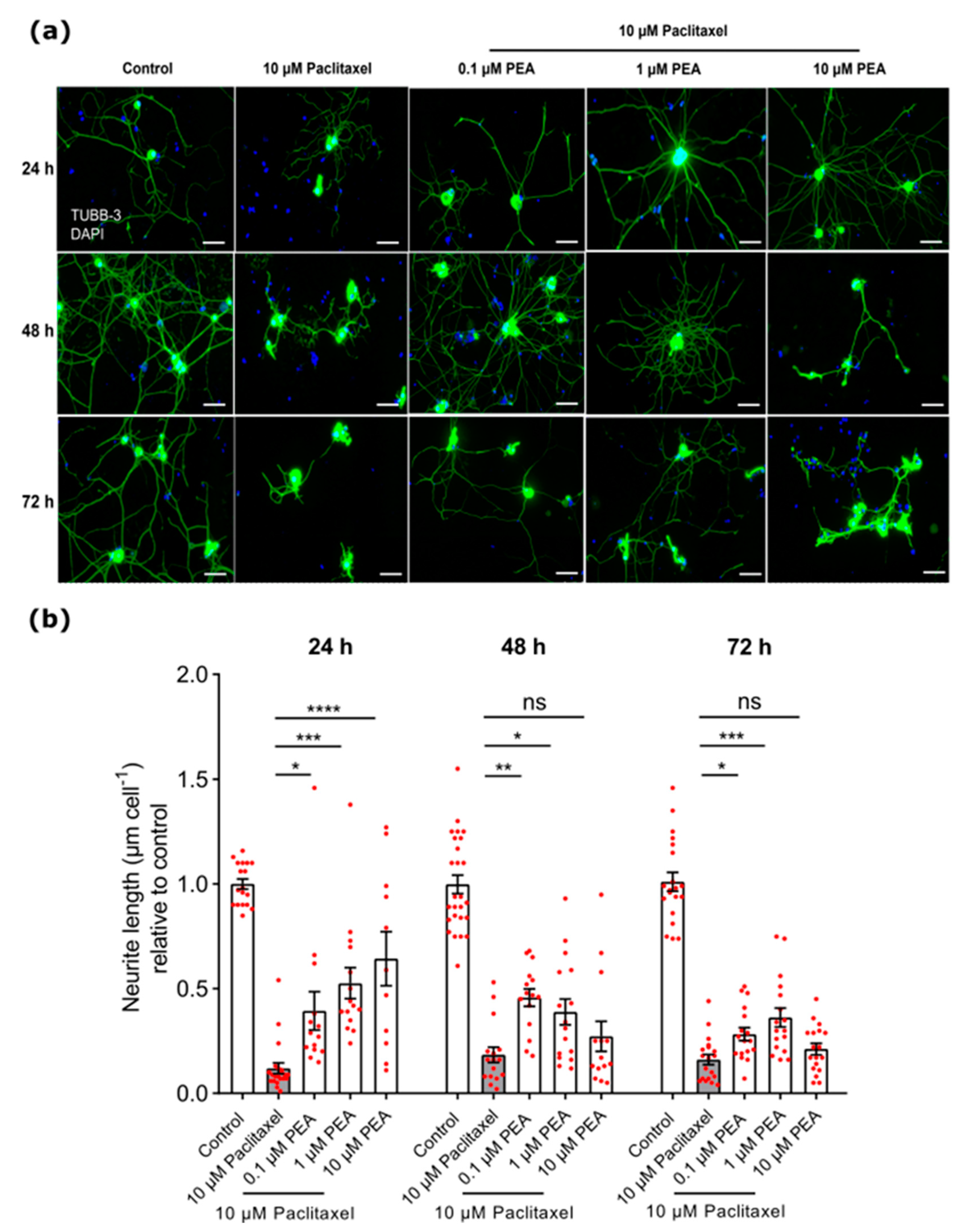
3.3.1. Neurite Length
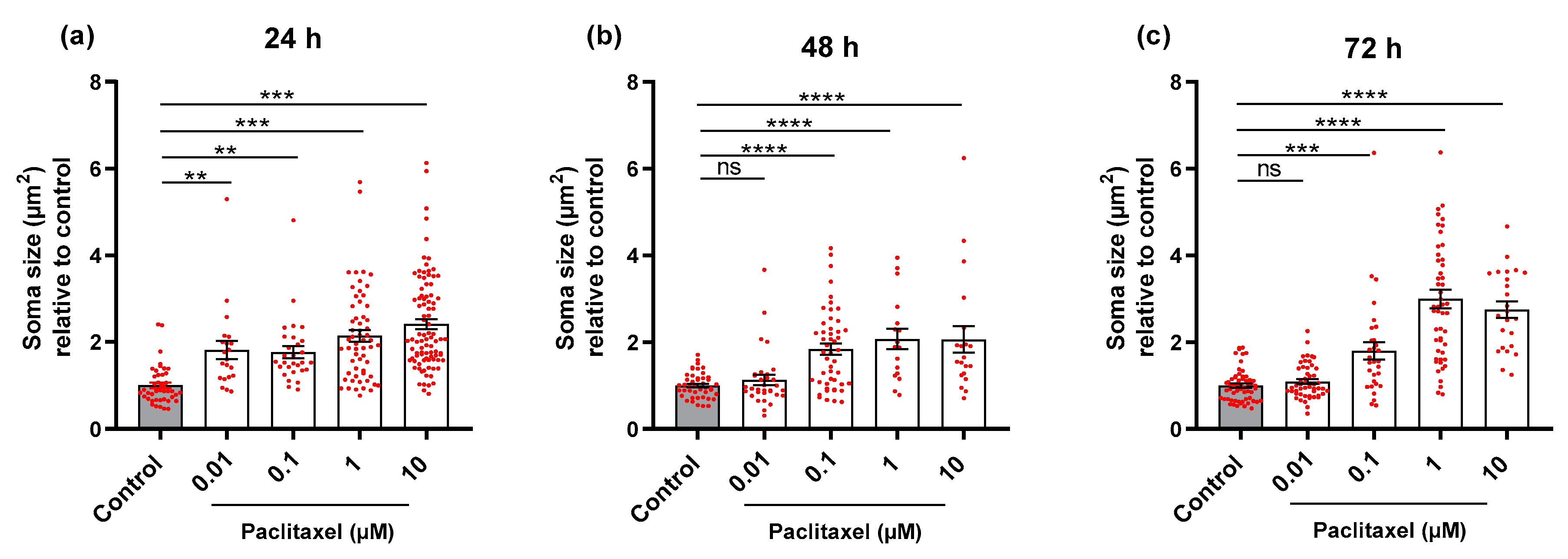
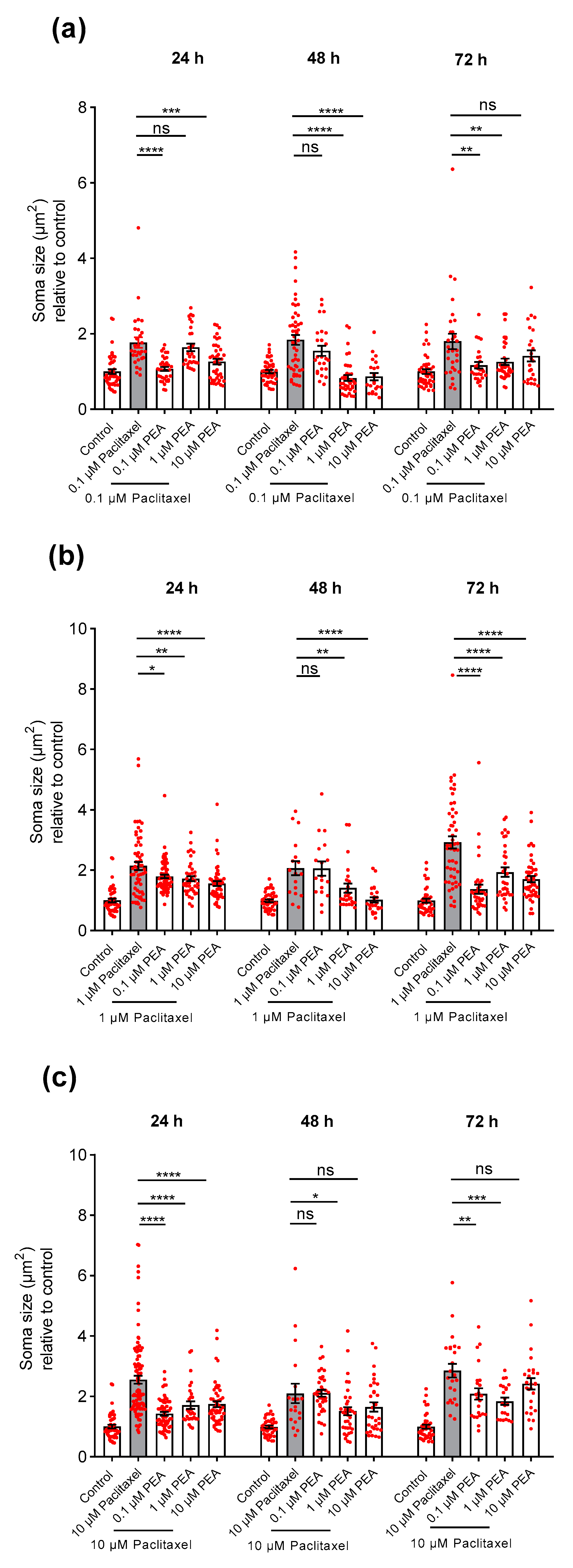
3.3.2. Soma Size
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grisold, W.; Cavaletti, G.; Windebank, A.J. Peripheral Neuropathies from Chemotherapeutics and Targeted Agents: Diagnosis, Treatment, and Prevention. Neuro. Oncol. 2012, 14 (Suppl. 4), iv45–iv54. [Google Scholar] [CrossRef] [Green Version]
- Masocha, W. Targeting the Endocannabinoid System for Prevention or Treatment of Chemotherapy-Induced Neuropathic Pain: Studies in Animal Models. Pain Res. Manag. 2018, 2018, 5234943. [Google Scholar] [CrossRef]
- Rowinsky, E.K.; Donehower, R.C. Paclitaxel (Taxol). N. Engl. J. Med. 1995, 332, 1004–1014. [Google Scholar] [CrossRef]
- Tankanow, R.M. Docetaxel: A Taxoid for the Treatment of Metastatic Breast Cancer. Am. J. Health Pharm. 1998, 55, 1777–1791. [Google Scholar] [CrossRef]
- Vyas, D.M.; Kadow, J.F. 6 Paclitaxel: A Unique Tubulin Interacting Anticancer Agent. Prog. Med. Chem. 1995, 32, 289–337. [Google Scholar] [CrossRef]
- Schiff, P.B.; Fant, J.; Horwitz, S.B. Promotion of Microtubule Assembly in Vitro by Taxol. Nature 1979, 277, 665–667. [Google Scholar] [CrossRef]
- Rowinsky, E.K.; Cazenave, L.A.; Donehower, R.C. Taxol: A Novel Investigational Antimicrotubule Agent. JNCI J. Natl. Cancer Inst. 1990, 82, 1247–1259. [Google Scholar] [CrossRef]
- Seretny, M.; Currie, G.L.; Sena, E.S.; Ramnarine, S.; Grant, R.; MacLeod, M.R.; Colvin, L.A.; Fallon, M. Incidence, Prevalence, and Predictors of Chemotherapy-Induced Peripheral Neuropathy: A Systematic Review and Meta-Analysis. Pain 2014, 155, 2461–2470. [Google Scholar] [CrossRef] [Green Version]
- Farquhar-Smith, P. Chemotherapy-Induced Neuropathic Pain. Curr. Opin. Support. Palliat. Care 2011, 5, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Dougherty, P.M.; Cata, J.P.; Cordella, J.V.; Burton, A.; Weng, H.-R. Taxol-Induced Sensory Disturbance Is Characterized by Preferential Impairment of Myelinated Fiber Function in Cancer Patients. Pain 2004, 109, 132–142. [Google Scholar] [CrossRef]
- Sharma, S.; Venkitaraman, R.; Vas, P.R.J.; Rayman, G. Assessment of Chemotherapy-Induced Peripheral Neuropathy Using the LDI FLARE Technique: A Novel Technique to Detect Neural Small Fiber Dysfunction. Brain Behav. 2015, 5, e00354. [Google Scholar] [CrossRef]
- Windebank, A.J.; Smith, A.G.; Russell, J.W. The Effect of Nerve Growth Factor, Ciliary Neurotrophic Factor, and ACTH Analogs on Cisplatin Neurotoxicity in Vitro. Neurology 1994, 44 Pt 1, 488–494. [Google Scholar] [CrossRef]
- Gill, J.S.; Windebank, A.J. Cisplatin-Induced Apoptosis in Rat Dorsal Root Ganglion Neurons Is Associated with Attempted Entry into the Cell Cycle. J. Clin. Investig. 1998, 101, 2842–2850. [Google Scholar] [CrossRef] [Green Version]
- Fischer, S.J.; McDonald, E.S.; Gross, L.; Windebank, A.J. Alterations in Cell Cycle Regulation Underlie Cisplatin Induced Apoptosis of Dorsal Root Ganglion Neurons in Vivo. Neurobiol. Dis. 2001, 8, 1027–1035. [Google Scholar] [CrossRef] [Green Version]
- Scuteri, A.; Nicolini, G.; Miloso, M.; Bossi, M.; Cavaletti, G.; Windebank, A.J.; Tredici, G. Paclitaxel Toxicity in Post-Mitotic Dorsal Root Ganglion (DRG) Cells. Anticancer Res. 2006, 26, 1065–1070. [Google Scholar]
- Guo, L.; Hamre, J.; Eldridge, S.; Behrsing, H.P.; Cutuli, F.M.; Mussio, J.; Davis, M. Multiparametric Image Analysis of Rat Dorsal Root Ganglion Cultures to Evaluate Peripheral Neuropathy-Inducing Chemotherapeutics. Toxicol. Sci. 2017, 156, kfw254. [Google Scholar] [CrossRef] [Green Version]
- Park, S.H.; Eber, M.R.; Fonseca, M.M.; Patel, C.M.; Cunnane, K.A.; Ding, H.; Hsu, F.-C.; Peters, C.M.; Ko, M.-C.; Strowd, R.E.; et al. Usefulness of the Measurement of Neurite Outgrowth of Primary Sensory Neurons to Study Cancer-Related Painful Complications. Biochem. Pharmacol. 2021, 188, 114520. [Google Scholar] [CrossRef]
- Akin, E.J.; Alsaloum, M.; Higerd, G.P.; Liu, S.; Zhao, P.; Dib-Hajj, F.B.; Waxman, S.G.; Dib-Hajj, S.D. Paclitaxel Increases Axonal Localization and Vesicular Trafficking of Nav1.7. Brain 2021, 144, 1727–1737. [Google Scholar] [CrossRef]
- Shin, G.J.; Pero, M.E.; Hammond, L.A.; Burgos, A.; Kumar, A.; Galindo, S.E.; Lucas, T.; Bartolini, F.; Grueber, W.B. Integrins Protect Sensory Neurons in Models of Paclitaxel-Induced Peripheral Sensory Neuropathy. Proc. Natl. Acad. Sci. USA 2021, 118, e2006050118. [Google Scholar] [CrossRef]
- Rao, S.S.C.; Seaton, K.; Miller, M.; Brown, K.; Nygaard, I.; Stumbo, P.; Zimmerman, B.; Schulze, K. Randomized Controlled Trial of Biofeedback, Sham Feedback, and Standard Therapy for Dyssynergic Defecation. Clin. Gastroenterol. Hepatol. 2007, 5, 331–338. [Google Scholar] [CrossRef]
- Kautio, A.-L.; Haanpää, M.; Leminen, A.; Kalso, E.; Kautiainen, H.; Saarto, T. Amitriptyline in the Prevention of Chemotherapy-Induced Neuropathic Symptoms. Anticancer Res. 2009, 29, 2601–2606. [Google Scholar]
- Hershman, D.L.; Lacchetti, C.; Loprinzi, C.L. Prevention and Management of Chemotherapy-Induced Peripheral Neuropathy in Survivors of Adult Cancers: American Society of Clinical Oncology Clinical Practice Guideline Summary. J. Oncol. Pract. 2014, 10, e421–e424. [Google Scholar] [CrossRef]
- Kim, J.H.; Dougherty, P.M.; Abdi, S. Basic Science and Clinical Management of Painful and Non-Painful Chemotherapy-Related Neuropathy. Gynecol. Oncol. 2015, 136, 453–459. [Google Scholar] [CrossRef] [Green Version]
- Aizpurua-Olaizola, O.; Elezgarai, I.; Rico-Barrio, I.; Zarandona, I.; Etxebarria, N.; Usobiaga, A. Targeting the Endocannabinoid System: Future Therapeutic Strategies. Drug Discov. Today 2017, 22, 105–110. [Google Scholar] [CrossRef]
- Pacher, P.; Kunos, G. Modulating the Endocannabinoid System in Human Health and Disease—Successes and Failures. FEBS J. 2013, 280, 1918–1943. [Google Scholar] [CrossRef] [Green Version]
- Skaper, S.D.; Facci, L.; Barbierato, M.; Zusso, M.; Bruschetta, G.; Impellizzeri, D.; Cuzzocrea, S.; Giusti, P. N-Palmitoylethanolamine and Neuroinflammation: A Novel Therapeutic Strategy of Resolution. Mol. Neurobiol. 2015, 52, 1034–1042. [Google Scholar] [CrossRef]
- di Marzo, V.D.; Skaper, S. Editorial (Hot Topic: Palmitoylethanolamide: Biochemistry, Pharmacology and Therapeutic Use of a Pleiotropic Anti-Inflammatory Lipid Mediator). CNS Neurol. Disord. Drug Targets 2013, 12, 4–6. [Google Scholar] [CrossRef]
- Re, G.; Barbero, R.; Miolo, A.; Di Marzo, V. Palmitoylethanolamide, Endocannabinoids and Related Cannabimimetic Compounds in Protection against Tissue Inflammation and Pain: Potential Use in Companion Animals. Vet. J. 2007, 173, 21–30. [Google Scholar] [CrossRef]
- Esposito, E.; Cuzzocrea, S. Palmitoylethanolamide in Homeostatic and Traumatic Central Nervous System Injuries. CNS Neurol. Disord. Drug Targets 2013, 12, 55–61. [Google Scholar] [CrossRef]
- Petrosino, S.; Schiano Moriello, A.; Cerrato, S.; Fusco, M.; Puigdemont, A.; De Petrocellis, L.; Di Marzo, V. The Anti-Inflammatory Mediator Palmitoylethanolamide Enhances the Levels of 2-Arachidonoyl-Glycerol and Potentiates Its Actions at TRPV1 Cation Channels. Br. J. Pharmacol. 2016, 173, 1154–1162. [Google Scholar] [CrossRef] [Green Version]
- Iannotti, F.A.; Vitale, R.M. The Endocannabinoid System and PPARs: Focus on Their Signalling Crosstalk, Action and Transcriptional Regulation. Cells 2021, 10, 586. [Google Scholar] [CrossRef]
- Ho, W.-S.V.; Barrett, D.A.; Randall, M.D. “Entourage” Effects of N-Palmitoylethanolamide and N-Oleoylethanolamide on Vasorelaxation to Anandamide Occur through TRPV1 Receptors. Br. J. Pharmacol. 2008, 155, 837–846. [Google Scholar] [CrossRef]
- Hohmann, U.; Pelzer, M.; Kleine, J.; Hohmann, T.; Ghadban, C.; Dehghani, F. Opposite Effects of Neuroprotective Cannabinoids, Palmitoylethanolamide, and 2-Arachidonoylglycerol on Function and Morphology of Microglia. Front. Neurosci. 2019, 13, 1180. [Google Scholar] [CrossRef]
- Lo Verme, J.; Fu, J.; Astarita, G.; La Rana, G.; Russo, R.; Calignano, A.; Piomelli, D. The Nuclear Receptor Peroxisome Proliferator-Activated Receptor-Alpha Mediates the Anti-Inflammatory Actions of Palmitoylethanolamide. Mol. Pharmacol. 2005, 67, 15–19. [Google Scholar] [CrossRef]
- Bougarne, N.; Weyers, B.; Desmet, S.J.; Deckers, J.; Ray, D.W.; Staels, B.; De Bosscher, K. Molecular Actions of PPARα in Lipid Metabolism and Inflammation. Endocr. Rev. 2018, 39, 760–802. [Google Scholar] [CrossRef] [Green Version]
- Koch, M.; Kreutz, S.; Böttger, C.; Benz, A.; Maronde, E.; Ghadban, C.; Korf, H.-W.; Dehghani, F. Palmitoylethanolamide Protects Dentate Gyrus Granule Cells via Peroxisome Proliferator-Activated Receptor-Alpha. Neurotox. Res. 2011, 19, 330–340. [Google Scholar] [CrossRef]
- Di Cesare Mannelli, L.; D’Agostino, G.; Pacini, A.; Russo, R.; Zanardelli, M.; Ghelardini, C.; Calignano, A. Palmitoylethanolamide Is a Disease-Modifying Agent in Peripheral Neuropathy: Pain Relief and Neuroprotection Share a PPAR-Alpha-Mediated Mechanism. Mediat. Inflamm. 2013, 2013, 328797. [Google Scholar] [CrossRef] [Green Version]
- Malin, S.A.; Davis, B.M.; Molliver, D.C. Production of Dissociated Sensory Neuron Cultures and Considerations for Their Use in Studying Neuronal Function and Plasticity. Nat. Protoc. 2007, 2, 152–160. [Google Scholar] [CrossRef]
- Tolkovsky, A.M.; Brelstaff, J. Sensory Neurons from Tau Transgenic Mice and Their Utility in Drug Screening. Methods Mol. Biol. 2018, 1727, 93–105. [Google Scholar] [CrossRef]
- Liebmann, J.; Cook, J.; Lipschultz, C.; Teague, D.; Fisher, J.; Mitchell, J. Cytotoxic Studies of Paclitaxel (Taxol®) in Human Tumour Cell Lines. Br. J. Cancer 1993, 68, 1104–1109. [Google Scholar] [CrossRef] [Green Version]
- Zasadil, L.M.; Andersen, K.A.; Yeum, D.; Rocque, G.B.; Wilke, L.G.; Tevaarwerk, A.J.; Raines, R.T.; Burkard, M.E.; Weaver, B.A. Cytotoxicity of Paclitaxel in Breast Cancer Is Due to Chromosome Missegregation on Multipolar Spindles. Sci. Transl. Med. 2014, 6, 229ra43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musella, A.; Fresegna, D.; Rizzo, F.R.; Gentile, A.; Bullitta, S.; De Vito, F.; Guadalupi, L.; Centonze, D.; Mandolesi, G. A Novel Crosstalk within the Endocannabinoid System Controls GABA Transmission in the Striatum. Sci. Rep. 2017, 7, 7363. [Google Scholar] [CrossRef] [PubMed]
- Scuderi, C.; Valenza, M.; Stecca, C.; Esposito, G.; Carratù, M.R.; Steardo, L. Palmitoylethanolamide Exerts Neuroprotective Effects in Mixed Neuroglial Cultures and Organotypic Hippocampal Slices via Peroxisome Proliferator-Activated Receptor-α. J. Neuroinflammation 2012, 9, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beggiato, S.; Tomasini, M.C.; Cassano, T.; Ferraro, L. Chronic Oral Palmitoylethanolamide Administration Rescues Cognitive Deficit and Reduces Neuroinflammation, Oxidative Stress, and Glutamate Levels in A Transgenic Murine Model of Alzheimer’s Disease. J. Clin. Med. 2020, 9, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pool, M.; Thiemann, J.; Bar-Or, A.; Fournier, A.E. NeuriteTracer: A Novel ImageJ Plugin for Automated Quantification of Neurite Outgrowth. J. Neurosci. Methods 2008, 168, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Klein, I.; Lehmann, H. Pathomechanisms of Paclitaxel-Induced Peripheral Neuropathy. Toxics 2021, 9, 229. [Google Scholar] [CrossRef]
- Mielke, S.; Sparreboom, A.; Mross, K. Peripheral Neuropathy: A Persisting Challenge in Paclitaxel-Based Regimes. Eur. J. Cancer 2006, 42, 24–30. [Google Scholar] [CrossRef]
- Fukuda, Y.; Li, Y.; Segal, R.A. A Mechanistic Understanding of Axon Degeneration in Chemotherapy-Induced Peripheral Neuropathy. Front. Neurosci. 2017, 11, 481. [Google Scholar] [CrossRef] [Green Version]
- Cavaletti, G.; Cavalletti, E.; Oggioni, N.; Sottani, C.; Minoia, C.; D’Incalci, M.; Zucchetti, M.; Marmiroli, P.; Tredici, G. Distribution of Paclitaxel within the Nervous System of the Rat after Repeated Intravenous Administration. Neurotoxicology 2000, 21, 389–393. [Google Scholar] [CrossRef]
- Staff, N.P.; Grisold, A.; Grisold, W.; Windebank, A.J. Chemotherapy-Induced Peripheral Neuropathy: A Current Review. Ann. Neurol. 2017, 81, 772–781. [Google Scholar] [CrossRef]
- Jamieson, S.M.F.; Liu, J.; Hsu, T.; Baguley, B.C.; McKeage, M.J. Paclitaxel Induces Nucleolar Enlargement in Dorsal Root Ganglion Neurons in vivo Reducing Oxaliplatin Toxicity. Br. J. Cancer 2003, 88, 1942–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaggi, A.S.; Singh, N. Mechanisms in Cancer-Chemotherapeutic Drugs-Induced Peripheral Neuropathy. Toxicology 2012, 291, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Carlson, K.; Ocean, A.J. Peripheral Neuropathy with Microtubule-Targeting Agents: Occurrence and Management Approach. Clin. Breast Cancer 2011, 11, 73–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, I.H.; Siddique, R.; Hosmane, S.; Thakor, N.; Höke, A. Compartmentalized Microfluidic Culture Platform to Study Mechanism of Paclitaxel-Induced Axonal Degeneration. Exp. Neurol. 2009, 218, 124–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbuti, A.M.; Chen, Z.-S. Paclitaxel Through the Ages of Anticancer Therapy: Exploring Its Role in Chemoresistance and Radiation Therapy. Cancers 2015, 7, 2360–2371. [Google Scholar] [CrossRef]
- Genovese, T.; Esposito, E.; Mazzon, E.; Paola, R.D.; Meli, R.; Bramanti, P.; Piomelli, D.; Calignano, A.; Cuzzocrea, S. Effects of Palmitoylethanolamide on Signaling Pathways Implicated in the Development of Spinal Cord Injury. J. Pharmacol. Exp. Ther. 2008, 326, 12–23. [Google Scholar] [CrossRef]
- Donvito, G.; Bettoni, I.; Comelli, F.; Colombo, A.; Costa, B. Palmitoylethanolamide Relieves Pain and Preserves Pancreatic Islet Cells in a Murine Model of Diabetes. CNS Neurol. Disord. Drug Targets 2015, 14, 452–462. [Google Scholar] [CrossRef]
- Ghafouri, N.; Ghafouri, B.; Larsson, B.; Stensson, N.; Fowler, C.J.; Gerdle, B. Palmitoylethanolamide and Stearoylethanolamide Levels in the Interstitium of the Trapezius Muscle of Women with Chronic Widespread Pain and Chronic Neck-Shoulder Pain Correlate with Pain Intensity and Sensitivity. Pain 2013, 154, 1649–1658. [Google Scholar] [CrossRef]
- D’Agostino, G.; Russo, R.; Avagliano, C.; Cristiano, C.; Meli, R.; Calignano, A. Palmitoylethanolamide Protects Against the Amyloid-Β25-35-Induced Learning and Memory Impairment in Mice, an Experimental Model of Alzheimer Disease. Neuropsychopharmacology 2012, 37, 1784–1792. [Google Scholar] [CrossRef] [Green Version]
- Lambert, D.M.; Vandevoorde, S.; Diependaele, G.; Govaerts, S.J.; Robert, A.R. Anticonvulsant Activity of N-Palmitoylethanolamide, a Putative Endocannabinoid, in Mice. Epilepsia 2002, 42, 321–327. [Google Scholar] [CrossRef]
- Andresen, S.R.; Bing, J.; Hansen, R.M.; Biering-Sørensen, F.; Johannesen, I.L.; Hagen, E.M.; Rice, A.S.C.; Nielsen, J.F.; Bach, F.W.; Finnerup, N.B. Ultramicronized Palmitoylethanolamide in Spinal Cord Injury Neuropathic Pain: A Randomized, Double-Blind, Placebo-Controlled Trial. Pain 2016, 157, 2097–2103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morsanuto, V.; Galla, R.; Molinari, C.; Uberti, F. A New Palmitoylethanolamide Form Combined with Antioxidant Molecules to Improve Its Effectivess on Neuronal Aging. Brain Sci. 2020, 10, 457. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare Mannelli, L.; Pacini, A.; Corti, F.; Boccella, S.; Luongo, L.; Esposito, E.; Cuzzocrea, S.; Maione, S.; Calignano, A.; Ghelardini, C. Antineuropathic Profile of N-Palmitoylethanolamine in a Rat Model of Oxaliplatin-Induced Neurotoxicity. PLoS ONE 2015, 10, e0128080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truini, A.; Biasiotta, A.; Di Stefano, G.; La Cesa, S.; Leone, C.; Cartoni, C.; Federico, V.T.; Petrucci, M.; Cruccu, G. Palmitoylethanolamide Restores Myelinated-Fibre Function in Patients with Chemotherapy-Induced Painful Neuropathy. CNS Neurol. Disord. Drug Targets 2011, 10, 916–920. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elfarnawany, A.; Dehghani, F. Palmitoylethanolamide Mitigates Paclitaxel Toxicity in Primary Dorsal Root Ganglion Neurons. Biomolecules 2022, 12, 1873. https://doi.org/10.3390/biom12121873
Elfarnawany A, Dehghani F. Palmitoylethanolamide Mitigates Paclitaxel Toxicity in Primary Dorsal Root Ganglion Neurons. Biomolecules. 2022; 12(12):1873. https://doi.org/10.3390/biom12121873
Chicago/Turabian StyleElfarnawany, Amira, and Faramarz Dehghani. 2022. "Palmitoylethanolamide Mitigates Paclitaxel Toxicity in Primary Dorsal Root Ganglion Neurons" Biomolecules 12, no. 12: 1873. https://doi.org/10.3390/biom12121873