
ImmunoPET Directed to the Brain: A New Tool for Preclinical and Clinical Neuroscience

Abstract
:1. Introduction
2. Strategies to Cross the BBB
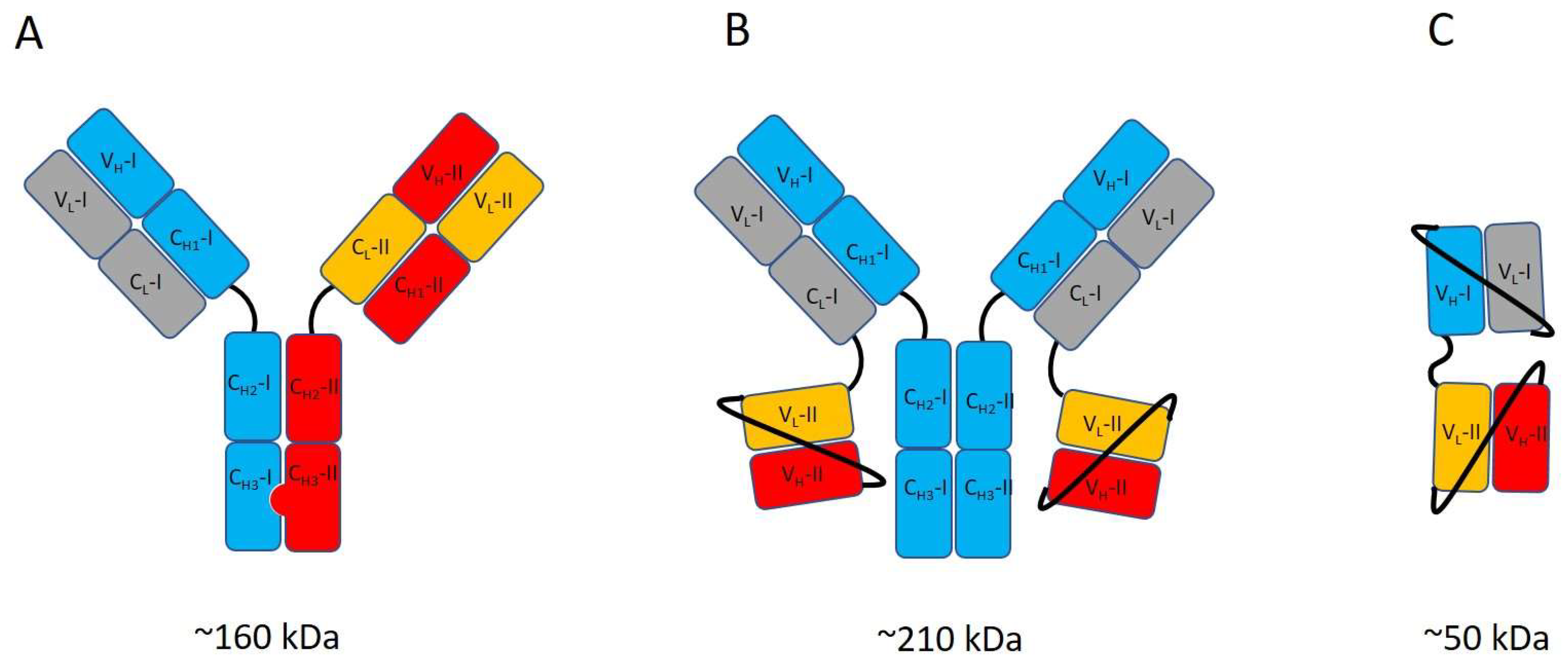
3. Antibody Engineering Strategies for ImmunoPET
4. Radiolabeling Strategies
5. Current Applications
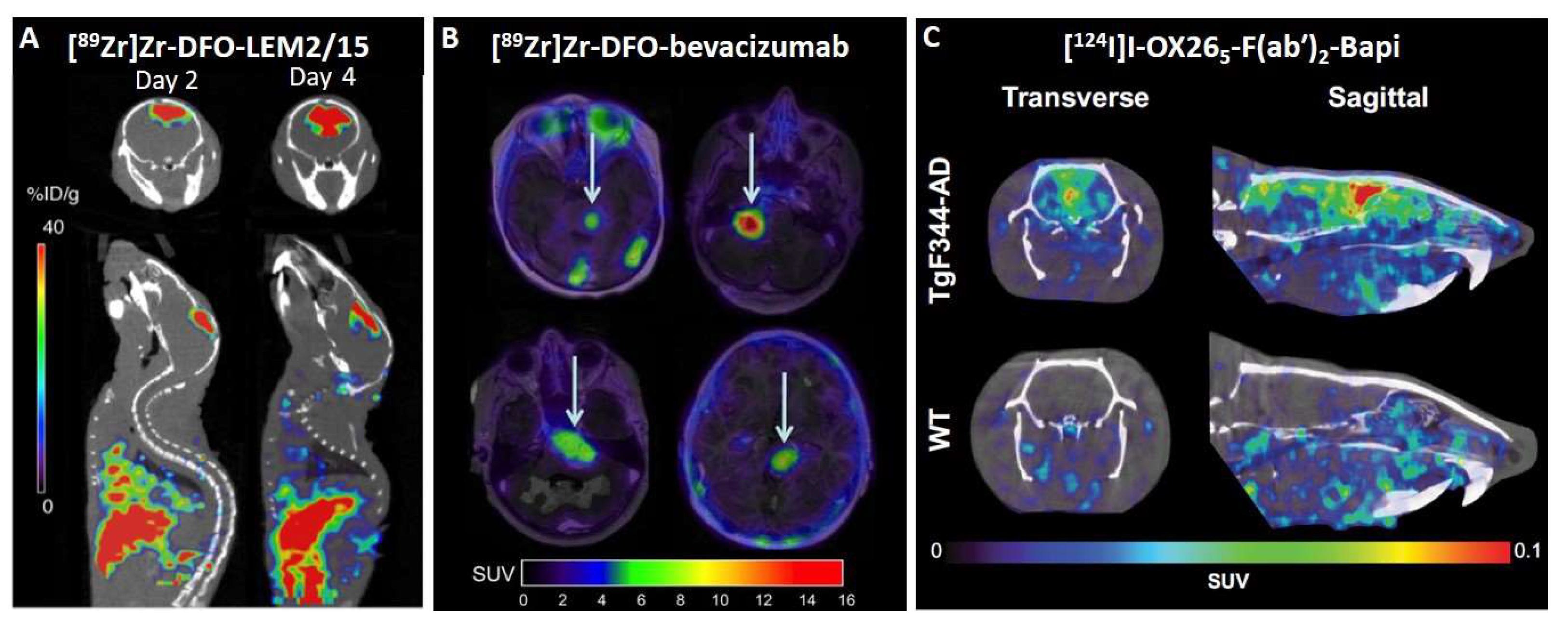
5.1. Neuro-Oncology
5.2. Neurological Diseases
6. Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Marik, J.; Junutula, J.R. Emerging role of immunoPET in receptor targeted cancer therapy. Curr. Drug Deliv. 2011, 8, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Fu, R.; Carroll, L.; Yahioglu, G.; Aboagye, E.O.; Miller, P.W. Antibody Fragment and Affibody ImmunoPET Imaging Agents: Radiolabelling Strategies and Applications. ChemMedChem 2018, 13, 2466–2478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, A.M.; Olafsen, T. Antibodies for molecular imaging of cancer. Cancer J. 2008, 14, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, D.M.; Nabi, H.A. Breast cancer imaging with radiolabeled antibodies. Semin. Nucl. Med. 1999, 29, 41–48. [Google Scholar] [CrossRef] [PubMed]
- McCabe, K.E.; Wu, A.M. Positive progress in immunoPET--not just a coincidence. Cancer Biother. Radiopharm. 2010, 25, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Sehlin, D.; Syvänen, S.; Ballanger, B.; Barthel, H.; Bischof, G.N.; Boche, D.; Boecker, H.; Bohn, K.P.; Borghammer, P.; Cross, D.; et al. Engineered antibodies: New possibilities for brain PET? Eur. J. Nucl. Med. Mol. Imaging 2019, 46, 2848–2858. [Google Scholar] [CrossRef] [Green Version]
- Mäger, I.; Meyer, A.H.; Li, J.; Lenter, M.; Hildebrandt, T.; Leparc, G.; Wood, M.J. Targeting blood-brain-barrier transcytosis-perspectives for drug delivery. Neuropharmacology 2017, 120, 4–7. [Google Scholar] [CrossRef] [Green Version]
- Banks, W.A.; Terrell, B.; Farr, S.A.; Robinson, S.M.; Nonaka, N.; Morley, J.E. Passage of amyloid beta protein antibody across the blood-brain barrier in a mouse model of Alzheimer‘s disease. Peptides 2002, 23, 2223–2226. [Google Scholar] [CrossRef]
- Mach, R.H.; Schwarz, S.W. Challenges for Developing PET Tracers: Isotopes, Chemistry, and Regulatory Aspects. PET Clin. 2010, 5, 131–153. [Google Scholar] [CrossRef]
- Pike, V.W. PET radiotracers: Crossing the blood-brain barrier and surviving metabolism. Trends Pharmacol. Sci. 2009, 30, 431–440. [Google Scholar] [CrossRef]
- Ruiz-López, E.; Calatayud-Pérez, J.; Castells-Yus, I.; Gimeno-Peribáñez, M.J.; Mendoza-Calvo, N.; Morcillo, M.; Schuhmacher, A.J. Diagnosis of Glioblastoma by Immuno-Positron Emission Tomography. Cancers 2021, 14, 74. [Google Scholar] [CrossRef] [PubMed]
- McLean, D.; Cooke, M.J.; Albay, R.; Glabe, C.; Shoichet, M.S. Positron emission tomography imaging of fibrillar parenchymal and vascular amyloid-β in TgCRND8 mice. ACS Chem. Neurosci. 2013, 4, 613–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLean, D.; Cooke, M.J.; Wang, Y.; Green, D.; Fraser, P.E.; George-Hyslop, P.S.; Shoichet, M.S. Anti-amyloid-β-mediated positron emission tomography imaging in Alzheimer’s disease mouse brains. PLoS ONE 2012, 7, e51958. [Google Scholar] [CrossRef] [PubMed]
- Hervé, F.; Ghinea, N.; Scherrmann, J.M. CNS delivery via adsorptive transcytosis. AAPS J. 2008, 10, 455–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Bourgeois, J.-P.; Celli, S.; Glacial, F.; Le Sourd, A.-M.; Mecheri, S.; Weksler, B.; Romero, I.; Couraud, P.-O.; Rougeon, F.; et al. Cell-penetrating anti-GFAP VHH and corresponding fluorescent fusion protein VHH-GFP spontaneously cross the blood-brain barrier and specifically recognize astrocytes: Application to brain imaging. FASEB J. 2012, 26, 3969–3979. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Vandesquille, M.; Koukouli, F.; Dudeffant, C.; Youssef, I.; Lenormand, P.; Ganneau, C.; Maskos, U.; Czech, C.; Grueninger, F. Camelid single-domain antibodies: A versatile tool for in vivo imaging of extracellular and intracellular brain targets. J. Control. Release 2016, 243, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Terstappen, G.C.; Meyer, A.H.; Bell, R.D.; Zhang, W. Strategies for delivering therapeutics across the blood-brain barrier. Nat. Rev. Drug Discov. 2021, 20, 362–383. [Google Scholar] [CrossRef]
- Pulgar, V.M. Transcytosis to Cross the Blood Brain Barrier, New Advancements and Challenges. Front. Neurosci. 2018, 12, 1019. [Google Scholar] [CrossRef]
- Kouhi, A.; Pachipulusu, V.; Kapenstein, T.; Hu, P.; Epstein, A.L.; Khawli, L.A. Brain Disposition of Antibody-Based Therapeutics: Dogma, Approaches and Perspectives. Int. J. Mol. Sci. 2021, 22, 6442. [Google Scholar] [CrossRef]
- Yu, Y.J.; Zhang, Y.; Kenrick, M.; Hoyte, K.; Luk, W.; Lu, Y.; Atwal, J.; Elliott, J.M.; Prabhu, S.; Watts, R.J.; et al. Boosting brain uptake of a therapeutic antibody by reducing its affinity for a transcytosis target. Sci. Transl. Med. 2011, 3, 84ra44. [Google Scholar] [CrossRef]
- Niewoehner, J.; Bohrmann, B.; Collin, L.; Urich, E.; Sade, H.; Maier, P.; Rueger, P.; Stracke, J.O.; Lau, W.; Tissot, A.C.; et al. Increased brain penetration and potency of a therapeutic antibody using a monovalent molecular shuttle. Neuron 2014, 81, 49–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehlin, D.; Fang, X.T.; Cato, L.; Antoni, G.; Lannfelt, L.; Syvänen, S. Antibody-based PET imaging of amyloid beta in mouse models of Alzheimer’s disease. Nat. Commun. 2016, 7, 10759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehlin, D.; Fang, X.T.; Meier, S.R.; Jansson, M.; Syvänen, S. Pharmacokinetics, biodistribution and brain retention of a bispecific antibody-based PET radioligand for imaging of amyloid-β. Sci. Rep. 2017, 7, 17254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hultqvist, G.; Syvänen, S.; Fang, X.T.; Lannfelt, L.; Sehlin, D. Bivalent Brain Shuttle Increases Antibody Uptake by Monovalent Binding to the Transferrin Receptor. Theranostics 2017, 7, 308–318. [Google Scholar] [CrossRef] [PubMed]
- Meier, S.R.; Syvänen, S.; Hultqvist, G.; Fang, X.T.; Roshanbin, S.; Lannfelt, L.; Neumann, U.; Sehlin, D. Antibody-Based In Vivo PET Imaging Detects Amyloid-β Reduction in Alzheimer Transgenic Mice After BACE-1 Inhibition. J. Nucl. Med. 2018, 59, 1885–1891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, S.R.; Sehlin, D.; Roshanbin, S.; Falk, V.L.; Saito, T.; Saido, T.C.; Neumann, U.; Rokka, J.; Eriksson, J.; Syvänen, S. (11)C-PiB and (124)I-Antibody PET Provide Differing Estimates of Brain Amyloid-β After Therapeutic Intervention. J. Nucl. Med. 2022, 63, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Syvänen, S.; Fang, X.T.; Faresjö, R.; Rokka, J.; Lannfelt, L.; E Olberg, D.; Eriksson, J.; Sehlin, D. Fluorine-18-Labeled Antibody Ligands for PET Imaging of Amyloid-β in Brain. ACS Chem. Neurosci. 2020, 11, 4460–4468. [Google Scholar] [CrossRef]
- Syvänen, S.; Fang, X.T.; Hultqvist, G.; Meier, S.R.; Lannfelt, L.; Sehlin, D. A bispecific Tribody PET radioligand for visualization of amyloid-beta protofibrils—A new concept for neuroimaging. Neuroimage 2017, 148, 55–63. [Google Scholar] [CrossRef]
- Fang, X.T.; Hultqvist, G.; Meier, S.R.; Antoni, G.; Sehlin, D.; Syvänen, S. High detection sensitivity with antibody-based PET radioligand for amyloid beta in brain. Neuroimage 2019, 184, 881–888. [Google Scholar] [CrossRef]
- Faresjö, R.; Bonvicini, G.; Fang, X.T.; Aguilar, X.; Sehlin, D.; Syvänen, S. Brain pharmacokinetics of two BBB penetrating bispecific antibodies of different size. Fluids Barriers CNS 2021, 18, 26. [Google Scholar] [CrossRef]
- Sehlin, D.; Stocki, P.; Gustavsson, T.; Hultqvist, G.; Walsh, F.S.; Rutkowski, J.L.; Syvänen, S. Brain delivery of biologics using a cross-species reactive transferrin receptor 1 VNAR shuttle. FASEB J. 2020, 34, 13272–13283. [Google Scholar] [CrossRef] [PubMed]
- Bonvicini, G.; Syvänen, S.; Andersson, K.G.; Haaparanta-Solin, M.; López-Picón, F.; Sehlin, D. ImmunoPET imaging of amyloid-beta in a rat model of Alzheimer’s disease with a bispecific, brain-penetrating fusion protein. Transl. Neurodegener. 2022, 11, 55. [Google Scholar] [CrossRef] [PubMed]
- Neuwelt, E.A.; Diehl, J.T.; Vu, L.H.; Hill, S.A.; Michael, A.J.; Frenkel, E.P. Monitoring of methotrexate delivery in patients with malignant brain tumors after osmotic blood-brain barrier disruption. Ann. Intern. Med. 1981, 94, 449–454. [Google Scholar] [CrossRef]
- Arif, W.M.; Elsinga, P.H.; Gasca-Salas, C.; Versluis, M.; Martínez-Fernández, R.; Dierckx, R.A.; Borra, R.J.; Luurtsema, G. Focused ultrasound for opening blood-brain barrier and drug delivery monitored with positron emission tomography. J. Control. Release 2020, 324, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Leinenga, G.; Bodea, L.G.; Koh, W.K.; Nisbet, R.M.; Götz, J. Delivery of Antibodies into the Brain Using Focused Scanning Ultrasound. J. Vis. Exp. 2020, 161, e61372. [Google Scholar] [CrossRef]
- Arvanitis, C.D.; Askoxylakis, V.; Guo, Y.; Datta, M.; Kloepper, J.; Ferraro, G.B.; Bernabeu, M.O.; Fukumura, D.; McDannold, N.; Jain, R.K. Mechanisms of enhanced drug delivery in brain metastases with focused ultrasound-induced blood-tumor barrier disruption. Proc. Natl. Acad. Sci. USA 2018, 115, E8717–E8726. [Google Scholar] [CrossRef] [Green Version]
- Tran, V.L.; Novell, A.; Tournier, N.; Gerstenmayer, M.; Schweitzer-Chaput, A.; Mateos, C.; Jego, B.; Bouleau, A.; Nozach, H.; Winkeler, A.; et al. Impact of blood-brain barrier permeabilization induced by ultrasound associated to microbubbles on the brain delivery and kinetics of cetuximab: An immunoPET study using. J. Control. Release 2020, 328, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Sheybani, N.; Breza, V.; Paul, S.; McCauley, K.; Berr, S.; Miller, W.; Neumann, K.; Price, R. ImmunoPET-informed sequence for focused ultrasound-targeted mCD47 blockade controls glioma. J. Control. Release 2021, 331, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Poduslo, J.F.; Curran, G.L.; Berg, C.T. Macromolecular permeability across the blood-nerve and blood-brain barriers. Proc. Natl. Acad. Sci. USA 1994, 91, 5705–5709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiess, C.; Zhai, Q.; Carter, P.J. Alternative molecular formats and therapeutic applications for bispecific antibodies. Mol. Immunol. 2015, 67, 95–106. [Google Scholar] [CrossRef]
- Merchant, A.M.; Zhu, Z.; Yuan, J.Q.; Goddard, A.; Adams, C.W.; Presta, L.G.; Carter, P. An efficient route to human bispecific IgG. Nat. Biotechnol. 1998, 16, 677–681. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, W.; Regula, J.T.; Bähner, M.; Schanzer, J.; Croasdale, R.; Dürr, H.; Gassner, C.; Georges, G.; Kettenberger, H.; Imhof-Jung, S.; et al. Immunoglobulin domain crossover as a generic approach for the production of bispecific IgG antibodies. Proc. Natl. Acad. Sci. USA 2011, 108, 11187–11192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, S.M.; Wu, X.; Pustilnik, A.; Sereno, A.; Huang, F.; Rick, H.L.; Guntas, G.; Leaver-Fay, A.; Smith, E.M.; Ho, C.; et al. Generation of bispecific IgG antibodies by structure-based design of an orthogonal Fab interface. Nat. Biotechnol. 2014, 32, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Coloma, M.J.; Morrison, S.L. Design and production of novel tetravalent bispecific antibodies. Nat. Biotechnol. 1997, 15, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Yazaki, P.J.; Lee, B.; Channappa, D.; Cheung, C.W.; Crow, D.; Chea, J.; Poku, E.; Li, L.; Andersen, J.T.; Sandlie, I.; et al. A series of anti-CEA/anti-DOTA bispecific antibody formats evaluated for pre-targeting: Comparison of tumor uptake and blood clearance. Protein Eng. Des. Sel. 2013, 26, 187–193. [Google Scholar] [CrossRef]
- Mallender, W.D.; Voss, E.W., Jr. Construction, expression, and activity of a bivalent bispecific single-chain antibody. J. Biol. Chem. 1994, 269, 199–206. [Google Scholar] [CrossRef]
- Wolf, E.; Hofmeister, R.; Kufer, P.; Schlereth, B.; Baeuerle, P.A. BiTEs: Bispecific antibody constructs with unique anti-tumor activity. Drug Discov. Today 2005, 10, 1237–1244. [Google Scholar] [CrossRef]
- Kariolis, M.S.; Wells, R.C.; Getz, J.A.; Kwan, W.; Mahon, C.S.; Tong, R.; Kim, D.J.; Srivastava, A.; Bedard, C.; Henne, K.R.; et al. Brain delivery of therapeutic proteins using an Fc fragment blood-brain barrier transport vehicle in mice and monkeys. Sci. Transl. Med. 2020, 12, eaay1359. [Google Scholar] [CrossRef]
- Meier, S.R.; Sehlin, D.; Syvänen, S. Passive and receptor mediated brain delivery of an anti-GFAP nanobody. Nucl. Med. Biol. 2022, 114–115, 128–134. [Google Scholar] [CrossRef]
- Vorobyeva, A.; Schulga, A.; Konovalova, E.; Mitran, B.; Garousi, J.; Rinne, S.; Orlova, A.; Deyev, S.; Tolmachev, V. Comparison of tumor-targeting properties of directly and indirectly radioiodinated designed ankyrin repeat protein (DARPin) G3 variants for molecular imaging of HER2. Int. J. Oncol. 2019, 54, 1209–1220. [Google Scholar] [CrossRef]
- Brinkmann, U.; Kontermann, R.E. The making of bispecific antibodies. MAbs 2017, 9, 182–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L. Pharmacokinetics of monoclonal antibodies and Fc-fusion proteins. Protein Cell 2018, 9, 15–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mammen, M.; Choi, S.K.; Whitesides, G.M. Polyvalent Interactions in Biological Systems: Implications for Design and Use of Multivalent Ligands and Inhibitors. Angew. Chem. Int. Ed. Engl. 1998, 37, 2754–2794. [Google Scholar] [CrossRef]
- Datta-Mannan, A.; Brown, R.M.; Fitchett, J.; Heng, A.R.; Balasubramaniam, D.; Pereira, J.; Croy, J.E. Modulation of the Biophysical Properties of Bifunctional Antibodies as a Strategy for Mitigating Poor Pharmacokinetics. Biochemistry 2019, 58, 3116–3132. [Google Scholar] [CrossRef] [PubMed]
- Shukla, A.K.; Kumar, U. Positron emission tomography: An overview. J. Med. Phys. 2006, 31, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.M. Antibodies and antimatter: The resurgence of immuno-PET. J. Nucl. Med. 2009, 50, 2–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, M.; Eriksson, L. Physics of pure and non-pure positron emitters for PET: A review and a discussion. EJNMMI Phys. 2016, 3, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunka, M.; Müller, C.; Vermeulen, C.; Haller, S.; Türler, A.; Schibli, R.; van der Meulen, N.P. Imaging quality of (44)Sc in comparison with five other PET radionuclides using Derenzo phantoms and preclinical PET. Appl. Radiat. Isot. 2016, 110, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Kręcisz, P.; Czarnecka, K.; Królicki, L.; Mikiciuk-Olasik, E.; Szymański, P. Radiolabeled Peptides and Antibodies in Medicine. Bioconjug Chem. 2021, 32, 25–42. [Google Scholar] [CrossRef]
- Oroujeni, M.; Garousi, J.; Andersson, K.G.; Löfblom, J.; Mitran, B.; Orlova, A.; Tolmachev, V. Preclinical Evaluation of [68Ga]Ga-DFO-ZEGFR:2377: A Promising Affibody-Based Probe for Noninvasive PET Imaging of EGFR Expression in Tumors. Cells 2018, 7, 141. [Google Scholar] [CrossRef]
- Chakravarty, R.; Goel, S.; Valdovinos, H.F.; Hernandez, R.; Hong, H.; Nickles, R.J.; Cai, W. Matching the decay half-life with the biological half-life: ImmunoPET imaging with 44Sc-labeled cetuximab Fab fragment. Bioconjug. Chem. 2014, 25, 2197–2204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stéen, E.J.L.; Edem, P.E.; Nørregaard, K.; Jørgensen, J.T.; Shalgunov, V.; Kjaer, A.; Herth, M.M. Pretargeting in nuclear imaging and radionuclide therapy: Improving efficacy of theranostics and nanomedicines. Biomaterials 2018, 179, 209–245. [Google Scholar] [CrossRef] [PubMed]
- Rossin, R.; Verkerk, P.R.; Bosch, S.M.V.D.; Vulders, R.C.M.; Verel, I.; Lub, J.; Robillard, M.S. In vivo chemistry for pretargeted tumor imaging in live mice. Angew. Chem. Int. Ed. Engl. 2010, 49, 3375–3378. [Google Scholar] [CrossRef] [PubMed]
- Carroll, L.; Evans, H.L.; Aboagye, E.O.; Spivey, A.C. Bioorthogonal chemistry for pre-targeted molecular imaging--progress and prospects. Org. Biomol. Chem. 2013, 11, 5772–5781. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, B.L.; Guo, Z.; Bernardes, G.J.L. Inverse electron demand Diels-Alder reactions in chemical biology. Chem. Soc. Rev. 2017, 46, 4895–4950. [Google Scholar] [CrossRef] [Green Version]
- Poulie, C.B.; Jørgensen, J.T.; Shalgunov, V.; Kougioumtzoglou, G.; Jeppesen, T.E.; Kjaer, A.; Herth, M.M. Evaluation of [64Cu]Cu-NOTA-PEG7-H-Tz for Pretargeted Imaging in LS174T Xenografts-Comparison to [111In]In-DOTA-PEG11-BisPy-Tz. Molecules 2021, 26, 544. [Google Scholar] [CrossRef]
- Stéen, E.J.L.; Jørgensen, J.T.; Denk, C.; Battisti, U.M.; Nørregaard, K.; Edem, P.E.; Bratteby, K.; Shalgunov, V.; Wilkovitsch, M.; Svatunek, D.; et al. Lipophilicity and Click Reactivity Determine the Performance of Bioorthogonal Tetrazine Tools in Pretargeted In Vivo Chemistry. ACS Pharmacol. Transl. Sci. 2021, 4, 824–833. [Google Scholar] [CrossRef]
- Shalgunov, V.; Broek, S.L.v.D.; Andersen, I.V.; Vázquez, R.G.; Raval, N.R.; Palner, M.; Mori, Y.; Schäfer, G.; Herrmann, B.; Mikula, H.; et al. Pretargeted imaging beyond the blood–brain barrier-Utopia or Feasible? Pharmaceuticals (Basel) 2022, 15, 1191. [Google Scholar]
- Dijkers, E.C.; Oude Munnink, T.H.; Kosterink, J.G.; Brouwers, A.H.; Jager, P.L.; De Jong, J.R.; Van Dongen, G.A.; Schroder, C.P.; Lub-de Hooge, M.N.; de Vries, E.G. Biodistribution of 89Zr-trastuzumab and PET imaging of HER2-positive lesions in patients with metastatic breast cancer. Clin. Pharmacol. Ther. 2010, 87, 586–592. [Google Scholar] [CrossRef]
- Ulaner, G.A.; Lyashchenko, S.K.; Riedl, C.; Ruan, S.; Zanzonico, P.B.; Lake, D.; Jhaveri, K.; Zeglis, B.; Lewis, J.S.; O’Donoghue, J.A. First-in-Human Human Epidermal Growth Factor Receptor 2-Targeted Imaging Using 89Zr-Pertuzumab PET/CT: Dosimetry and Clinical Application in Patients with Breast Cancer. J. Nucl. Med. 2018, 59, 900–906. [Google Scholar] [CrossRef] [Green Version]
- Rousseau, C.; Goldenberg, D.; Colombié, M.; Sébille, J.-C.; Meingan, P.; Ferrer, L.; Baumgartner, P.; Cerato, E.; Masson, D.; Campone, M.; et al. Initial Clinical Results of a Novel Immuno-PET Theranostic Probe in Human Epidermal Growth Factor Receptor 2-Negative Breast Cancer. J. Nucl. Med. 2020, 61, 1205–1211. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; McDougald, D.; Meshaw, R.; Balyasnikova, I.; Zalutsky, M.R.; Vaidyanathan, G. Labeling single domain antibody fragments with (18)F using a novel residualizing prosthetic agent-N-succinimidyl 3-(1-(2-(2-(2-(2-[(18)F]fluoroethoxy)ethoxy)ethoxy)ethyl)-1H-1,2,3-triazol-4-yl)-5-(guanidinomethyl)benzoate. Nucl. Med. Biol. 2021, 100–101, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Chen, W. Clinical applications of PET in brain tumors. J. Nucl. Med. 2007, 48, 1468–1481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, W.; Wu, Y.; Chen, K.; Cao, Q.; Tice, D.A.; Chen, X. In vitro and in vivo characterization of 64Cu-labeled Abegrin, a humanized monoclonal antibody against integrin alpha v beta 3. Cancer Res. 2006, 66, 9673–9681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Hernandez, R.; Hong, H.; Graves, S.A.; Yang, Y.; England, C.G.; Theuer, C.P.; Nickles, R.J.; Cai, W. Noninvasive brain cancer imaging with a bispecific antibody fragment, generated via click chemistry. Proc. Natl. Acad. Sci. USA 2015, 112, 12806–12811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Hu, Y.; Liu, W.; Chen, L.; Zhao, Y.; Ma, H.; Yang, J.; Yang, Y.; Liao, J.; Cai, J.; et al. A radiopharmaceutical [89Zr]Zr-DFO-nimotuzumab for immunoPET with epidermal growth factor receptor expression in vivo. Nucl. Med. Biol. 2019, 70, 23–31. [Google Scholar] [CrossRef]
- Zhou, B.; Wang, H.; Liu, R.; Wang, M.; Deng, H.; Giglio, B.C.; Gill, P.S.; Shan, H.; Li, Z. PET Imaging of Dll4 Expression in Glioblastoma and Colorectal Cancer Xenografts Using (64)Cu-Labeled Monoclonal Antibody 61B. Mol. Pharm. 2015, 12, 3527–3534. [Google Scholar] [CrossRef] [Green Version]
- Pandya, D.; Sinha, A.; Yuan, H.; Mutkus, L.; Stumpf, K.; Marini, F.; Wadas, T. Imaging of Fibroblast Activation Protein Alpha Expression in a Preclinical Mouse Model of Glioma Using Positron Emission Tomography. Molecules 2020, 25, 3672. [Google Scholar] [CrossRef]
- Hernandez, R.; Sun, H.; England, C.G.; Valdovinos, H.F.; Barnhart, T.E.; Yang, Y.; Cai, W. ImmunoPET Imaging of CD146 Expression in Malignant Brain Tumors. Mol. Pharm. 2016, 13, 2563–2570. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Hernandez, R.; Rao, J.; Yin, L.; Qu, Y.; Wu, J.; England, C.G.; Graves, S.A.; Lewis, C.M.; Wang, P.; et al. Targeting CD146 with a 64Cu-labeled antibody enables in vivo immunoPET imaging of high-grade gliomas. Proc. Natl. Acad. Sci. USA 2015, 112, E6525–E6534. [Google Scholar] [CrossRef] [Green Version]
- De Lucas, A.G.; Schuhmacher, A.J.; Oteo, M.; Romero, E.; Cámara, J.A.; de Martino, A.; Arroyo, A.G.; Morcillo, M.Á.; Squatrito, M.; Martinez-Torrecuadrada, J.L.; et al. Targeting MT1-MMP as an ImmunoPET-Based Strategy for Imaging Gliomas. PLoS ONE 2016, 11, e0158634. [Google Scholar] [CrossRef] [PubMed]
- Nigam, S.; McCarl, L.; Kumar, R.; Edinger, R.S.; Kurland, B.F.; Anderson, C.J.; Panigrahy, A.; Kohanbash, G.; Edwards, W.B. Preclinical ImmunoPET Imaging of Glioblastoma-Infiltrating Myeloid Cells Using Zirconium-89 Labeled Anti-CD11b Antibody. Mol. Imaging Biol. 2020, 22, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Jansen, M.H.; Lagerweij, T.; Sewing, A.C.P.; Vugts, D.J.; van Vuurden, D.G.; Molthoff, C.F.; Caretti, V.; Veringa, S.J.; Petersen, N.; Carcaboso, A.M.; et al. Bevacizumab Targeting Diffuse Intrinsic Pontine Glioma: Results of 89Zr-Bevacizumab PET Imaging in Brain Tumor Models. Mol. Cancer Ther. 2016, 15, 2166–2174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, M.H.; van Zanten, S.E.M.V.; van Vuurden, D.G.; Huisman, M.C.; Vugts, D.J.; Hoekstra, O.S.; van Dongen, G.A.; Kaspers, G.-J.L. Molecular Drug Imaging: (89)Zr-Bevacizumab PET in Children with Diffuse Intrinsic Pontine Glioma. J. Nucl. Med. 2017, 58, 711–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Zanten, S.E.V.; Sewing, A.C.P.; Van Lingen, A.; Hoekstra, O.S.; Wesseling, P.; Meel, M.H.; Van Vuurden, D.G.; Kaspers, G.J.; Hulleman, E.; Bugiani, M. Multiregional Tumor Drug-Uptake Imaging by PET and Microvascular Morphology in End-Stage Diffuse Intrinsic Pontine Glioma. J. Nucl. Med. 2018, 59, 612–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- den Hollander, M.W.; Bensch, F.; Glaudemans, A.W.; Munnink, T.H.O.; Enting, R.H.; den Dunnen, W.F.; Heesters, M.A.; Kruyt, F.A.; Lub-de Hooge, M.N.; de Groot, J.C.; et al. TGF-β Antibody Uptake in Recurrent High-Grade Glioma Imaged with 89Zr-Fresolimumab PET. J. Nucl. Med. 2015, 56, 1310–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Junca, A.; Reiners, O.; Borrero-Garcia, L.D.; Beckford-Vera, D.; Lazar, A.A.; Chou, W.; Braunstein, S.; VanBrocklin, H.; Franc, B.L.; Barcellos-Hoff, M.H. Positron Emission Tomography Imaging of Functional Transforming Growth Factor β (TGFβ) Activity and Benefit of TGFβ Inhibition in Irradiated Intracranial Tumors. Int J Radiat Oncol Biol Phys. 2021, 109, 527–539. [Google Scholar] [CrossRef]
- Krebs, S.; Grommes, C.; McDevitt, M.R.; Carlin, S.D.; O’Donoghue, J.A.; Graham, M.S.; Young, R.J.; Schöder, H.; Gutin, P.H.; Bander, N.H.; et al. [89Zr]Zr-huJ591 immuno-PET targeting PSMA in IDH mutant anaplastic oligodendroglioma. Eur. J. Nucl. Med. Mol. Imaging 2022, 49, 783–785. [Google Scholar] [CrossRef]
- Matsuda, M.; Ishikawa, E.; Yamamoto, T.; Hatano, K.; Joraku, A.; Iizumi, Y.; Masuda, Y.; Nishiyama, H.; Matsumura, A. Potential use of prostate specific membrane antigen (PSMA) for detecting the tumor neovasculature of brain tumors by PET imaging with 89Zr-Df-IAB2M anti-PSMA minibody. J. Neurooncol. 2018, 138, 581–589. [Google Scholar] [CrossRef]
- Sarkaria, J.N.; Hu, L.S.; Parney, I.F.; Pafundi, D.H.; Brinkmann, D.H.; Laack, N.N.; Giannini, C.; Burns, T.C.; Kizilbash, S.; Laramy, J.K.; et al. Is the blood-brain barrier really disrupted in all glioblastomas? A critical assessment of existing clinical data. Neuro Oncol. 2018, 20, 184–191. [Google Scholar] [CrossRef]
- Zaky, W.; Wellner, M.; Brown, R.J.; Blüml, S.; Finlay, J.L.; Dhall, G. Treatment of children with diffuse intrinsic pontine gliomas with chemoradiotherapy followed by a combination of temozolomide, irinotecan, and bevacizumab. Pediatr. Hematol. Oncol. 2013, 30, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, D.G.; Mazewski, C.; Hayes, L.; Jordan, C.; Esiashivilli, N.; Janns, A.; MacDonald, T.J. Prolonged survival after treatment of diffuse intrinsic pontine glioma with radiation, temozolamide, and bevacizumab: Report of 2 cases. J. Pediatr. Hematol. Oncol. 2013, 35, e42-6. [Google Scholar] [CrossRef] [PubMed]
- Fissers, J.; Waldron, A.-M.; De Vijlder, T.; Van Broeck, B.; Pemberton, D.J.; Mercken, M.; Van Der Veken, P.; Joossens, J.; Augustyns, K.; Dedeurwaerdere, S.; et al. Synthesis and Evaluation of a Zr-89-Labeled Monoclonal Antibody for Immuno-PET Imaging of Amyloid-β Deposition in the Brain. Mol. Imaging Biol. 2016, 18, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, K.; Sehlin, D.; Syvänen, S.; Svedberg, M.M.; Philipson, O.; Söderberg, L.; Tegerstedt, K.; Holmquist, M.; Gellerfors, P.; Tolmachev, V.; et al. Specific uptake of an amyloid-β protofibril-binding antibody-tracer in AβPP transgenic mouse brain. J. Alzheimers Dis. 2013, 37, 29–40. [Google Scholar] [CrossRef]
- Roshanbin, S.; Xiong, M.; Hultqvist, G.; Söderberg, L.; Zachrisson, O.; Meier, S.; Ekmark-Lewén, S.; Bergström, J.; Ingelsson, M.; Sehlin, D.; et al. In vivo imaging of alpha-synuclein with antibody-based PET. Neuropharmacolog 2022, 208, 108985. [Google Scholar] [CrossRef] [PubMed]
- Morito, T.; Harada, R.; Iwata, R.; Du, Y.; Okamura, N.; Kudo, Y.; Yanai, K. Synthesis and pharmacokinetic characterisation of a fluorine-18 labelled brain shuttle peptide fusion dimeric affibody. Sci. Rep. 2021, 11, 2588. [Google Scholar] [CrossRef] [PubMed]
- Meier, S.R.; Sehlin, D.; Hultqvist, G.; Syvänen, S. Pinpointing Brain TREM2 Levels in Two Mouse Models of Alzheimer’s Disease. Mol. Imaging Biol. 2021, 23, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Bar-Or, A.; Calabresi, P.A.J.; Arnold, D.; Markowitz, C.; Shafer, S.; Kasper, L.H.; Waubant, E.; Gazda, S.; Fox, R.J.; Panzara, M.; et al. Rituximab in relapsing-remitting multiple sclerosis: A 72-week, open-label, phase I trial. Ann Neurol. 2008, 63, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Bar-Or, A.; Grove, R.A.; Austin, D.J.; Tolson, J.M.; VanMeter, S.A.; Lewis, E.W.; Derosier, F.J.; Lopez, M.C.; Kavanagh, S.T.; Miller, A.E.; et al. Subcutaneous ofatumumab in patients with relapsing-remitting multiple sclerosis: The MIRROR study. Neurology 2018, 90, e1805–e1814. [Google Scholar] [CrossRef]
- Hauser, S.L.; Bar-Or, A.; Comi, G.; Giovannoni, G.; Hartung, H.-P.; Hemmer, B.; Lublin, F.; Montalban, X.; Rammohan, K.W.; Selmaj, K.; et al. Ocrelizumab versus Interferon Beta-1a in Relapsing Multiple Sclerosis. N. Engl. J. Med. 2017, 376, 221–234. [Google Scholar] [CrossRef]
- James, M.L.; Hoehne, A.; Mayer, A.T.; Lechtenberg, K.; Moreno, M.; Gowrishankar, G.; Ilovich, O.; Natarajan, A.; Johnson, E.M.; Nguyen, J.; et al. Imaging B Cells in a Mouse Model of Multiple Sclerosis Using (64)Cu-Rituximab PET. J. Nucl. Med. 2017, 58, 1845–1851. [Google Scholar] [CrossRef] [PubMed]
- Migotto, M.-A.; Mardon, K.; Orian, J.; Weckbecker, G.; Kneuer, R.; Bhalla, R.; Reutens, D.C. Efficient Distribution of a Novel Zirconium-89 Labeled Anti-cd20 Antibody Following Subcutaneous and Intravenous Administration in Control and Experimental Autoimmune Encephalomyelitis-Variant Mice. Front. Immunol. 2019, 10, 2437. [Google Scholar] [CrossRef] [PubMed]
- Werner, J.M.; Lohmann, P.; Fink, G.R.; Langen, K.J.; Galldiks, N. Current Landscape and Emerging Fields of PET Imaging in Patients with Brain Tumors. Molecules 2020, 25, 1471. [Google Scholar] [CrossRef] [Green Version]
- Dupont, A.C.; Largeau, B.; Guilloteau, D.; Santiago Ribeiro, M.J.; Arlicot, N. The Place of PET to Assess New Therapeutic Effectiveness in Neurodegenerative Diseases. Contrast Media Mol. Imaging 2018, 2018, 7043578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werry, E.L.; Bright, F.M.; Piguet, O.; Ittner, L.M.; Halliday, G.M.; Hodges, J.R.; Kiernan, M.C.; Loy, C.T.; Kril, J.J.; Kassiou, M. Recent Developments in TSPO PET Imaging as A Biomarker of Neuroinflammation in Neurodegenerative Disorders. Int. J. Mol. Sci. 2019, 20, 3161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaino, W.; Janssen, B.; Vugts, D.J.; de Vries, H.E.; Windhorst, A.D. Towards PET imaging of the dynamic phenotypes of microglia. Clin. Exp. Immunol. 2021, 206, 282–300. [Google Scholar] [CrossRef]
- Narayanaswami, V.; Dahl, K.; Bernard-Gauthier, V.; Josephson, L.; Cumming, P.; Vasdev, N. Emerging PET Radiotracers and Targets for Imaging of Neuroinflammation in Neurodegenerative Diseases: Outlook Beyond TSPO. Mol. Imaging 2018, 17, 1536012118792317. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Hu, K.; Shao, T.; Hou, L.; Zhang, S.; Ye, W.; Josephson, L.; Meyer, J.H.; Zhang, M.-R.; Vasdev, N.; et al. Recent developments on PET radiotracers for TSPO and their applications in neuroimaging. Acta Pharm. Sin. B. 2021, 11, 373–393. [Google Scholar] [CrossRef]
- Tiepolt, S.; Meyer, P.M.; Patt, M.; Deuther-Conrad, W.; Hesse, S.; Barthel, H.; Sabri, O. PET Imaging of Cholinergic Neurotransmission in Neurodegenerative Disorders. J. Nucl. Med. 2022, 63, 33s–44s. [Google Scholar] [CrossRef]
- Ozenil, M.; Aronow, J.; Millard, M.; Langer, T.; Wadsak, W.; Hacker, M.; Pichler, V. Update on PET Tracer Development for Muscarinic Acetylcholine Receptors. Pharmaceuticals 2021, 14, 530. [Google Scholar] [CrossRef]
- Tolosa, E.; Garrido, A.; Scholz, S.W.; Poewe, W. Challenges in the diagnosis of Parkinson’s disease. Lancet Neurol. 2021, 20, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Waszkiewicz, N. Mentally Sick or Not-(Bio)Markers of Psychiatric Disorders Needed. J. Clin. Med. 2020, 9, 2375. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.F.; Tso, I.F. GABA abnormalities in schizophrenia: A methodological review of in vivo studies. Schizophr. Res. 2015, 167, 84–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parot, V.; Herraiz, J.L.; Dave, S.R.; Udías, J.M.; Moore, S.C.; Park, M.-A.; Vaquero, J.J.; Lage, E. A New Approach for Multiplexed PET Imaging; IEEE NSS-MIC: Seoul, Korea, 2013. [Google Scholar]
- Slart, R.H.J.A.; Tsoumpas, C.; Glaudemans, A.W.J.M.; Noordzij, W.; Willemsen, A.T.M.; Borra, R.J.H.; Dierckx, R.A.J.O.; Lammertsma, A.A. Long axial field of view PET scanners: A road map to implementation and new possibilities. Eur. J. Nucl. Med. Mol. Imaging 2021, 48, 4236–4245. [Google Scholar] [CrossRef]
- Surti, S.; Pantel, A.R.; Karp, J.S. Total Body PET: Why, How, What for? IEEE Trans. Radiat. Plasma Med. Sci. 2020, 4, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Miller-Thomas, M.M.; Benzinger, T.L. Neurologic Applications of PET/MR Imaging. Magn. Reson. Imaging Clin. N. Am. 2017, 25, 297–313. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Strategy | Mechanism | References |
|---|---|---|
| Physicochemical properties modification | Poly(ethylene glycol) conjugation to increase the circulation half-life | [12,13] |
| Through physiological mechanism | Cationic proteins or nanobodies that trigger adsorption-mediated transcytosis (AMT) | [14,15,16] |
| Ligands or antibodies that trigger receptor-mediated transcytosis (RMT) | [17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32] | |
| BBB permeability changes | Open BBB with solvents such as mannitol | [19,33] |
| Focused ultrasound (FUS) with microbubbles | [34,35,36,37,38] |
| Radionuclide | Half-Life | Branching Ratio (Β+) (%) | Positron Energy–E Max [Mev] | Mean Positron Range (mm) |
|---|---|---|---|---|
| 11C | 20.4 min | 99 | 0.97 | 1.2 |
| 18F | 109.7 min | 97 | 0.65 | 0.6 |
| 68Ga | 67.7 min | 89 | 1.90 | 3.5 |
| 44Sc | 3.97 h | 94 | 1.47 | 2.3 |
| 64Cu | 12.7 h | 18 | 0.65 | 0.7 |
| 89Zr | 78.4 h | 23 | 0.91 | 1.3 |
| 124I | 100.2 h | 23 | 1.54 | 4.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Lucas, Á.G.; Lamminmäki, U.; López-Picón, F.R. ImmunoPET Directed to the Brain: A New Tool for Preclinical and Clinical Neuroscience. Biomolecules 2023, 13, 164. https://doi.org/10.3390/biom13010164
de Lucas ÁG, Lamminmäki U, López-Picón FR. ImmunoPET Directed to the Brain: A New Tool for Preclinical and Clinical Neuroscience. Biomolecules. 2023; 13(1):164. https://doi.org/10.3390/biom13010164
Chicago/Turabian Stylede Lucas, Ángel García, Urpo Lamminmäki, and Francisco R. López-Picón. 2023. "ImmunoPET Directed to the Brain: A New Tool for Preclinical and Clinical Neuroscience" Biomolecules 13, no. 1: 164. https://doi.org/10.3390/biom13010164