
Palmitoylethanolamide and Related ALIAmides for Small Animal Health: State of the Art


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
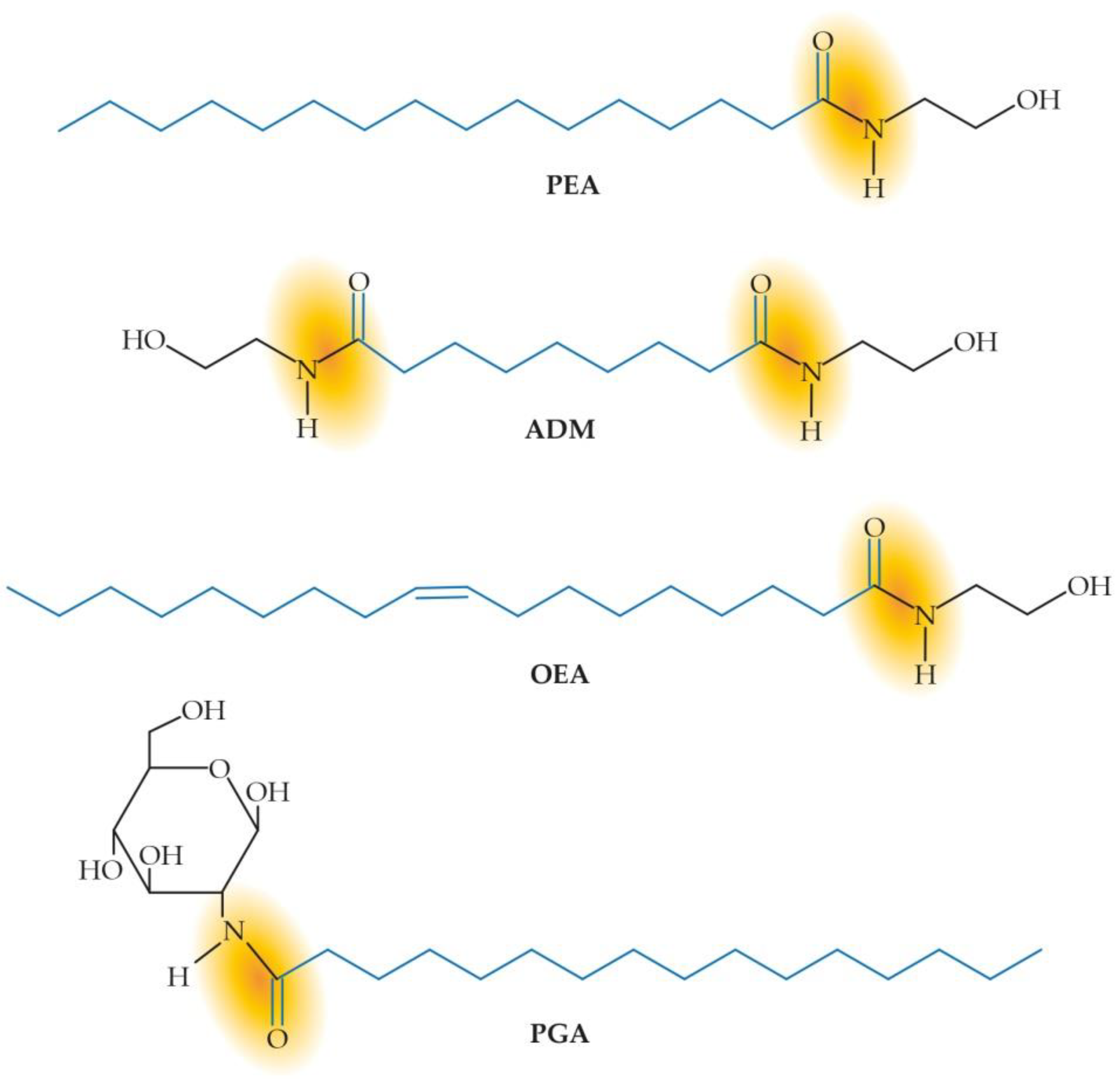
2. Mimicking and Supporting the Healing Power of Nature
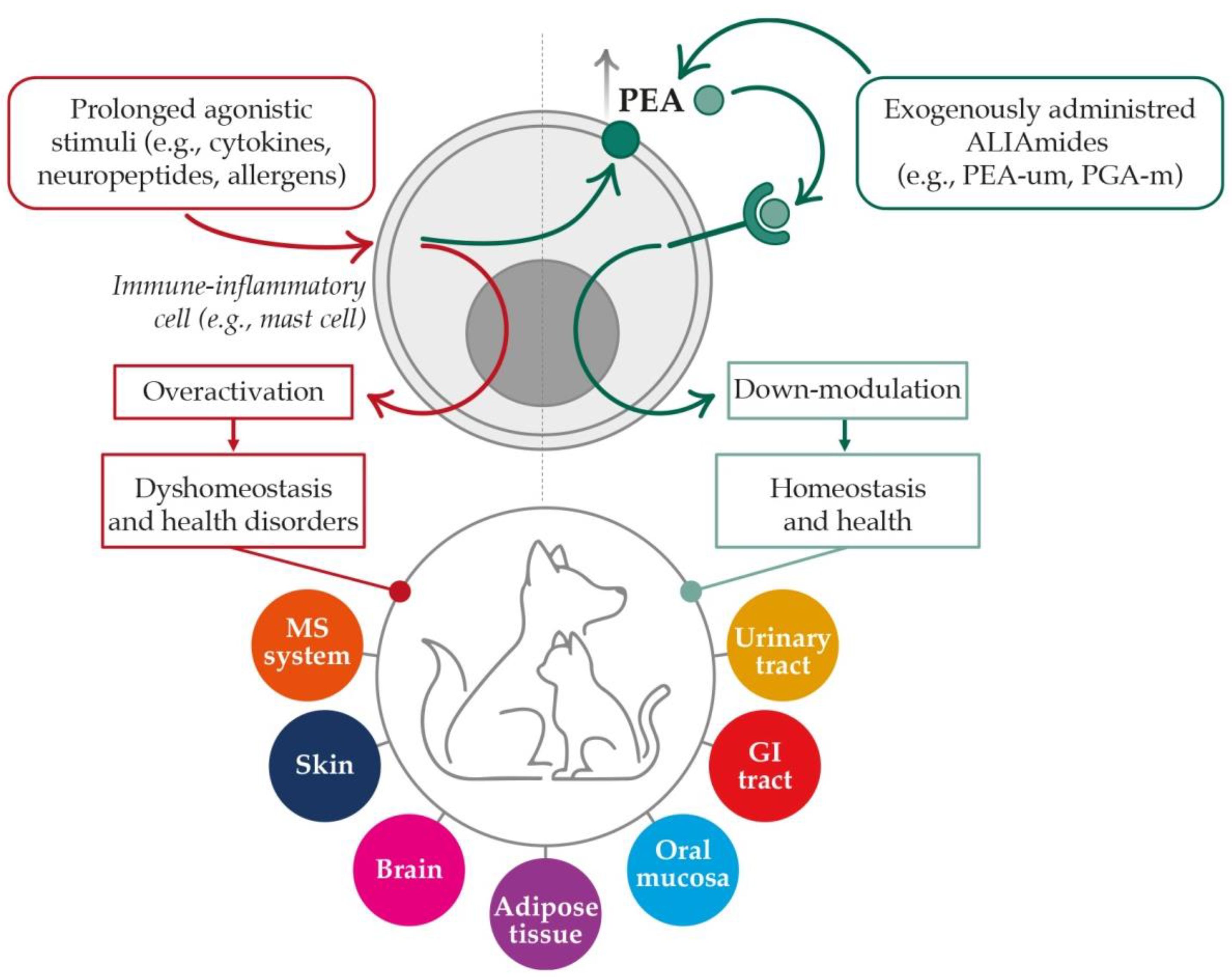
3. A Brief Insight into PEA Metabolism and Molecular Targets
4. Key Pharmacokinetic Issues
5. Preclinical and Clinical Results in Small Animals
5.1. Dermatological Field
5.2. Other Health Needs
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aloe, L.; Leon, A.; Levi-Montalcini, R. A Proposed Autacoid Mechanism Controlling Mastocyte Behaviour. Agents Actions 1993, 39, C145–C147. [Google Scholar] [CrossRef] [PubMed]
- Melmon, K.L.; Rocklin, R.E.; Rosenkranz, R.P. Autacoids as Modulators of the Inflammatory and Immune Response. Am. J. Med. 1981, 71, 100–106. [Google Scholar] [CrossRef]
- Gugliandolo, E.; Peritore, A.F.; Piras, C.; Cuzzocrea, S.; Crupi, R. Palmitoylethanolamide and Related ALIAmides: Prohomeostatic Lipid Compounds for Animal Health and Wellbeing. Vet. Sci. 2020, 7, 78. [Google Scholar] [CrossRef] [PubMed]
- Mazzari, S.; Canella, R.; Petrelli, L.; Marcolongo, G.; Leon, A. N-(2-Hydroxyethyl)Hexadecanamide Is Orally Active in Reducing Edema Formation and Inflammatory Hyperalgesia by down-Modulating Mast Cell Activation. Eur. J. Pharmacol. 1996, 300, 227–236. [Google Scholar] [CrossRef]
- Cerrato, S.; Brazis, P.; della Valle, M.F.; Miolo, A.; Puigdemont, A. Effects of Palmitoylethanolamide on Immunologically Induced Histamine, PGD2 and TNFalpha Release from Canine Skin Mast Cells. Vet. Immunol. Immunopathol. 2010, 133, 9–15. [Google Scholar] [CrossRef]
- Abramo, F.; Lazzarini, G.; Pirone, A.; Lenzi, C.; Albertini, S.; Della Valle, M.F.; Schievano, C.; Vannozzi, I.; Miragliotta, V. Ultramicronized Palmitoylethanolamide Counteracts the Effects of Compound 48/80 in a Canine Skin Organ Culture Model. Vet. Dermatol. 2017, 28, 456-e104. [Google Scholar] [CrossRef]
- Scarampella, F.; Abramo, F.; Noli, C. Clinical and Histological Evaluation of an Analogue of Palmitoylethanolamide, PLR 120 (Comicronized Palmidrol INN) in Cats with Eosinophilic Granuloma and Eosinophilic Plaque: A Pilot Study. Vet. Dermatol. 2001, 12, 29–39. [Google Scholar] [CrossRef]
- Rinne, P.; Guillamat-Prats, R.; Rami, M.; Bindila, L.; Ring, L.; Lyytikäinen, L.-P.; Raitoharju, E.; Oksala, N.; Lehtimäki, T.; Weber, C.; et al. Palmitoylethanolamide Promotes a Proresolving Macrophage Phenotype and Attenuates Atherosclerotic Plaque Formation. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 2562–2575. [Google Scholar] [CrossRef]
- Petrosino, S.; Cristino, L.; Karsak, M.; Gaffal, E.; Ueda, N.; Tüting, T.; Bisogno, T.; De Filippis, D.; D’Amico, A.; Saturnino, C.; et al. Protective Role of Palmitoylethanolamide in Contact Allergic Dermatitis. Allergy 2010, 65, 698–711. [Google Scholar] [CrossRef]
- Bettoni, I.; Comelli, F.; Colombo, A.; Bonfanti, P.; Costa, B. Non-Neuronal Cell Modulation Relieves Neuropathic Pain: Efficacy of the Endogenous Lipid Palmitoylethanolamide. CNS Neurol. Disord. Drug Targets 2013, 12, 34–44. [Google Scholar] [CrossRef]
- Luongo, L.; Guida, F.; Boccella, S.; Bellini, G.; Gatta, L.; Rossi, F.; de Novellis, V.; Maione, S. Palmitoylethanolamide Reduces Formalin-Induced Neuropathic-like Behaviour through Spinal Glial/Microglial Phenotypical Changes in Mice. CNS Neurol. Disord. Drug Targets 2013, 12, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Guida, F.; Luongo, L.; Marmo, F.; Romano, R.; Iannotta, M.; Napolitano, F.; Belardo, C.; Marabese, I.; D’Aniello, A.; De Gregorio, D.; et al. Palmitoylethanolamide Reduces Pain-Related Behaviors and Restores Glutamatergic Synapses Homeostasis in the Medial Prefrontal Cortex of Neuropathic Mice. Mol. Brain 2015, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Gabrielsson, L.; Gouveia-Figueira, S.; Häggström, J.; Alhouayek, M.; Fowler, C.J. The Anti-Inflammatory Compound Palmitoylethanolamide Inhibits Prostaglandin and Hydroxyeicosatetraenoic Acid Production by a Macrophage Cell Line. Pharmacol. Res. Perspect. 2017, 5, e00300. [Google Scholar] [CrossRef] [PubMed]
- Chiurchiù, V.; Leuti, A.; Smoum, R.; Mechoulam, R.; Maccarrone, M. Bioactive Lipids ALIAmides Differentially Modulate Inflammatory Responses of Distinct Subsets of Primary Human T Lymphocytes. FASEB J. 2018, 32, 5716–5723. [Google Scholar] [CrossRef]
- Bronzuoli, M.R.; Facchinetti, R.; Steardo, L.; Romano, A.; Stecca, C.; Passarella, S.; Steardo, L.; Cassano, T.; Scuderi, C. Palmitoylethanolamide Dampens Reactive Astrogliosis and Improves Neuronal Trophic Support in a Triple Transgenic Model of Alzheimer’s Disease: In Vitro and In Vivo Evidence. Oxidative Med. Cell. Longev. 2018, 2018, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Scuderi, C.; Esposito, G.; Blasio, A.; Valenza, M.; Arietti, P.; Steardo, L.; Carnuccio, R.; De Filippis, D.; Petrosino, S.; Iuvone, T.; et al. Palmitoylethanolamide Counteracts Reactive Astrogliosis Induced by β-Amyloid Peptide. J. Cell Mol. Med. 2011, 15, 2664–2674. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, T.; Kamiyama, N.; Saechue, B.; Soga, Y.; Gotoh, R.; Nakayama, T.; Fukuda, C.; Dewayani, A.; Chalalai, T.; Ariki, S.; et al. Comprehensive Lipidomics of Lupus-Prone Mice Using LC-MS/MS Identifies the Reduction of Palmitoylethanolamide That Suppresses TLR9-Mediated Inflammation. Genes Cells 2022, 27, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Facchinetti, R.; Valenza, M.; Gomiero, C.; Mancini, G.F.; Steardo, L.; Campolongo, P.; Scuderi, C. Co-Ultramicronized Palmitoylethanolamide/Luteolin Restores Oligodendrocyte Homeostasis via Peroxisome Proliferator-Activated Receptor-α in an In Vitro Model of Alzheimer’s Disease. Biomedicines 2022, 10, 1236. [Google Scholar] [CrossRef]
- Bisogno, T.; Maurelli, S.; Melck, D.; De Petrocellis, L.; Di Marzo, V. Biosynthesis, Uptake, and Degradation of Anandamide and Palmitoylethanolamide in Leukocytes. J. Biol. Chem. 1997, 272, 3315–3323. [Google Scholar] [CrossRef]
- Muccioli, G.G.; Stella, N. Microglia Produce and Hydrolyze Palmitoylethanolamide. Neuropharmacology 2008, 54, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Walter, L.; Franklin, A.; Witting, A.; Moller, T.; Stella, N. Astrocytes in Culture Produce Anandamide and Other Acylethanolamides. J. Biol. Chem. 2002, 277, 20869–20876. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, V.; Reddy, P.V.; Schmid, P.C.; Schmid, H.H. On the Biosynthesis and Metabolism of N-Acylethanolamine Phospholipids in Infarcted Dog Heart. Biochim. Biophys. Acta 1981, 664, 445–448. [Google Scholar] [CrossRef]
- Epps, D.E.; Schmid, P.C.; Natarajan, V.; Schmid, H.H.O. N-Acylethanolamine Accumulation in Infarcted Myocardium. Biochem. Biophys. Res. Commun. 1979, 90, 628–633. [Google Scholar] [CrossRef]
- Natarajan, V.; Schmid, P.C.; Reddy, P.V.; Schmid, H.H. Catabolism of N-Acylethanolamine Phospholipids by Dog Brain Preparations. J. Neurochem. 1984, 42, 1613–1619. [Google Scholar] [CrossRef] [PubMed]
- Devane, W.A.; Hanus, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and Structure of a Brain Constituent That Binds to the Cannabinoid Receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef]
- della Valle, F.; Gambardella, A. ‘Biological’ Revolution and Strategies for Innovation in Pharmaceutical Companies. R&D Manag. 1993, 23, 287–302. [Google Scholar] [CrossRef]
- Aloe, L. Rita Levi-Montalcini: The Discovery of Nerve Growth Factor and Modern Neurobiology. Trends Cell. Biol. 2004, 14, 395–399. [Google Scholar] [CrossRef]
- Grayson, D.R.; Guidotti, A. Erminio Costa. Neuropsychopharmacology 2010, 35, 2646. [Google Scholar] [CrossRef]
- Costa, E. An Early Attempt to Foster Neuroscience Globalization: An Autobiography; Good Life Press, Incorporated: Singapore, 2003; ISBN 978-0-9726121-1-1. [Google Scholar]
- Levi-Montalcini, R.; Skaper, S.D.; Dal Toso, R.; Petrelli, L.; Leon, A. Nerve Growth Factor: From Neurotrophin to Neurokine. Trends Neurosci. 1996, 19, 514–520. [Google Scholar] [CrossRef]
- Costa, E. Rrespecting Nature as a Strategy for the Development of New Drugs of the Nervous System. In Proceedings of the Drugs for Human Life Conference, Rome, Italy, 23–25 October 1986. [Google Scholar]
- della Rocca, G.; Gamba, D. Chronic Pain in Dogs and Cats: Is There Place for Dietary Intervention with Micro-Palmitoylethanolamide? Animals 2021, 11, 952. [Google Scholar] [CrossRef]
- Re, G.; Barbero, R.; Miolo, A.; Di Marzo, V. Palmitoylethanolamide, Endocannabinoids and Related Cannabimimetic Compounds in Protection against Tissue Inflammation and Pain: Potential Use in Companion Animals. Vet. J. 2007, 173, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Petrosino, S.; Schiano Moriello, A. Palmitoylethanolamide: A Nutritional Approach to Keep Neuroinflammation within Physiological Boundaries-A Systematic Review. Int. J. Mol. Sci. 2020, 21, 9526. [Google Scholar] [CrossRef] [PubMed]
- Petrosino, S.; Di Marzo, V. The Pharmacology of Palmitoylethanolamide and First Data on the Therapeutic Efficacy of Some of Its New Formulations. Br. J. Pharmacol. 2017, 174, 1349–1365. [Google Scholar] [CrossRef] [PubMed]
- Esposito, E.; Cuzzocrea, S. Palmitoylethanolamide Is a New Possible Pharmacological Treatment for the Inflammation Associated with Trauma. Mini Rev. Med. Chem. 2013, 13, 237–255. [Google Scholar] [PubMed]
- Scuderi, C.; Golini, L. Successful and Unsuccessful Brain Aging in Pets: Pathophysiological Mechanisms behind Clinical Signs and Potential Benefits from Palmitoylethanolamide Nutritional Intervention. Animals 2021, 11, 2584. [Google Scholar] [CrossRef]
- Hussain, Z.; Uyama, T.; Tsuboi, K.; Ueda, N. Mammalian Enzymes Responsible for the Biosynthesis of N-Acylethanolamines. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 1546–1561. [Google Scholar] [CrossRef]
- Tsuboi, K.; Uyama, T.; Okamoto, Y.; Ueda, N. Endocannabinoids and Related N-Acylethanolamines: Biological Activities and Metabolism. Inflamm. Regen. 2018, 38, 28. [Google Scholar] [CrossRef]
- Sun, Y.-X.; Tsuboi, K.; Okamoto, Y.; Tonai, T.; Murakami, M.; Kudo, I.; Ueda, N. Biosynthesis of Anandamide and N-Palmitoylethanolamine by Sequential Actions of Phospholipase A2 and Lysophospholipase D. Biochem. J. 2004, 380, 749–756. [Google Scholar] [CrossRef]
- Ueda, N.; Tsuboi, K.; Uyama, T. N-Acylethanolamine Metabolism with Special Reference to N-Acylethanolamine-Hydrolyzing Acid Amidase (NAAA). Prog. Lipid Res. 2010, 49, 299–315. [Google Scholar] [CrossRef]
- Alhouayek, M.; Muccioli, G.G. Harnessing the Anti-Inflammatory Potential of Palmitoylethanolamide. Drug Discov. Today 2014, 19, 1632–1639. [Google Scholar] [CrossRef]
- Roviezzo, F.; Rossi, A.; Caiazzo, E.; Orlando, P.; Riemma, M.A.; Iacono, V.M.; Guarino, A.; Ialenti, A.; Cicala, C.; Peritore, A.; et al. Palmitoylethanolamide Supplementation during Sensitization Prevents Airway Allergic Symptoms in the Mouse. Front. Pharmacol. 2017, 8, 857. [Google Scholar] [CrossRef] [PubMed]
- Skaper, S.D.; Facci, L.; Barbierato, M.; Zusso, M.; Bruschetta, G.; Impellizzeri, D.; Cuzzocrea, S.; Giusti, P. N-Palmitoylethanolamine and Neuroinflammation: A Novel Therapeutic Strategy of Resolution. Mol. Neurobiol. 2015, 52, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Petrosino, S.; Palazzo, E.; de Novellis, V.; Bisogno, T.; Rossi, F.; Maione, S.; Di Marzo, V. Changes in Spinal and Supraspinal Endocannabinoid Levels in Neuropathic Rats. Neuropharmacology 2007, 52, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Charrua, A.; Matos, R.; Oliveira, R.; Marczylo, T.; Nagy, I.; Cruz, F. Fatty Acid Amide Hydrolase Inhibition Normalises Bladder Function and Reduces Pain through Normalising the Anandamide/Palmitoylethanolamine Ratio in the Inflamed Bladder of Rats. Naunyn. Schmiedebergs Arch. Pharmacol. 2020, 393, 263–272. [Google Scholar] [CrossRef]
- Zhou, P.; Xiang, L.; Yang, Y.; Wu, Y.; Hu, T.; Liu, X.; Lin, F.; Xiu, Y.; Wu, K.; Lu, C.; et al. N-Acylethanolamine Acid Amidase (NAAA) Inhibitor F215 as a Novel Therapeutic Agent for Osteoarthritis. Pharmacol. Res. 2019, 145, 104264. [Google Scholar] [CrossRef]
- Fichna, J.; Wood, J.T.; Papanastasiou, M.; Vadivel, S.K.; Oprocha, P.; Sałaga, M.; Sobczak, M.; Mokrowiecka, A.; Cygankiewicz, A.I.; Zakrzewski, P.K.; et al. Endocannabinoid and Cannabinoid-like Fatty Acid Amide Levels Correlate with Pain-Related Symptoms in Patients with IBS-D and IBS-C: A Pilot Study. PLoS ONE 2013, 8, e85073. [Google Scholar] [CrossRef]
- Sarnelli, G.; Pesce, M.; Seguella, L.; Lu, J.; Efficie, E.; Tack, J.; Elisa De Palma, F.D.; D’Alessandro, A.; Esposito, G. Impaired Duodenal Palmitoylethanolamide Release Underlies Acid-Induced Mast Cell Activation in Functional Dyspepsia. Cell Mol. Gastroenterol. Hepatol. 2021, 11, 841–855. [Google Scholar] [CrossRef]
- Richardson, D.; Pearson, R.G.; Kurian, N.; Latif, M.L.; Garle, M.J.; Barrett, D.A.; Kendall, D.A.; Scammell, B.E.; Reeve, A.J.; Chapman, V. Characterisation of the Cannabinoid Receptor System in Synovial Tissue and Fluid in Patients with Osteoarthritis and Rheumatoid Arthritis. Arthritis Res. Ther. 2008, 10, R43. [Google Scholar] [CrossRef]
- Berdyshev, E.V.; Schmid, P.C.; Dong, Z.; Schmid, H.H. Stress-Induced Generation of N-Acylethanolamines in Mouse Epidermal JB6 P+ Cells. Biochem. J. 2000, 346 Pt 2, 369–374. [Google Scholar] [CrossRef]
- Abramo, F.; Campora, L.; Albanese, F.; della Valle, M.F.; Cristino, L.; Petrosino, S.; Di Marzo, V.; Miragliotta, V. Increased Levels of Palmitoylethanolamide and Other Bioactive Lipid Mediators and Enhanced Local Mast Cell Proliferation in Canine Atopic Dermatitis. BMC Vet. Res. 2014, 10, 21. [Google Scholar] [CrossRef] [Green Version]
- Pengo, G.; Miolo, A. Utilizzo Di Palmitoiletanolamide Micronizzata Nell’infiammazione Gastrointestinale Idiopatica (IBD) Del Cane: Descrizione Di 7 Casi Clinici. In Proceedings of the 72 International SCIVAC Congress, Milan, Italy, 23–25 March 2012; pp. 299–301. [Google Scholar]
- Skaper, S.D.; Facci, L.; Giusti, P. Mast Cells, Glia and Neuroinflammation: Partners in Crime? Immunology 2014, 141, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Del Re, A.; Palenca, I.; Seguella, L.; Pesce, M.; Corpetti, C.; Steardo, L.; Rurgo, S.; Sarnelli, G.; Esposito, G. Oral Adelmidrol Administration Up-Regulates Palmitoylethanolamide Production in Mice Colon and Duodenum through a PPAR-γ Independent Action. Metabolites 2022, 12, 457. [Google Scholar] [CrossRef] [PubMed]
- Petrosino, S.; Puigdemont, A.; Della Valle, M.F.; Fusco, M.; Verde, R.; Allarà, M.; Aveta, T.; Orlando, P.; Di Marzo, V. Adelmidrol Increases the Endogenous Concentrations of Palmitoylethanolamide in Canine Keratinocytes and Down-Regulates an Inflammatory Reaction in an in Vitro Model of Contact Allergic Dermatitis. Vet. J. 2016, 207, 85–91. [Google Scholar] [CrossRef] [PubMed]
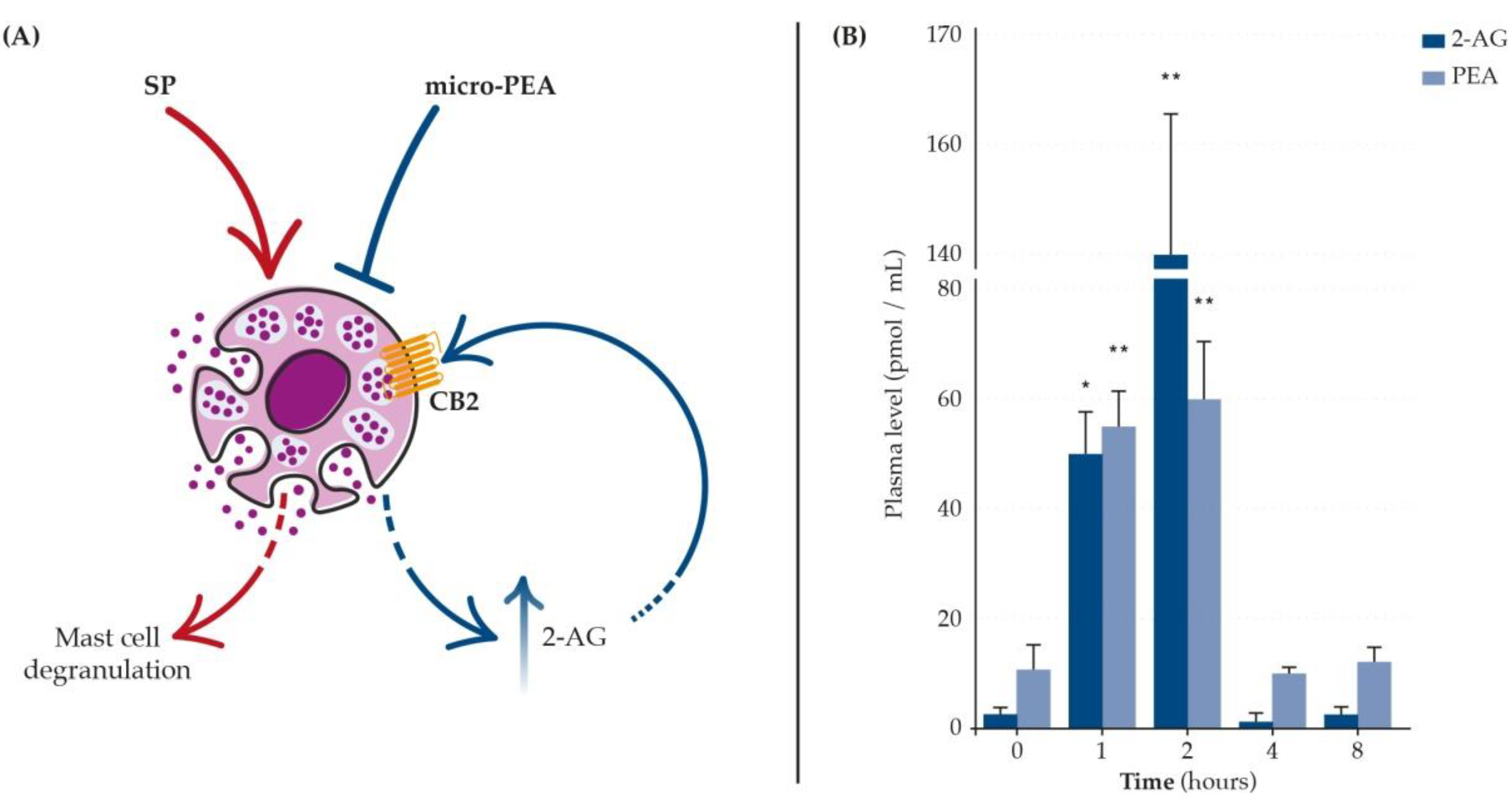
- Petrosino, S.; Schiano Moriello, A.; Verde, R.; Allarà, M.; Imperatore, R.; Ligresti, A.; Mahmoud, A.M.; Peritore, A.F.; Iannotti, F.A.; Di Marzo, V. Palmitoylethanolamide Counteracts Substance P-Induced Mast Cell Activation in Vitro by Stimulating Diacylglycerol Lipase Activity. J. Neuroinflammation 2019, 16, 274. [Google Scholar] [CrossRef] [PubMed]
- Jacobsson, S.O.; Fowler, C.J. Characterization of Palmitoylethanolamide Transport in Mouse Neuro-2a Neuroblastoma and Rat RBL-2H3 Basophilic Leukaemia Cells: Comparison with Anandamide. Br. J. Pharmacol. 2001, 132, 1743–1754. [Google Scholar] [CrossRef] [PubMed]
- Carta, G.; Murru, E.; Banni, S.; Manca, C. Palmitic Acid: Physiological Role, Metabolism and Nutritional Implications. Front. Physiol. 2017, 8, 902. [Google Scholar] [CrossRef]
- Bojesen, I.N.; Hansen, H.S. Membrane Transport of Anandamide through Resealed Human Red Blood Cell Membranes. J. Lipid Res. 2005, 46, 1652–1659. [Google Scholar] [CrossRef]
- Kaczocha, M.; Glaser, S.T.; Maher, T.; Clavin, B.; Hamilton, J.; O’Rourke, J.; Rebecchi, M.; Puopolo, M.; Owada, Y.; Thanos, P.K. Fatty Acid Binding Protein Deletion Suppresses Inflammatory Pain through Endocannabinoid/N-Acylethanolamine-Dependent Mechanisms. Mol. Pain 2015, 11, 52. [Google Scholar] [CrossRef]
- Kaczocha, M.; Vivieca, S.; Sun, J.; Glaser, S.T.; Deutsch, D.G. Fatty Acid-Binding Proteins Transport N-Acylethanolamines to Nuclear Receptors and Are Targets of Endocannabinoid Transport Inhibitors. J. Biol. Chem. 2012, 287, 3415–3424. [Google Scholar] [CrossRef]
- Oddi, S.; Fezza, F.; Pasquariello, N.; D’Agostino, A.; Catanzaro, G.; De Simone, C.; Rapino, C.; Finazzi-Agrò, A.; Maccarrone, M. Molecular Identification of Albumin and Hsp70 as Cytosolic Anandamide-Binding Proteins. Chem. Biol. 2009, 16, 624–632. [Google Scholar] [CrossRef]
- Korbecki, J.; Bobiński, R.; Dutka, M. Self-Regulation of the Inflammatory Response by Peroxisome Proliferator-Activated Receptors. Inflamm. Res. 2019, 68, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Lo Verme, J.; Fu, J.; Astarita, G.; La Rana, G.; Russo, R.; Calignano, A.; Piomelli, D. The Nuclear Receptor Peroxisome Proliferator-Activated Receptor-Alpha Mediates the Anti-Inflammatory Actions of Palmitoylethanolamide. Mol. Pharmacol. 2005, 67, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Lo Verme, J.; La Rana, G.; Russo, R.; Calignano, A.; Piomelli, D. The Search for the Palmitoylethanolamide Receptor. Life Sci. 2005, 77, 1685–1698. [Google Scholar] [CrossRef] [PubMed]
- LoVerme, J.; Russo, R.; La Rana, G.; Fu, J.; Farthing, J.; Mattace-Raso, G.; Meli, R.; Hohmann, A.; Calignano, A.; Piomelli, D. Rapid Broad-Spectrum Analgesia through Activation of Peroxisome Proliferator-Activated Receptor-Alpha. J. Pharmacol. Exp. Ther. 2006, 319, 1051–1061. [Google Scholar] [CrossRef]
- Zhou, G.; Fu, X.; Wang, L.; Cao, Y.; Zhuang, J.; Hu, J.; Li, Y.; Xu, C.; Gao, S.; Shao, A.; et al. Palmitoylethanolamide Ameliorates Neuroinflammation via Modulating PPAR-α to Promote the Functional Outcome after Intracerebral Hemorrhage. Neurosci. Lett. 2022, 781, 136648. [Google Scholar] [CrossRef]
- Annunziata, C.; Pirozzi, C.; Lama, A.; Senzacqua, M.; Comella, F.; Bordin, A.; Monnolo, A.; Pelagalli, A.; Ferrante, M.C.; Mollica, M.P.; et al. Palmitoylethanolamide Promotes White-to-Beige Conversion and Metabolic Reprogramming of Adipocytes: Contribution of PPAR-α. Pharmaceutics 2022, 14, 338. [Google Scholar] [CrossRef]
- Im, D.-S. GPR119 and GPR55 as Receptors for Fatty Acid Ethanolamides, Oleoylethanolamide and Palmitoylethanolamide. Int. J. Mol. Sci. 2021, 22, 1034. [Google Scholar] [CrossRef]
- Ryberg, E.; Larsson, N.; Sjögren, S.; Hjorth, S.; Hermansson, N.-O.; Leonova, J.; Elebring, T.; Nilsson, K.; Drmota, T.; Greasley, P.J. The Orphan Receptor GPR55 Is a Novel Cannabinoid Receptor. Br. J. Pharmacol. 2007, 152, 1092–1101. [Google Scholar] [CrossRef]
- Petrosino, S.; Schiano Moriello, A.; Cerrato, S.; Fusco, M.; Puigdemont, A.; De Petrocellis, L.; Di Marzo, V. The Anti-Inflammatory Mediator Palmitoylethanolamide Enhances the Levels of 2-Arachidonoyl-Glycerol and Potentiates Its Actions at TRPV1 Cation Channels. Br. J. Pharmacol. 2016, 173, 1154–1162. [Google Scholar] [CrossRef]
- Di Marzo, V.; Melck, D.; Orlando, P.; Bisogno, T.; Zagoory, O.; Bifulco, M.; Vogel, Z.; De Petrocellis, L. Palmitoylethanolamide Inhibits the Expression of Fatty Acid Amide Hydrolase and Enhances the Anti-Proliferative Effect of Anandamide in Human Breast Cancer Cells. Biochem. J. 2001, 358, 249–255. [Google Scholar] [CrossRef]
- Julius, D. TRP Channels and Pain. Annu. Rev. Cell Dev. Biol. 2013, 29, 355–384. [Google Scholar] [CrossRef] [PubMed]
- Costa, B.; Comelli, F.; Bettoni, I.; Colleoni, M.; Giagnoni, G. The Endogenous Fatty Acid Amide, Palmitoylethanolamide, Has Anti-Allodynic and Anti-Hyperalgesic Effects in a Murine Model of Neuropathic Pain: Involvement of CB(1), TRPV1 and PPARgamma Receptors and Neurotrophic Factors. Pain 2008, 139, 541–550. [Google Scholar] [CrossRef] [PubMed]
- De Petrocellis, L.; Davis, J.B.; Di Marzo, V. Palmitoylethanolamide Enhances Anandamide Stimulation of Human Vanilloid VR1 Receptors. FEBS Lett. 2001, 506, 253–256. [Google Scholar] [CrossRef]
- Ambrosino, P.; Soldovieri, M.V.; Russo, C.; Taglialatela, M. Activation and Desensitization of TRPV1 Channels in Sensory Neurons by the PPARα Agonist Palmitoylethanolamide. Br. J. Pharmacol. 2013, 168, 1430–1444. [Google Scholar] [CrossRef] [PubMed]
- Ho, W.-S.V.; Barrett, D.A.; Randall, M.D. “Entourage” Effects of N-Palmitoylethanolamide and N-Oleoylethanolamide on Vasorelaxation to Anandamide Occur through TRPV1 Receptors. Br. J. Pharmacol. 2008, 155, 837–846. [Google Scholar] [CrossRef]
- Campora, L.; Miragliotta, V.; Ricci, E.; Cristino, L.; Di Marzo, V.; Albanese, F.; Federica Della Valle, M.; Abramo, F. Cannabinoid Receptor Type 1 and 2 Expression in the Skin of Healthy Dogs and Dogs with Atopic Dermatitis. Am. J. Vet. Res. 2012, 73, 988–995. [Google Scholar] [CrossRef]
- Barbero, R.; Vercelli, C.; Cuniberti, B.; Della Valle, M.F.; Martano, M.; Re, G. Expression of Functional TRPV1 Receptor in Primary Culture of Canine Keratinocytes. J. Vet. Pharmacol. Ther. 2018, 41, 795–804. [Google Scholar] [CrossRef]
- Dall’Aglio, C.; Mercati, F.; Pascucci, L.; Boiti, C.; Pedini, V.; Ceccarelli, P. Immunohistochemical Localization of CB1 Receptor in Canine Salivary Glands. Vet. Res. Commun 2010, 34 (Suppl. S1), S9–S12. [Google Scholar] [CrossRef]
- Mercati, F.; Dall’Aglio, C.; Pascucci, L.; Boiti, C.; Ceccarelli, P. Identification of Cannabinoid Type 1 Receptor in Dog Hair Follicles. Acta Histochem. 2012, 114, 68–71. [Google Scholar] [CrossRef]
- Fernández-Trapero, M.; Espejo-Porras, F.; Rodríguez-Cueto, C.; Coates, J.R.; Pérez-Díaz, C.; de Lago, E.; Fernández-Ruiz, J. Upregulation of CB2 Receptors in Reactive Astrocytes in Canine Degenerative Myelopathy, a Disease Model of Amyotrophic Lateral Sclerosis. Dis. Model. Mech. 2017, 10, 551–558. [Google Scholar] [CrossRef] [Green Version]
- Freundt-Revilla, J.; Kegler, K.; Baumgärtner, W.; Tipold, A. Spatial Distribution of Cannabinoid Receptor Type 1 (CB1) in Normal Canine Central and Peripheral Nervous System. PLoS ONE 2017, 12, e0181064. [Google Scholar] [CrossRef]
- Freundt-Revilla, J.; Heinrich, F.; Zoerner, A.; Gesell, F.; Beyerbach, M.; Shamir, M.; Oevermann, A.; Baumgärtner, W.; Tipold, A. The Endocannabinoid System in Canine Steroid-Responsive Meningitis-Arteritis and Intraspinal Spirocercosis. PLoS ONE 2018, 13, e0187197. [Google Scholar] [CrossRef]
- Galiazzo, G.; Giancola, F.; Stanzani, A.; Fracassi, F.; Bernardini, C.; Forni, M.; Pietra, M.; Chiocchetti, R. Localization of Cannabinoid Receptors CB1, CB2, GPR55, and PPARα in the Canine Gastrointestinal Tract. Histochem. Cell Biol. 2018, 150, 187–205. [Google Scholar] [CrossRef] [PubMed]
- Stanzani, A.; Galiazzo, G.; Giancola, F.; Tagliavia, C.; De Silva, M.; Pietra, M.; Fracassi, F.; Chiocchetti, R. Localization of Cannabinoid and Cannabinoid Related Receptors in the Cat Gastrointestinal Tract. Histochem. Cell Biol. 2020, 153, 339–356. [Google Scholar] [CrossRef]
- Gebremedhin, D.; Lange, A.R.; Campbell, W.B.; Hillard, C.J.; Harder, D.R. Cannabinoid CB1 Receptor of Cat Cerebral Arterial Muscle Functions to Inhibit L-Type Ca2+ Channel Current. Am. J. Physiol. 1999, 276, H2085–H2093. [Google Scholar] [CrossRef]
- Miragliotta, V.; Ricci, P.L.; Albanese, F.; Pirone, A.; Tognotti, D.; Abramo, F. Cannabinoid Receptor Types 1 and 2 and Peroxisome Proliferator-Activated Receptor-α: Distribution in the Skin of Clinically Healthy Cats and Cats with Hypersensitivity Dermatitis. Vet. Dermatol. Online ahead of print. 2018. [Google Scholar] [CrossRef]
- Pirone, A.; Cantile, C.; Miragliotta, V.; Lenzi, C.; Giannessi, E.; Cozzi, B. Immunohistochemical Distribution of the Cannabinoid Receptor 1 and Fatty Acid Amide Hydrolase in the Dog Claustrum. J. Chem. Neuroanat. 2016, 74, 21–27. [Google Scholar] [CrossRef]
- Pirone, A.; Lenzi, C.; Briganti, A.; Abbate, F.; Levanti, M.; Abramo, F.; Miragliotta, V. Spatial Distribution of Cannabinoid Receptor 1 and Fatty Acid Amide Hydrolase in the Cat Ovary and Oviduct. Acta Histochem. 2017, 119, 417–422. [Google Scholar] [CrossRef]
- Ndong, C.; O’Donnell, D.; Ahmad, S.; Groblewski, T. Cloning and Pharmacological Characterization of the Dog Cannabinoid CB2 receptor. Eur. J. Pharmacol. 2011, 669, 24–31. [Google Scholar] [CrossRef]
- Silver, R.J. The Endocannabinoid System of Animals. Animals 2019, 9, 686. [Google Scholar] [CrossRef] [Green Version]
- Iannotta, M.; Belardo, C.; Trotta, M.C.; Iannotti, F.A.; Vitale, R.M.; Maisto, R.; Boccella, S.; Infantino, R.; Ricciardi, F.; Mirto, B.F.; et al. N-Palmitoyl-D-Glucosamine, a Natural Monosaccharide-Based Glycolipid, Inhibits TLR4 and Prevents LPS-Induced Inflammation and Neuropathic Pain in Mice. Int. J. Mol. Sci. 2021, 22, 1491. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, D.; Di Paola, R.; Cordaro, M.; Gugliandolo, E.; Casili, G.; Morittu, V.M.; Britti, D.; Esposito, E.; Cuzzocrea, S. Adelmidrol, a Palmitoylethanolamide Analogue, as a New Pharmacological Treatment for the Management of Acute and Chronic Inflammation. Biochem. Pharmacol. 2016, 119, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Cerrato, S.; Brazis, P.; Della Valle, M.F.; Miolo, A.; Puigdemont, A. Inhibitory Effect of Topical Adelmidrol on Antigen-Induced Skin Wheal and Mast Cell Behavior in a Canine Model of Allergic Dermatitis. BMC Vet. Res. 2012, 8, 230. [Google Scholar] [CrossRef] [PubMed]
- Cordaro, M.; Siracusa, R.; Impellizzeri, D.; D’ Amico, R.; Peritore, A.F.; Crupi, R.; Gugliandolo, E.; Fusco, R.; Di Paola, R.; Schievano, C.; et al. Safety and Efficacy of a New Micronized Formulation of the ALIAmide Palmitoylglucosamine in Preclinical Models of Inflammation and Osteoarthritis Pain. Arthritis Res. Ther. 2019, 21, 254. [Google Scholar] [CrossRef]
- Petrosino, S.; Cordaro, M.; Verde, R.; Schiano Moriello, A.; Marcolongo, G.; Schievano, C.; Siracusa, R.; Piscitelli, F.; Peritore, A.F.; Crupi, R.; et al. Oral Ultramicronized Palmitoylethanolamide: Plasma and Tissue Levels and Spinal Anti-Hyperalgesic Effect. Front. Pharmacol. 2018, 9, 249. [Google Scholar] [CrossRef] [PubMed]
- Bilia, A.R.; Piazzini, V.; Guccione, C.; Risaliti, L.; Asprea, M.; Capecchi, G.; Bergonzi, M.C. Improving on Nature: The Role of Nanomedicine in the Development of Clinical Natural Drugs. Planta Med. 2017, 83, 366–381. [Google Scholar] [CrossRef]
- Takano, R.; Furumoto, K.; Shiraki, K.; Takata, N.; Hayashi, Y.; Aso, Y.; Yamashita, S. Rate-Limiting Steps of Oral Absorption for Poorly Water-Soluble Drugs in Dogs; Prediction from a Miniscale Dissolution Test and a Physiologically-Based Computer Simulation. Pharm. Res. 2008, 25, 2334–2344. [Google Scholar] [CrossRef]
- Dhiman, A.; Prabhakar, P.K. Micronization in Food Processing: A Comprehensive Review of Mechanistic Approach, Physicochemical, Functional Properties and Self-Stability of Micronized Food Materials. J. Food Eng. 2021, 292, 110248. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Bruschetta, G.; Cordaro, M.; Crupi, R.; Siracusa, R.; Esposito, E.; Cuzzocrea, S. Micronized/Ultramicronized Palmitoylethanolamide Displays Superior Oral Efficacy Compared to Nonmicronized Palmitoylethanolamide in a Rat Model of Inflammatory Pain. J. Neuroinflammat. 2014, 11, 136. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Campolo, M.; Di Paola, R.; Bruschetta, G.; de Stefano, D.; Esposito, E.; Cuzzocrea, S. Ultramicronized Palmitoylethanolamide Reduces Inflammation an a Th1-Mediated Model of Colitis. Eur. J. Inflamm. 2015, 13, 14–31. [Google Scholar] [CrossRef] [Green Version]
- Cerrato, S.; Brazis, P.; Della Valle, M.F.; Miolo, A.; Petrosino, S.; Di Marzo, V.; Puigdemont, A. Effects of Palmitoylethanolamide on the Cutaneous Allergic Inflammatory Response in Ascaris Hypersensitive Beagle Dogs. Vet. J. 2012, 191, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Miragliotta, V.; Noli, C. Dermatology: Endocannabinoids and Related N-Acylethanolamines in the Skin. In Cannabis Therapy in Veterinary Medicine: A Complete Guide; Cital, S., Kramer, K., Hughston, L., Gaynor, J.S., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 207–230. ISBN 978-3-030-68317-7. [Google Scholar]
- Abramo, F.; Salluzzi, D.; Leotta, R.; Auxilia, S.; Noli, C.; Miolo, A.; Mantis, P.; Lloyd, D.H. Mast Cell Morphometry and Densitometry in Experimental Skin Wounds Treated with a Gel Containing Adelmidrol: A Placebo Controlled Study. Wounds 2008, 20, 149–157. [Google Scholar] [PubMed]
- Mantis, P.; Lloyd, D.H.; Pfeiffer, D.; Stevens, K.; Auxilia, S.; Noli, C.; Auxilia, S.; Noli, C.; Abramo, F.; Miolo, A. Assessment of the Effect of an Aliamide-Containing Topical Gel by Evaluation of the Reduction of Wound Volume Measured by High R. Wounds 2007, 19, 113–119. [Google Scholar] [PubMed]
- Marsella, R.; Joyce, J.; Nicklin, C.; Lopez, J. Evaluation of the Effects of Palmitoylethanolamide on Clinical Signs in House Dust Mite Allergic High IgE Beagle Dogs Using a Randomized, Double Blinded, Placebo Controlled Design. Vet. Dermatol. 2005, 16, 202. [Google Scholar]
- Noli, C.; Della Valle, M.F.; Miolo, A.; Medori, C.; Schievano, C. Skinalia Clinical Research Group Effect of Dietary Supplementation with Ultramicronized Palmitoylethanolamide in Maintaining Remission in Cats with Nonflea Hypersensitivity Dermatitis: A Double-Blind, Multicentre, Randomized, Placebo-Controlled Study. Vet. Dermatol. 2019, 30, 387-e117. [Google Scholar] [CrossRef]
- Waisglass, S.; Araujio, J.; Della Valle, M.F.; Milgram, N.W. Palmitoylethanolamide in the Management of Canine Atopic Dermatitis. Randomized, Double-Blind, Placebo Controlled Study. In Proceedings of the International Congress of Scivac, Rimini, Italy, 29–31 May 2009; pp. 56–61. [Google Scholar]
- Noli, C.; Della Valle, M.F.; Miolo, A.; Medori, C.; Schievano, C. Skinalia Clinical Research Group Efficacy of Ultra-Micronized Palmitoylethanolamide in Canine Atopic Dermatitis: An Open-Label Multi-Centre Study. Vet. Dermatol. 2015, 26, 432–440, e101. [Google Scholar] [CrossRef]
- Fabbrini, F.; Leone, F. Applicazione Topica Di Adelmidrol (2%) per La Gestione Del Prurito Associato a Dermatite Atopica Del Cane: Studio Osservazionale—Veterinaria. Veterinaria 2013, 27. [Google Scholar]
- Schiano Moriello, A.; Di Marzo, V.; Petrosino, S. Mutual Links between the Endocannabinoidome and the Gut Microbiome, with Special Reference to Companion Animals: A Nutritional Viewpoint. Animals 2022, 12, 348. [Google Scholar] [CrossRef]
- Russo, R.; Cristiano, C.; Avagliano, C.; De Caro, C.; La Rana, G.; Raso, G.M.; Canani, R.B.; Meli, R.; Calignano, A. Gut-Brain Axis: Role of Lipids in the Regulation of Inflammation, Pain and CNS Diseases. Curr. Med. Chem. 2018, 25, 3930–3952. [Google Scholar] [CrossRef]
- Hedlund, P.; Gratzke, C. The Endocannabinoid System - a Target for the Treatment of LUTS? Nat. Rev. Urol. 2016, 13, 463–470. [Google Scholar] [CrossRef]
- Zhou, S.; Ling, X.; Meng, P.; Liang, Y.; Shen, K.; Wu, Q.; Zhang, Y.; Chen, Q.; Chen, S.; Liu, Y.; et al. Cannabinoid Receptor 2 Plays a Central Role in Renal Tubular Mitochondrial Dysfunction and Kidney Ageing. J. Cell Mol. Med. 2021, 25, 8957–8972. [Google Scholar] [CrossRef] [PubMed]
- Chua, J.T.; Argueta, D.A.; DiPatrizio, N.V.; Kovesdy, C.P.; Vaziri, N.D.; Kalantar-Zadeh, K.; Moradi, H. Endocannabinoid System and the Kidneys: From Renal Physiology to Injury and Disease. Cannabis Cannabinoid Res. 2019, 4, 10–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klawitter, J.; Sempio, C.; Jackson, M.J.; Smith, P.H.; Hopp, K.; Chonchol, M.; Gitomer, B.Y.; Christians, U.; Klawitter, J. Endocannabinoid System in Polycystic Kidney Disease. Am. J. Nephrol. 2022, 53, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Britti, D.; Crupi, R.; Impellizzeri, D.; Gugliandolo, E.; Fusco, R.; Schievano, C.; Morittu, V.M.; Evangelista, M.; Di Paola, R.; Cuzzocrea, S. A Novel Composite Formulation of Palmitoylethanolamide and Quercetin Decreases Inflammation and Relieves Pain in Inflammatory and Osteoarthritic Pain Models. BMC Vet. Res. 2017, 13, 229. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, E.; Peritore, A.F.; Impellizzeri, D.; Cordaro, M.; Siracusa, R.; Fusco, R.; D’Amico, R.; Paola, R.D.; Schievano, C.; Cuzzocrea, S.; et al. Dietary Supplementation with Palmitoyl-Glucosamine Co-Micronized with Curcumin Relieves Osteoarthritis Pain and Benefits Joint Mobility. Animals 2020, 10, E1827. [Google Scholar] [CrossRef]
- Annunziata, C.; Lama, A.; Pirozzi, C.; Cavaliere, G.; Trinchese, G.; Di Guida, F.; Nitrato Izzo, A.; Cimmino, F.; Paciello, O.; De Biase, D.; et al. Palmitoylethanolamide Counteracts Hepatic Metabolic Inflexibility Modulating Mitochondrial Function and Efficiency in Diet-Induced Obese Mice. FASEB J. 2020, 34, 350–364. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.M.K.; Uyama, T.; Hussain, Z.; Ueda, N. Roles of Endocannabinoids and Endocannabinoid-Like Molecules in Energy Homeostasis and Metabolic Regulation: A Nutritional Perspective. Annu Rev. Nutr. 2021, 41, 177–202. [Google Scholar] [CrossRef]
- Sihag, J.; Di Marzo, V. (Wh)Olistic (E)Ndocannabinoidome-Microbiome-Axis Modulation through (N)Utrition (WHEN) to Curb Obesity and Related Disorders. Lipids Health Dis. 2022, 21, 9. [Google Scholar] [CrossRef]
- Esposito, G.; Capoccia, E.; Turco, F.; Palumbo, I.; Lu, J.; Steardo, A.; Cuomo, R.; Sarnelli, G.; Steardo, L. Palmitoylethanolamide Improves Colon Inflammation through an Enteric Glia/Toll like Receptor 4-Dependent PPAR-α Activation. Gut 2014, 63, 1300–1312. [Google Scholar] [CrossRef]
- Karwad, M.A.; Macpherson, T.; Wang, B.; Theophilidou, E.; Sarmad, S.; Barrett, D.A.; Larvin, M.; Wright, K.L.; Lund, J.N.; O’Sullivan, S.E. Oleoylethanolamine and Palmitoylethanolamine Modulate Intestinal Permeability in Vitro via TRPV1 and PPARα. FASEB J. 2017, 31, 469–481. [Google Scholar] [CrossRef]
- Sarnelli, G.; Seguella, L.; Pesce, M.; Lu, J.; Gigli, S.; Bruzzese, E.; Lattanzi, R.; D’Alessandro, A.; Cuomo, R.; Steardo, L.; et al. HIV-1 Tat-Induced Diarrhea Is Improved by the PPARalpha Agonist, Palmitoylethanolamide, by Suppressing the Activation of Enteric Glia. J. Neuroinflammat. 2018, 15, 94. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, F.; Romano, B.; Petrosino, S.; Pagano, E.; Capasso, R.; Coppola, D.; Battista, G.; Orlando, P.; Di Marzo, V.; Izzo, A.A. Palmitoylethanolamide, a Naturally Occurring Lipid, Is an Orally Effective Intestinal Anti-Inflammatory Agent. Br. J. Pharmacol. 2015, 172, 142–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capasso, R.; Orlando, P.; Pagano, E.; Aveta, T.; Buono, L.; Borrelli, F.; Di Marzo, V.; Izzo, A.A. Palmitoylethanolamide Normalizes Intestinal Motility in a Model of Post-Inflammatory Accelerated Transit: Involvement of CB1 Receptors and TRPV1 Channels. Br. J. Pharmacol. 2014, 171, 4026–4037. [Google Scholar] [CrossRef] [PubMed]
- Couch, D.G.; Cook, H.; Ortori, C.; Barrett, D.; Lund, J.N.; O’Sullivan, S.E. Palmitoylethanolamide and Cannabidiol Prevent Inflammation-Induced Hyperpermeability of the Human Gut In Vitro and In Vivo-A Randomized, Placebo-Controlled, Double-Blind Controlled Trial. Inflammat. Bowel Dis. 2019, 25, 1006–1018. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, R.; Impellizzeri, D.; Torre, A.; Mazzon, E.; Cappellani, A.; Faggio, C.; Esposito, E.; Trischitta, F.; Cuzzocrea, S. Effects of Palmitoylethanolamide on Intestinal Injury and Inflammation Caused by Ischemia-Reperfusion in Mice. J. Leukoc. Biol. 2012, 91, 911–920. [Google Scholar] [CrossRef]
- Petrini, D.; Di Giuseppe, M.; Deli, G.; De Caro Carella, C. Cystolithiasis in a Syrian Hamster: A Different Outcome. Open Vet. J. 2016, 6, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, E.; Crupi, R.; Peritore, A.F.; Licata, P.; Piras, C.; Cuzzocrea, S.; Britti, D. Interstitial Cystitis/Bladder Pain Syndrome (IC/BPS): Role of N-Palmitoyl-D-Glucosamine-Hesperidin. In Proceedings of the 74th SISVET, Virtual Congress, 23–26 June 2021; p. 243. [Google Scholar]
- Vezzoni, A.; Crupi, F.; Boiocchi, S.; Boano, S. Effect of Palmitoylethanolamide Co-Ultra Micronized with Quercetin in Dogs with Osteoarthritis by Means of Dynamic Gate Analysis and Canine Brief Pain Inventory Questionnaire. In Proceedings of the 5th World Veterinary Orthopaedic Congress ESVOT-VOS, Barcelona, Spain, 12–15 October 2018; 2018; pp. 771–772. [Google Scholar]
- Asperio, R.M. Integrazione dietetica con PGA-Cur: Indagine osservazionale su 181 cani con osteoartrite [Dietary supplementation with PGA-cur: A survey on 181 osteoarthritis dogs]. Summa 2020, 8, 39–48. [Google Scholar]
- Bonello, D.; Squarzoni, P. Effect of a Mucoadhesive Gel and Dental Scaling on Gingivitis in Dogs. J. Vet. Dent. 2008, 25, 28–32. [Google Scholar] [CrossRef]
- De Luca, L.; Ferracane, R.; Vitaglione, P. Food Database of N-Acyl-Phosphatidylethanolamines, N-Acylethanolamines and Endocannabinoids and Daily Intake from a Western, a Mediterranean and a Vegetarian Diet. Food Chem. 2019, 300, 125218. [Google Scholar] [CrossRef]
- Nestmann, E.R. Safety of Micronized Palmitoylethanolamide (MicroPEA): Lack of Toxicity and Genotoxic Potential. Food Sci. Nutr. 2017, 5, 292–309. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
della Rocca, G.; Re, G. Palmitoylethanolamide and Related ALIAmides for Small Animal Health: State of the Art. Biomolecules 2022, 12, 1186. https://doi.org/10.3390/biom12091186
della Rocca G, Re G. Palmitoylethanolamide and Related ALIAmides for Small Animal Health: State of the Art. Biomolecules. 2022; 12(9):1186. https://doi.org/10.3390/biom12091186
Chicago/Turabian Styledella Rocca, Giorgia, and Giovanni Re. 2022. "Palmitoylethanolamide and Related ALIAmides for Small Animal Health: State of the Art" Biomolecules 12, no. 9: 1186. https://doi.org/10.3390/biom12091186