
Molecular Characterization of Kunitz-Type Protease Inhibitors from Blister Beetles (Coleoptera, Meloidae)

 , , , ,
, , , ,  , and
, and 
Abstract
:1. Introduction
1.1. Proteases’ Functions in Living Organisms
1.2. Protease Inhibitors as Drugs for Clinical Applications
1.3. Kunitz-Type Protease Inhibitors (KTPI)
1.4. Kunitz-Type Protease Inhibitors (KTPI) in Insects
1.5. KTPI in the Subfamily Meloinae
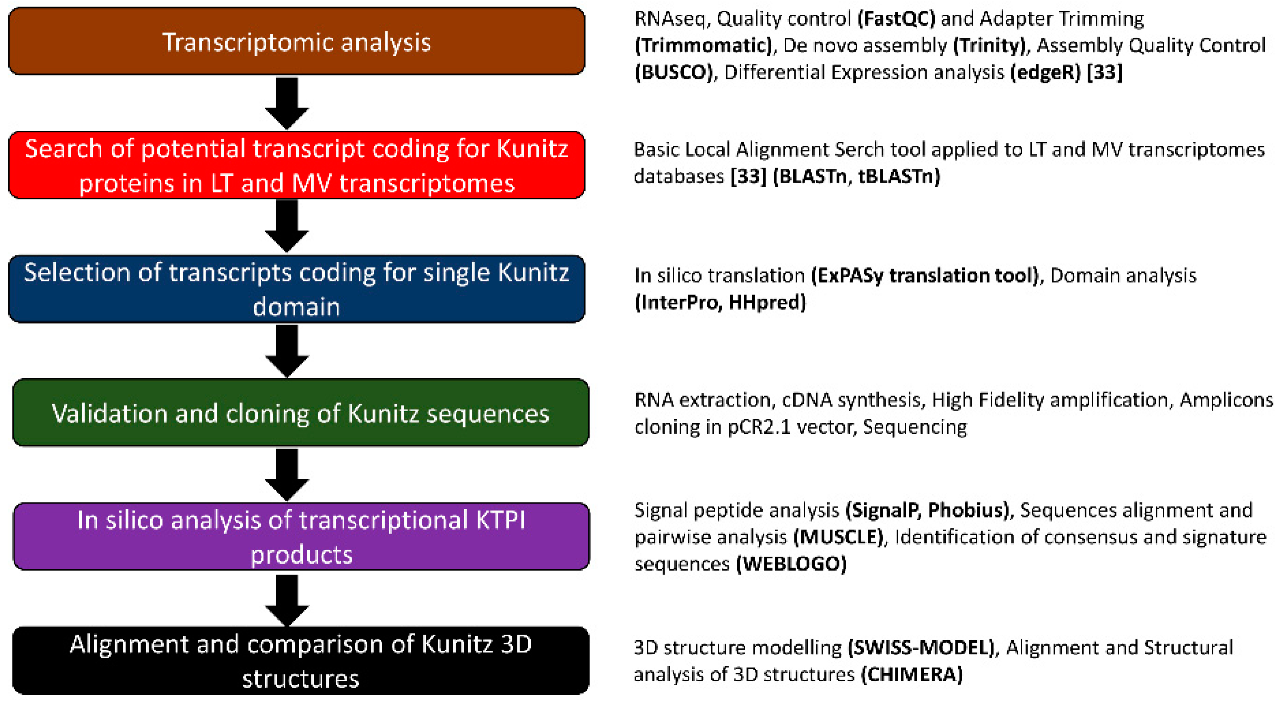
2. Materials and Methods
2.1. BLAST Search for Kunitz-Containing Domain Protein in the Transcriptome of L. trimaculatus and M. variabilis
2.2. Molecular Cloning
2.3. In Silico Analyses of the Identified Sequences
3. Results
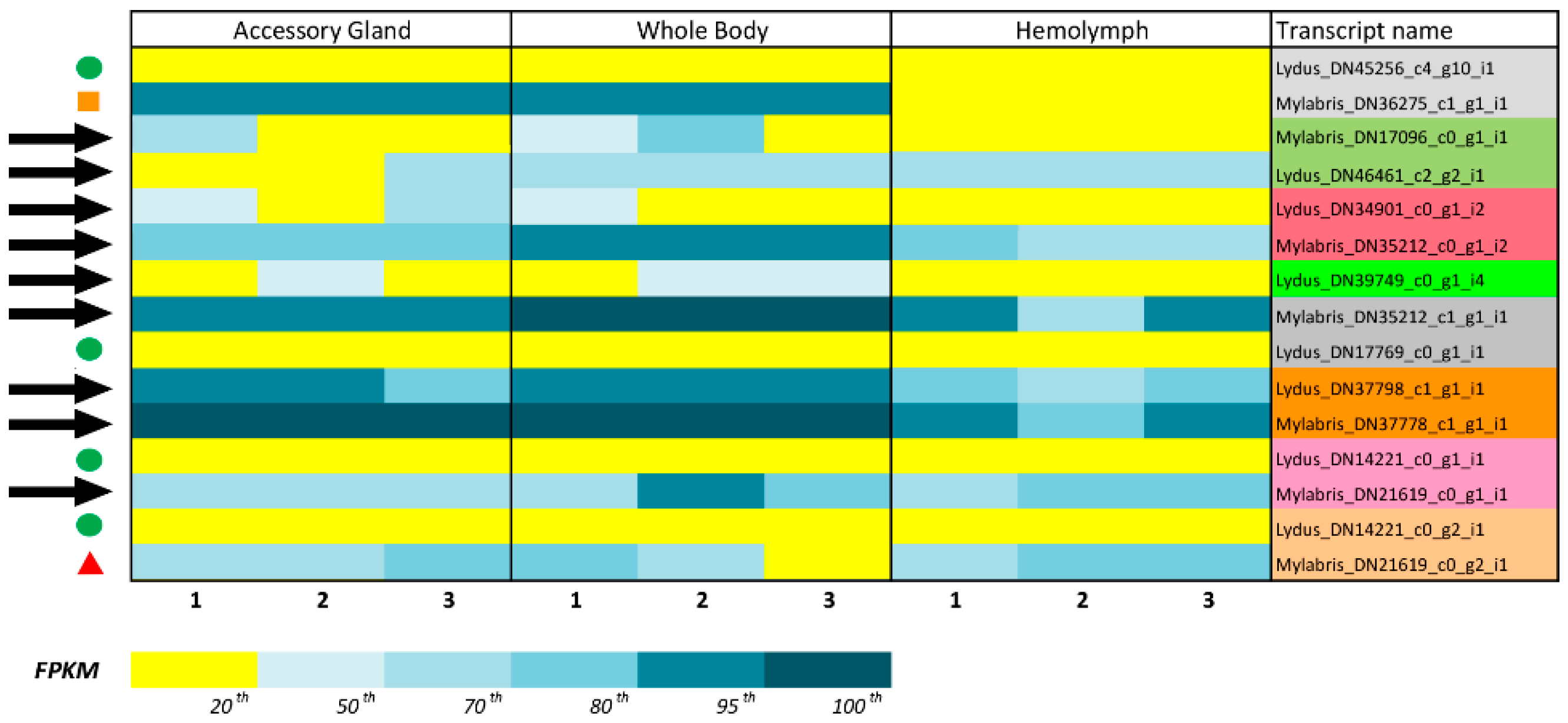
3.1. Identification of Translational KTPI Products from Lydus trimaculatus and Mylabris variabilis
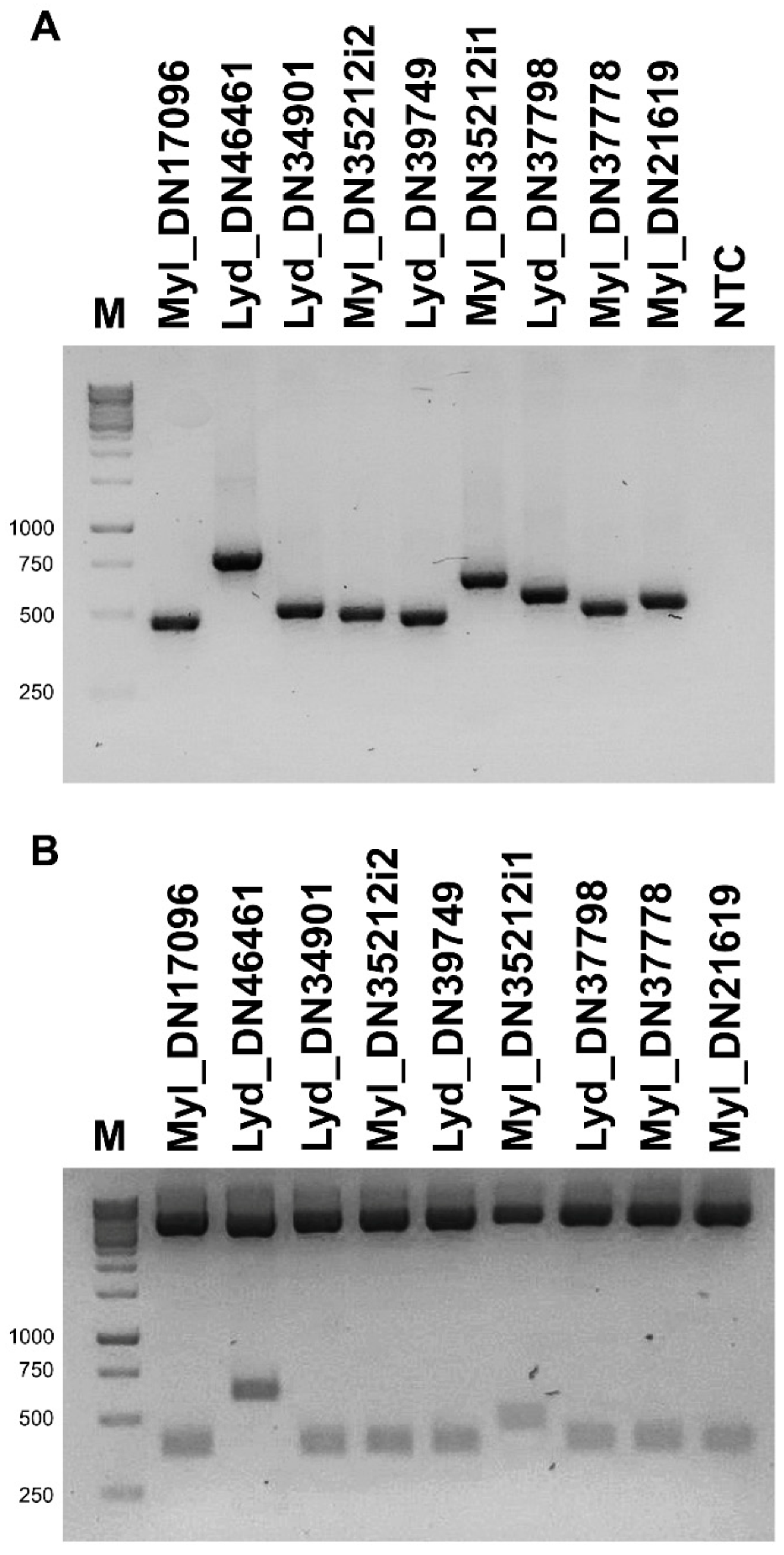
3.2. Validation and Cloning of Kunitz Sequences from Lydus trimaculatus and Mylabris variabilis
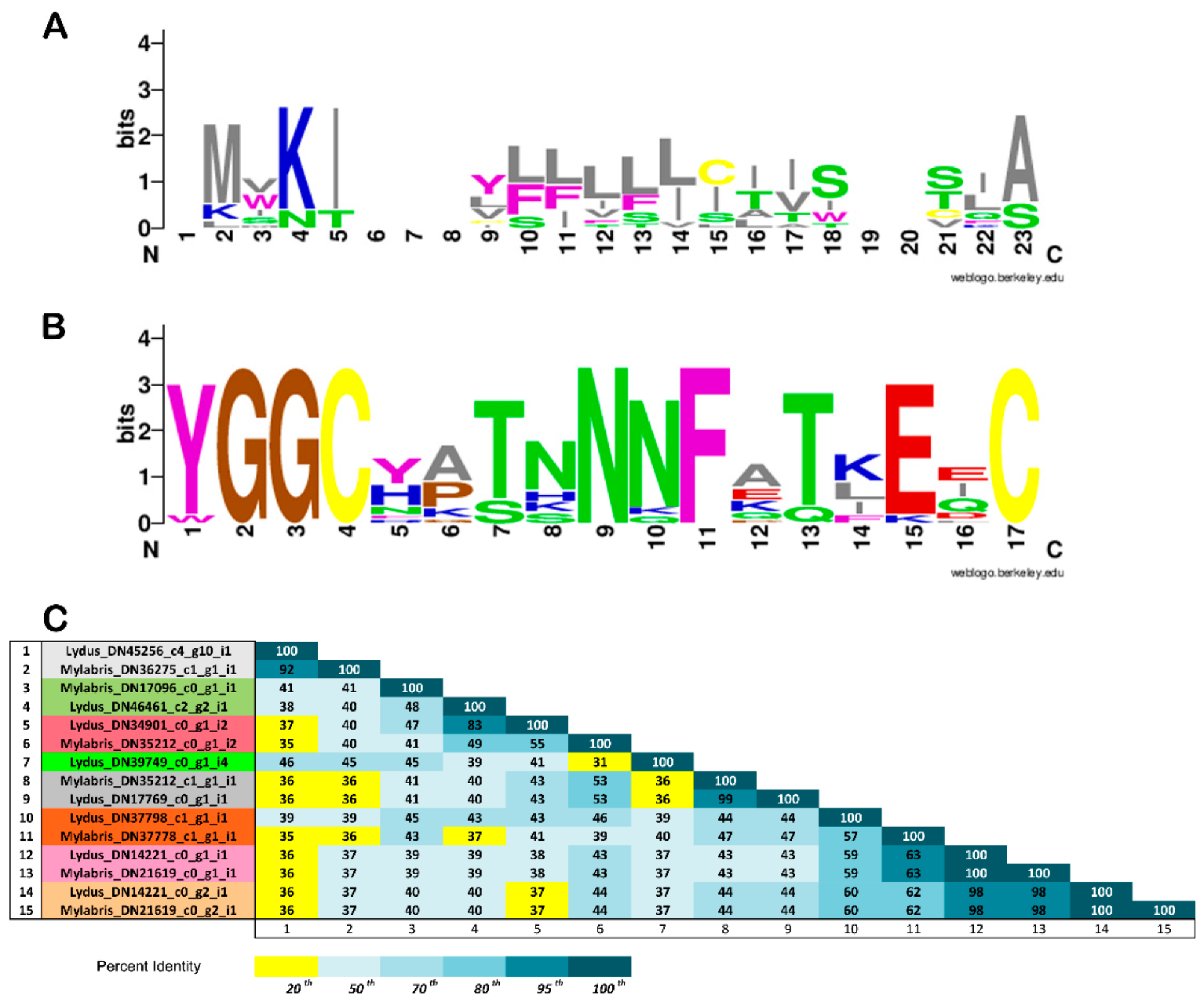
3.3. In Silico Analysis of Translational KTPI Products from Lydus trimaculatus and Mylabris variabilis
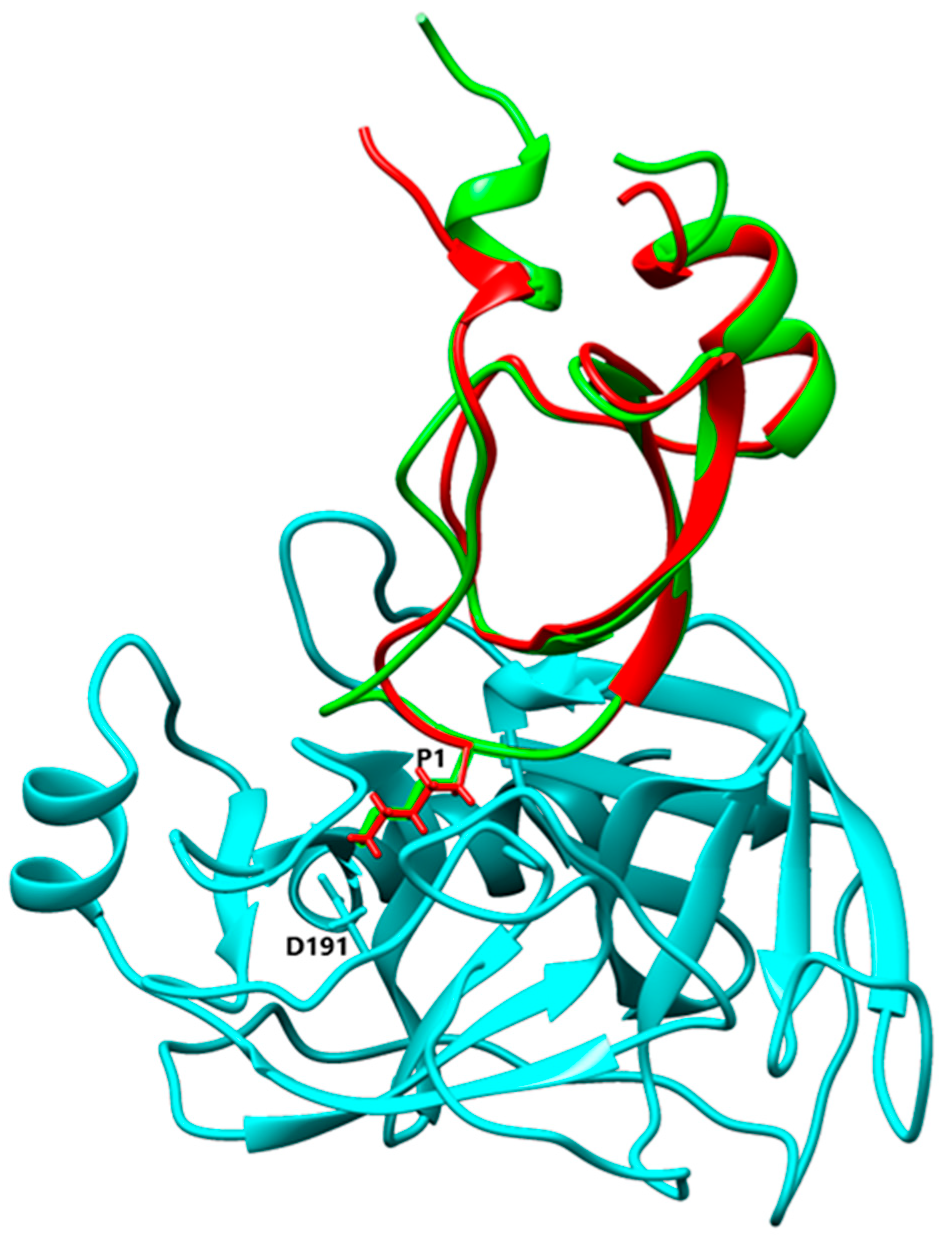
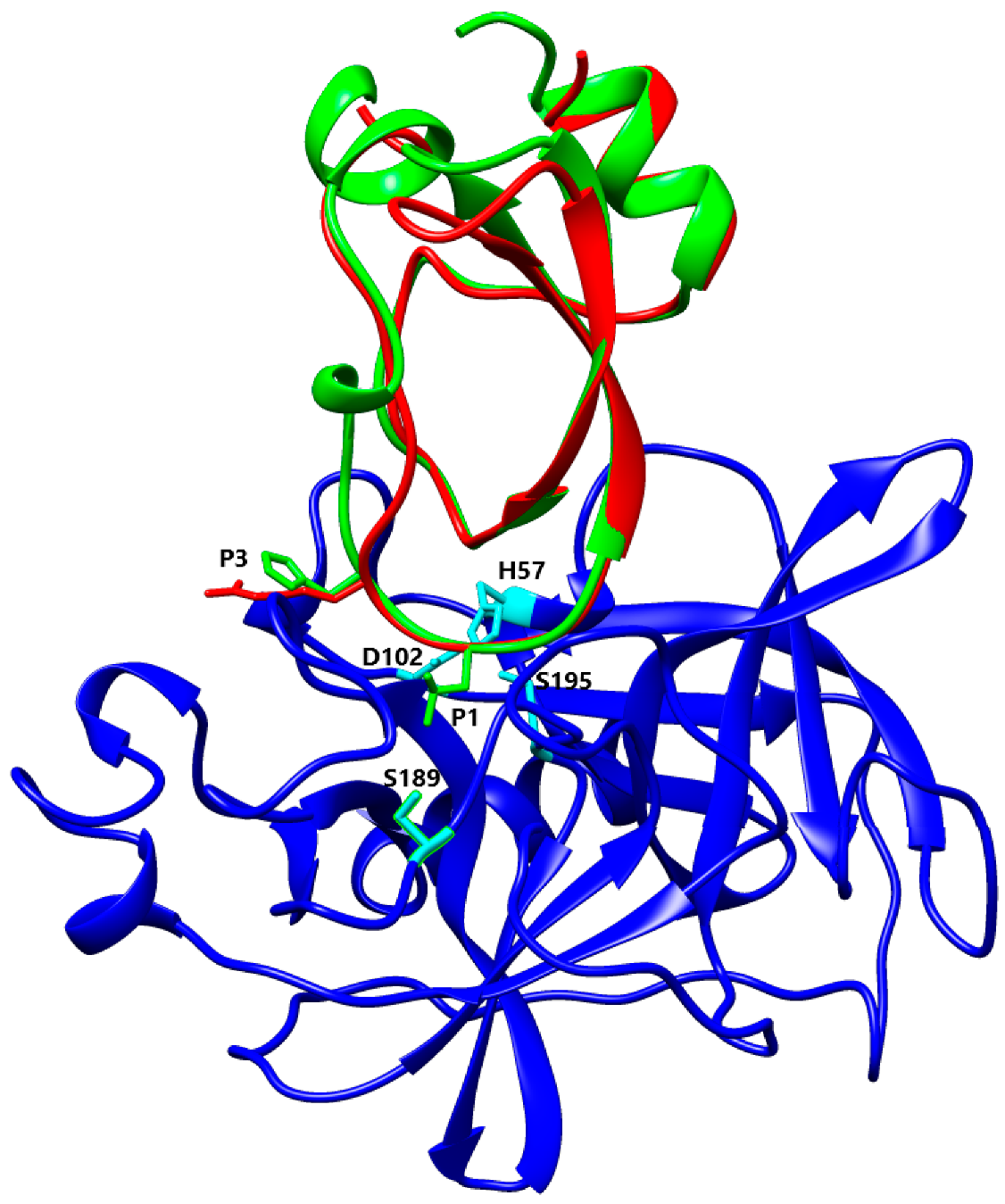
3.4. Alignment and Comparison of Kunitz 3D Structures
4. Discussion
4.1. Meloidae KTPI Identification by Transcriptome Analysis
4.2. P1 Residue Characteristic and Target Protease Specificities
4.3. Basic and Nonpolar P1 Residue
4.4. Acidic P1 Residue
4.5. Natural-KTPI Pharmacological Application
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Rawlings, N.D.; Tolle, D.P.; Barrett, A.J. MEROPS: The peptidase database. Nucleic Acids Res. 2004, 32, D160–D164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drag, M.; Salvesen, G.S. Emerging principles in protease-based drug discovery. Nat. Rev. Drug Discov. 2010, 9, 690–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chattopadhyay, A.; Gray, L.R.; Patton, L.L.; Caplan, D.J.; Slade, G.D.; Tien, H.C.; Shugars, D.C. Salivary secretory leukocyte protease inhibitor and oral candidiasis in human immunodeficiency virus type 1-infected persons. Infect. Immun. 2004, 72, 1956–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potera, R.M.; Jensen, M.J.; Hilkin, B.M.; South, G.K.; Hook, J.S.; Gross, E.A.; Moreland, J.G. Neutrophil azurophilic granule exocytosis is primed by TNF-α and partially regulated by NADPH oxidase. Innate Immun. 2016, 22, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Moraes, T.J.; Chow, C.W.; Downey, G.P. Proteases and lung injury. Crit. Care Med. 2003, 31, S189–S194. [Google Scholar] [CrossRef]
- Kauffman, H.F.; Tamm, M.; Timmerman, J.A.; Borger, P. House dust mite major allergens Der p 1 and Der p 5 activate human airway-derived epithelial cells by protease-dependent and protease-independent mechanisms. Clin. Mol. Allergy 2006, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Polverino, E.; Rosales-Mayor, E.; Dale, G.E.; Dembowsky, K.; Torres, A. The Role of Neutrophil Elastase Inhibitors in Lung Diseases. Chest 2017, 152, 249–262. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, M.M.A.; El-Shimy, I.A.; Hadi, M.A. Neutrophil Elastase Inhibitors: A potential prophylactic treatment option for SARS-CoV-2-induced respiratory complications? Crit. Care 2020, 24, 311. [Google Scholar] [CrossRef]
- Turgeon, V.L.; Houenou, L.J. The role of thrombin-like (serine) proteases in the development, plasticity and pathology of the nervous system. Brain Res. Rev. 1997, 25, 85–95. [Google Scholar] [CrossRef]
- Smith, C.G.; Vane, J.R. The discovery of captopril. FASEB J. 2003, 17, 788–789. [Google Scholar] [CrossRef]
- Flexner, C.; Bate, G.; Kirkpatrick, P. Tipranavir. Nature reviews. Drug Discov. 2005, 4, 955–956. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.S.; Weerapana, E.; Cravatt, B.F. Strategies for discovering and derisking covalent, irreversible enzyme inhibitors. Future Med. Chem. 2010, 2, 949–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junren, C.; Xiaofang, X.; Huiqiong, Z.; Gangmin, L.; Yanpeng, Y.; Xiaoyu, C.; Yuqing, G.; Yanan, L.; Yue, Z.; Fu, P.; et al. Pharmacological Activities and Mechanisms of Hirudin and Its Derivatives-A Review. Front. Pharmacol. 2021, 12, 660757. [Google Scholar] [CrossRef]
- Stoop, A.A.; Craik, C.S. Engineering of a macromolecular scaffold to develop specific protease inhibitors. Nat. Biotechnol. 2003, 21, 1063–1068. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genom. Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef] [Green Version]
- García-Fernández, R.; Peigneur, S.; Pons, T.; Alvarez, C.; González, L.; Chávez, M.A.; Tytgat, J. The Kunitz-Type Protein ShPI-1 Inhibits Serine Proteases and Voltage-Gated Potassium Channels. Toxins 2016, 8, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, A.L.; Robertson, B. Dendrotoxins: Structure-activity relationships and effects on potassium ion channels. Curr. Med. Chem. 2004, 11, 3065–3072. [Google Scholar] [CrossRef]
- Ikeo, K.; Takahashi, K.; Gojobori, T. Evolutionary origin of a Kunitz-type trypsin inhibitor domain inserted in the amyloid beta precursor protein of Alzheimer’s disease. J. Mol. Evol. 1992, 34, 536–543. [Google Scholar] [CrossRef]
- Ben Khalifa, N.; Tyteca, D.; Courtoy, P.J.; Renauld, J.C.; Constantinescu, S.N.; Octave, J.N.; Kienlen-Campard, P. Contribution of Kunitz protease inhibitor and transmembrane domains to amyloid precursor protein homodimerization. Neuro-Degener. Dis. 2012, 10, 92–95. [Google Scholar] [CrossRef]
- Kumthekar, P.; Tang, S.C.; Brenner, A.J.; Kesari, S.; Piccioni, D.E.; Anders, C.; Carrillo, J.; Chalasani, P.; Kabos, P.; Puhalla, S.; et al. ANG1005, a Brain-Penetrating Peptide-Drug Conjugate, Shows Activity in Patients with Breast Cancer with Leptomeningeal Carcinomatosis and Recurrent Brain Metastases. Clin. Cancer Res. 2020, 26, 2789–2799. [Google Scholar] [CrossRef] [Green Version]
- Habib, S.; Singh, M. Angiopep-2-Modified Nanoparticles for Brain-Directed Delivery of Therapeutics: A Review. Polymers 2022, 14, 712. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.H.; O’Donnell, P.S. The therapeutic potential of a kallikrein inhibitor for treating hereditary angioedema. Expert Opin. Investig. Drugs 2006, 15, 1077–1090. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Goenaga, J.; López-Abán, J.; Protasio, A.V.; Vicente Santiago, B.; del Olmo, E.; Vanegas, M.; Fernández-Soto, P.; Patarroyo, M.A.; Muro, A. Peptides Derived of Kunitz-Type Serine Protease Inhibitor as Potential Vaccine Against Experimental Schistosomiasis. Front. Immunol. 2019, 10, 2498. [Google Scholar] [CrossRef] [Green Version]
- Kanost, M.R. Serine proteinase inhibitors in arthropod immunity. Dev. Comp. Immunol. 1999, 23, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, M. Inhibition of the Fungal Protease by Haemolymph Protease Inhibitors of the Silkworm, Bombyx mori L. (Lepidoptera: Bombycidae). Appl. Entomol. Zool. 1982, 17, 589–590. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; Yamashita, M.; Yonehara, S.; Eguchi, M. Properties of fungal protease inhibitors from the integument and haemolymph of the silkworm and effect of an inhibitor on the fungal growth. Comp. Biochem. Physiol. Part B Comp. Biochem. 1990, 95, 559–564. [Google Scholar] [CrossRef]
- Bologna, M.A.; Oliverio, M.; Pitzalis, M.; Mariottini, P. Phylogeny and evolutionary history of the blister beetles (Coleoptera, Meloidae). Mol. Phylogenet. Evol. 2008, 48, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Bologna, M.A.; Turco, F.; Pinto, J.D. Coleoptera, Beetles, Volume 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim) Arthropoda. Isecta. In Handbook of Zoology; Leschen, R.A.B., Lawrence, J.F., Leschen, R.A.B., Beutel, R.G., Lawrence, J.F., Eds.; De Gruyter: Berlin, Germany, 2010. [Google Scholar]
- Bologna, M.A. Coleoptera Meloidae. In Fauna d’Italia; Calderini: Bologna, Italy, 1991; Volume XXVIII. [Google Scholar]
- Riccieri, A.; Mancini, E.; Pitzalis, M.; Salvi, D.; Bologna, M.A. Multigene phylogeny of blister beetles (Coleoptera, Meloidae) reveals an extensive polyphyly of the tribe Lyttini and allows redefining its boundaries. Syst. Entomol. 2022, 1–12. [Google Scholar] [CrossRef]
- Resh, V.H. Autohemorrhage. In Encyclopedia of Insects, 2nd ed.; Resh, V., Carde, R., Eds.; Academic Press: New York, NY, USA, 2009. [Google Scholar]
- Boevé, J.L.; Ducarme, V.; Mertens, T.; Bouillard, P.; Angeli, S. Surface structure, model and mechanism of an insect integument adapted to be damaged easily. J. Nanobiotechnol. 2004, 2, 10. [Google Scholar] [CrossRef] [Green Version]
- Fratini, E.; Salvemini, M.; Lombardo, F.; Muzzi, M.; Molfini, M.; Gisondi, S.; Roma, E.; D’Ezio, V.; Persichini, T.; Gasperi, T.; et al. Unraveling the role of male reproductive tract and haemolymph in cantharidin-exuding Lydus trimaculatus and Mylabris variabilis (Coleoptera: Meloidae): A comparative transcriptomics approach. BMC Genom. 2021, 22, 808. [Google Scholar] [CrossRef]
- Salvi, D.; Maura, M.; Pan, Z.; Bologna, M.A. Phylogenetic systematics of Mylabris blister beetles (Coleoptera, Meloidae): A molecular assessment using species trees and total evidence. Cladistics Int. J. Willi Hennig Soc. 2019, 35, 243–268. [Google Scholar] [CrossRef] [PubMed]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Gallo Cassarino, T.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef]
- Kiefer, F.; Arnold, K.; Künzli, M.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository and associated resources. Nucleic Acids Res. 2009, 37, D387–D392. [Google Scholar] [CrossRef] [Green Version]
- Arnold, K.; Bordoli, L.; Kopp, J.; Schwede, T. The SWISS-MODEL workspace: A web-based environment for protein structure homology modelling. Bioinformatics 2006, 22, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Guex, N.; Peitsch, M.C.; Schwede, T. Automated comparative protein structure modeling with SWISS-MODEL and Swiss-PdbViewer: A historical perspective. Electrophoresis 2009, 30, S162–S173. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Bendre, A.D.; Ramasamy, S.; Suresh, C.G. Analysis of Kunitz inhibitors from plants for comprehensive structural and functional insights. Int. J. Biol. Macromol. 2018, 113, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Tikhonova, I.G.; Jewhurst, H.L.; Drysdale, O.C.; Dvořák, J.; Robinson, M.W.; Cwiklinski, K.; Dalton, J.P. Unexpected Activity of a Novel Kunitz-type Inhibitor: Inhibition of cysteine proteases but not serine proteases. J. Biol. Chem. 2016, 291, 19220–19234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, C.H.; Chen, P.J.; Wang, H.L. Evidence that highly conserved residues of Delonix regia trypsin inhibitor are important for activity. Biochemistry 2010, 75, 1388–1392. [Google Scholar] [CrossRef] [PubMed]
- Yao, P.L.; Hwang, M.J.; Chen, Y.M.; Yeh, K.W. Site-directed mutagenesis evidence for a negatively charged trypsin inhibitory loop in sweet potato sporamin. FEBS Lett. 2001, 496, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Brandsdal, B.O.; Smalås, A.O.; Aqvist, J. Free energy calculations show that acidic P1 variants undergo large pKa shifts upon binding to trypsin. Proteins 2006, 64, 740–748. [Google Scholar] [CrossRef]
- Li, M.; Phylip, L.H.; Lees, W.E.; Winther, J.R.; Dunn, B.M.; Wlodawer, A.; Kay, J.; Gustchina, A. The aspartic proteinase from Saccharomyces cerevisiae folds its own inhibitor into a helix. Nat. Struct. Biol. 2000, 7, 113–117. [Google Scholar] [CrossRef]
- Guo, J.; Erskine, P.T.; Coker, A.R.; Wood, S.P.; Cooper, J.B. Structure of a Kunitz-type potato cathepsin D inhibitor. J. Struct. Biol. 2015, 192, 554–560. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- García-Fernández, R.; Perbandt, M.; Rehders, D.; Ziegelmüller, P.; Piganeau, N.; Hahn, U.; Betzel, C.; Chávez Mde, L.; Redecke, L. Three-dimensional Structure of a Kunitz-type Inhibitor in Complex with an Elastase-like Enzyme. J. Biol. Chem. 2015, 290, 14154–14165. [Google Scholar] [CrossRef] [Green Version]
- Ye, S.; Loll, B.; Berger, A.A.; Mülow, U.; Alings, C.; Wahl, M.C.; Koksch, B. Fluorine teams up with water to restore inhibitor activity to mutant BPTI. Chem. Sci. 2015, 6, 5246–5254. [Google Scholar] [CrossRef] [Green Version]
- Czapinska, H.; Helland, R.; Smalås, A.O.; Otlewski, J. Crystal structures of five bovine chymotrypsin complexes with P1 BPTI variants. J. Mol. Biol. 2004, 344, 1005–1020. [Google Scholar] [CrossRef] [PubMed]
- Helland, R.; Czapinska, H.; Leiros, I.; Olufsen, M.; Otlewski, J.; Smalås, A.O. Structural consequences of accommodation of four non-cognate amino acid residues in the S1 pocket of bovine trypsin and chymotrypsin. J. Mol. Biol. 2003, 333, 845–861. [Google Scholar] [CrossRef] [PubMed]
- Mishra, M. Evolutionary Aspects of the Structural Convergence and Functional Diversification of Kunitz-Domain Inhibitors. J. Mol. Evol. 2020, 88, 537–548. [Google Scholar] [CrossRef]
- Ascenzi, P.; Bocedi, A.; Bolognesi, M.; Spallarossa, A.; Coletta, M.; De Cristofaro, R.; Menegatti, E. The bovine basic pancreatic trypsin inhibitor (Kunitz inhibitor): A milestone protein. Curr. Protein Pept. Sci. 2003, 4, 231–251. [Google Scholar] [CrossRef]
- Hernández González, J.E.; García-Fernández, R.; Valiente, P.A. Polar Desolvation and Position 226 of Pancreatic and Neutrophil Elastases Are Crucial to their Affinity for the Kunitz-Type Inhibitors ShPI-1 and ShPI-1/K13L. PLoS ONE 2015, 10, e0137787. [Google Scholar] [CrossRef]
- Vincent, J.P.; Lazdunski, M. The interaction between alpha-chymotrypsin and pancreatic trypsin inhibitor (Kunitz inhibitor). Kinetic and thermodynamic properties. Eur. J. Biochem. 1973, 38, 365–372. [Google Scholar] [CrossRef]
- Malfertheiner, P.; Büchler, M.; Stanescu, A.; Uhl, W.; Ditschuneit, H. Serum elastase 1 in inflammatory pancreatic and gastrointestinal diseases and in renal insufficiency. A comparison with other serum pancreatic enzymes. Int. J. Pancreatol. Off. J. Int. Assoc. Pancreatol. 1987, 2, 159–170. [Google Scholar] [CrossRef]
- Chughtai, B.; O’Riordan, T.G. Potential role of inhibitors of neutrophil elastase in treating diseases of the airway. J. Aerosol Med. 2004, 17, 289–298. [Google Scholar] [CrossRef]
- Moroy, G.; Alix, A.J.; Sapi, J.; Hornebeck, W.; Bourguet, E. Neutrophil elastase as a target in lung cancer. Anti-Cancer Agents Med. Chem. 2012, 12, 565–579. [Google Scholar] [CrossRef]
- Krowarsch, D.; Dadlez, M.; Buczek, O.; Krokoszynska, I.; Smalas, A.O.; Otlewski, J. Interscaffolding additivity: Binding of P1 variants of bovine pancreatic trypsin inhibitor to four serine proteases. J. Mol. Biol. 1999, 289, 175–186. [Google Scholar] [CrossRef]
- Amin, A. Choosing Non-Vitamin K Antagonist Oral Anticoagulants: Practical Considerations We Need to Know. Ochsner J. 2016, 16, 531–541. [Google Scholar] [PubMed]
- Zucoloto, A.Z.; Jenne, C.N. Platelet-Neutrophil Interplay: Insights Into Neutrophil Extracellular Trap (NET)-Driven Coagulation in Infection. Front. Cardiovasc. Med. 2019, 6, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, C.C.; Shih, T.P.; Ko, W.C.; Tang, H.J.; Hsueh, P.R. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and coronavirus disease-2019 (COVID-19): The epidemic and the challenges. Int. J. Antimicrob. Agents 2020, 55, 105924. [Google Scholar] [CrossRef] [PubMed]
- Turk, B. Targeting proteases: Successes, failures and future prospects. Nature reviews. Drug Discov. 2006, 5, 785–799. [Google Scholar] [CrossRef]
- Hellinger, R.; Gruber, C.W. Peptide-based protease inhibitors from plants. Drug Discov. Today 2019, 24, 1877–1889. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcript | Primers | Primer Name | Fragment Length |
|---|---|---|---|
| Kunitz_Myl_DN17096 | 5′-TAATAAGAGTTGAACCCCAGC-3′ | Kunitz_Myl_DN17096 for | 302 bp |
| 5′-ATCGATCAAAGTACAAATTGCG-3′ | Kunitz_Myl_DN17096 rev | ||
| Kunitz_Lyd_DN46461 | 5′-CTCATCGGTGTATATAAACATC-3′ | Kunitz_Lyd_DN46461 for | 633 bp |
| 5′-TTGAGAAAGTTTATTTAATTTTTTTG-3′ | Kunitz_Lyd_DN46461 rev | ||
| Kunitz_Lyd_DN34901 | 5′-CTCATCGGTGTATATAAACATC-3′ | Kunitz_Lyd_DN34901 for | 380 bp |
| 5′-TTGAGAAAGTTTATTTAATTTTTTTG-3′ | Kunitz_Lyd_DN34901 rev | ||
| Kunitz_Myl_DN35212i2 | 5′-AAAAGTTGCTTATAAGTAACCAAA-3′ | Kunitz_Myl_DN35212i2 for | 324 bp |
| 5′-AACAATTTGGTAAGTTTTTATTATG-3′ | Kunitz_Myl_DN35212i2 rev | ||
| Kunitz_Lyd_DN39749 | 5′-TAATTACACAGCAATAATGTTTAC-3′ | Kunitz_Lyd_DN39749 for | 301 bp |
| 5′-GTACTCTACTTTGCTTACCAAAA-3′ | Kunitz_Lyd_DN39749 rev | ||
| Kunitz_Myl_DN35212i1 | 5′-AATCGTAATTATTGTTGTGTATTG-3′ | Kunitz_Myl_DN35212i1 for | 469 bp |
| 5′-GACAATTGGTGGGTTATAGTTG-3′ | Kunitz_Myl_DN35212i1 rev | ||
| Kunitz_Lyd_DN37798 | 5′-CATCATAAGATTTTTACATATTGC-3′ | Kunitz_Lyd_DN37798 for | 320 bp |
| 5′-GTGAAAATTCAAAATTCCCTCAA-3′ | Kunitz_Lyd_DN37798 rev | ||
| Kunitz_ Myl_DN37778 | 5′-ATTCTAATATCAACAACAATAGCA-3′ | Kunitz_Myl_DN37778 for | 319 bp |
| 5′-GTAAAATTTGAATTTATCAATGCTA-3′ | Kunitz_Myl_DN37778 rev | ||
| Kunitz_Myl_DN21619 | 5′-TAATAACACAGCAATAATGTTTAC-3′ | Kunitz_Myl_DN21619 for | 301 bp |
| 5′-TTTTGGTAAGCAAAGTAGAGTAC-3′ | Kunitz_Myl_DN21619 rev |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fratini, E.; Rossi, M.N.; Spagoni, L.; Riccieri, A.; Mancini, E.; Polticelli, F.; Bologna, M.A.; Mariottini, P.; Cervelli, M. Molecular Characterization of Kunitz-Type Protease Inhibitors from Blister Beetles (Coleoptera, Meloidae). Biomolecules 2022, 12, 988. https://doi.org/10.3390/biom12070988
Fratini E, Rossi MN, Spagoni L, Riccieri A, Mancini E, Polticelli F, Bologna MA, Mariottini P, Cervelli M. Molecular Characterization of Kunitz-Type Protease Inhibitors from Blister Beetles (Coleoptera, Meloidae). Biomolecules. 2022; 12(7):988. https://doi.org/10.3390/biom12070988
Chicago/Turabian StyleFratini, Emiliano, Marianna Nicoletta Rossi, Lucrezia Spagoni, Alessandra Riccieri, Emiliano Mancini, Fabio Polticelli, Marco Alberto Bologna, Paolo Mariottini, and Manuela Cervelli. 2022. "Molecular Characterization of Kunitz-Type Protease Inhibitors from Blister Beetles (Coleoptera, Meloidae)" Biomolecules 12, no. 7: 988. https://doi.org/10.3390/biom12070988