
Metabolomic Analysis of Human Astrocytes in Lipotoxic Condition: Potential Biomarker Identification by Machine Learning Modeling

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Treatment of Human Astrocytes with Palmitic Acid
2.3. Endogenous Metabolite Extraction (Fingerprinting)
2.4. Exogenous Metabolite Extraction (Footprinting)
2.5. Derivatization
2.6. Metabolomics Analysis
2.6.1. Instrumental Analysis
2.6.2. Data Processing and Annotation
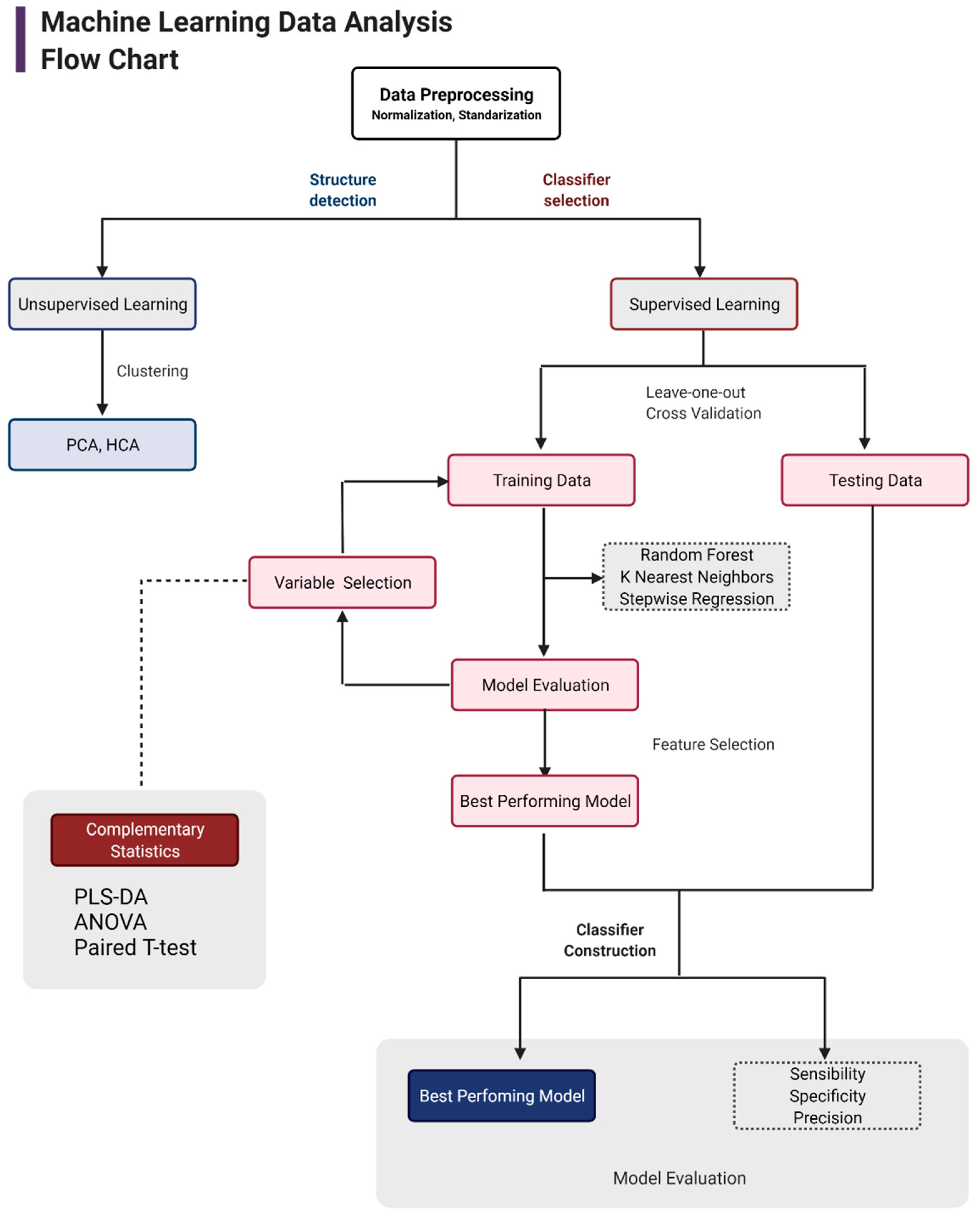
2.7. Metabolomics Data Analysis with Machine Learning
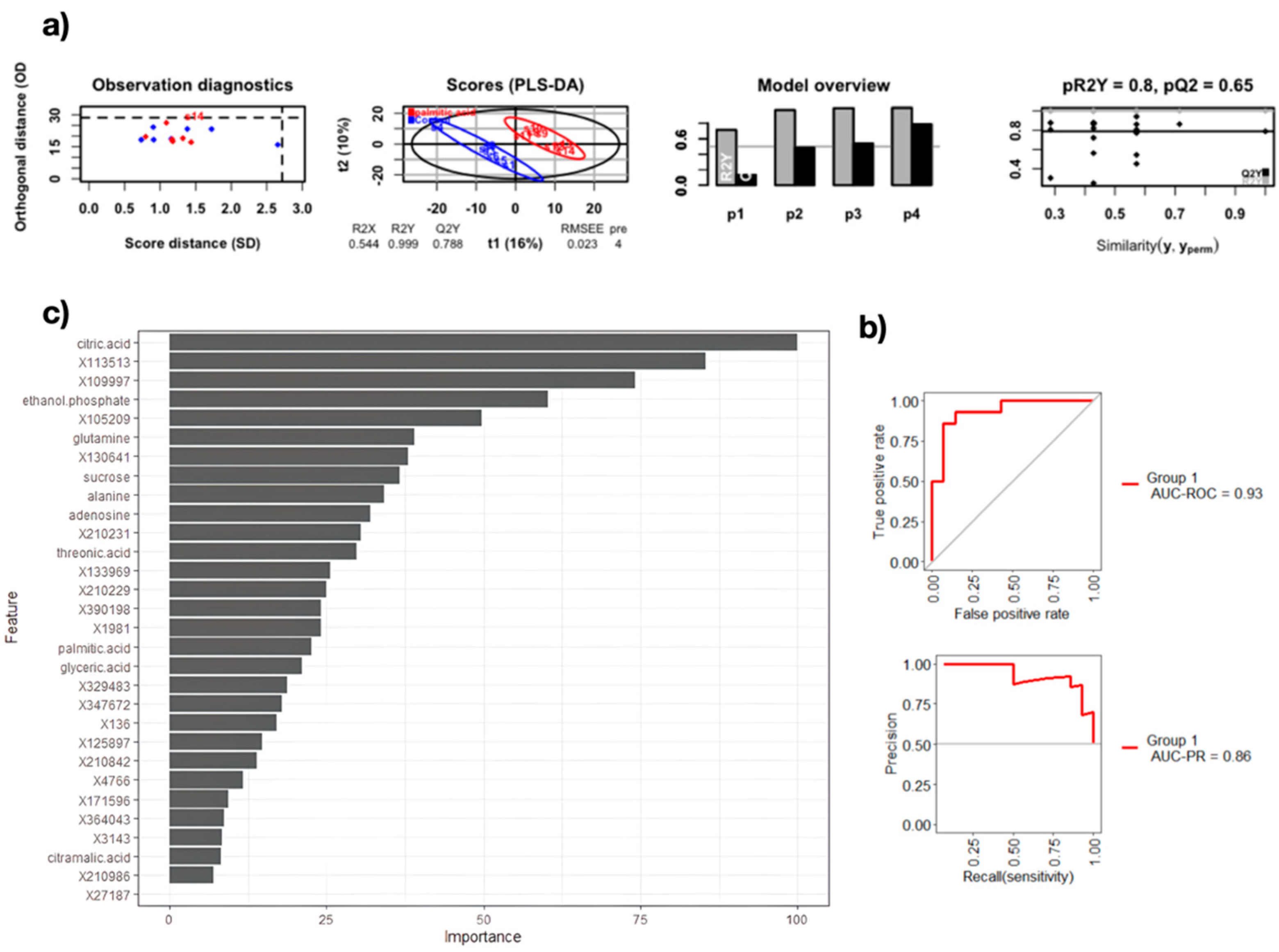
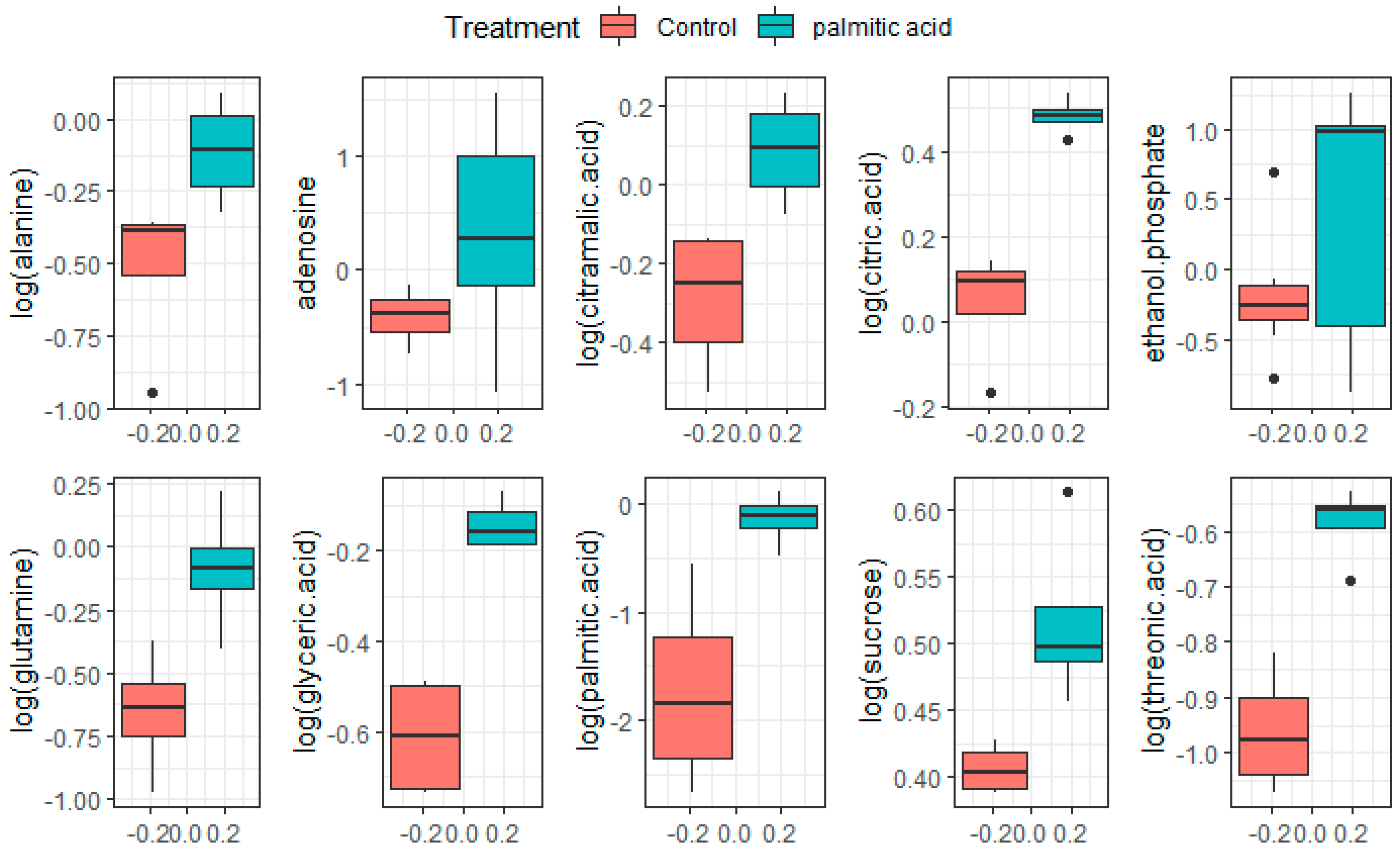
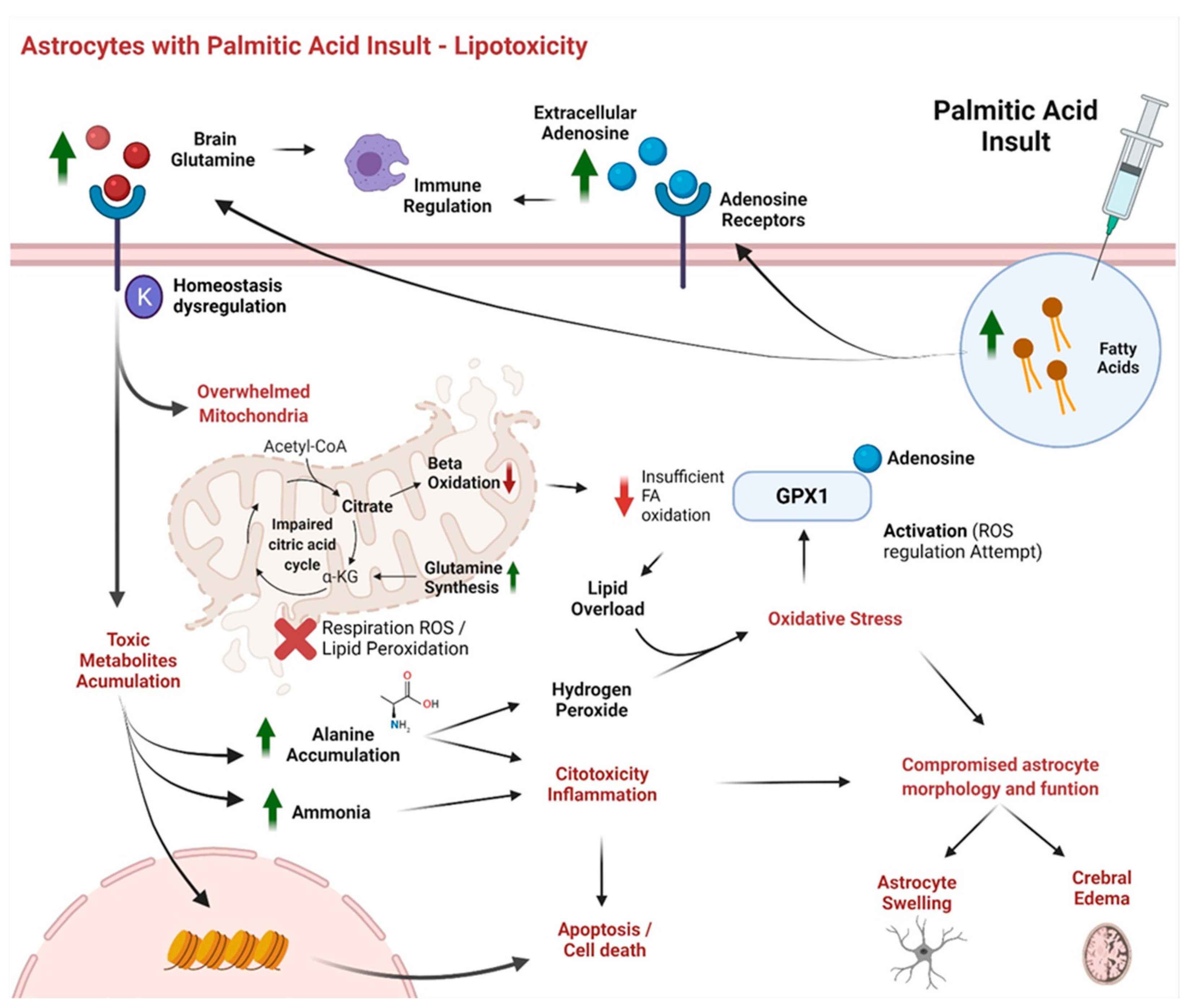
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 4 April 2022).
- Jung, U.J.; Choi, M.S. Obesity and Its Metabolic Complications: The Role of Adipokines and the Relationship between Obesity, Inflammation, Insulin Resistance, Dyslipidemia and Nonalcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lelliott, C.; Vidal-Puig, A.J. Lipotoxicity, an Imbalance between Lipogenesis de Novo and Fatty Acid Oxidation. Int. J. Obes. 2004, 28, S22–S28. [Google Scholar] [CrossRef] [Green Version]
- Unger, R.H. Lipotoxicity in the Pathogenesis of Obesity-Dependent NIDDM: Genetic and Clinical Implications. Diabetes 1995, 44, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.W.; Zhang, X.; Huang, W.-J. Role of Neuroinflammation in Neurodegenerative Diseases (Review). Mol. Med. Rep. 2016, 13, 3391–3396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laske, C.; Stellos, K.; Hoffmann, N.; Stransky, E.; Straten, G.; Eschweiler, G.W.; Leyhe, T. Higher BDNF Serum Levels Predict Slower Cognitive Decline in Alzheimer’s Disease Patients. Int. J. Neuropsychopharmacol. 2011, 14, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Molteni, R.; Barnard, R.J.; Ying, Z.; Roberts, C.K.; Gómez-Pinilla, F. A High-Fat, Refined Sugar Diet Reduces Hippocampal Brain-Derived Neurotrophic Factor, Neuronal Plasticity, and Learning. Neuroscience 2002, 112, 803–814. [Google Scholar] [CrossRef] [Green Version]
- De Zwaan, M.; Mitchell, J.E.; Howell, L.M.; Monson, N.; Swan-Kremeier, L.; Roerig, J.L.; Kolotkin, R.L.; Crosby, R.D. Two Measures of Health-Related Quality of Life in Morbid Obesity. Obes. Res. 2002, 10, 1143–1151. [Google Scholar] [CrossRef] [Green Version]
- Kien, L.; Bunn, J.Y.; Ugrasbul, F. Increasing Dietary Palmitic Acid Decreases Fat Oxidation and Daily Energy Expenditure. Am. J. Clin. Nutr. 2005, 82, 320–326. [Google Scholar] [CrossRef]
- Ajuwon, K.M.; Spurlock, M.E. Palmitate Activates the NF-ΚB Transcription Factor and Induces IL-6 and TNFα Expression in 3T3-L1 Adipocytes. J. Nutr. 2005, 135, 1841–1846. [Google Scholar] [CrossRef]
- Maloney, E.; Sweet, I.R.; Hockenbery, D.M.; Pham, M.; Rizzo, N.O.; Tateya, S.; Handa, P.; Schwartz, M.W.; Kim, F. Activation of NF-ΚB by Palmitate in Endothelial Cells: A Key Role for NADPH Oxidase-Derived Superoxide in Response to TLR4 Activation. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1370–1375. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Martin, R.; Chan, C. Palmitate-Activated Astrocytes via Serine Palmitoyltransferase Increase BACE1 in Primary Neurons by Sphingomyelinases. Neurobiol. Aging 2013, 34, 540–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, C.M.; Belsham, D.D. Palmitate Attenuates Insulin Signaling and Induces Endoplasmic Reticulum Stress and Apoptosis in Hypothalamic Neurons: Rescue of Resistance and Apoptosis through Adenosine 5′ Monophosphate-Activated Protein Kinase Activation. Endocrinology 2010, 151, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Contreras, C.; González-García, I.; Martínez-Sánchez, N.; Seoane-Collazo, P.; Jacas, J.; Morgan, D.A.; Serra, D.; Gallego, R.; Gonzalez, F.; Casals, N.; et al. Central Ceramide-Induced Hypothalamic Lipotoxicity and ER Stress Regulate Energy Balance. Cell Rep. 2014, 9, 366–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Wang, D.; Topczewski, F.; Pagliassotti, M.J. Saturated Fatty Acids Induce Endoplasmic Reticulum Stress and Apoptosis Independently of Ceramide in Liver Cells. Am. J. Physiol. Endocrinol. Metab. 2006, 291, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Phatnani, H.; Maniatis, T. Astrocytes in Neurodegenerative Disease. Cold Spring Harb. Perspect. Biol. 2015, 7, a020628. [Google Scholar] [CrossRef] [Green Version]
- Pekny, M.; Pekna, M. Astrocyte Reactivity and Reactive Astrogliosis: Costs and Benefits. Physiol. Rev. 2014, 94, 1077–1098. [Google Scholar] [CrossRef]
- De Keyser, J.; Mostert, J.P.; Koch, M.W. Dysfunctional Astrocytes as Key Players in the Pathogenesis of Central Nervous System Disorders. J. Neurol. Sci. 2008, 267, 3–16. [Google Scholar] [CrossRef]
- Sofroniew, M.V. Astrocyte Barriers to Neurotoxic Inflammation. Nat. Rev. Neurosci. 2015, 16, 249–263. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, J.; Toku, K.; Zhang, B.O.; Ishihara, K.; Sakanaka, M.; Maeda, N. Astrocytes Prevent Neuronal Death Induced by Reactive Oxygen and Nitrogen Species. Glia 1999, 28, 85–96. [Google Scholar] [CrossRef]
- Abbott, N.J. Astrocyte-Endothelial Interactions and Blood-Brain Barrier Permeability. J. Anat. 2002, 200, 523–534. [Google Scholar] [CrossRef]
- Cabezas, R.; Ávila, M.; Gonzalez, J.; El-Bachá, R.S.; Báez, E.; García-Segura, L.M.; Coronel, J.C.J.; Capani, F.; Cardona-Gomez, G.P.; Barreto, G.E. Astrocytic Modulation of Blood Brain Barrier: Perspectives on Parkinson’s Disease. Front. Cell. Neurosci. 2014, 8, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Benveniste, E.N. Immune Function of Astrocytes. Glia 2001, 36, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Guillamón-Vivancos, T.; Gómez-Pinedo, U.; Matías-Guiu, J. Astrocytes in Neurodegenerative Diseases (I): Function and Molecular Description. Neurologia 2015, 30, 119–129. [Google Scholar] [CrossRef]
- Montgomery, D.L. Astrocytes: Form, Functions, and Roles in Disease. Vet. Pathol. 1994, 31, 145–167. [Google Scholar] [CrossRef] [PubMed]
- Koehler, R.C.; Gebremedhin, D.; Harder, D.R. Role of Astrocytes in Cerebrovascular Regulation. J. Appl. Physiol. 2006, 100, 307–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson, R.; Ying, W.; Kauppinen, T. Astrocyte Influences on Ischemic Neuronal Death. Curr. Mol. Med. 2005, 4, 193–205. [Google Scholar] [CrossRef]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and Pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [Green Version]
- Barreto, G.E.; Gonzalez, J.; Torres, Y.; Morales, L. Astrocytic-Neuronal Crosstalk: Implications for Neuroprotection from Brain Injury. Neurosci. Res. 2011, 71, 107–113. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics—the Link between Genotypes and Phenotypes. In Functional Genomics; Springer: Berlin/Heidelberg, Germany, 2002; pp. 155–171. [Google Scholar] [CrossRef]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond Biomarkers and towards Mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, P.; Johansson, A.I.; Gullberg, J.; Trygg, J.; Jiye, A.; Grung, B.; Marklund, S.; Sjöström, M.; Antti, H.; Moritz, T. High-Throughput Data Analysis for Detecting and Identifying Differences between Samples in GC/MS-Based Metabolomic Analyses. Anal. Chem. 2005, 77, 5635–5642. [Google Scholar] [CrossRef]
- Rozen, S.; Cudkowicz, M.E.; Bogdanov, M.; Matson, W.R.; Kristal, B.S.; Beecher, C.; Harrison, S.; Vouros, P.; Flarakos, J.; Vigneau-Callahan, K.; et al. Metabolomic Analysis and Signatures in Motor Neuron Disease. Metabolomics 2005, 1, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Du, H.; Xu, L.; Wang, J.; Tang, L.; Cao, Y.; Li, C.; Wang, Q.; Liu, Y.; Shan, F.; et al. Metabolomic Analysis Revealed Glycylglycine Accumulation in Astrocytes after Methionine Enkephalin Administration Exhibiting Neuron Protective Effects. J. Pharm. Biomed. Anal. 2015, 115, 48–54. [Google Scholar] [CrossRef]
- Aliferis, K.A.; Jabaji, S. Metabolomics–A Robust Bioanalytical Approach for the Discovery of the Modes-of-Action of Pesticides: A Review. Pestic. Biochem. Physiol. 2011, 100, 105–117. [Google Scholar] [CrossRef]
- Lämmerhofer, M.; Weckwerth, W. Metabolomics in Practice: Successful Strategies to Generate and Analyze-Google Libros; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Oliver, A.H.J. Metabolomics and Systems Biology in Human Health and Medicine; Cabi: Boston, MA, USA, 2014. [Google Scholar]
- Čuperlović-Culf, M.; Barnett, D.A.; Culf, A.S.; Chute, I. Cell Culture Metabolomics: Applications and Future Directions. Drug Discov. Today 2010, 15, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Castellanos, D.B.; Martín-Jiménez, C.A.; Rojas-Rodríguez, F.; Barreto, G.E.; González, J. Brain Lipidomics as a Rising Field in Neurodegenerative Contexts: Perspectives with Machine Learning Approaches. Front. Neuroendocrinol. 2021, 61, 100899. [Google Scholar] [CrossRef] [PubMed]
- Ausim Azizi, S.; Krynska, B. Derivation of Neuronal Cells from Fetal Normal Human Astrocytes (NHA). Methods Mol. Biol. 2013, 1078, 89–96. [Google Scholar] [CrossRef]
- Shao, H.; Chung, J.; Balaj, L.; Charest, A.; Bigner, D.D.; Carter, B.S.; Hochberg, F.H.; Breakefield, X.O.; Weissleder, R.; Lee, H. Protein Typing of Circulating Microvesicles Allows Real-Time Monitoring of Glioblastoma Therapy. Nat. Med. 2012, 18, 1835–1840. [Google Scholar] [CrossRef]
- Sutinen, E.M.; Pirttilä, T.; Anderson, G.; Salminen, A.; Ojala, J.O. Pro-Inflammatory Interleukin-18 Increases Alzheimer’s Disease-Associated Amyloid-β Production in Human Neuron-like Cells. J. Neuroinflammation 2012, 9, 199. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.; Wu, M.; Zhang, L.; Thompson, R.; Nath, A.; Chan, C. Signaling Dynamics of Palmitate-Induced ER Stress Responses Mediated by ATF4 in HepG2 Cells. BMC Syst. Biol. 2013, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Dunn, W.B.; Ellis, D.I. Metabolomics: Current Analytical Platforms and Methodologies. TrAC Trends Anal. Chem. 2005, 24, 285–294. [Google Scholar] [CrossRef]
- Weckwerth, W. Metabolomics: Methods and Protocols; Humana Press: Totowa, NJ, USA, 2007. [Google Scholar]
- Van den Berg, R.A.; Hoefsloot, H.C.J.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, Scaling, and Transformations: Improving the Biological Information Content of Metabolomics Data. BMC Genom. 2006, 7, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdi, H.; Williams, L.J. Principal Component Analysis. Wiley Interdiscip. Rev. Comput. Stat. 2010, 2, 433–459. [Google Scholar] [CrossRef]
- Murtagh, F.; Contreras, P. Methods of Hierarchical Clustering. arXiv 2011, arXiv:1105.0121. [Google Scholar] [CrossRef]
- Burkov, A. Machine Learning Kompakt: Alles, Was Sie Wissen Müssen; MITP: Frechen, Germany, 2019. [Google Scholar]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A Web Server for Metabolomic Data Analysis and Interpretation. Nucleic Acids Res. 2009, 37 (Suppl. 2), W652–W660. [Google Scholar] [CrossRef] [Green Version]
- Manzoni, A.G.; Passos, D.F.; Leitemperger, J.W.; Storck, T.R.; Doleski, P.H.; Jantsch, M.H.; Loro, V.L.; Leal, D.B.R. Hyperlipidemia-Induced Lipotoxicity and Immune Activation in Rats Are Prevented by Curcumin and Rutin. Int. Immunopharmacol. 2020, 81, 106217. [Google Scholar] [CrossRef]
- Brusilow, S.W.; Koehler, R.C.; Traystman, R.J.; Cooper, A.J.L. Astrocyte Glutamine Synthetase: Importance in Hyperammonemic Syndromes and Potential Target for Therapy. Neurotherapeutics 2010, 7, 452–470. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Weber, K.J.; Schilling, J.D. Glutamine Modulates Macrophage Lipotoxicity. Nutrients 2016, 8, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haskó, G.; Csóka, B.; Németh, Z.H.; Vizi, E.S.; Pacher, P. A2B Adenosine Receptors in Immunity and Inflammation. Trends Immunol. 2009, 30, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Stegman, L.D.; Zheng, H.; Neal, E.R.; Ben-Yoseph, O.; Pollegioni, L.; Pilone, M.S.; Ross, B.D. Induction of Cytotoxic Oxidative Stress by D-Alanine in Brain Tumor Cells Expressing Rhodotorula Gracilis D-Amino Acid Oxidase: A Cancer Gene Therapy Strategy. Hum. Gene Ther. 1998, 9, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Trezzi, J.P.; Galozzi, S.; Jaeger, C.; Barkovits, K.; Brockmann, K.; Maetzler, W.; Berg, D.; Marcus, K.; Betsou, F.; Hiller, K.; et al. Distinct Metabolomic Signature in Cerebrospinal Fluid in Early Parkinson’s Disease. Mov. Disord. 2017, 32, 1401–1408. [Google Scholar] [CrossRef]
- Trépanier, G.; Furling, D.; Puymirat, J.; Mirault, M.E. Immunocytochemical Localization of Seleno-Glutathione Peroxidase in the Adult Mouse Brain. Neuroscience 1996, 75, 231–243. [Google Scholar] [CrossRef]
- Tapiero, H.; Townsend, D.M.; Tew, K.D. The Antioxidant Role of Selenium and Seleno-Compounds. Biomed. Pharmacother. 2003, 57, 134–144. [Google Scholar] [CrossRef]
- Lei, X.G.; Cheng, W.H.; McClung, J.P. Metabolic Regulation and Function of Glutathione Peroxidase-1. Annu. Rev. Nutr. 2007, 27, 41–61. [Google Scholar] [CrossRef] [PubMed]
- Osorio, D.; Pinzón, A.; Martín-Jiménez, C.; Barreto, G.E.; González, J. Multiple Pathways Involved in Palmitic Acid-Induced Toxicity: A System Biology Approach. Front. Neurosci. 2020, 13, 1410. [Google Scholar] [CrossRef] [Green Version]
- Gray, C.W.; Patel, A.J. Neurodegeneration Mediated by Glutamate and β-Amyloid Peptide: A Comparison and Possible Interaction. Brain Res. 1995, 691, 169–179. [Google Scholar] [CrossRef]
- Kim, A.Y.; Baik, E.J. Glutamate Dehydrogenase as a Neuroprotective Target Against Neurodegeneration. Neurochem. Res. 2019, 44, 147–153. [Google Scholar] [CrossRef]
- Lewerenz, J.; Maher, P. Chronic Glutamate Toxicity in Neurodegenerative Diseases-What Is the Evidence? Front. Neurosci. 2015, 9, 469. [Google Scholar] [CrossRef]
- Pioro, E.P.; Majors, A.W.; Mitsumoto, H.; Nelson, D.R.; Ng, T.C. 1H-MRS Evidence of Neurodegeneration and Excess Glutamate Glutamine in ALS Medulla. Neurology 1999, 53, 71. [Google Scholar] [CrossRef]
- Gropman, A.L.; Summar, M.; Leonard, J.V. Neurological Implications of Urea Cycle Disorders. J. Inherit. Metab. Dis. 2007, 30, 865–879. [Google Scholar] [CrossRef]
- Haffar, T.; Akoumi, A.; Bousette, N. Lipotoxic Palmitate Impairs the Rate of β-Oxidation and Citric Acid Cycle Flux in Rat Neonatal Cardiomyocytes. Cell. Physiol. Biochem. 2016, 40, 969–981. [Google Scholar] [CrossRef]
- Waagepetersen, H.S.; Sonnewald, U.; Larsson, O.M.; Schousboe, A. A possible role of alanine for ammonia transfer between astrocytes and glutamatergic neurons. J. Neurochem. 2000, 75, 471–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boison, D.; Chen, J.F.; Fredholm, B.B. Adenosine signaling and function in glial cells. Cell Death Differ. 2010, 17, 1071–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hines, D.J.; Haydon, P.G. Astrocytic adenosine: From synapses to psychiatric disorders. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2014, 369, 20130594. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metric | Random Forest | K-Nearest Neighbor | Stepwise Regression |
|---|---|---|---|
| Sensibility (%) | 80.57 | 92.85 | 92.85 |
| Specificity (%) | 85.71 | 85.71 | 71.42 |
| Area under the ROC curve (%) | 93.06 | 93.62 | 79.84 |
| Area under the precision recall curve | 86 | 44 | 25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castellanos, D.B.; Martín-Jiménez, C.A.; Pinzón, A.; Barreto, G.E.; Padilla-González, G.F.; Aristizábal, A.; Zuluaga, M.; González Santos, J. Metabolomic Analysis of Human Astrocytes in Lipotoxic Condition: Potential Biomarker Identification by Machine Learning Modeling. Biomolecules 2022, 12, 986. https://doi.org/10.3390/biom12070986
Castellanos DB, Martín-Jiménez CA, Pinzón A, Barreto GE, Padilla-González GF, Aristizábal A, Zuluaga M, González Santos J. Metabolomic Analysis of Human Astrocytes in Lipotoxic Condition: Potential Biomarker Identification by Machine Learning Modeling. Biomolecules. 2022; 12(7):986. https://doi.org/10.3390/biom12070986
Chicago/Turabian StyleCastellanos, Daniel Báez, Cynthia A. Martín-Jiménez, Andrés Pinzón, George E. Barreto, Guillermo Federico Padilla-González, Andrés Aristizábal, Martha Zuluaga, and Janneth González Santos. 2022. "Metabolomic Analysis of Human Astrocytes in Lipotoxic Condition: Potential Biomarker Identification by Machine Learning Modeling" Biomolecules 12, no. 7: 986. https://doi.org/10.3390/biom12070986