
Gamma-Linolenic Acid (GLA) Protects against Ionizing Radiation-Induced Damage: An In Vitro and In Vivo Study

 ,
, 
Abstract
:1. Introduction
2. Material and Methods: Reagents
2.1. Cell Culture Conditions
2.2. Experimental Animals
2.3. Irradiation Procedure
2.4. In Vitro Studies
2.4.1. Exposure of RAW 264.7 Cells to Radiation
2.4.2. Study of Effect of PUFAs on Viability of RAW 264.7 Cells In Vitro
2.4.3. Determination of Radio-Protective Efficacy of PUFAs by MTT Assay
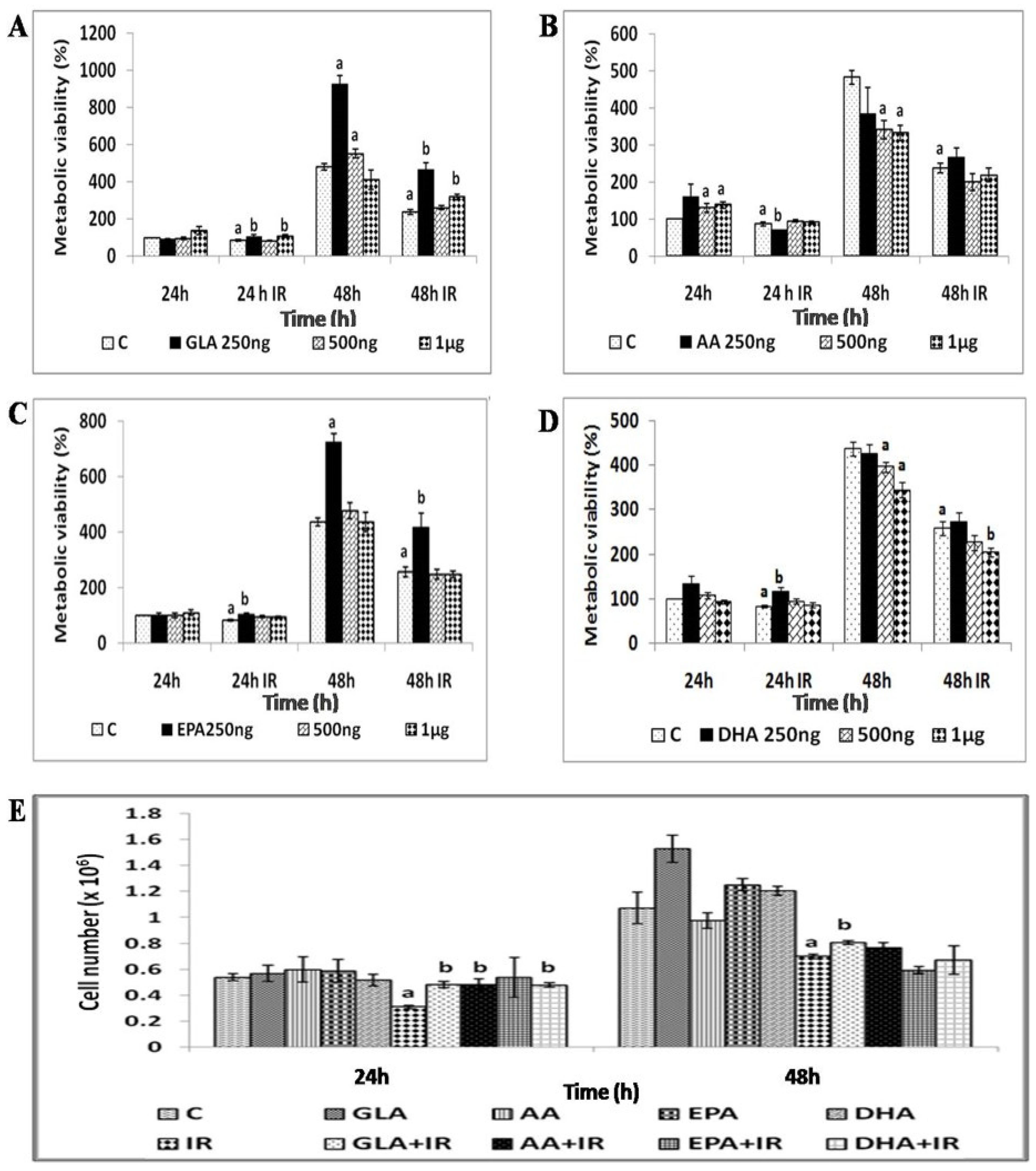
2.4.4. Radio-Protective Efficacy of PUFAs by Growth Kinetics Assay
2.4.5. Effect of PUFAs on Generation of Intracellular ROS
2.4.6. Effect of PUFAs on Antioxidant Enzymes, Lipid Peroxides, and Nitric Oxide (NO)
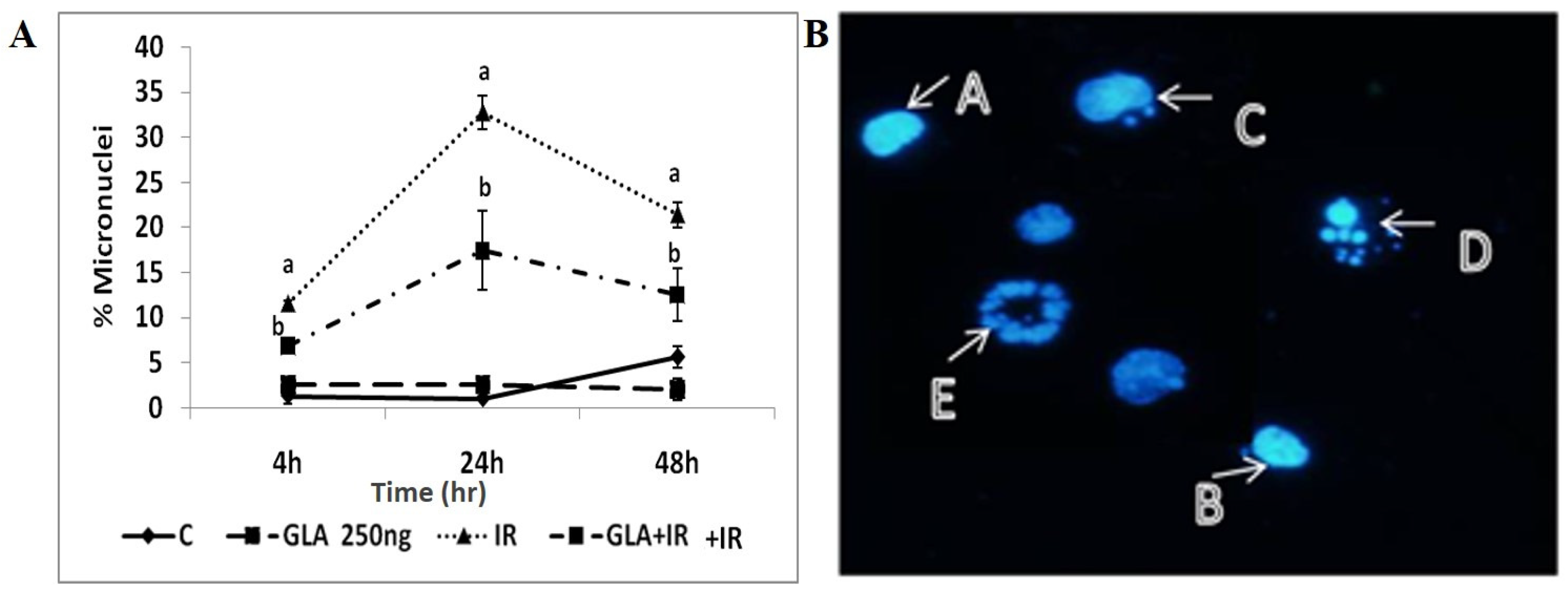
2.4.7. Micronuclei Analysis
2.4.8. Gene Expression Analysis in RAW Cells
2.5. In Vivo Studies
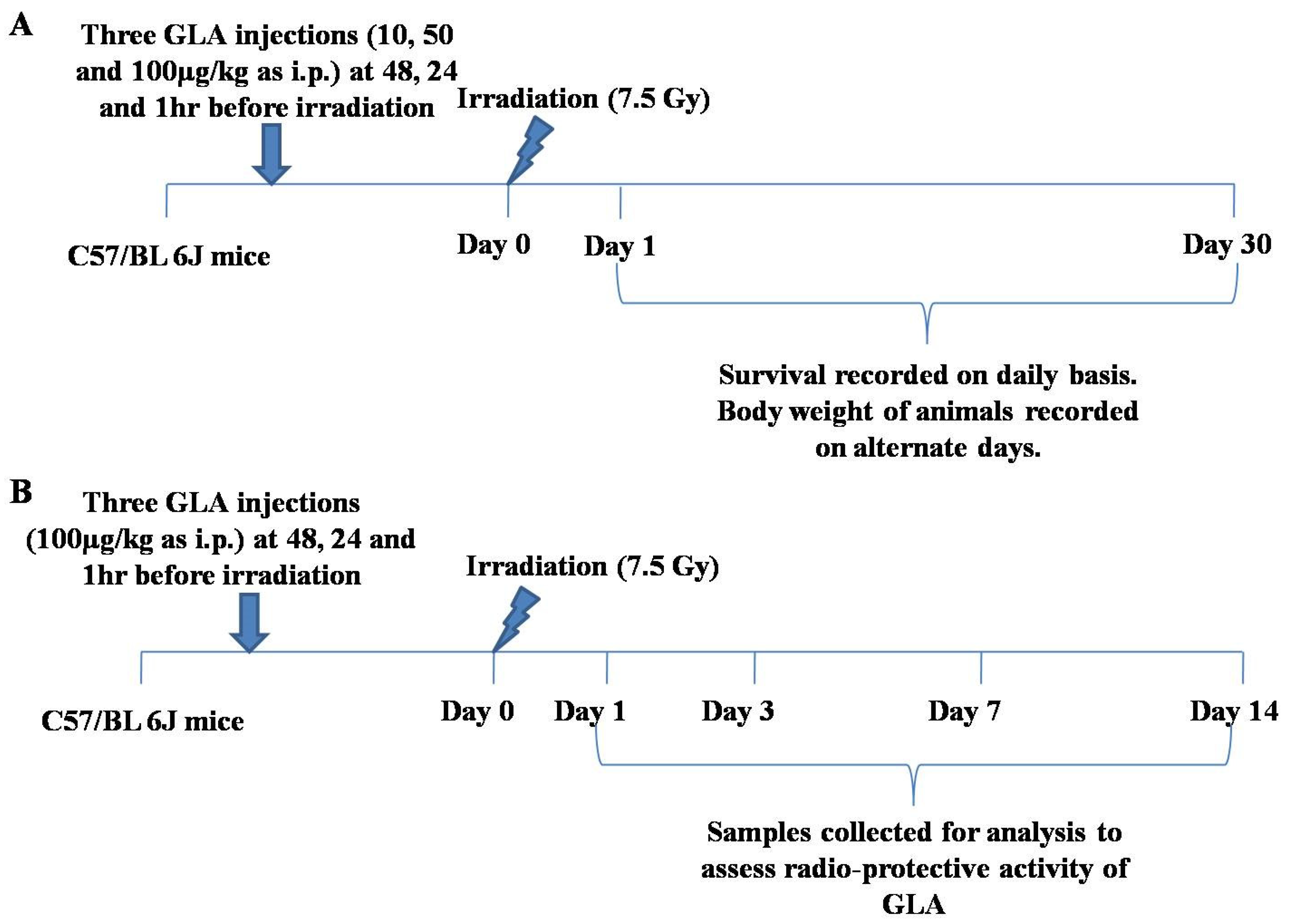
2.5.1. Dose Standardization of GLA for In Vivo Studies
2.5.2. Survival Study for Radio-Protection with PUFAs and LXA4
2.5.3. Mechanism of GLA-Mediated Radio-Protection Studies
- Antioxidant status in mouse duodenum
- b.
- Measurement of inflammatory and anti-inflammatory mediators in duodenum
- c.
- Gene expression studies on mouse duodenum
- d.
- Protein expression studies on mouse duodenum
- e.
- Histopathology of duodenum
2.6. Statistical Analysis
3. Results
3.1. In Vitro Studies
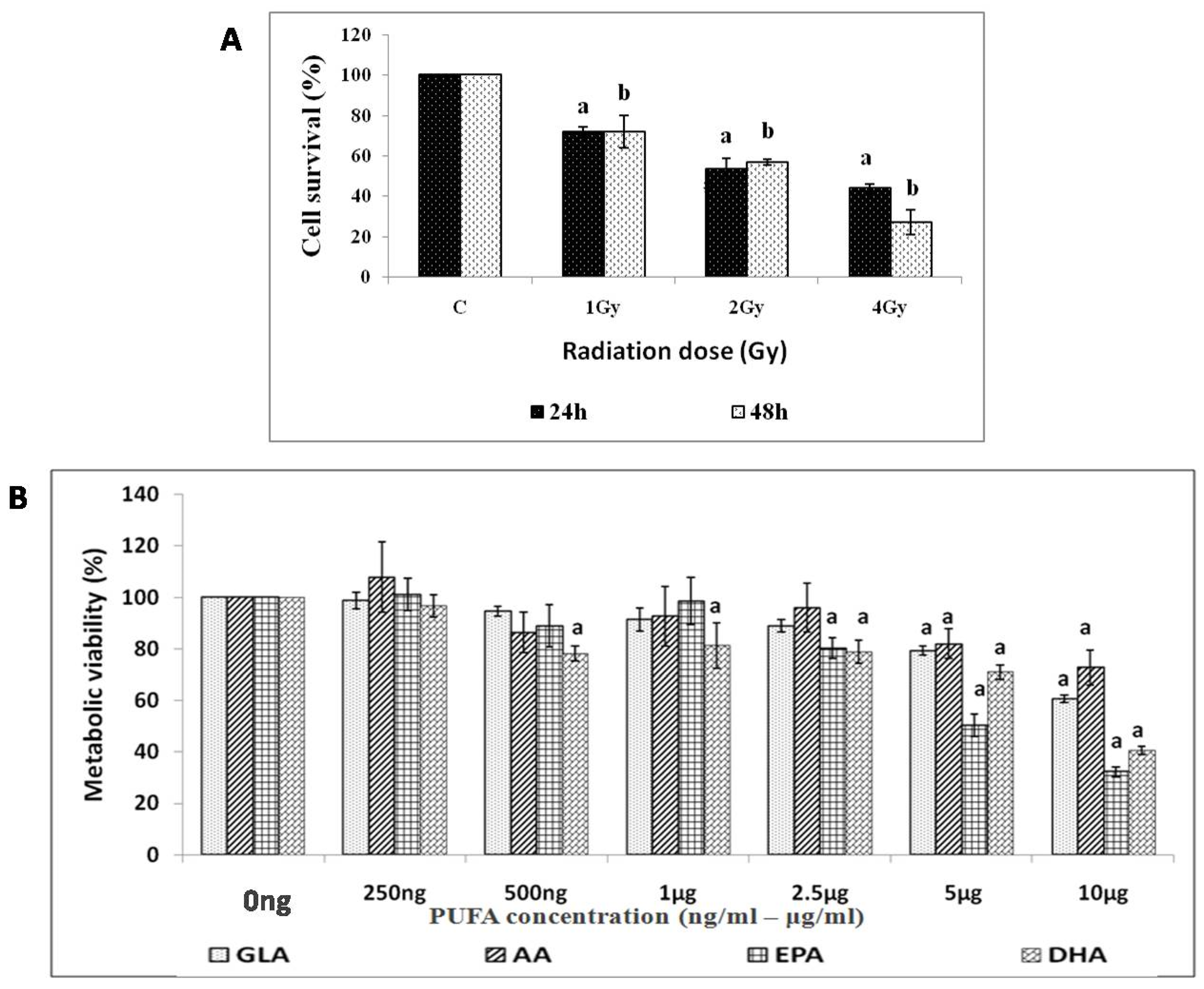
3.1.1. Effect of Radiation on RAW 264.7 Cells
3.1.2. Effect of PUFAs on Viability of RAW 264.7 Cells
3.1.3. GLA Protects RAW 264.7 Cells from Radiation-Induced Toxicity
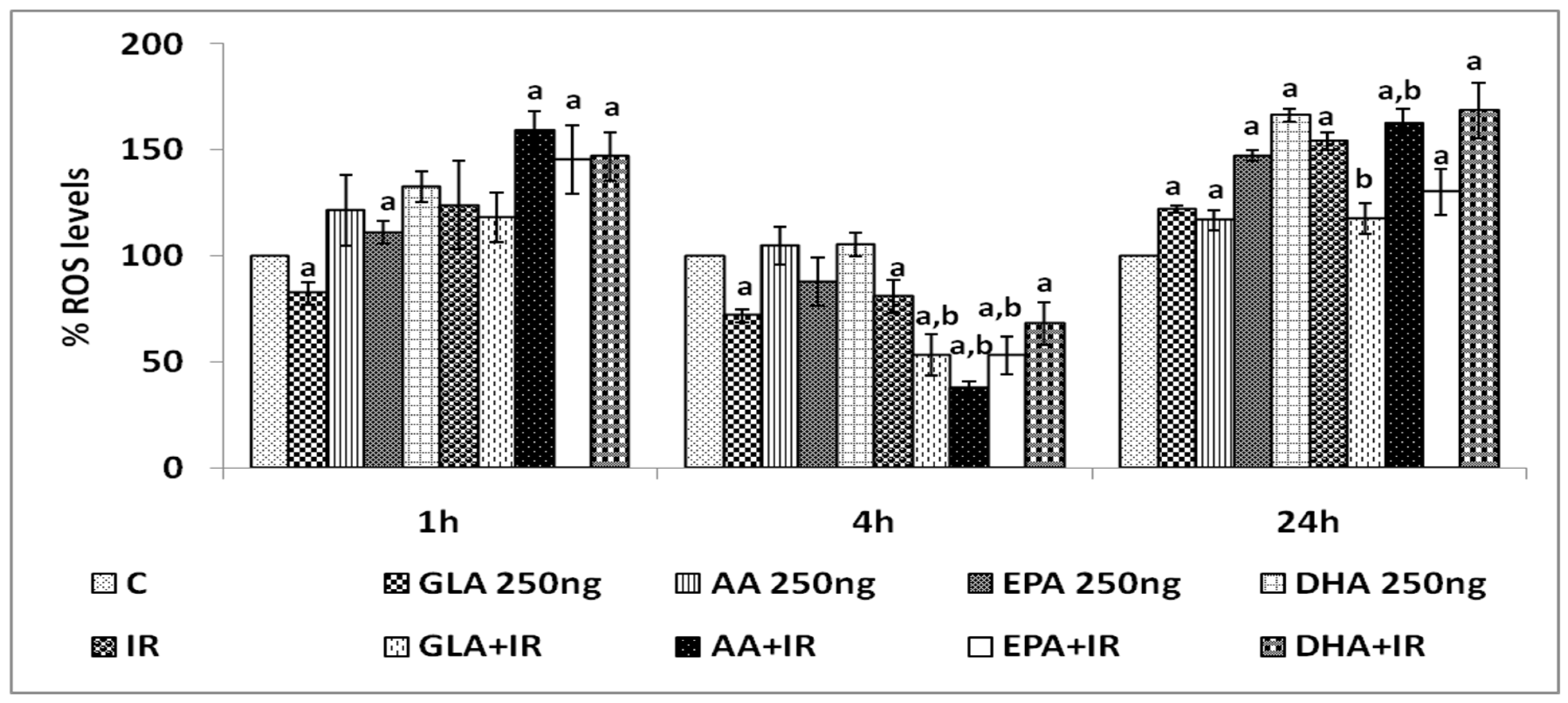
3.1.4. Effect of PUFAs on Intracellular Reactive Oxygen Species (ROS)
3.1.5. Effect of GLA on Antioxidant Enzymes, Lipid Peroxides, and Nitric Oxide (NO)
3.1.6. GLA Decreased Radiation-Induced Cytogenetic Damage
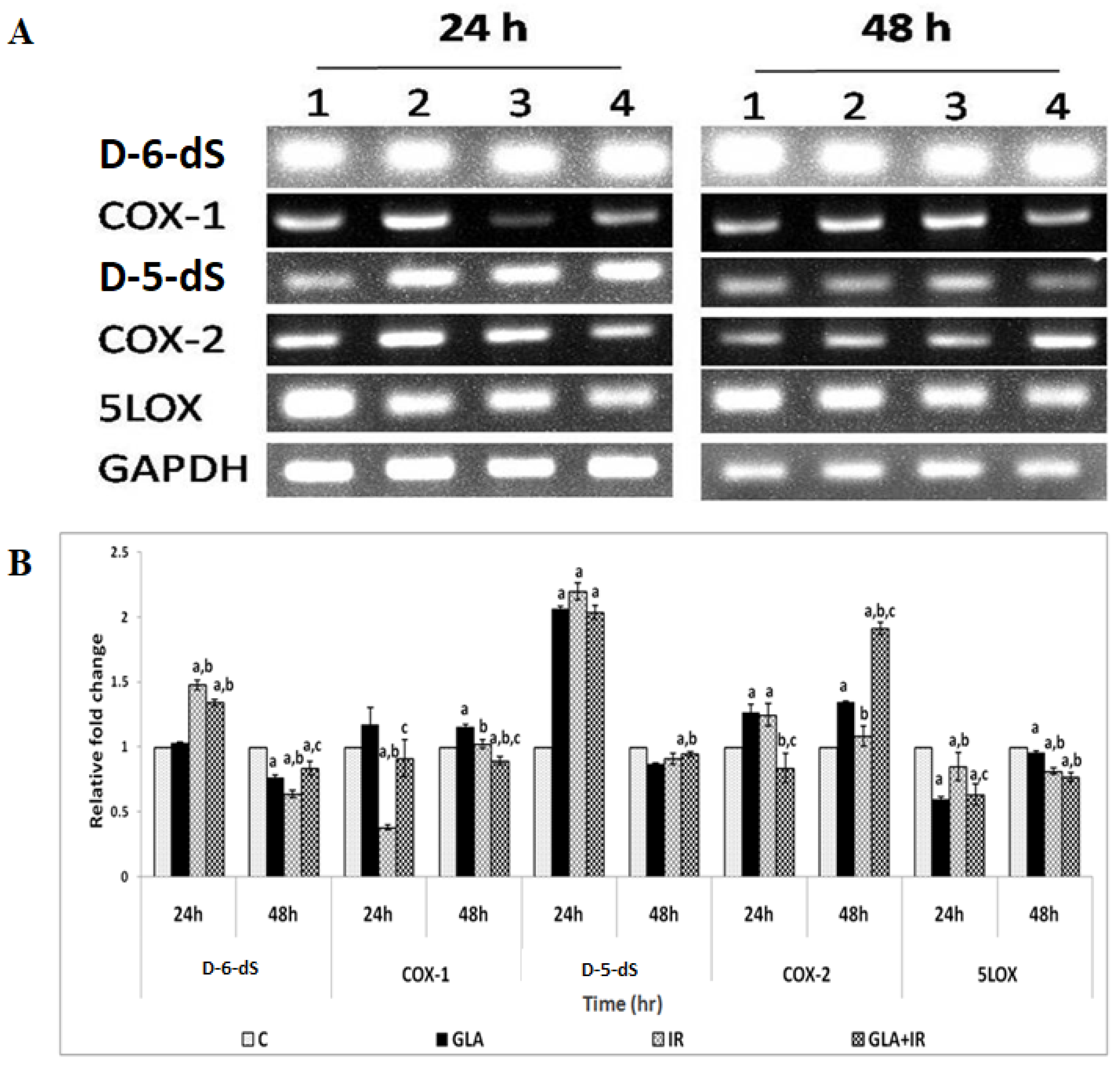
3.1.7. PUFAs Modify Radiation-Induced Changes in Gene(s) Expression
3.2. In Vivo Studies
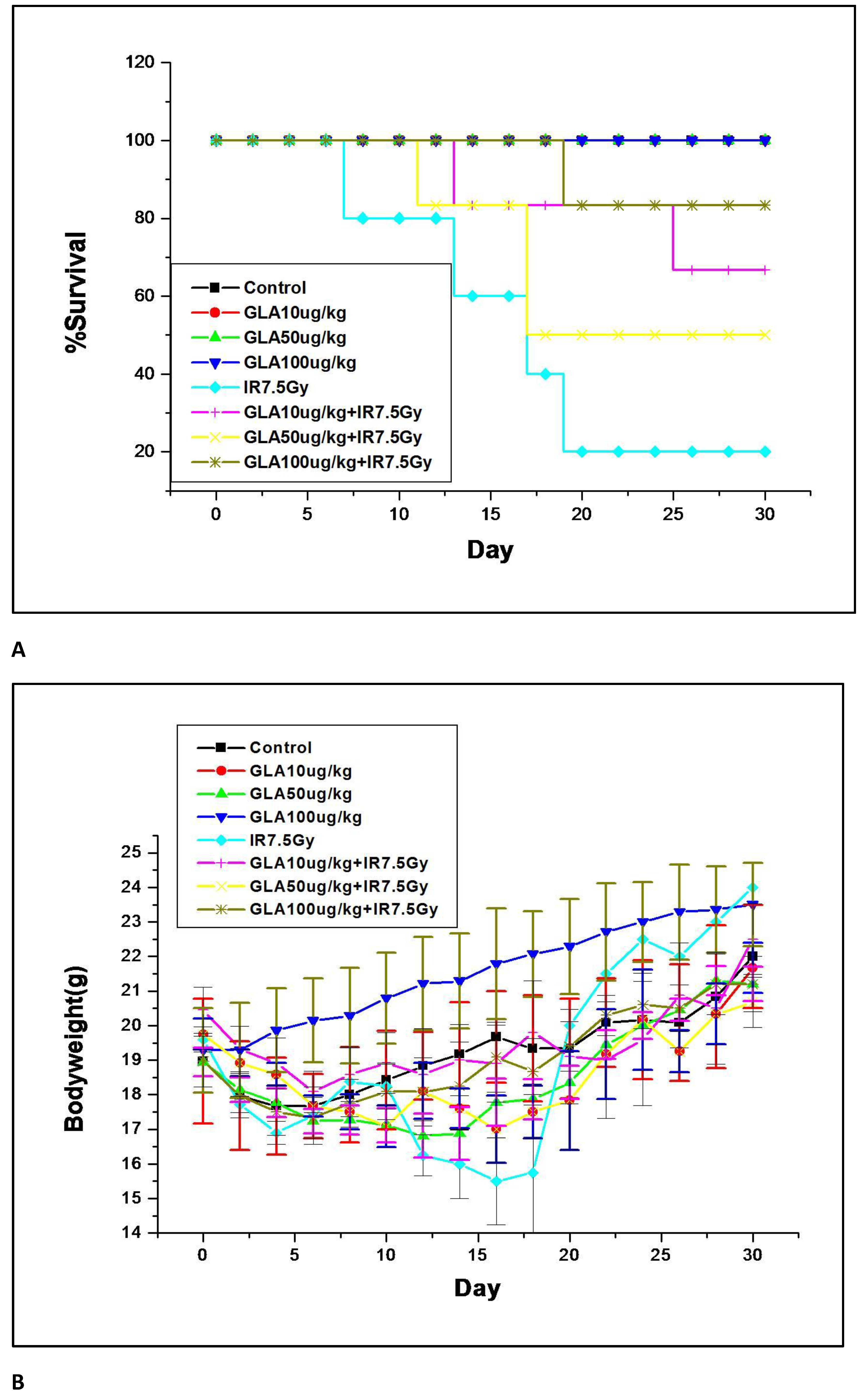
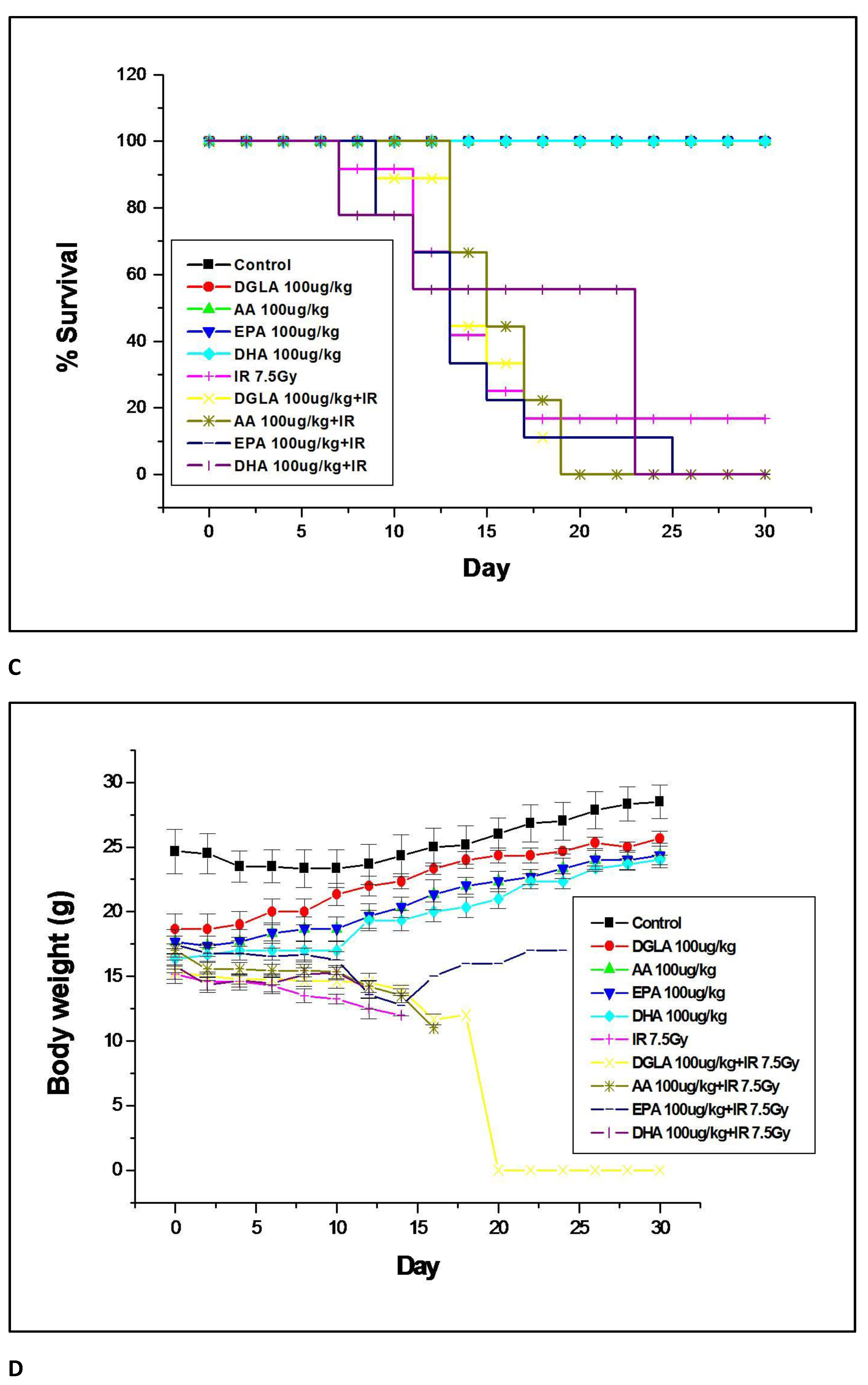
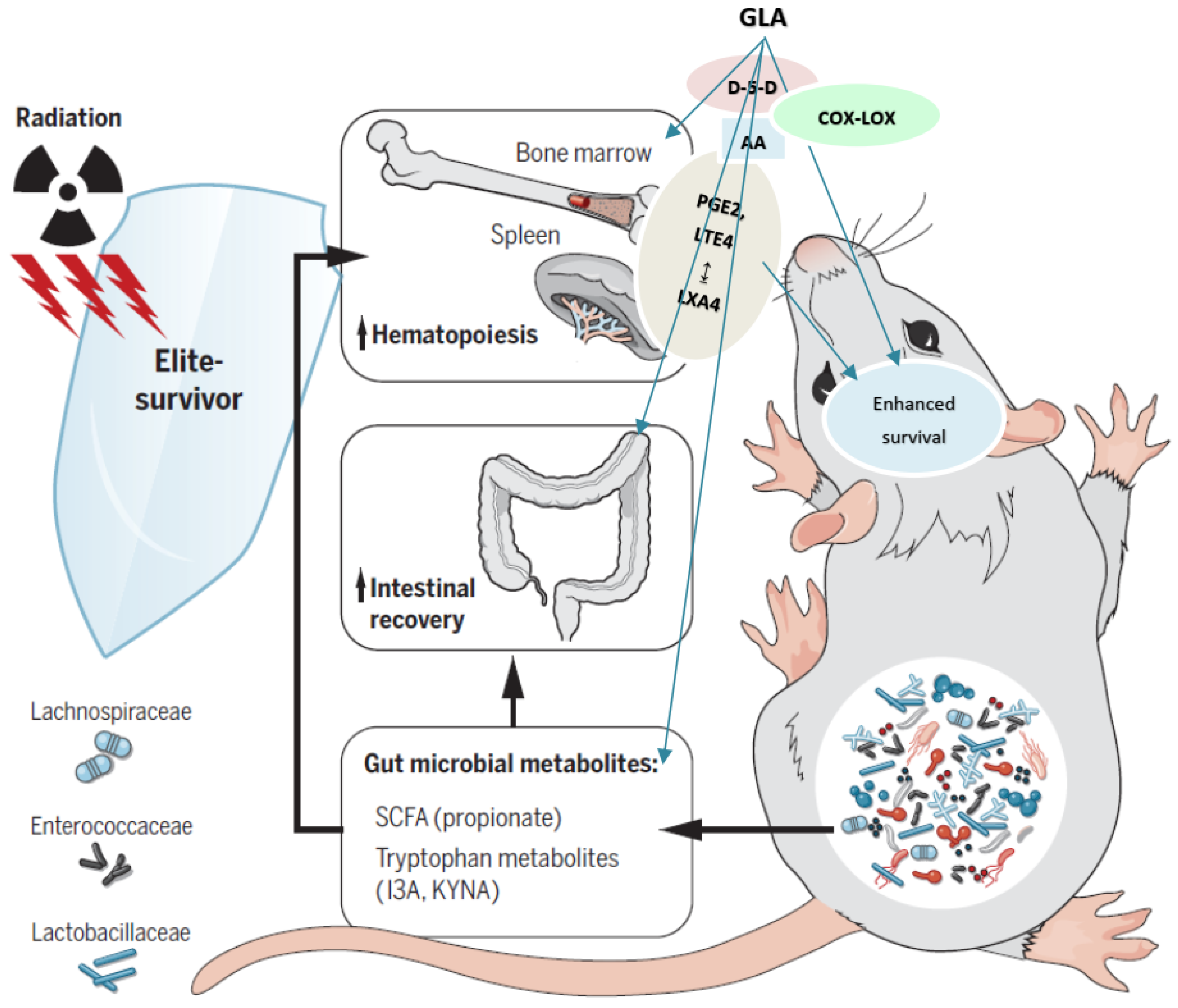
3.2.1. GLA Protects Mice against Whole-Body Lethal Radiation
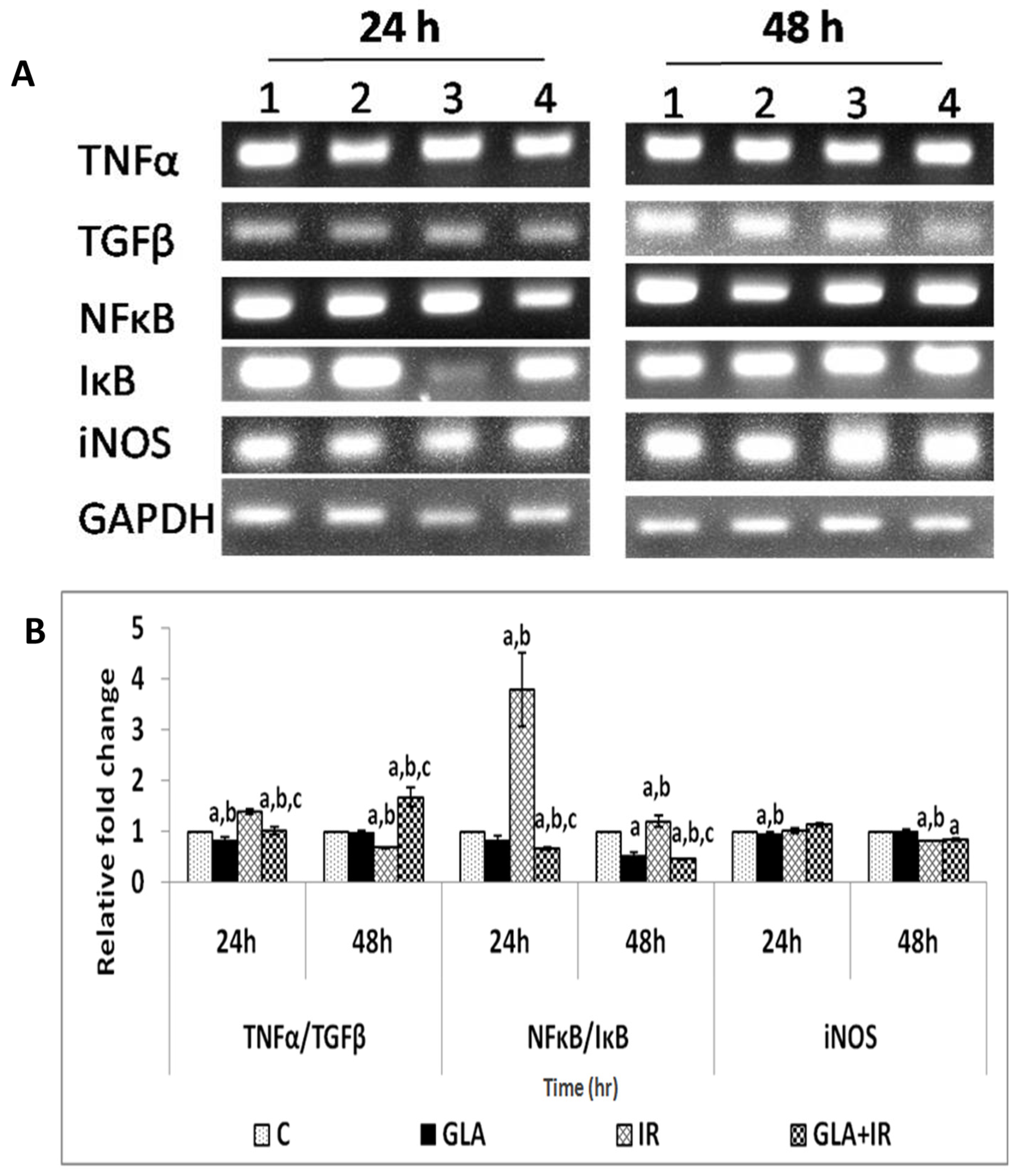
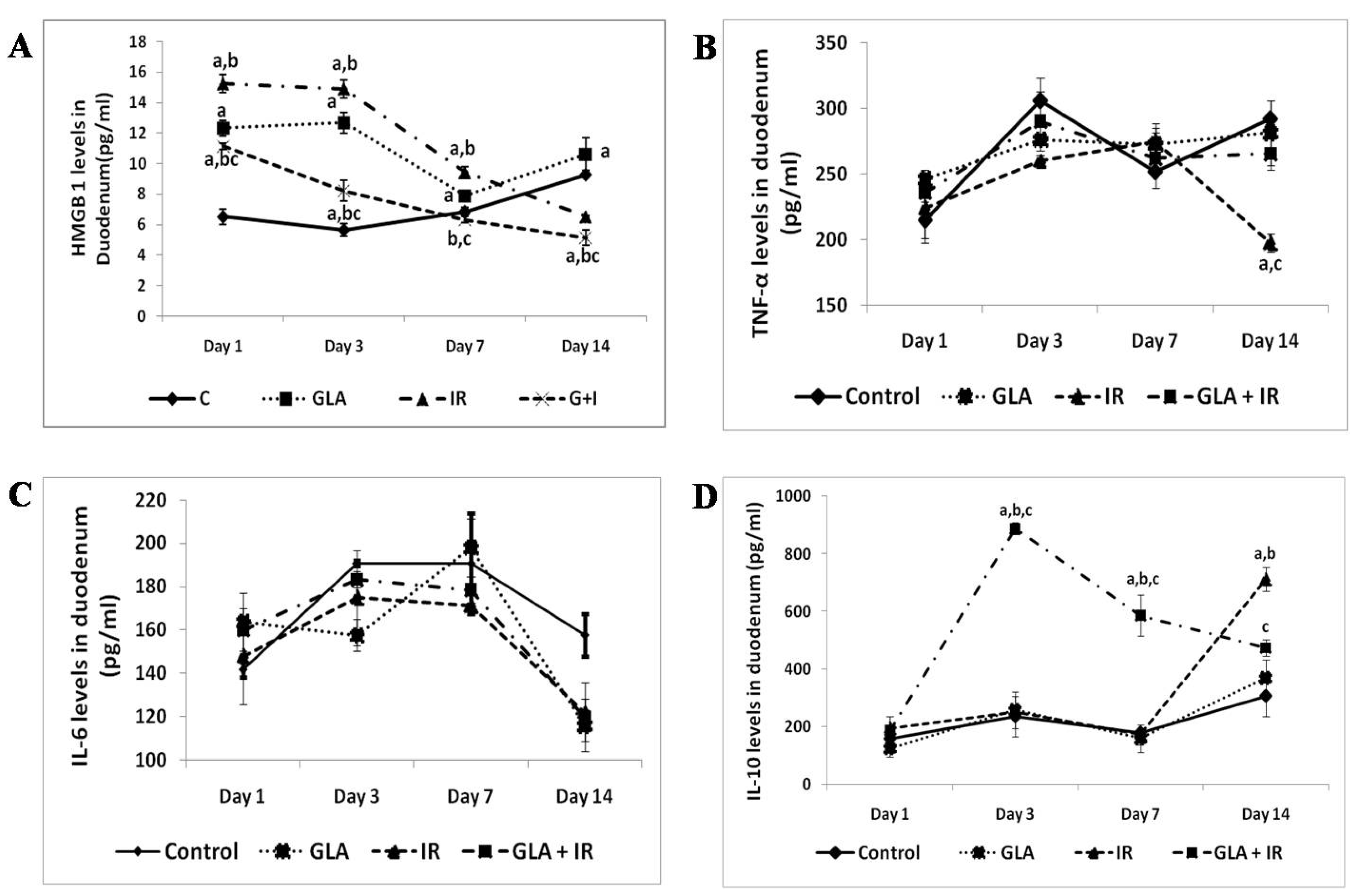
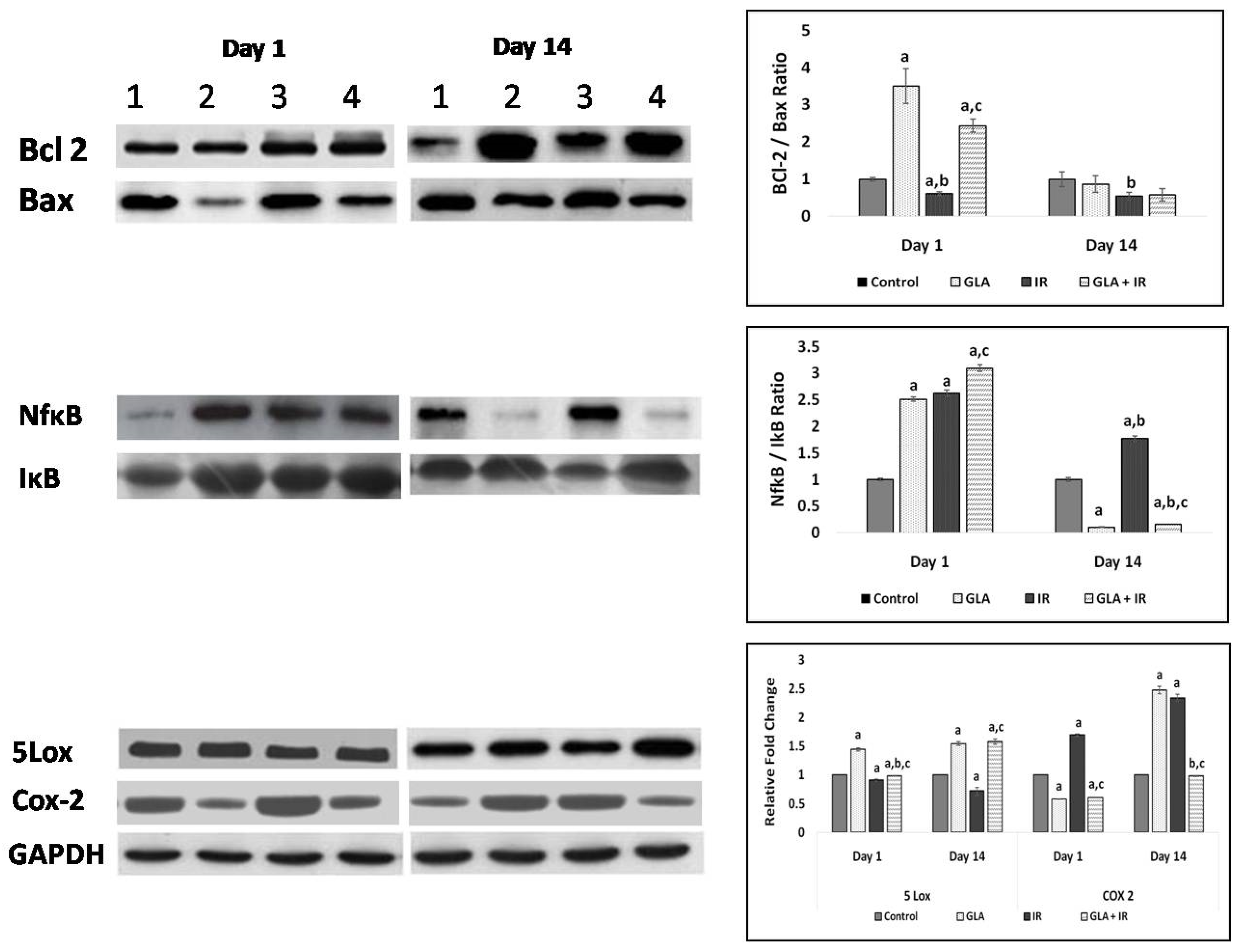
3.2.2. Effect of GLA and Radiation on Cytokines in Mouse Duodenum
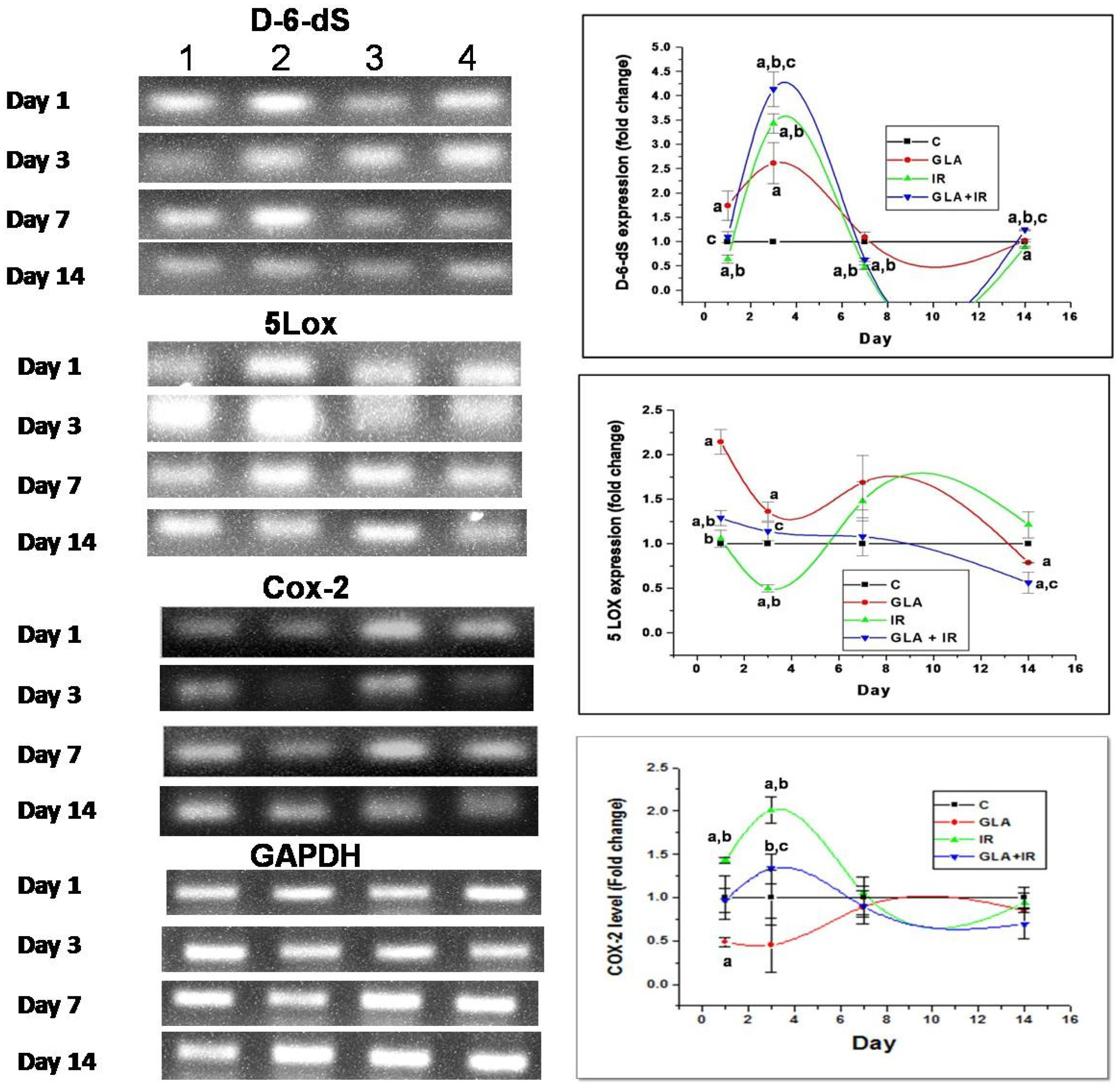
3.2.3. Changes in EFA Metabolism-Associated Genes in Mouse Duodenum
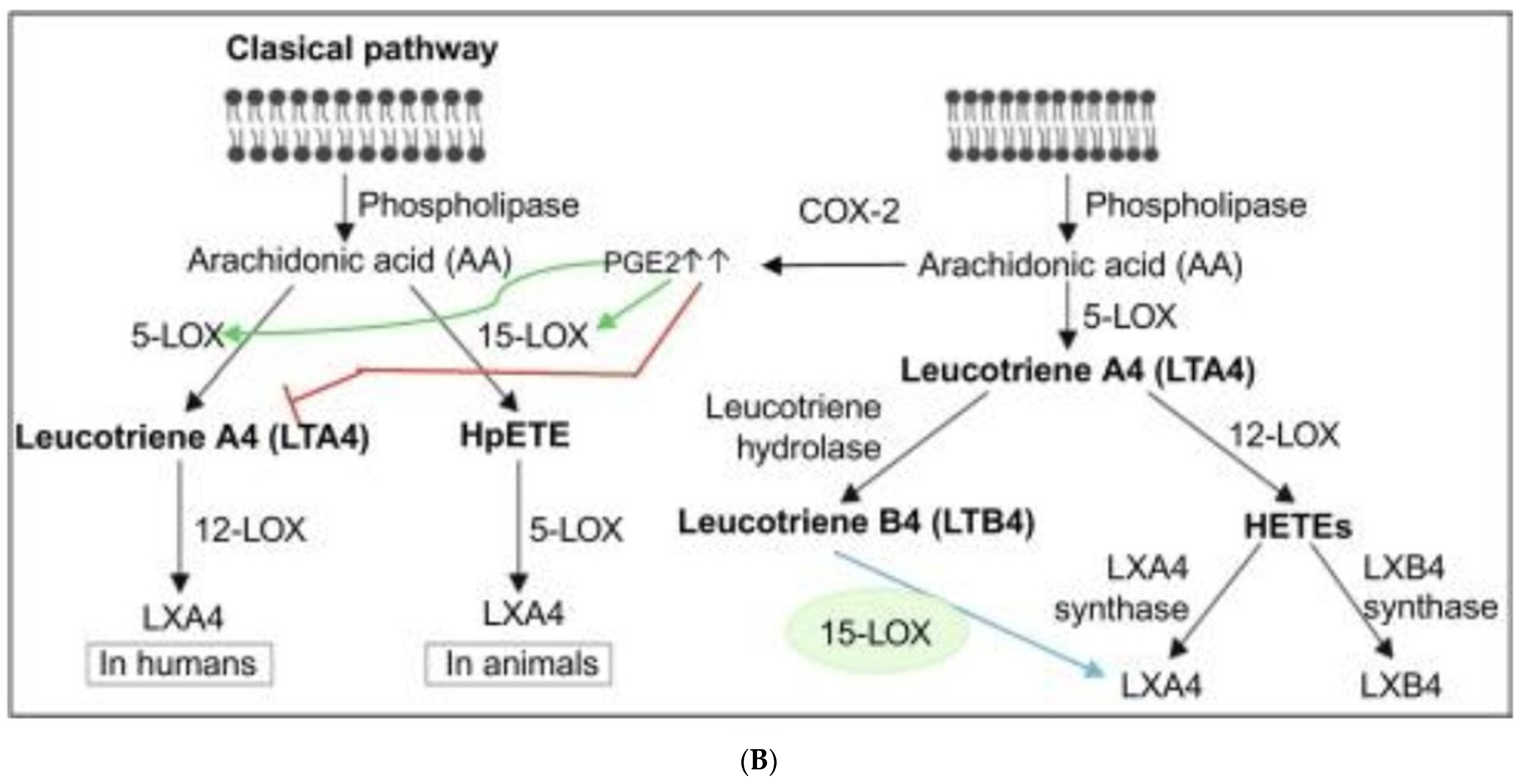
3.2.4. Genes of EFA Metabolic Pathway
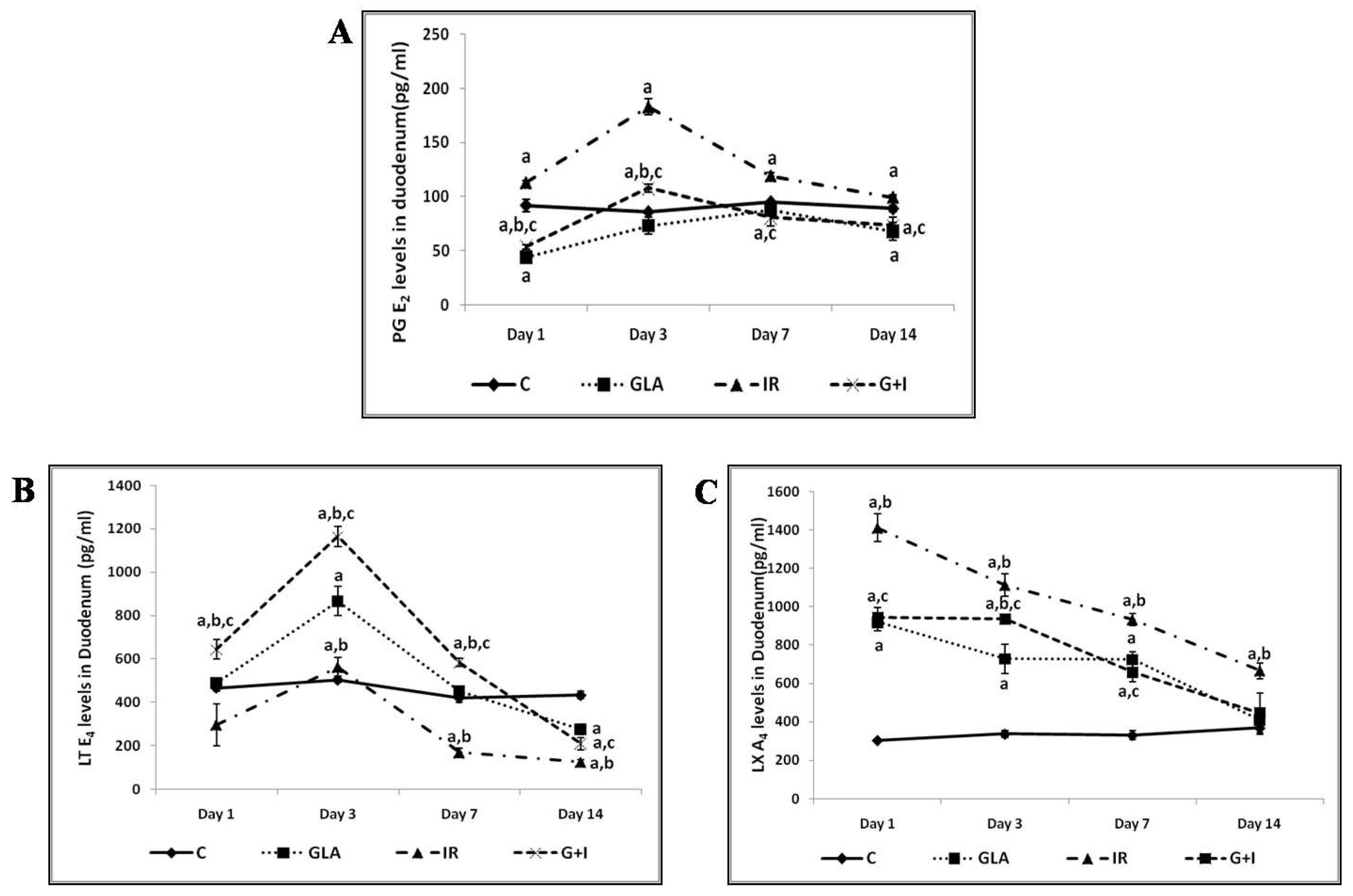
3.2.5. Effect of GLA on Radiation-Induced Changes in Eicosanoids
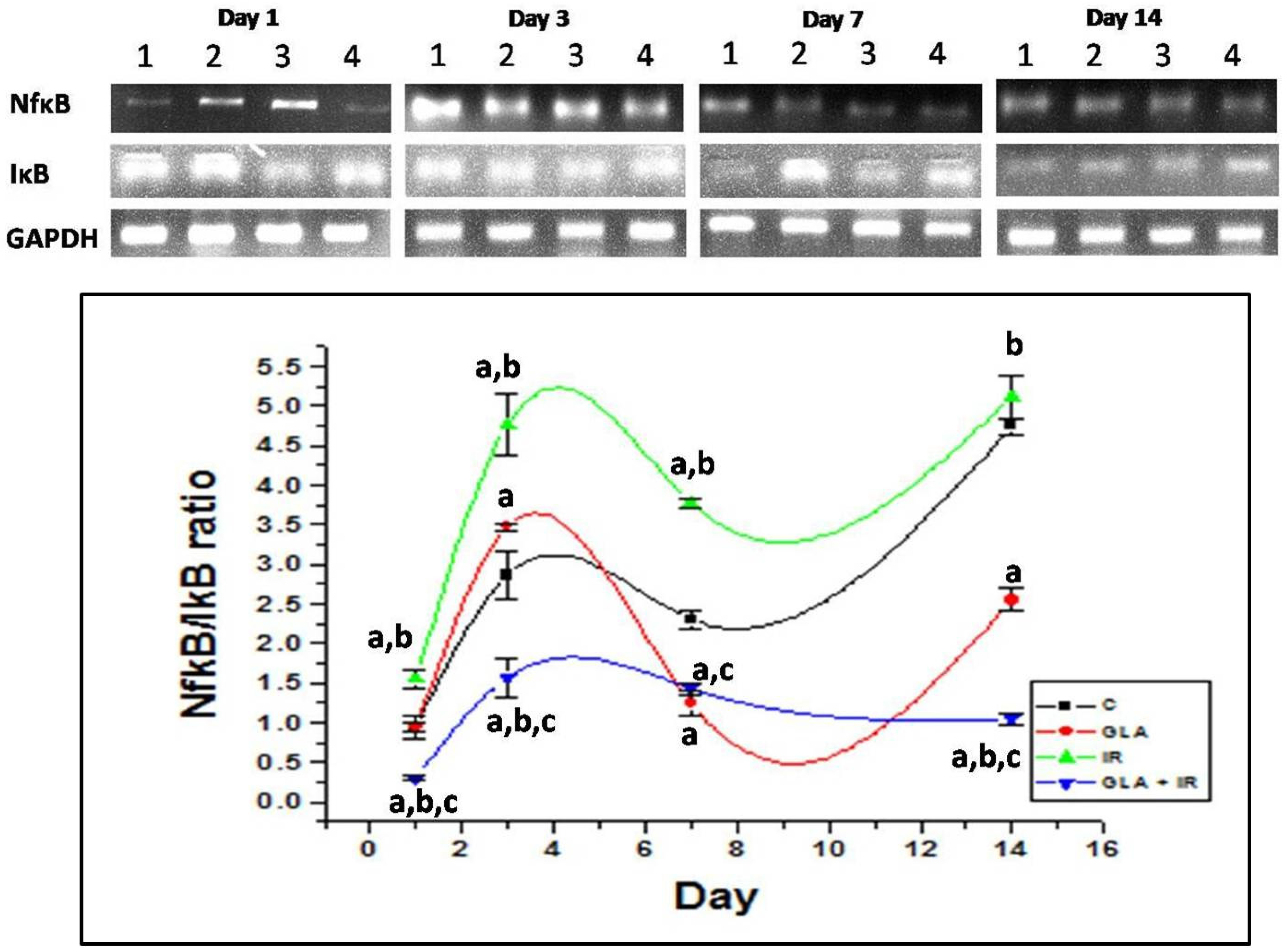
3.2.6. Genes Associated with Inflammation
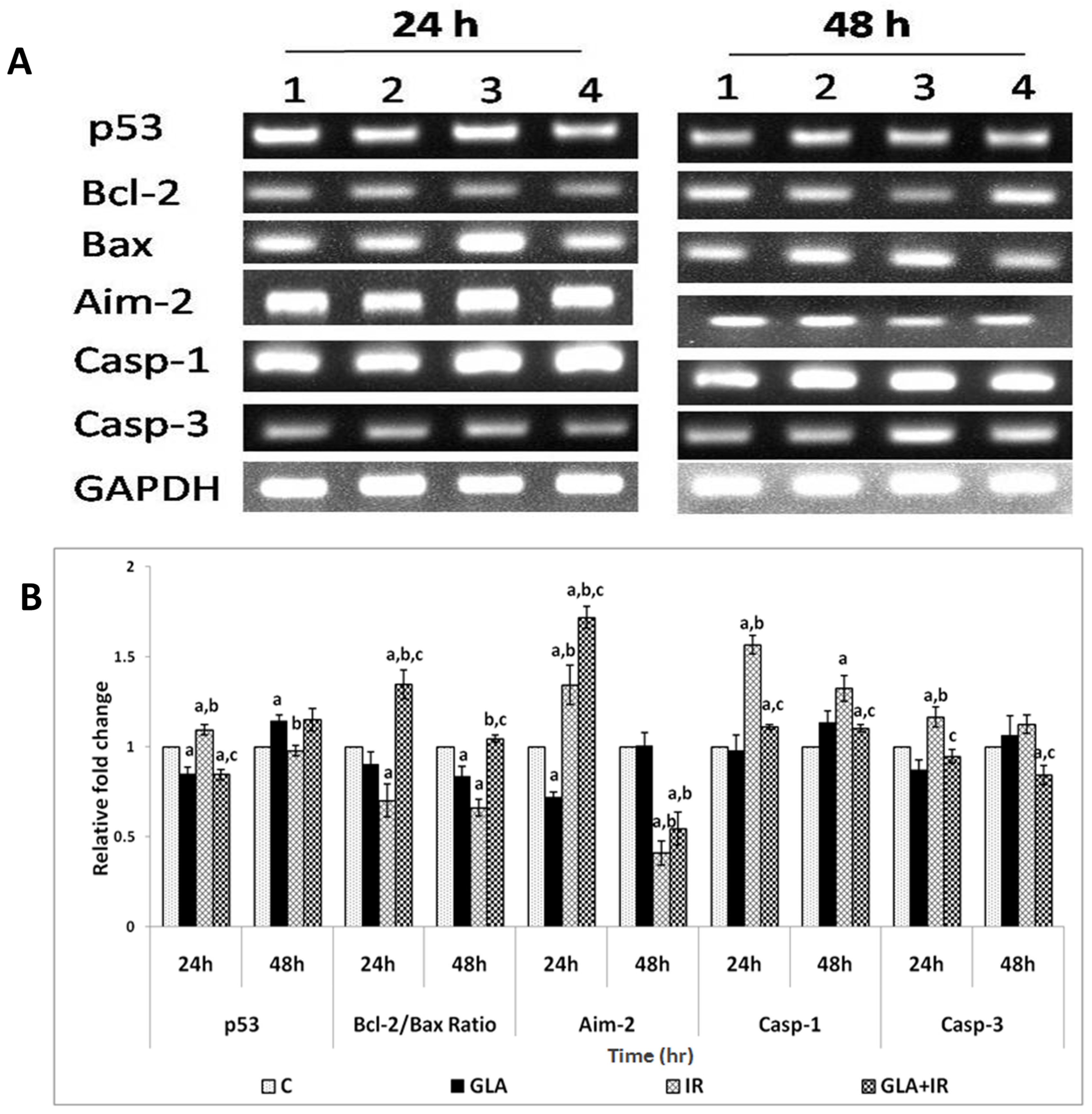
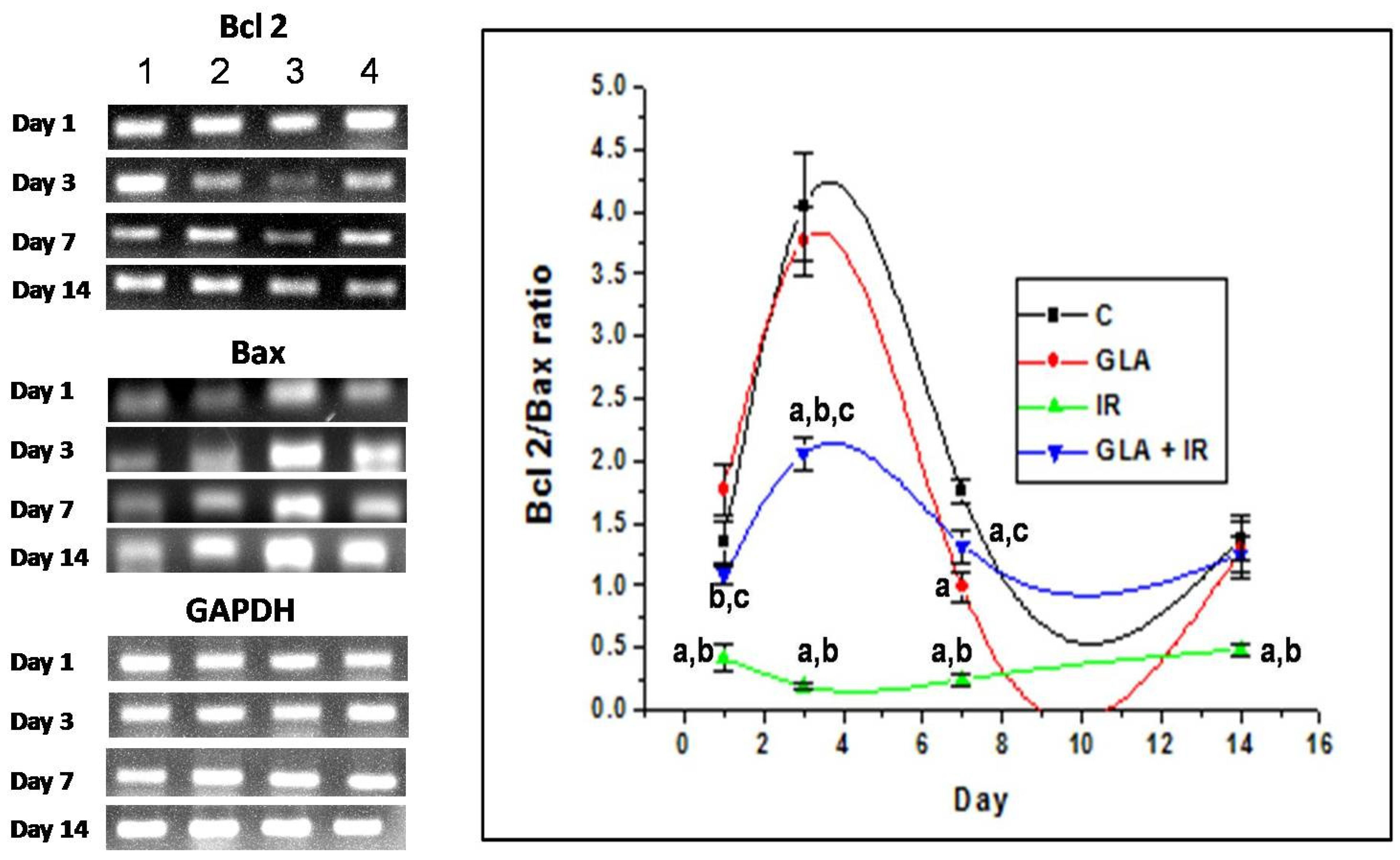
3.2.7. Genes Associated with Apoptotic Pathways
3.2.8. Protein Expression Studies on Mouse Duodenum
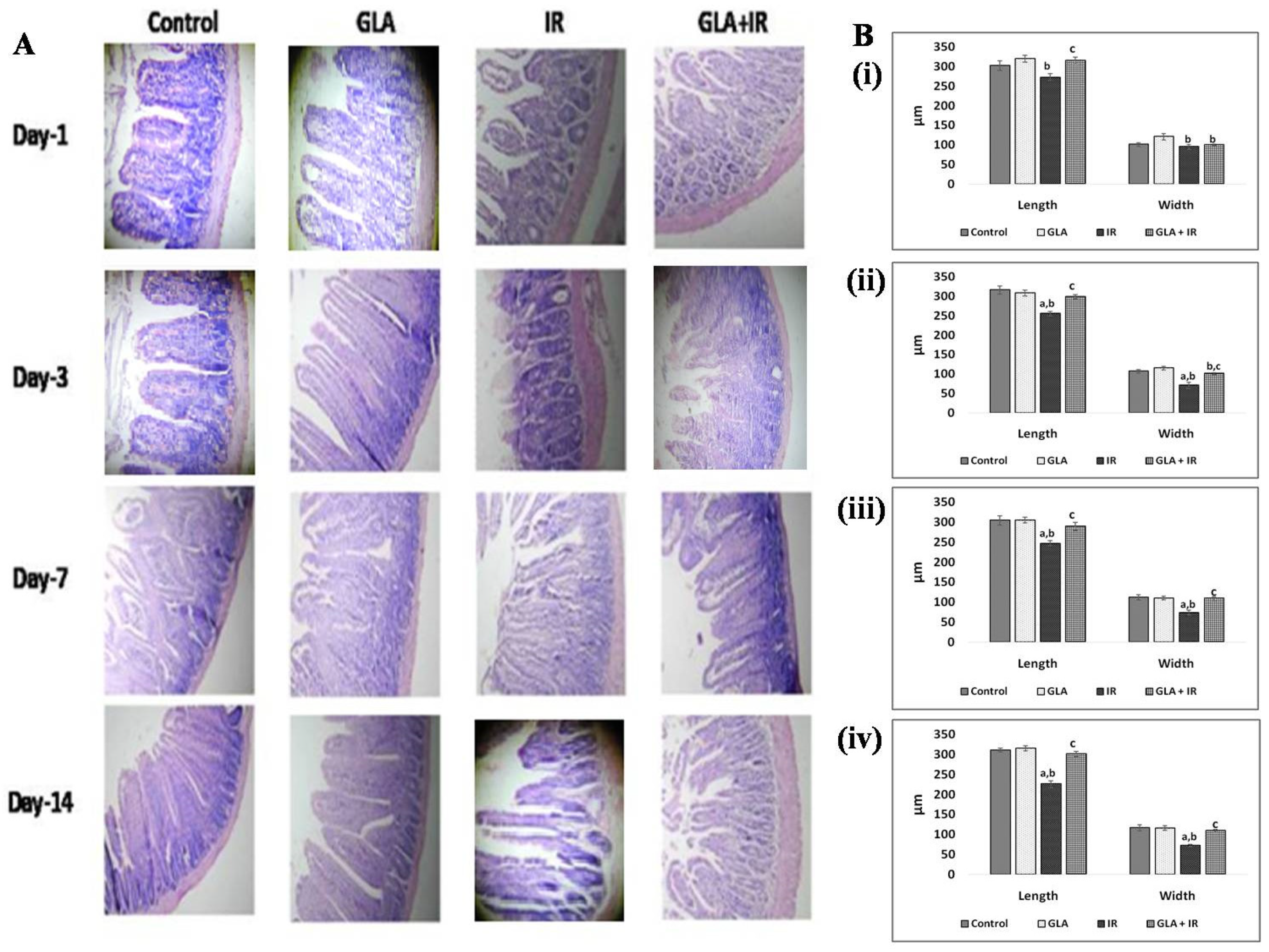
3.2.9. Histopathological Analysis of Mouse Duodenum
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Molecular Weights of Fatty Acids and Their Metabolites Mentioned in the Present Study
- Linoleic acid (LA, 18:2 n-6)-Mol. Wt. 280.4472.
- Gamma-linolenic acid (GLA, 18:3 n-6)-Mol. Wt. 278.43.
- Di-homo-gamma-linoleic acid (DGLA, 20:3 n-6)-Mol. Wt. 306.482.
- Arachidonic acid (AA, 20:4 n-6)-Mol. Wt. 304.47.
- Alpha-linolenic acid (ALA, 18:3 n-3)-Mol. Wt. 278.43.
- Eicosapentaenoic acid (EPA, 20:5 n-3)-Mol. Wt. 302.451.
- Docosahexaenoic acid (DHA, 22:6 n-3)-Mol. Wt. 328.488.
- Prostaglandin E1 (PGE1)-Mol. Wt. 354.481.
- Prostaglandin E2 (PGE2)-Mol. Wt. 352.4651.
- Leukotriene E4 (LTE4)-Mol. Wt. 439.6.
- Lipoxin A4 (LXA4)-Mol. Wt. 352.5.
References
- Sia, J.; Szmyd, R.; Hau, E.; Gee, H.E. Molecular Mechanisms of Radiation-Induced Cancer Cell Death: A Primer. Front. Cell Dev. Biol. 2020, 8, 41. [Google Scholar] [CrossRef]
- Steel, L.K.; Hughes, H.N.; Walden, T.L., Jr. Quantitative, functional and biochemical alterations in the peritoneal cells of mice exposed to whole-body gamma-irradiation. I. Changes in cellular protein, adherence properties and enzymatic activities associated with platelet-activating factor formation and inactivation, and arachidonate metabolism. Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 1988, 53, 943–964. [Google Scholar] [PubMed]
- Bito, L.Z.; Klein, E.M. The role of the arachidonic acid cascade in the species-specific X-ray-induced inflammation of the rabbit eye. Investig. Ophthalmol. Vis. Sci. 1982, 22, 579–587. [Google Scholar]
- Martinez, R.M.; Fattori, V.; Saito, P.; Melo, C.B.P.; Borghi, S.M.; Pinto, I.C.; Bussmann, A.J.C.; Baracat, M.M.; Georgetti, S.R.; Verri, W.A., Jr.; et al. Lipoxin A4 inhibits UV radiation-induced skin inflammation and oxidative stress in mice. J. Dermatol. Sci. 2018, S0923–S1811, 30201–30209. [Google Scholar] [CrossRef] [Green Version]
- da-Palma-Cruz, M.; da Silva, R.F.; Monteiro, D.; Rehim, H.M.M.A.; Grabulosa, C.C.; de Oliveira, A.P.L.; Lino-Dos-Santos-Franco, A. Photo biomodulation modulates the resolution of inflammation during acute lung injury induced by sepsis. Lasers Med. Sci. 2019, 34, 191–199. [Google Scholar] [CrossRef]
- Cohen, D.; DeLeo, V.A. Ultraviolet radiation-induced phospholipase A2 activation occurs in mammalian cell membrane preparations. Photochem. Photobiol. 1993, 57, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Hanson, D.; DeLeo, V. Long-wave ultraviolet light induces phospholipase activation in cultured human epidermal keratinocytes. J. Investig. Dermatol. 1990, 95, 158–163. [Google Scholar] [CrossRef] [Green Version]
- Hanson, D.L.; DeLeo, V.A. Long wave ultraviolet radiation stimulates arachidonic acid release and cyclooxygenase activity in mammalian cells in culture. Photochem. Photobiol. 1989, 49, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Poorani, R.; Bhatt, A.N.; Dwarakanath, B.S.; Das, U.N. COX-2, aspirin and metabolism of arachidonic, eicosapentaenoic and docosahexaenoic acids and their physiological and clinical significance. Eur. J. Pharmacol. 2016, 785, 116–132. [Google Scholar] [CrossRef]
- Cao, Y.; Guan, Y.; Xu, Y.Y.; Hao, C.M. Endothelial prostacyclin protects the kidney from ischemia-reperfusion injury. Pflugers. Arch. 2019, 471, 543–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, H.; Cheng, L.; Langenbach, R.; Ju, C. Prostaglandin I(2) and E(2) mediate the protective effects of cyclooxygenase-2 in a mouse model of immune-mediated liver injury. Hepatology 2007, 45, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, M.; Lamb, J.; Ebert, B.L.; Lynch, M.; Neil, C.; Schmidt, E.; Golub, T.R.; Iliopoulos, O. The connectivity map links iron regulatory protein-1-mediated inhibition of hypoxia-inducible factor-2a translation to the anti-inflammatory 15-deoxy-delta12,14-prostaglandin J2. Cancer Res. 2010, 70, 3071–3079. [Google Scholar] [CrossRef] [Green Version]
- Jung, W.K.; Lee, C.M.; Lee, D.S.; Na, G.; Lee, D.Y.; Choi, I.; Park, S.G.; Seo, S.K.; Yang, J.W.; Choi, J.S.; et al. The 15-deoxy-δ12,14-prostaglandin J2 inhibits LPS-stimulated inflammation via enhancement of the platelet-activating factor acetylhydrolase activity in human retinal pigment epithelial cells. Int. J. Mol. Med. 2014, 33, 449–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, T.; Ohara, O.; Teraoka, H.; Arita, H. Glucocorticoids suppress group II phospholipase A2 production by blocking mRNA synthesis and post-transcriptional expression. J. Biol. Chem. 1990, 265, 12745–12748. [Google Scholar] [CrossRef]
- Das, U.N. “Cell Membrane Theory of Senescence” and the Role of Bioactive Lipids in Aging, and Aging Associated Diseases and Their Therapeutic Implications. Biomolecules 2021, 11, 241. [Google Scholar] [CrossRef]
- Das, U.N. Current and emerging strategies for the treatment and management of systemic lupus erythematosus based on molecular signatures of acute and chronic inflammation. J. Inflamm. Res. 2010, 3, 143–170. [Google Scholar] [CrossRef] [Green Version]
- Das, U.N. Bioactive Lipids in Age-Related Disorders. Adv. Exp. Med. Biol. 2020, 1260, 33–83. [Google Scholar] [PubMed]
- Loynes, C.A.; Lee, J.A.; Robertson, A.L.; Steel, M.J.; Ellett, F.; Feng, Y.; Levy, B.D.; Whyte, M.K.B.; Renshaw, S.A. PGE2 production at sites of tissue injury promotes an anti-inflammatory neutrophil phenotype and determines the outcome of inflammation resolution in vivo. Sci. Adv. 2018, 4, eaar8320. [Google Scholar] [CrossRef] [Green Version]
- Tang, E.H.; Libby, P.; Vanhoutte, P.M.; Xu, A. Anti-inflammation therapy by activation of prostaglandin EP4 receptor in cardiovascular and other inflammatory diseases. J. Cardiovasc. Pharmacol. 2012, 59, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Walden, T.L., Jr.; Patchen, M.; Snyder, S.L. 16,16-Dimethyl prostaglandin E2 increases survival in mice following irradiation. Radiat. Res. 1987, 109, 440–448. [Google Scholar] [CrossRef]
- Walden, T.L., Jr.; Farzaneh, N.K. Radioprotection by 16,16 dimethyl prostaglandin E2 is equally effective in male and female mice. J. Radiat. Res. 1995, 36, 1–7. [Google Scholar] [CrossRef]
- Walden, T.L., Jr. Leukotriene C4-induced radioprotection: The role of hypoxia. Radiat. Res. 1992, 132, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Hanson, W.R.; Ainsworth, E.J. 16,16-Dimethyl prostaglandin E2 induces radioprotection in murine intestinal and hematopoietic stem cells. Radiat. Res. 1985, 103, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Walden, T.L., Jr.; Patchen, M.L.; MacVittie, T.J. Leukotriene-induced radioprotection of hematopoietic stem cells in mice. Radiat. Res. 1988, 113, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Walden, T.L., Jr. Radioprotection of mouse hematopoietic stem cells by leukotriene A4 and lipoxin B4. J. Radiat. Res. 1988, 29, 255–260. [Google Scholar] [CrossRef]
- Zhang, Y.; Desai, A.; Yang, S.Y.; Bae, K.B.; Antczak, M.I.; Fink, S.P.; Tiwari, S.; Willis, J.E.; Williams, N.S.; Dawson, D.M.; et al. TISSUE REGENERATION. Inhibition of the prostaglandin-degrading enzyme 15-PGDH potentiates tissue regeneration. Science 2015, 348, aaa2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palla, A.R.; Ravichandran, M.; Wang, Y.X.; Alexandrova, L.; Yang, A.V.; Kraft, P.; Holbrook, C.A.; Schürch, C.M.; Ho, A.T.V.; Blau, H.M. Inhibition of prostaglandin-degrading enzyme 15-PGDH rejuvenates aged muscle mass and strength. Science. 2021, 371, eabc8059. [Google Scholar] [CrossRef] [PubMed]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. Curr. Protoc. Immunol. 2015, 111, A3.B.1–A3.B.3. [Google Scholar] [CrossRef]
- Vistica, V.T.; Skehan, P.; Scudiero, D.; Monks, A.; Pittman, A.; Boyd, M. Tetrazolium-based assays for cellular viability: A critical examination of selected parameters affecting formazan production. Cancer Res. 1991, 51, 2515–2520. [Google Scholar]
- Reula, A.; Pellicer, D.; Castillo, S.; Magallón, M.; Armengot, M.; Herrera, G.; O’Connor, J.E.; Bañuls, L.; Navarro-García, M.M.; Escribano, A.; et al. New Laboratory Protocol to Determine the Oxidative Stress Profile of Human Nasal Epithelial Cells Using Flow Cytometry. J. Clin. Med. 2021, 10, 1172. [Google Scholar] [CrossRef]
- Naveen, K.V.G.; Naidu, V.G.M.; Das, U.N. Arachidonic acid and lipoxin A4 attenuate streptozotocin-induced cytotoxicity to RIN5F cells in vitro and type 1 and type 2 diabetes mellitus in vivo. Nutrition 2017, 35, 61–80. [Google Scholar]
- Polavarapu, S.; Mani, A.M.; Gundala, N.K.; Hari, A.D.; Bathina, S.; Das, U.N. Effect of polyunsaturated fatty acids and their metabolites on bleomycin-induced cytotoxic action on human neuroblastoma cells in vitro. PLoS ONE 2014, 9, e114766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, M. The micronucleus test—most widely used in vivo genotoxicity test. Genes Environ. 2016, 38, 18. [Google Scholar] [CrossRef] [Green Version]
- Das, U.N.; Ramadevi, G.; Rao, K.P.; Rao, M.S. Prostaglandins and their precursors can modify genetic damage-induced by gamma-radiation and benzo(a)pyrene. Prostaglandins 1985, 29, 911–920. [Google Scholar] [CrossRef]
- Devi, G.R.; Das, U.N.; Rao, K.P.; Rao, M.S. Prostaglandins and mutagenesis: Prevention and/or reversibility of genetic damage induced by benzo (a) pyrene in the bone marrow cells of mice by prostaglandins El. Prostaglandins Leukot. Med. 1984, 15, 287–292. [Google Scholar]
- Devi, G.R.; Das, U.N.; Rao, K.P.; Rao, M.S. Prostaglandins and mutagenesis: Modification of phenytoin-induced genetic damage by Prostaglandins in lymphocyte cultures. Prostaglandins Leukot. Med. 1984, 15, 109–113. [Google Scholar]
- Jacobson, L.K.; Johnson, M.B.; Dedhia, R.D.; Niknam-Bienia, S.; Wong, A.K. Impaired wound healing after radiation therapy: A systematic review of pathogenesis and treatment. JPRAS Open 2017, 13, 92–105. [Google Scholar] [CrossRef]
- Dormand, E.L.; Banwell, P.E.; Goodacre, T.E. Radiotherapy and wound healing. Int. Wound J. 2005, 2, 112–127. [Google Scholar] [CrossRef]
- Denham, J.W.; Hauer-Jensen, M. The radiotherapeutic injury—A complex ‘wound’. Radiother Oncol. 2002, 63, 129–145. [Google Scholar] [CrossRef]
- Herskind, C.; Bamberg, M.; Rodemann, H.P. The role of cytokines in the development of normal-tissue reactions after radiotherapy. Strahlenther. Onkol. 1998, 174 (Suppl. 3), 12–15. [Google Scholar]
- Shi, H.P.; Efron, D.T.; Most, D.; Barbul, A. The role of iNOS in wound healing. Surgery 2001, 130, 225. [Google Scholar] [CrossRef]
- Schaffer, M.; Weimer, W.; Wider, S.; Stulten, C.; Bongartz, M.; Budach, W.; Becker, H.D. Differential expression of inflammatory mediators in radiation-impaired wound healing. J. Surg. Res. 2002, 107, 93–100. [Google Scholar] [CrossRef]
- Regan, M.C.; Kirk, S.J.; Hurson, M.; Sodeyama, M.; Wasserkrug, H.L.; Barbul, A. Tumor necrosis factor-alpha inhibits in vivo collagen synthesis. Surgery 1993, 113, 173–177. [Google Scholar] [PubMed]
- Miles, R.H.; Paxton, T.P.; Zachais, D.; Dries, D.J.; Gamelli, R.L. Systemic administration of interferon-gamma impairs wound healing. J. Surg. Res. 1994, 56, 288–294. [Google Scholar] [CrossRef]
- Schaffer, M.; Barbul, A. Lymphocyte function in wound healing and following injury. Br. J. Surg. 1998, 85, 444–460. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Wang, D.; Gao, Y.; Zhou, J.; Peng, R.; Cui, Y.; Xia, G.; Qing, Q.; Yang, H.; Liu, J.; et al. Expression of MMP1 in surgical and radiation-impaired wound healing and its effects on the healing process. J. Environ. Pathol. Toxicol Oncol. 2002, 21, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Poorani, R.; Bhatt, A.N.; Das, U.N. Modulation of pro-inflammatory and pro-resolution mediators by γ-linolenic acid: An important element in radioprotection against ionizing radiation. Arch. Med. Sci. 2020, 29, 1448–1456. [Google Scholar]
- Devi, G.R.; Das, U.N.; Rao, K.P.; Rao, M.S. Prevention of radiation-induced poly-chromatophilia by prostaglandin El and colchicine. IRCS Med. Sci. 1983, 11, 863–864. [Google Scholar]
- Das, U.N.; Devi, G.R.; Rao, K.P.; Rao, M.S. Modification of benzo (a) pyrene induced genetic damage to the bone marrow cells of mice by prostaglandins. IRCS Med. Sci. 1983, 11, 823–824. [Google Scholar]
- Das, U.N.; Devi, G.R.; Rao, K.P.; Rao, M.S. Prevention and/or reversibility of genetic damage induced by diphenylhydantoin to the bone marrow cells of mice by colchicine: Relevance to prostaglandin involvement. IRCS Med. Sci. 1983, 11, 122–123. [Google Scholar]
- Das, U.N. Nutrients, essential fatty acids and prostaglandins interact to augment immune responses and prevent genetic damage and cancer. Nutrition. 1989, 5, 106–110. [Google Scholar] [PubMed]
- Das, U.N.; Ramadevi, G.; Rao, K.P.; Rao, M.S. Prostaglandins can modify gamma-radiation and chemical induced cytotoxicity and genetic damage in vitro and in vivo. Prostaglandins 1989, 38, 689–716. [Google Scholar] [CrossRef]
- Das, U.N.; Rao, K.P. Effect of gamma-linolenic acid and prostaglandins E1 on gamma-radiation and chemical-induced genetic damage to the bone marrow cells of mice. Prostaglandins Leukot. Essent. Fat. Acids 2006, 74, 165–173. [Google Scholar] [CrossRef]
- Das, U.N. Colchicine can prevent and/or reverse mutagenesis: Possible role for prostaglandins. IRCS J. Med. Sci. 1983, 11, 300–301. [Google Scholar]
- Shivani, P.; Rao, K.P.; Chaudhury, J.R.; Ahmed, J.; Rao, B.R.; Kanjilal, S.; Hasan, Q.; Das, U.N. Effect of polyunsaturated fatty acids on diphenyl hydantoin-induced genetic damage in vitro and in vivo. Prostaglandins Leukot. Essen. Fat. Acids 2009, 80, 43–50. [Google Scholar]
- Suresh, Y.; Das, U.N. Protective action of arachidonic acid against alloxan-induced cytotoxicity and diabetes mellitus. Prostaglandins Leuk. Essen. Fat. Acids 2001, 64, 37–52. [Google Scholar] [CrossRef]
- Suresh, Y.; Das, U.N. Differential effect of saturated, monounsaturated, and polyunsaturated fatty acids on alloxan-induced diabetes mellitus. Prostaglandins Leukot. Essen. Fat. Acids 2006, 74, 199–213. [Google Scholar] [CrossRef] [PubMed]
- Suresh, Y.; Das, U.N. Long-chain polyunsaturated fatty acids and chemically induced diabetes mellitus: Effect of omega-6 fatty acids. Nutrition 2003, 19, 93–114. [Google Scholar] [CrossRef]
- Suresh, Y.; Das, U.N. Long-chain polyunsaturated fatty acids and chemically induced diabetes mellitus: Effect of omega-3 fatty acids. Nutrition 2003, 19, 213–228. [Google Scholar] [CrossRef]
- Naveen, K.V.G.; Naidu, V.G.M.; Das, U.N. Arachidonic acid and lipoxin A4 attenuate alloxan-induced cytotoxicity to RIN5F cells in vitro and type 1 diabetes mellitus in vivo. BioFactors 2017, 43, 251–271. [Google Scholar]
- Das, U.N. Tumoricidal action of cis-unsaturated fatty acids and its relationship to free radicals and lipid peroxidation. Cancer Lett. 1991, 56, 235–243. [Google Scholar] [CrossRef]
- Das, U.N. Essential fatty acids, lipid peroxidation and apoptosis. Prostaglandins Leukot. Essent. Fat. Acids 1999, 61, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. Essential fatty acids enhance free radical generation and lipid peroxidation to induce apoptosis of tumor cells. Clin. Lipidol. 2011, 6, 463–489. [Google Scholar] [CrossRef]
- Leaver, H.A.; Bell, H.S.; Rizzo, M.T.; Ironside, J.W.; Gregor, A.; Wharton, S.B.; Whittle, I.R. Antitumour and pro-apoptotic actions of highly unsaturated fatty acids in glioma. Prostaglandins Leukot. Essent. Fat. Acids 2002, 66, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Leaver, H.A.; Wharton, S.B.; Bell, H.S.; Leaver-Yap, I.M.; Whittle, I.R. Highly unsaturated fatty acid induced tumour regression in glioma pharmacodynamics and bioavailability of gamma linolenic acid in an implantation glioma model: Effects on tumour biomass, apoptosis and neuronal tissue histology. Prostaglandins Leukot. Essent. Fat. Acids 2002, 67, 283–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, U.N. Gamma-linolenic acid therapy of human glioma-a review of in vitro, in vivo, and clinical studies. Med. Sci. Monit. 2007, 13, RA119–RA131. [Google Scholar]
- Das, U.N. Gamma-linolenic acid, arachidonic acid, and eicosapentaenoic acid as potential anticancer drugs. Nutrition 1990, 6, 429–434. [Google Scholar] [PubMed]
- Andreoli Miyake, J.; Nascimento Gomes, R.; Colquhoun, A. Gamma-Linolenic acid alters migration, proliferation and apoptosis in human and rat glioblastoma cells. Prostaglandins Other Lipid Mediat. 2020, 150, 106452. [Google Scholar] [CrossRef]
- Benadiba, M.; Miyake, J.A.; Colquhoun, A. Gamma-linolenic acid alters Ku80, E2F1, and bax expression and induces micronucleus formation in C6 glioma cells in vitro. IUBMB Life 2009, 61, 244–251. [Google Scholar] [CrossRef]
- Miyake, J.A.; Benadiba, M.; Colquhoun, A. Gamma-linolenic acid inhibits both tumour cell cycle progression and angiogenesis in the orthotopic C6 glioma model through changes in VEGF, Flt1, ERK1/2, MMP2, cyclin D1, pRb, p53 and p27 protein expression. Lipids Health Dis. 2009, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Efraim, S. Interactions between macrophage cytokines and eicosanoids in expression of antitumour activity. Mediat. Inflamm. 1992, 1, 295–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, J.A.; Shacter, E. Regulation of Macrophage Cytokine Production by Prostaglandin E2. J. Biol. Chem. 1997, 272, 25693–25699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, A.; Biswas, A.; Srivastav, S.; Mukherjee, M.; Das, P.K.; Ukil, A. Prostaglandin E2 Negatively Regulates the Production of Inflammatory Cytokines/Chemokines and IL-17 in Visceral Leishmaniasis. J. Immunol. 2014, 193, 2330–2339. [Google Scholar] [CrossRef] [Green Version]
- Das, U.N. Essential Fatty Acids and Their Metabolites in the Pathobiology of Inflammation and Its Resolution. Biomolecules 2021, 11, 1873. [Google Scholar] [CrossRef]
- Das, U.N. Bioactive lipids in intervertebral disc degeneration and its therapeutic implications. Biosci. Rep. 2019, 39, BSR20192117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treffkorn, L.; Scheibe, R.; Maruyama, T.; Dieter, P. PGE2 exerts its effect on the LPS-induced release of TNF-alpha, ET-1, IL-1alpha, IL-6 and IL-10 via the EP2 and EP4 receptor in rat liver macrophages. Prostaglandins Other Lipid Mediat. 2004, 74, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Sravan Kumar, G.; Das, U.N. Effect of prostaglandins and their precursors on the proliferation of human lymphocytes and their secretion of tumor necrosis factor and various interleukins. Prostaglandins Leukot. Essen. Fat. Acids 1994, 50, 331–334. [Google Scholar] [CrossRef]
- Strong, V.E.; Mackrell, P.J.; Concannon, E.M.; Naama, H.A.; Schaefer, P.A.; Shaftan, G.W.; Stapleton, P.P.; Daly, J.M. Blocking prostaglandin E2 after trauma attenuates pro-inflammatory cytokines and improves survival. Shock 2000, 14, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Knöferl, M.W.; Diodato, M.D.; Schwacha, M.G.; Cioffi, W.G.; Bland, K.I.; Chaudry, I.H. Cyclooxygenase-2-mediated regulation of Kupffer cell interleukin-6 production following trauma-hemorrhage and subsequent sepsis. Shock 2001, 16, 479–483. [Google Scholar] [CrossRef]
- Anderson, G.D.; Hauser, S.D.; McGarity, K.L.; Bremer, M.E.; Isakson, P.C.; Gregory, S.A. Selective inhibition of cyclooxygenase (COX)-2 reverses inflammation and expression of COX-2 and interleukin 6 in rat adjuvant arthritis. J. Clin. Investig. 1996, 97, 2672–2679. [Google Scholar] [CrossRef] [Green Version]
- Dakin, S.G.; Colas, R.A.; Newton, J.; Gwilym, S.; Jones, N.; Reid, H.A.B.; Wood, S.; Appleton, L.; Wheway, K.; Watkins, B.; et al. 15-Epi-LXA4 and MaR1 counter inflammation in stromal cells from patients with Achilles tendinopathy and rupture. FASEB J. 2019, 33, 8043–8054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, J.; Dichter, E.; Lacorte, G.; Kerner, D.; Spur, B.; Rodriguez, A.; Yin, K. Lipoxin a4 increases survival by decreasing systemic inflammation and bacterial load in sepsis. Shock 2011, 36, 410–416. [Google Scholar] [CrossRef]
- Martinez, R.M.; Fattori, V.; Saito, P.; Pinto, I.C.; Rodrigues, C.C.A.; Melo, C.P.B.; Bussmann, A.J.C.; Staurengo-Ferrari, L.; Bezerra, J.R.; Vignoli, J.A.; et al. The Lipoxin Receptor/FPR2 Agonist BML-111 Protects Mouse Skin Against Ultraviolet B Radiation. Molecules. 2020, 25, 2953. [Google Scholar] [CrossRef]
- Shi, Y.; Pan, H.; Zhang, H.Z.; Zhao, X.Y.; Jin, J.; Wang, H.Y. Lipoxin A4 mitigates experimental autoimmune myocarditis by regulating inflammatory response, NF-κB and PI3K/Akt signaling pathway in mice. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 1850–1859. [Google Scholar]
- Kurtoğlu, E.L.; Kayhan, B.; Gül, M.; Kayhan, B.; Akdoğan Kayhan, M.; Karaca, Z.M.; Yeşilada, E.; Yılmaz, S. A bioactive product lipoxin A4 attenuates liver fibrosis in an experimental model by regulating immune response and modulating the expression of regeneration genes. Turk. J. Gastroenterol. 2019, 30, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Li, Z.; Jiang, S.; Tong, X.; Zou, X.; Wang, W.; Zhang, Z.; Wu, L.; Tian, D. Lipoxin A4 exerts protective effects against experimental acute liver failure by inhibiting the NF-κB pathway. Int. J. Mol. Med. 2016, 37, 773–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortier, S.M.; Penke, L.R.; Peters-Golden, M. Illuminating the lung regenerative potential of prostanoids. Sci. Adv. 2022, 8, eabp8322. [Google Scholar] [CrossRef]
- Tateishi, N.; Kakutani, S.; Kawashima, H.; Shibata, H.; Morita, I. Dietary supplementation of arachidonic acid increases arachidonic acid and lipoxin A₄ contents in colon but does not affect severity or prostaglandin E₂ content in murine colitis model. Lipids Health Dis. 2014, 13, 30. [Google Scholar] [CrossRef] [Green Version]
- Tateishi, N.; Kaneda, Y.; Kakutani, S.; Kawashima, H.; Shibata, H.; Morita, I. Dietary supplementation with arachidonic acid increases arachidonic acid content in paw, but does not affect arthritis severity or prostaglandin E2 content in rat adjuvant-induced arthritis model. Lipids Health Dis. 2015, 14, 3. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.S.; Xie, L.W.; Cai, S.; Xu, J.Y.; Zhou, H.; Tang, L.F.; Yang, C.; Fang, S.; Li, M.; Tian, Y. Dysbiosis of Gut Microbiota Is Associated With the Progression of Radiation-Induced Intestinal Injury and Is Alleviated by Oral Compound Probiotics in Mouse Model. Front. Cell Infect Microbiol. 2021, 11, 717636. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, Y.; Lu, P.; Wang, X.; Li, W.; Dong, H.; Fan, S.; Li, D. Gut metabolite Urolithin A mitigates ionizing radiation-induced intestinal damage. J. Cell Mol. Med. 2021, 25, 10306–10312. [Google Scholar] [CrossRef] [PubMed]
- Sittipo, P.; Pham, H.Q.; Park, C.E.; Kang, G.U.; Zhi, Y.; Ji, H.J.; Jang, A.; Seo, H.S.; Shin, J.H.; Lee, Y.K. Irradiation-Induced Intestinal Damage Is Recovered by the Indigenous Gut Bacteria Lactobacillus acidophilus. Front. Cell Infect. Microbiol. 2020, 10, 415. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Chou, W.C.; Lai, Y.; Liang, K.; Tam, J.W.; Brickey, W.J.; Chen, L.; Montgomery, N.D.; Li, X.; Bohannon, L.M.; et al. Multi-omics analyses of radiation survivors identify radioprotective microbes and metabolites. Science 2020, 370, eaay9097. [Google Scholar] [CrossRef]
- Ma, X.; Xu, T.; Qian, M.; Zhang, Y.; Yang, Z.; Han, X. Faecal microbiota transplantation alleviates early-life antibiotic-induced gut microbiota dysbiosis and mucosa injuries in a neonatal piglet model. Microbiol. Res. 2021, 255, 126942. [Google Scholar] [CrossRef]
- Nagatake, T.; Kishino, S.; Urano, E.; Murakami, H.; Kitamura, N.; Konishi, K.; Ohno, H.; Tiwari, P.; Morimoto, S.; Node, E.; et al. Intestinal microbe-dependent ω3 lipid metabolite αKetoA prevents inflammatory diseases in mice and cynomolgus macaques. Mucosal. Immunol. 2022, 15, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.N.; Zhu, J.; Pan, W.S.; Shen, S.R.; Shan, W.G.; Das, U.N. Effects of fish oil with a high content of n-3 polyunsaturated fatty acids on mouse gut microbiota. Arch. Med. Res. 2014, 45, 195–202. [Google Scholar] [CrossRef]
- Kindt, A.; Liebisch, G.; Clavel, T.; Haller, D.; Hörmannsperger, G.; Yoon, H.; Kolmeder, D.; Sigruener, A.; Krautbauer, S.; Seeliger, C.; et al. The gut microbiota promotes hepatic fatty acid desaturation and elongation in mice. Nat. Commun. 2018, 9, 3760. [Google Scholar] [CrossRef] [PubMed] [Green Version]






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′-3′) | Annealing Temp (°C) | Product Size (bp) |
|---|---|---|---|---|
| p53 | F | GCTGGTTCATCACTCCTCCC | 58.4 | 216 |
| R | GCTTCCCCATTTCACTCTGG | |||
| Bcl-2 | F | CTCGTCGCTACCGTCGTGACTTCG | 66 | 242 |
| R | CAGATGCCGGTTCAGGTACTCAGTC | |||
| Bax | F | CGGCGAATTGGAGATGAACTG | 58 | 161 |
| R | GCAAAGTAGAAGAGGGCAACC | |||
| AIM-2 | F | ACAAAGTGCGAGGAAGGAGA | 53.7 | 125 |
| R | TTTGGCTTTGCAGCCTTAAT | |||
| Caspase-1 | F | GAGAAGAGAGTGCTGAATCAG | 55.5 | 400 |
| R | CAAGACGTGTACGAGTGGTTG | |||
| Caspase-3 | F | TGTCATCTCGCTCTGGTACG | 55.5 | 392 |
| R | AGCCTCCACCGGTATCTTCT | |||
| TNF-α | F | CTGAACTTCGGGGTGATCGG | 58.4 | 122 |
| R | GGCTTGTCACTCGAATTTTGAGA | |||
| TGF-β | F | GGATACCAACTATTGCTTCAGCTCC | 65 | 156 |
| R | AGGCTCCAAATATAGGGGCAGGGTC | |||
| NF-κB | F | CAGCACTGATGGCATGGGGGACACTGACA | 68 | 588 |
| R | CCCAATGCATAGCCATTACACGTTT TTCACCTTAAATCTGCTT | |||
| IκB | F | CTTGGTGACTTTGGGTGCTGAT | 59 | 101 |
| R | GCGAAACCAGGTCAGGATTC | |||
| iNOS | F | CCCTTCCGAAGTTTCTGGCAGCAGC | 60 | 499 |
| R | GGCTGTCAGAGAGCCT CGTGGCTTTGG | |||
| COX-1 | F | AGGAGATGGCTGCTGAGTTGG | 57.5 | 601 |
| R | AATCTGACTTTCTGAGTTGCC | |||
| COX-2 | F | ACACACTCTATCACTGGCACC | 57.5 | 274 |
| R | TTCAGGGAGAAGCGTTTGC | |||
| D-6-dS | F | TGCCTGGGTCATCCTCTCGTA | 58 | 58 |
| R | GGCTGTGACGAGGGTAGGAA | |||
| D-5-dS | F | TGTGTGGGTGACACAGATGA | 53.5 | 115 |
| R | GTTGAAGGCTGATTGGTGAA | |||
| 5-LOX | F | ATGTTGGCATCTAGGTGCAGTGTG | 62 | 114 |
| R | ATCATGGCTTCCTTCACTGGCTTC | |||
| GAPDH | F | AACTTTGGCATTGTGGAAGG | 54 | 223 |
| R | ACACATTGGGGGTAGGAACA |
| Time | Group | LPO/NO Ratio | SOD (Units/mg ptn) | Catalase (µM H2O2/min/g ptn) | GST (µM/min/g ptn) | GPX (µM/min/g ptn) |
|---|---|---|---|---|---|---|
| 24 h | Control | 0.042 ± 0.004 | 198.68 ± 26.62 | 12,189.58 ± 1374.44 | 12.12 ± 2.96 | 93,868.97 ± 11,205.50 |
| GLA 250 ng | 0.05 ± 0.006 | 65.83 ± 9.10 a | 14,755.81 ± 1295.06 | 33.18 ± 0.69 a | 56,963.98 ± 2681.21 | |
| IR 2 Gy | 0.058 ± 0.006 a | 22.38 ± 2.04 a,b | 2318.51 ± 252.59 a,b | 2.45 ± 0.13 b | 11,870.93 ±477.72 a,b | |
| GLA + IR | 0.043 ± 0.004 c | 44.71 ± 5.95 a,c | 9107.89 ± 1237.38 b,c | 3.83 ± 0.57 b | 30,631.42 ± 1366.69 a,b,c | |
| 48 h | Control | 0.056 ± 0.007 | 96.63.84 ± 15.95 | 2013.84 ± 3079.58 | 17.07 ± 1.14 | 94,664.32 ± 11,809.67 |
| GLA 250 ng | 0.079 ± 0.006 | 38.24 ± 10.19 | 20,689.78 ± 1258.46 | 30.45 ± 2.26 a | 52,692.46 ± 1713.91 | |
| IR 2 Gy | 0.222 ± 0.013 a,b | 6.9 ± 1.6 a,b | 859.86 ± 61.84 a,b | 3.34 ± 0.34 a,b | 10,702.34 ± 391.29 b,c | |
| GLA + IR | 0.085 ± 0.012c | 28.54 ± 2.20 a,c | 6322.51 ± 1398.04 a,b,c | 13.57 ± 1.13 b,c | 28,497.30 ± 1218.81 a,b,c |
| Group | Nitric Oxide (µM) | Lipid Peroxide (µM) | NO/LPO Ratio | SOD (Units/g Protein) | Catalase (µM H2O2/min/g Protein) | GST (µM/min/g Protein) | GPX (µM/min/g Protein) | |
|---|---|---|---|---|---|---|---|---|
| Day 1 | Control | 1.92 ± 0.07 | 1.26 ± 0.06 | 1.52 ± 0.02 | 5.44 ± 0.81 | 853.02 ± 61.10 | 10.87 ± 0.50 | 29.19 ± 0.59 |
| GLA 100 µg | 2.56 ± 0.39 | 1.05 ± 0.10 | 2.72 ± 0.63 | 7.51 ± 1.35 | 804.58 ± 119.91 | 12.37 ± 1.48 | 43.63 ± 4.31 a | |
| IR 7.5 Gy | 2.70 ± 0.46 | 1.38 ± 0.29 | 2.25 ± 0.34 | 10.41 ± 2.17 | 944.28 ± 98.88 | 10.43 ± 0.84 | 32.87 ± 4.03 | |
| GLA + IR | 3.41 ± 0.43 a | 1.44 ± 0.4 | 3.33 ± 0.94 a | 13.11 ± 1.55 a | 2052.89 ± 544.06 a,b,c | 7.97 ± 1.24 b | 44.65 ± 5.19 a | |
| Day 3 | Control | 3.77 ± 0.61 | 1.43 ± 0.18 | 2.96 ± 0.57 | 6.60 ± 1.30 | 1613.74 ± 512.70 | 4.36 ± 0.31 | 38.48 ± 4.97 |
| GLA 100 µg | 2.47 ± 0.20 | 1.07 ± 0.08 | 2.41 ± 0.31 | 7.93 ± 1.53 | 1981.82 ± 417.53 | 6.97 ± 0.30 a | 59.04 ± 4.38 a | |
| IR 7.5 Gy | 3.31 ± 0.63 | 0.88 ± 0.22 | 5.43 ± 1.72 | 4.23 ± 1.27 | 1319.49 ± 78.28 | 4.08 ± 0.16 b | 47.81 ± 1.69 b | |
| GLA + IR | 2.98 ± 0.65 | 0.66 ± 0.14 b | 5.95 ± 1.86 | 6.64 ± 1.54 | 2242.91 ± 284.87 c | 6.79 ± 0.91 a,c | 53.48 ± 6.43 | |
| Day 7 | Control | 4.36 ± 0.39 | 1.13 ± 0.07 | 3.94 ± 0.44 | 55.91 ± 0.84 | 869.36 ± 53.96 | 0.49 ± 0.01 | 33.52 ± 0.78 |
| GLA 100 µg | 5.55 ± 0.06 a | 1.27 ± 0.09 | 4.52 ± 0.45 | 52.09 ± 0.08 a | 1171.99 ± 118.60 a | 0.39 ± 0.02 a | 28.78 ± 0.58 a | |
| IR 7.5 Gy | 4.20 ± 0.23 b | 1.04 ± 0.10 | 4.19 ± 0.38 | 35.45 ± 8.70a | 422.73 ± 124.27 a,b | 0.70 ± 0.13 b | 38.80 ± 5.59 | |
| GLA + IR | 5.79 ± 0.15 a,c | 0.17 ± 0.07 a,b,c | 77.53 ± 21.07 a,b,c | 64.71 ± 2.16 a,b,c | 1096.76 ± 259.34 a,c | 1.44 ± 0.31 a,b,c | 42.80 ± 2.72 a,b | |
| Day 14 | Control | 2.25 ± 0.34 | 0.89 ± 0.01 | 2.53 ± 0.40 | 19.23 ± 3.54 | 454.17 ± 82.91 | 1.18 ± 0.20 | 9.46 ± 0.24 |
| GLA 100 µg | 7.23 ± 0.09 a | 1.98 ± 0.45 a | 4.90 ± 1.08 | 16.37 ± 0.25 | 739.23 ± 62.85 a | 0.99 ± 0.04 | 8.69 ± 0.93 | |
| IR 7.5 Gy | 7.29 ± 0.18 a | 0.29 ± 0.12 a,b | 235.95 ± 99.33 a,b | 8.03 ± 3.07 a,b | 469.72 ± 31.52 b | 1.43 ± 0.15 b | 12.10 ± 0.56 a,b | |
| GLA + IR | 2.32 ± 0.26 b,c | 0.67 ± 0.26 b | 16.48 ± 8.86 | 20.63 ± 0.53 b,c | 316.00 ± 76.15 b,c | 1.26 ± 0.24 | 9.50 ± 0.25 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rengachar, P.; Bhatt, A.N.; Polavarapu, S.; Veeramani, S.; Krishnan, A.; Sadananda, M.; Das, U.N. Gamma-Linolenic Acid (GLA) Protects against Ionizing Radiation-Induced Damage: An In Vitro and In Vivo Study. Biomolecules 2022, 12, 797. https://doi.org/10.3390/biom12060797
Rengachar P, Bhatt AN, Polavarapu S, Veeramani S, Krishnan A, Sadananda M, Das UN. Gamma-Linolenic Acid (GLA) Protects against Ionizing Radiation-Induced Damage: An In Vitro and In Vivo Study. Biomolecules. 2022; 12(6):797. https://doi.org/10.3390/biom12060797
Chicago/Turabian StyleRengachar, Poorani, Anant Narayan Bhatt, Sailaja Polavarapu, Senthil Veeramani, Anand Krishnan, Monika Sadananda, and Undurti N. Das. 2022. "Gamma-Linolenic Acid (GLA) Protects against Ionizing Radiation-Induced Damage: An In Vitro and In Vivo Study" Biomolecules 12, no. 6: 797. https://doi.org/10.3390/biom12060797