
Utility of the Specialized Pro-Resolving Mediators as Diagnostic and Prognostic Biomarkers in Disease


Abstract
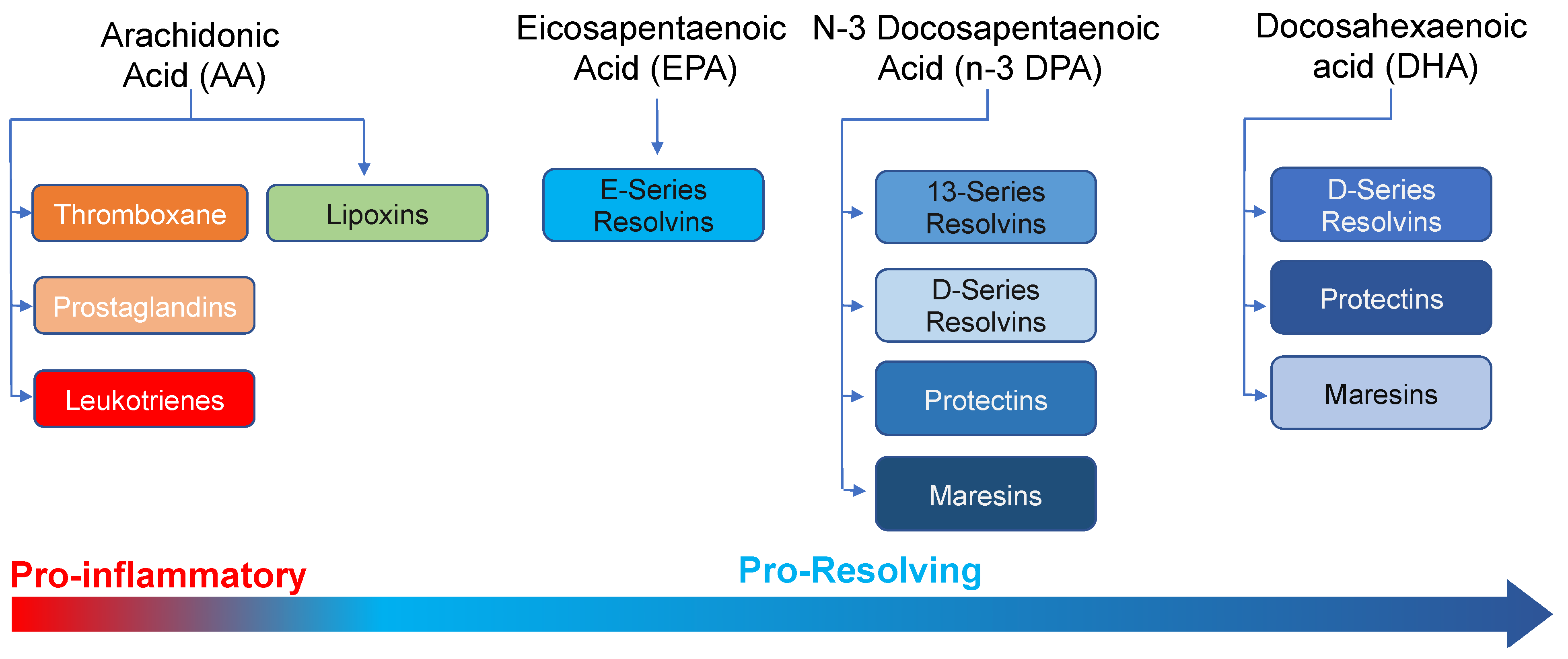
:1. Introduction
2. Additional Quantitation Methodologies
3. Sample Preparation and Quantitation Methodologies
4. Diagnostic Utility of SPM in Infections
5. Pro-Resolving Mediators as Biomarkers in Chronic Inflammatory Conditions
5.1. Periodontal Disease
5.2. Rheumatoid Arthritis
5.3. Vascular Disease
5.4. Allergic Inflammation
5.5. COVID-19
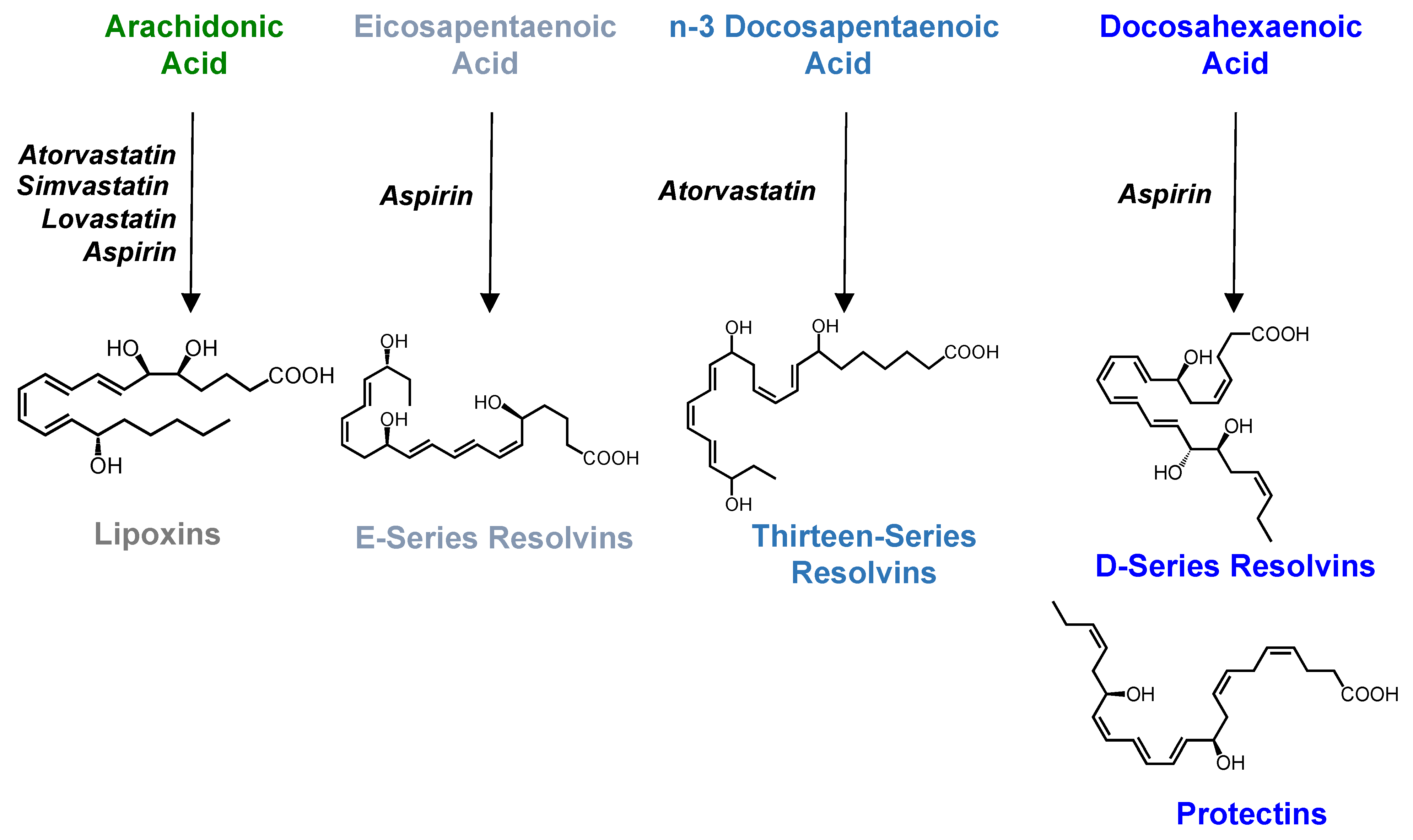
6. SPM as Biomarkers for Determining Therapeutic Efficacy of Anti-Inflammatory Drugs
7. Potential for the Use of SPM as Biomarkers for Determining the Utility of Omega-3 Supplements in Regulating Inflammation
8. Utility of Machine Learning in Biomarker Identification
9. Current Limitations and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collins, F.S.; Varmus, H. A new initiative on precision medicine. N. Engl. J. Med. 2015, 372, 793–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jameson, J.L.; Longo, D.L. Precision medicine—Personalized, problematic, and promising. N. Engl. J. Med. 2015, 372, 2229–2234. [Google Scholar] [CrossRef] [Green Version]
- Aletaha, D.; Smolen, J.S. Diagnosis and Management of Rheumatoid Arthritis: A Review. JAMA 2018, 320, 1360–1372. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.; Ahmed, M.; Conway, R.; Carey, J.J. Risk of Infection with Methotrexate Therapy in Inflammatory Diseases: A Systematic Review and Meta-Analysis. J. Clin. Med. 2018, 8, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, B.D.; Clish, C.B.; Schmidt, B.; Gronert, K.; Serhan, C.N. Lipid mediator class switching during acute inflammation: Signals in resolution. Nat. Immunol. 2001, 2, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Clish, C.B.; Brannon, J.; Colgan, S.P.; Chiang, N.; Gronert, K. Novel functional sets of lipid-derived mediators with antiinflammatory actions generated from omega-3 fatty acids via cyclooxygenase 2-nonsteroidal antiinflammatory drugs and transcellular processing. J. Exp. Med. 2000, 192, 1197–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serhan, C.N.; Hong, S.; Gronert, K.; Colgan, S.P.; Devchand, P.R.; Mirick, G.; Moussignac, R.L. Resolvins: A family of bioactive products of omega-3 fatty acid transformation circuits initiated by aspirin treatment that counter proinflammation signals. J. Exp. Med. 2002, 196, 1025–1037. [Google Scholar] [CrossRef] [Green Version]
- Chiang, N.; Serhan, C.N. Specialized pro-resolving mediator network: An update on production and actions. Essays Biochem. 2020, 64, 443–462. [Google Scholar] [CrossRef]
- Serhan, C.N.; Dalli, J.; Colas, R.A.; Winkler, J.W.; Chiang, N. Protectins and maresins: New pro-resolving families of mediators in acute inflammation and resolution bioactive metabolome. Biochim. Biophys. Acta 2015, 1851, 397–413. [Google Scholar] [CrossRef] [Green Version]
- Dalli, J.; Chiang, N.; Serhan, C.N. Identification of 14-series sulfido-conjugated mediators that promote resolution of infection and organ protection. Proc. Natl. Acad. Sci. USA 2014, 111, E4753–E4761. [Google Scholar] [CrossRef] [Green Version]
- Dalli, J.; Chiang, N.; Serhan, C.N. Elucidation of novel 13-series resolvins that increase with atorvastatin and clear infections. Nat. Med. 2015, 21, 1071–1075. [Google Scholar] [CrossRef] [PubMed]
- Dalli, J.; Colas, R.A.; Serhan, C.N. Novel n-3 immunoresolvents: Structures and actions. Sci. Rep. 2013, 3, 1940. [Google Scholar] [CrossRef] [PubMed]
- Dalli, J.; Ramon, S.; Norris, P.C.; Colas, R.A.; Serhan, C.N. Novel proresolving and tissue-regenerative resolvin and protectin sulfido-conjugated pathways. FASEB J. 2015, 29, 2120–2136. [Google Scholar] [CrossRef] [Green Version]
- Freedman, C.; Tran, A.; Tourdot, B.E.; Kalyanaraman, C.; Perry, S.; Holinstat, M.; Jacobson, M.P.; Holman, T.R. Biosynthesis of the Maresin Intermediate, 13S,14S-Epoxy-DHA, by Human 15-Lipoxygenase and 12-Lipoxygenase and Its Regulation through Negative Allosteric Modulators. Biochemistry 2020, 59, 1832–1844. [Google Scholar] [CrossRef]
- Perry, S.C.; Kalyanaraman, C.; Tourdot, B.E.; Conrad, W.S.; Akinkugbe, O.; Freedman, J.C.; Holinstat, M.; Jacobson, M.P.; Holman, T.R. 15-Lipoxygenase-1 biosynthesis of 7S,14S-diHDHA implicates 15-lipoxygenase-2 in biosynthesis of resolvin D5. J. Lipid Res. 2020, 61, 1087–1103. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.C.; Kalyanaraman, C.; Yamaguchi, A.; Holinstat, M.; Jacobson, M.P.; Holman, T.R. In Vitro Biosynthetic Pathway Investigations of Neuroprotectin D1 (NPD1) and Protectin DX (PDX) by Human 12-Lipoxygenase, 15-Lipoxygenase-1, and 15-Lipoxygenase-2. Biochemistry 2021, 60, 1741–1754. [Google Scholar] [CrossRef]
- Gerlach, B.D.; Marinello, M.; Heinz, J.; Rymut, N.; Sansbury, B.E.; Riley, C.O.; Sadhu, S.; Hosseini, Z.; Kojima, Y.; Tang, D.D.; et al. Resolvin D1 promotes the targeting and clearance of necroptotic cells. Cell Death Differ. 2020, 27, 525–539. [Google Scholar] [CrossRef]
- Rymut, N.; Heinz, J.; Sadhu, S.; Hosseini, Z.; Riley, C.O.; Marinello, M.; Maloney, J.; MacNamara, K.C.; Spite, M.; Fredman, G. Resolvin D1 promotes efferocytosis in aging by limiting senescent cell-induced MerTK cleavage. FASEB J. 2020, 34, 597–609. [Google Scholar] [CrossRef] [Green Version]
- Sansbury, B.E.; Li, X.; Wong, B.; Patsalos, A.; Giannakis, N.; Zhang, M.J.; Nagy, L.; Spite, M. Myeloid ALX/FPR2 regulates vascularization following tissue injury. Proc. Natl. Acad. Sci. USA 2020, 117, 14354–14364. [Google Scholar] [CrossRef]
- Giannakis, N.; Sansbury, B.E.; Patsalos, A.; Hays, T.T.; Riley, C.O.; Han, X.; Spite, M.; Nagy, L. Dynamic changes to lipid mediators support transitions among macrophage subtypes during muscle regeneration. Nat. Immunol. 2019, 20, 626–636. [Google Scholar] [CrossRef]
- De Gaetano, M.; McEvoy, C.; Andrews, D.; Cacace, A.; Hunter, J.; Brennan, E.; Godson, C. Specialized Pro-resolving Lipid Mediators: Modulation of Diabetes-Associated Cardio-, Reno-, and Retino-Vascular Complications. Front. Pharmacol. 2018, 9, 1488. [Google Scholar] [CrossRef] [PubMed]
- Mattoscio, D.; Isopi, E.; Lamolinara, A.; Patruno, S.; Medda, A.; De Cecco, F.; Chiocca, S.; Iezzi, M.; Romano, M.; Recchiuti, A. Resolvin D1 reduces cancer growth stimulating a protective neutrophil-dependent recruitment of anti-tumor monocytes. J. Exp. Clin. Cancer Res. 2021, 40, 129. [Google Scholar] [CrossRef] [PubMed]
- Gobbetti, T.; Dalli, J.; Colas, R.A.; Federici Canova, D.; Aursnes, M.; Bonnet, D.; Alric, L.; Vergnolle, N.; Deraison, C.; Hansen, T.V.; et al. Protectin D1n-3 DPA and resolvin D5n-3 DPA are effectors of intestinal protection. Proc. Natl. Acad. Sci. USA 2017, 114, 3963–3968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, R.M.; Gonzalez-Nunez, M.; Walker, M.E.; Gomez, E.A.; Colas, R.A.; Montero-Melendez, T.; Perretti, M.; Dalli, J. Loss of 15-lipoxygenase disrupts Treg differentiation altering their pro-resolving functions. Cell Death Differ. 2021, 28, 3140–3160. [Google Scholar] [CrossRef]
- Libreros, S.; Shay, A.E.; Nshimiyimana, R.; Fichtner, D.; Martin, M.J.; Wourms, N.; Serhan, C.N. A New E-Series Resolvin: RvE4 Stereochemistry and Function in Efferocytosis of Inflammation-Resolution. Front. Immunol. 2020, 11, 631319. [Google Scholar] [CrossRef]
- Wei, J.; Mattapallil, M.J.; Horai, R.; Jittayasothorn, Y.; Modi, A.P.; Sen, H.N.; Gronert, K.; Caspi, R.R. A novel role for lipoxin A4 in driving a lymph node-eye axis that controls autoimmunity to the neuroretina. Elife 2020, 9, e51102. [Google Scholar] [CrossRef]
- Alvarez, C.; Abdalla, H.; Sulliman, S.; Rojas, P.; Wu, Y.C.; Almarhoumi, R.; Huang, R.Y.; Galindo, M.; Vernal, R.; Kantarci, A. RvE1 Impacts the Gingival Inflammatory Infiltrate by Inhibiting the T Cell Response in Experimental Periodontitis. Front. Immunol. 2021, 12, 664756. [Google Scholar] [CrossRef]
- Chiurchiu, V.; Leuti, A.; Dalli, J.; Jacobsson, A.; Battistini, L.; Maccarrone, M.; Serhan, C.N. Proresolving lipid mediators resolvin D1, resolvin D2, and maresin 1 are critical in modulating T cell responses. Sci. Transl. Med. 2016, 8, 353ra111. [Google Scholar] [CrossRef] [Green Version]
- Bhat, T.A.; Kalathil, S.G.; Miller, A.; Thatcher, T.H.; Sime, P.J.; Thanavala, Y. Specialized Proresolving Mediators Overcome Immune Suppression Induced by Exposure to Secondhand Smoke. J. Immunol. 2020, 205, 3205–3217. [Google Scholar] [CrossRef]
- Ramon, S.; Gao, F.; Serhan, C.N.; Phipps, R.P. Specialized proresolving mediators enhance human B cell differentiation to antibody-secreting cells. J. Immunol. 2012, 189, 1036–1042. [Google Scholar] [CrossRef] [Green Version]
- Quiros, M.; Feier, D.; Birkl, D.; Agarwal, R.; Zhou, D.W.; Garcia, A.J.; Parkos, C.A.; Nusrat, A. Resolvin E1 is a pro-repair molecule that promotes intestinal epithelial wound healing. Proc. Natl. Acad. Sci. USA 2020, 117, 9477–9482. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Yang, H.; Pan, Z.; Wang, Z.; Wolosin, J.M.; Gjorstrup, P.; Reinach, P.S. Dependence of resolvin-induced increases in corneal epithelial cell migration on EGF receptor transactivation. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5601–5609. [Google Scholar] [CrossRef] [PubMed]
- Levy, O.; Canny, G.; Serhan, C.N.; Colgan, S.P. Expression of BPI (bactericidal/permeability-increasing protein) in human mucosal epithelia. Biochem. Soc. Trans. 2003, 31, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Canny, G.; Levy, O.; Furuta, G.T.; Narravula-Alipati, S.; Sisson, R.B.; Serhan, C.N.; Colgan, S.P. Lipid mediator-induced expression of bactericidal/ permeability-increasing protein (BPI) in human mucosal epithelia. Proc. Natl. Acad. Sci. USA 2002, 99, 3902–3907. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, R.; Raghavan, S.; Rao, G.N. Resolvin D1 via prevention of ROS-mediated SHP2 inactivation protects endothelial adherens junction integrity and barrier function. Redox Biol. 2017, 12, 438–455. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, T.; Hosur, K.; Abe, T.; Kantarci, A.; Ziogas, A.; Wang, B.; Van Dyke, T.E.; Chavakis, T.; Hajishengallis, G. Antagonistic effects of IL-17 and D-resolvins on endothelial Del-1 expression through a GSK-3beta-C/EBPbeta pathway. Nat. Commun. 2015, 6, 8272. [Google Scholar] [CrossRef] [Green Version]
- Chiang, N.; Serhan, C.N. Structural elucidation and physiologic functions of specialized pro-resolving mediators and their receptors. Mol. Aspects Med. 2017, 58, 114–129. [Google Scholar] [CrossRef]
- Chiang, N.; Dalli, J.; Colas, R.A.; Serhan, C.N. Identification of resolvin D2 receptor mediating resolution of infections and organ protection. J. Exp. Med. 2015, 212, 1203–1217. [Google Scholar] [CrossRef] [Green Version]
- Tourki, B.; Kain, V.; Pullen, A.B.; Norris, P.C.; Patel, N.; Arora, P.; Leroy, X.; Serhan, C.N.; Halade, G.V. Lack of resolution sensor drives age-related cardiometabolic and cardiorenal defects and impedes inflammation-resolution in heart failure. Mol. Metab. 2020, 31, 138–149. [Google Scholar] [CrossRef]
- Gomez, E.A.; Colas, R.A.; Souza, P.R.; Hands, R.; Lewis, M.J.; Bessant, C.; Pitzalis, C.; Dalli, J. Blood pro-resolving mediators are linked with synovial pathology and are predictive of DMARD responsiveness in rheumatoid arthritis. Nat. Commun. 2020, 11, 5420. [Google Scholar] [CrossRef]
- Barden, A.E.; Moghaddami, M.; Mas, E.; Phillips, M.; Cleland, L.G.; Mori, T.A. Specialised pro-resolving mediators of inflammation in inflammatory arthritis. Prostaglandins Leukot. Essent. Fatty Acids 2016, 107, 24–29. [Google Scholar] [CrossRef]
- Freire, M.O.; Dalli, J.; Serhan, C.N.; Van Dyke, T.E. Neutrophil Resolvin E1 Receptor Expression and Function in Type 2 Diabetes. J. Immunol. 2017, 198, 718–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elabdeen, H.R.; Mustafa, M.; Szklenar, M.; Ruhl, R.; Ali, R.; Bolstad, A.I. Ratio of pro-resolving and pro-inflammatory lipid mediator precursors as potential markers for aggressive periodontitis. PLoS ONE 2013, 8, e70838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.W.; Colas, R.A.; Dalli, J.; Arnardottir, H.H.; Nguyen, D.; Hasturk, H.; Chiang, N.; Van Dyke, T.E.; Serhan, C.N. Maresin 1 Biosynthesis and Proresolving Anti-infective Functions with Human-Localized Aggressive Periodontitis Leukocytes. Infect. Immun. 2015, 84, 658–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claria, J.; Serhan, C.N. Aspirin triggers previously undescribed bioactive eicosanoids by human endothelial cell-leukocyte interactions. Proc. Natl. Acad. Sci. USA 1995, 92, 9475–9479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilligan, M.M.; Gartung, A.; Sulciner, M.L.; Norris, P.C.; Sukhatme, V.P.; Bielenberg, D.R.; Huang, S.; Kieran, M.W.; Serhan, C.N.; Panigrahy, D. Aspirin-triggered proresolving mediators stimulate resolution in cancer. Proc. Natl. Acad. Sci. USA 2019, 116, 6292–6297. [Google Scholar] [CrossRef] [Green Version]
- Planaguma, A.; Pfeffer, M.A.; Rubin, G.; Croze, R.; Uddin, M.; Serhan, C.N.; Levy, B.D. Lovastatin decreases acute mucosal inflammation via 15-epi-lipoxin A4. Mucosal. Immunol. 2010, 3, 270–279. [Google Scholar] [CrossRef] [Green Version]
- Birnbaum, Y.; Ye, Y.; Lin, Y.; Freeberg, S.Y.; Nishi, S.P.; Martinez, J.D.; Huang, M.H.; Uretsky, B.F.; Perez-Polo, J.R. Augmentation of myocardial production of 15-epi-lipoxin-a4 by pioglitazone and atorvastatin in the rat. Circulation 2006, 114, 929–935. [Google Scholar] [CrossRef] [Green Version]
- Radmark, O.; Malmsten, C.; Samuelsson, B.; Clark, D.A.; Goto, G.; Marfat, A.; Corey, E.J. Leukotriene A: Stereochemistry and enzymatic conversion to leukotriene B. Biochem. Biophys. Res. Commun. 1980, 92, 954–961. [Google Scholar] [CrossRef]
- Pfeiffer, F.; Mayer, G. Selection and Biosensor Application of Aptamers for Small Molecules. Front. Chem. 2016, 4, 25. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Cohen, L.; Wang, J.; Walt, D.R. Competitive Immunoassays for the Detection of Small Molecules Using Single Molecule Arrays. J. Am. Chem. Soc. 2018, 140, 18132–18139. [Google Scholar] [CrossRef] [PubMed]
- Liakh, I.; Pakiet, A.; Sledzinski, T.; Mika, A. Modern Methods of Sample Preparation for the Analysis of Oxylipins in Biological Samples. Molecules 2019, 24, 1639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koenis, D.S.; Beegun, I.; Jouvene, C.C.; Aguirre, G.A.; Souza, P.R.; Gonzalez-Nunez, M.; Ly, L.; Pistorius, K.; Kocher, H.M.; Ricketts, W.; et al. Disrupted Resolution Mechanisms Favor Altered Phagocyte Responses in COVID-19. Circ. Res. 2021, 129, e54–e71. [Google Scholar] [CrossRef] [PubMed]
- Diaz, L.A.; Altman, N.H.; Khan, W.N.; Serhan, C.N.; Adkins, B. Specialized Proresolving Mediators Rescue Infant Mice from Lethal Citrobacter rodentium Infection and Promote Immunity against Reinfection. Infect. Immun. 2017, 85, e00464-17. [Google Scholar] [CrossRef] [Green Version]
- Chiang, N.; Fredman, G.; Backhed, F.; Oh, S.F.; Vickery, T.; Schmidt, B.A.; Serhan, C.N. Infection regulates pro-resolving mediators that lower antibiotic requirements. Nature 2012, 484, 524–528. [Google Scholar] [CrossRef] [Green Version]
- Palmer, C.D.; Mancuso, C.J.; Weiss, J.P.; Serhan, C.N.; Guinan, E.C.; Levy, O. 17(R)-Resolvin D1 differentially regulates TLR4-mediated responses of primary human macrophages to purified LPS and live E. coli. J. Leukoc. Biol. 2011, 90, 459–470. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.; Luo, L.; Wang, Q.; Yan, S.; Lin, J.; Li, D.; Cao, B.; Mei, H.; Ying, B.; Bin, L.; et al. Maresin 1 attenuates mitochondrial dysfunction through the ALX/cAMP/ROS pathway in the cecal ligation and puncture mouse model and sepsis patients. Lab. Investig. 2018, 98, 715–733. [Google Scholar] [CrossRef]
- Motwani, M.P.; Colas, R.A.; George, M.J.; Flint, J.D.; Dalli, J.; Richard-Loendt, A.; De Maeyer, R.P.; Serhan, C.N.; Gilroy, D.W. Pro-resolving mediators promote resolution in a human skin model of UV-killed Escherichia coli-driven acute inflammation. JCI Insight 2018, 3, e94463. [Google Scholar] [CrossRef] [Green Version]
- Norris, P.C.; Skulas-Ray, A.C.; Riley, I.; Richter, C.K.; Kris-Etherton, P.M.; Jensen, G.L.; Serhan, C.N.; Maddipati, K.R. Identification of specialized pro-resolving mediator clusters from healthy adults after intravenous low-dose endotoxin and omega-3 supplementation: A methodological validation. Sci. Rep. 2018, 8, 18050. [Google Scholar] [CrossRef] [Green Version]
- Dalli, J.; Colas, R.A.; Quintana, C.; Barragan-Bradford, D.; Hurwitz, S.; Levy, B.D.; Choi, A.M.; Serhan, C.N.; Baron, R.M. Human Sepsis Eicosanoid and Proresolving Lipid Mediator Temporal Profiles: Correlations with Survival and Clinical Outcomes. Crit. Care Med. 2017, 45, 58–68. [Google Scholar] [CrossRef]
- Colas, R.A.; Nhat, L.T.H.; Thuong, N.T.T.; Gomez, E.A.; Ly, L.; Thanh, H.H.; Mai, N.T.H.; Phu, N.H.; Thwaites, G.E.; Dalli, J. Proresolving mediator profiles in cerebrospinal fluid are linked with disease severity and outcome in adults with tuberculous meningitis. FASEB J. 2019, 33, 13028–13039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramon, S.; Baker, S.F.; Sahler, J.M.; Kim, N.; Feldsott, E.A.; Serhan, C.N.; Martinez-Sobrido, L.; Topham, D.J.; Phipps, R.P. The specialized proresolving mediator 17-HDHA enhances the antibody-mediated immune response against influenza virus: A new class of adjuvant? J. Immunol. 2014, 193, 6031–6040. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Kuba, K.; Ichikawa, A.; Nakayama, M.; Katahira, J.; Iwamoto, R.; Watanebe, T.; Sakabe, S.; Daidoji, T.; Nakamura, S.; et al. The lipid mediator protectin D1 inhibits influenza virus replication and improves severe influenza. Cell 2013, 153, 112–125. [Google Scholar] [CrossRef] [Green Version]
- Rajasagi, N.K.; Reddy, P.B.; Suryawanshi, A.; Mulik, S.; Gjorstrup, P.; Rouse, B.T. Controlling herpes simplex virus-induced ocular inflammatory lesions with the lipid-derived mediator resolvin E1. J. Immunol. 2011, 186, 1735–1746. [Google Scholar] [CrossRef] [PubMed]
- Recchiuti, A.; Patruno, S.; Mattoscio, D.; Isopi, E.; Pomilio, A.; Lamolinara, A.; Iezzi, M.; Pecce, R.; Romano, M. Resolvin D1 and D2 reduce SARS-CoV-2-induced inflammatory responses in cystic fibrosis macrophages. FASEB J. 2021, 35, e21441. [Google Scholar] [CrossRef] [PubMed]
- Archambault, A.S.; Zaid, Y.; Rakotoarivelo, V.; Turcotte, C.; Dore, E.; Dubuc, I.; Martin, C.; Flamand, O.; Amar, Y.; Cheikh, A.; et al. High levels of eicosanoids and docosanoids in the lungs of intubated COVID-19 patients. FASEB J. 2021, 35, e21666. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, B.; Sharma, L.; Roberts, L.; Peng, X.; Bermejo, S.; Leighton, I.; Casanovas-Massana, A.; Minasyan, M.; Farhadian, S.; Ko, A.I.; et al. Cutting Edge: Severe SARS-CoV-2 Infection in Humans Is Defined by a Shift in the Serum Lipidome, Resulting in Dysregulation of Eicosanoid Immune Mediators. J. Immunol. 2021, 206, 329–334. [Google Scholar] [CrossRef]
- Serhan, C.N.; Levy, B.D. Resolvins in inflammation: Emergence of the pro-resolving superfamily of mediators. J. Clin. Investig. 2018, 128, 2657–2669. [Google Scholar] [CrossRef]
- Panigrahy, D.; Gilligan, M.M.; Serhan, C.N.; Kashfi, K. Resolution of inflammation: An organizing principle in biology and medicine. Pharmacol. Ther. 2021, 227, 107879. [Google Scholar] [CrossRef]
- Carracedo, M.; Artiach, G.; Arnardottir, H.; Back, M. The resolution of inflammation through omega-3 fatty acids in atherosclerosis, intimal hyperplasia, and vascular calcification. Semin. Immunopathol. 2019, 41, 757–766. [Google Scholar] [CrossRef] [Green Version]
- Halade, G.V.; Tourki, B. Specialized Pro-resolving Mediators Directs Cardiac Healing and Repair with Activation of Inflammation and Resolution Program in Heart Failure. Adv. Exp. Med. Biol. 2019, 1161, 45–64. [Google Scholar] [CrossRef] [PubMed]
- Fredman, G. Can Inflammation-Resolution Provide Clues to Treat Patients According to Their Plaque Phenotype? Front. Pharmacol. 2019, 10, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Zhang, Y.Q.; Qadri, Y.J.; Serhan, C.N.; Ji, R.R. Microglia in Pain: Detrimental and Protective Roles in Pathogenesis and Resolution of Pain. Neuron 2018, 100, 1292–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Miao, Z.; Xu, X.; Schultzberg, M.; Zhao, Y. Reduced Levels of Plasma Lipoxin A4 Are Associated with Post-Stroke Cognitive Impairment. J. Alzheimers Dis. 2021, 79, 607–613. [Google Scholar] [CrossRef]
- Wang, X.; Zhu, M.; Hjorth, E.; Cortes-Toro, V.; Eyjolfsdottir, H.; Graff, C.; Nennesmo, I.; Palmblad, J.; Eriksdotter, M.; Sambamurti, K.; et al. Resolution of inflammation is altered in Alzheimer’s disease. Alzheimers Dement. 2015, 11, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Wang, X.; Hjorth, E.; Colas, R.A.; Schroeder, L.; Granholm, A.C.; Serhan, C.N.; Schultzberg, M. Pro-Resolving Lipid Mediators Improve Neuronal Survival and Increase Abeta42 Phagocytosis. Mol. Neurobiol. 2016, 53, 2733–2749. [Google Scholar] [CrossRef]
- Fosshaug, L.E.; Colas, R.A.; Anstensrud, A.K.; Gregersen, I.; Nymo, S.; Sagen, E.L.; Michelsen, A.; Vinge, L.E.; Oie, E.; Gullestad, L.; et al. Early increase of specialized pro-resolving lipid mediators in patients with ST-elevation myocardial infarction. EBioMedicine 2019, 46, 264–273. [Google Scholar] [CrossRef]
- Halade, G.V.; Kain, V.; Dillion, C.; Beasley, M.; Dudenbostel, T.; Oparil, S.; Limdi, N.A. Race-based and sex-based differences in bioactive lipid mediators after myocar.rdial infarction. ESC Heart Fail. 2020, 7, 1700–1710. [Google Scholar] [CrossRef]
- Grenon, S.M.; Owens, C.D.; Nosova, E.V.; Hughes-Fulford, M.; Alley, H.F.; Chong, K.; Perez, S.; Yen, P.K.; Boscardin, J.; Hellmann, J.; et al. Short-Term, High-Dose Fish Oil Supplementation Increases the Production of Omega-3 Fatty Acid-Derived Mediators in Patients With Peripheral Artery Disease (the OMEGA-PAD I Trial). J. Am. Heart Assoc. 2015, 4, e002034. [Google Scholar] [CrossRef] [Green Version]
- Schaller, M.S.; Zahner, G.J.; Gasper, W.J.; Harris, W.S.; Conte, M.S.; Hills, N.K.; Grenon, S.M. Relationship between the omega-3 index and specialized pro-resolving lipid mediators in patients with peripheral arterial disease taking fish oil supplements. J. Clin. Lipidol. 2017, 11, 1289–1295. [Google Scholar] [CrossRef]
- Fredman, G.; Hellmann, J.; Proto, J.D.; Kuriakose, G.; Colas, R.A.; Dorweiler, B.; Connolly, E.S.; Solomon, R.; Jones, D.M.; Heyer, E.J.; et al. An imbalance between specialized pro-resolving lipid mediators and pro-inflammatory leukotrienes promotes instability of atherosclerotic plaques. Nat. Commun. 2016, 7, 12859. [Google Scholar] [CrossRef] [PubMed]
- Tobon-Arroyave, S.I.; Isaza-Guzman, D.M.; Gomez-Ortega, J.; Florez-Alzate, A.A. Salivary levels of specialized pro-resolving lipid mediators as indicators of periodontal health/disease status. J. Clin. Periodontol. 2019, 46, 978–990. [Google Scholar] [CrossRef]
- Vickery, T.W.; Armstrong, M.; Kofonow, J.M.; Robertson, C.E.; Kroehl, M.E.; Reisdorph, N.A.; Ramakrishnan, V.R.; Frank, D.N. Altered tissue specialized pro-resolving mediators in chronic rhinosinusitis. Prostaglandins Leukot. Essent. Fatty Acids 2021, 164, 102218. [Google Scholar] [CrossRef] [PubMed]
- Shivakoti, R.; Dalli, J.; Kadam, D.; Gaikwad, S.; Barthwal, M.; Colas, R.A.; Mazzacuva, F.; Lokhande, R.; Dharmshale, S.; Bharadwaj, R.; et al. Lipid mediators of inflammation and Resolution in individuals with tuberculosis and tuberculosis-Diabetes. Prostaglandins Other Lipid Mediat. 2020, 147, 106398. [Google Scholar] [CrossRef]
- Polus, A.; Zapala, B.; Razny, U.; Gielicz, A.; Kiec-Wilk, B.; Malczewska-Malec, M.; Sanak, M.; Childs, C.E.; Calder, P.C.; Dembinska-Kiec, A. Omega-3 fatty acid supplementation influences the whole blood transcriptome in women with obesity, associated with pro-resolving lipid mediator production. Biochim. Biophys. Acta 2016, 1861, 1746–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barden, A.; Shinde, S.; Tsai, I.J.; Croft, K.D.; Beilin, L.J.; Puddey, I.B.; Mori, T.A. Effect of weight loss on neutrophil resolvins in the metabolic syndrome. Prostaglandins Leukot. Essent. Fatty Acids 2019, 148, 25–29. [Google Scholar] [CrossRef]
- Lopez-Vicario, C.; Titos, E.; Walker, M.E.; Alcaraz-Quiles, J.; Casulleras, M.; Duran-Guell, M.; Flores-Costa, R.; Perez-Romero, N.; Forne, M.; Dalli, J.; et al. Leukocytes from obese individuals exhibit an impaired SPM signature. FASEB J. 2019, 33, 7072–7083. [Google Scholar] [CrossRef] [Green Version]
- Schulte, F.; Asbeutah, A.A.; Benotti, P.N.; Wood, G.C.; Still, C.; Bistrian, B.R.; Hardt, M.; Welty, F.K. The relationship between specialized pro-resolving lipid mediators, morbid obesity and weight loss after bariatric surgery. Sci. Rep. 2020, 10, 20128. [Google Scholar] [CrossRef]
- Xia, F.; He, C.; Ren, M.; Xu, F.G.; Wan, J.B. Quantitative profiling of eicosanoids derived from n-6 and n-3 polyunsaturated fatty acids by twin derivatization strategy combined with LC-MS/MS in patients with type 2 diabetes mellitus. Anal. Chim. Acta 2020, 1120, 24–35. [Google Scholar] [CrossRef]
- Sano, Y.; Toyoshima, S.; Miki, Y.; Taketomi, Y.; Ito, M.; Lee, H.; Saito, S.; Murakami, M.; Okayama, Y. Activation of inflammation and resolution pathways of lipid mediators in synovial fluid from patients with severe rheumatoid arthritis compared with severe osteoarthritis. Asia Pac. Allergy 2020, 10, e21. [Google Scholar] [CrossRef]
- Song, J.; Sun, R.; Zhang, Y.; Fu, Y.; Zhao, D. Role of the Specialized Pro-resolving Mediator Resolvin D1 in Hashimoto’s Thyroiditis. Exp. Clin. Endocrinol. Diabetes 2021, 129, 791–797. [Google Scholar] [CrossRef] [PubMed]
- See, V.H.L.; Mas, E.; Prescott, S.L.; Beilin, L.J.; Burrows, S.; Barden, A.E.; Huang, R.C.; Mori, T.A. Effects of postnatal omega-3 fatty acid supplementation on offspring pro-resolving mediators of inflammation at 6 months and 5 years of age: A double blind, randomized controlled clinical trial. Prostaglandins Leukot. Essent. Fatty Acids 2017, 126, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Lamon-Fava, S.; So, J.; Mischoulon, D.; Ziegler, T.R.; Dunlop, B.W.; Kinkead, B.; Schettler, P.J.; Nierenberg, A.A.; Felger, J.C.; Maddipati, K.R.; et al. Dose- and time-dependent increase in circulating anti-inflammatory and pro-resolving lipid mediators following eicosapentaenoic acid supplementation in patients with major depressive disorder and chronic inflammation. Prostaglandins Leukot. Essent. Fatty Acids 2021, 164, 102219. [Google Scholar] [CrossRef] [PubMed]
- So, J.; Wu, D.; Lichtenstein, A.H.; Tai, A.K.; Matthan, N.R.; Maddipati, K.R.; Lamon-Fava, S. EPA and DHA differentially modulate monocyte inflammatory response in subjects with chronic inflammation in part via plasma specialized pro-resolving lipid mediators: A randomized, double-blind, crossover study. Atherosclerosis 2021, 316, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Norling, L.V.; Headland, S.E.; Dalli, J.; Arnardottir, H.H.; Haworth, O.; Jones, H.R.; Irimia, D.; Serhan, C.N.; Perretti, M. Proresolving and cartilage-protective actions of resolvin D1 in inflammatory arthritis. JCI Insight 2016, 1, e85922. [Google Scholar] [CrossRef] [PubMed]
- Arnardottir, H.H.; Dalli, J.; Norling, L.V.; Colas, R.A.; Perretti, M.; Serhan, C.N. Resolvin D3 Is Dysregulated in Arthritis and Reduces Arthritic Inflammation. J. Immunol. 2016, 197, 2362–2368. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.; Chen, H.; Li, Y.; Zhong, H.; Sun, W.; Wang, J.; Zhang, T.; Ma, J.; Yan, S.; Zhang, J.; et al. Maresin 1 improves the Treg/Th17 imbalance in rheumatoid arthritis through miR-21. Ann. Rheum. Dis. 2018, 77, 1644–1652. [Google Scholar] [CrossRef]
- Allen, B.L.; Montague-Cardoso, K.; Simeoli, R.; Colas, R.A.; Oggero, S.; Vilar, B.; McNaughton, P.A.; Dalli, J.; Perretti, M.; Sher, E.; et al. Imbalance of proresolving lipid mediators in persistent allodynia dissociated from signs of clinical arthritis. Pain 2020, 161, 2155–2166. [Google Scholar] [CrossRef]
- Ozgul Ozdemir, R.B.; Soysal Gunduz, O.; Ozdemir, A.T.; Akgul, O. Low levels of pro-resolving lipid mediators lipoxin-A4, resolvin-D1 and resolvin-E1 in patients with rheumatoid arthritis. Immunol. Lett. 2020, 227, 34–40. [Google Scholar] [CrossRef]
- Merched, A.J.; Ko, K.; Gotlinger, K.H.; Serhan, C.N.; Chan, L. Atherosclerosis: Evidence for impairment of resolution of vascular inflammation governed by specific lipid mediators. FASEB J. 2008, 22, 3595–3606. [Google Scholar] [CrossRef]
- Ho, K.J.; Spite, M.; Owens, C.D.; Lancero, H.; Kroemer, A.H.; Pande, R.; Creager, M.A.; Serhan, C.N.; Conte, M.S. Aspirin-triggered lipoxin and resolvin E1 modulate vascular smooth muscle phenotype and correlate with peripheral atherosclerosis. Am. J. Pathol. 2010, 177, 2116–2123. [Google Scholar] [CrossRef] [PubMed]
- Viola, J.R.; Lemnitzer, P.; Jansen, Y.; Csaba, G.; Winter, C.; Neideck, C.; Silvestre-Roig, C.; Dittmar, G.; Doring, Y.; Drechsler, M.; et al. Resolving Lipid Mediators Maresin 1 and Resolvin D2 Prevent Atheroprogression in Mice. Circ. Res. 2016, 119, 1030–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colas, R.A.; Souza, P.R.; Walker, M.E.; Burton, M.; Zaslona, Z.; Curtis, A.M.; Marques, R.M.; Dalli, J. Impaired Production and Diurnal Regulation of Vascular RvDn-3 DPA Increase Systemic Inflammation and Cardiovascular Disease. Circ. Res. 2018, 122, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Aoki-Saito, H.; Fukuda, H.; Ikeda, H.; Koga, Y.; Yatomi, M.; Tsurumaki, H.; Maeno, T.; Saito, T.; Nakakura, T.; et al. Resolvin E3 attenuates allergic airway inflammation via the interleukin-23-interleukin-17A pathway. FASEB J. 2019, 33, 12750–12759. [Google Scholar] [CrossRef] [Green Version]
- Flesher, R.P.; Herbert, C.; Kumar, R.K. Resolvin E1 promotes resolution of inflammation in a mouse model of an acute exacerbation of allergic asthma. Clin. Sci. 2014, 126, 805–814. [Google Scholar] [CrossRef] [Green Version]
- Rogerio, A.P.; Haworth, O.; Croze, R.; Oh, S.F.; Uddin, M.; Carlo, T.; Pfeffer, M.A.; Priluck, R.; Serhan, C.N.; Levy, B.D. Resolvin D1 and aspirin-triggered resolvin D1 promote resolution of allergic airways responses. J. Immunol. 2012, 189, 1983–1991. [Google Scholar] [CrossRef] [Green Version]
- Zambalde, E.P.; Teixeira, M.M.; Favarin, D.C.; de Oliveira, J.R.; Magalhaes, M.L.; Cunha, M.M.; Silva, W.C.J.; Okuma, C.H.; Rodrigues, V.J.; Levy, B.D.; et al. The anti-inflammatory and pro-resolution effects of aspirin-triggered RvD1 (AT-RvD1) on peripheral blood mononuclear cells from patients with severe asthma. Int. Immunopharmacol. 2016, 35, 142–148. [Google Scholar] [CrossRef]
- Miyata, J.; Fukunaga, K.; Iwamoto, R.; Isobe, Y.; Niimi, K.; Takamiya, R.; Takihara, T.; Tomomatsu, K.; Suzuki, Y.; Oguma, T.; et al. Dysregulated synthesis of protectin D1 in eosinophils from patients with severe asthma. J. Allergy Clin. Immunol. 2013, 131, 353–360. [Google Scholar] [CrossRef]
- Hottz, E.D.; Azevedo-Quintanilha, I.G.; Palhinha, L.; Teixeiraet, L.; Barreto, E.A.; Pão, C.R.R.; Righy, C.; Franco, S.; Souza, T.M.L.; Kurtz, P.; et al. Platelet activation and platelet-monocyte aggregate formation trigger tissue factor expression in patients with severe COVID-19. Blood 2020, 136, 1330–1341. [Google Scholar] [CrossRef]
- Sun, Y.P.; Oh, S.F.; Uddin, J.; Yang, R.; Gotlinger, K.; Campbell, E.; Colgan, S.P.; Petasis, N.A.; Serhan, C.N. Resolvin D1 and its aspirin-triggered 17R epimer. Stereochemical assignments, anti-inflammatory properties, and enzymatic inactivation. J. Biol. Chem. 2007, 282, 9323–9334. [Google Scholar] [CrossRef] [Green Version]
- Chiang, N.; Bermudez, E.A.; Ridker, P.M.; Hurwitz, S.; Serhan, C.N. Aspirin triggers antiinflammatory 15-epi-lipoxin A4 and inhibits thromboxane in a randomized human trial. Proc. Natl. Acad. Sci. USA 2004, 101, 15178–15183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, N.; Hurwitz, S.; Ridker, P.M.; Serhan, C.N. Aspirin has a gender-dependent impact on antiinflammatory 15-epi-lipoxin A4 formation: A randomized human trial. Arterioscler. Thromb. Vasc. Biol. 2006, 26, e14–e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bali, S.K.; Madaiah, H.; Dharmapalan, J.; Janarthanam, S.; Tarannum, F. Effect of systemic long-term, low-dose aspirin on periodontal status and soluble CD14 in gingival crevicular fluid: A case-control study. J. Investig. Clin. Dent. 2018, 9, e12353. [Google Scholar] [CrossRef]
- Walker, M.E.; Souza, P.R.; Colas, R.A.; Dalli, J. 13-Series resolvins mediate the leukocyte-platelet actions of atorvastatin and pravastatin in inflammatory arthritis. FASEB J. 2017, 31, 3636–3648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyrillou, K.; Chairakaki, A.D.; Tamvakopoulos, C.; Andreakos, E. Dexamethasone induces omega3-derived immunoresolvents driving resolution of allergic airway inflammation. J. Allergy Clin. Immunol. 2018, 142, 691–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdulrazaq, M.; Innes, J.K.; Calder, P.C. Effect of omega-3 polyunsaturated fatty acids on arthritic pain: A systematic review. Nutrition 2017, 39, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Yates, C.M.; Calder, P.C.; Ed Rainger, G. Pharmacology and therapeutics of omega-3 polyunsaturated fatty acids in chronic inflammatory disease. Pharmacol. Ther. 2014, 141, 272–282. [Google Scholar] [CrossRef]
- Fiala, M.; Halder, R.C.; Sagong, B.; Ross, O.; Sayre, J.; Porter, V.; Bredesen, D.E. omega-3 Supplementation increases amyloid-beta phagocytosis and resolvin D1 in patients with minor cognitive impairment. FASEB J. 2015, 29, 2681–2689. [Google Scholar] [CrossRef] [Green Version]
- Terrando, N.; Gomez-Galan, M.; Yang, T.; Carlstrom, M.; Gustavsson, D.; Harding, R.E.; Lindskog, M.; Eriksson, L.I. Aspirin-triggered resolvin D1 prevents surgery-induced cognitive decline. FASEB J. 2013, 27, 3564–3571. [Google Scholar] [CrossRef] [Green Version]
- Barden, A.E.; Shinde, S.; Burke, V.; Puddey, I.B.; Beilin, L.J.; Irish, A.B.; Watts, G.F.; Mori, T.A. The effect of n-3 fatty acids and coenzyme Q10 supplementation on neutrophil leukotrienes, mediators of inflammation resolution and myeloperoxidase in chronic kidney disease. Prostaglandins Other Lipid Mediat. 2018, 136, 1–8. [Google Scholar] [CrossRef]
- Souza, P.R.; Marques, R.M.; Gomez, E.A.; Colas, R.A.; De Matteis, R.; Zak, A.; Patel, M.; Collier, D.J.; Dalli, J. Enriched Marine Oil Supplements Increase Peripheral Blood Specialized Pro-Resolving Mediators Concentrations and Reprogram Host Immune Responses: A Randomized Double-Blind Placebo-Controlled Study. Circ. Res. 2020, 126, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Claria, J.; Dalli, J.; Yacoubian, S.; Gao, F.; Serhan, C.N. Resolvin D1 and resolvin D2 govern local inflammatory tone in obese fat. J. Immunol. 2012, 189, 2597–2605. [Google Scholar] [CrossRef] [PubMed]
- Colas, R.A.; Dalli, J.; Chiang, N.; Vlasakov, I.; Sanger, J.M.; Riley, I.R.; Serhan, C.N. Identification and Actions of the Maresin 1 Metabolome in Infectious Inflammation. J. Immunol. 2016, 197, 4444–4452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaller, M.S.; Chen, M.; Colas, R.A.; Sorrentino, T.A.; Lazar, A.A.; Grenon, S.M.; Dalli, J.; Conte, M.S. Treatment With a Marine Oil Supplement Alters Lipid Mediators and Leukocyte Phenotype in Healthy Patients and Those With Peripheral Artery Disease. J. Am. Heart Assoc. 2020, 9, e016113. [Google Scholar] [CrossRef]
- Mamoshina, P.; Volosnikova, M.; Ozerov, I.V.; Putin, E.; Skibina, E.; Cortese, F.; Zhavoronkov, A. Machine Learning on Human Muscle Transcriptomic Data for Biomarker Discovery and Tissue-Specific Drug Target Identification. Front. Genet. 2018, 9, 242. [Google Scholar] [CrossRef]
- Bishop, C.M. Pattern Recognition and Machine Learning; Springer: New York, NY, USA, 2006; p. 738. [Google Scholar]
- Swan, A.L.; Mobasheri, A.; Allaway, D.; Liddell, S.; Bacardit, J. Application of machine learning to proteomics data: Classification and biomarker identification in postgenomics biology. OMICS 2013, 17, 595–610. [Google Scholar] [CrossRef] [Green Version]
- Zitnik, M.; Nguyen, F.; Wang, B.; Leskovec, J.; Goldenberg, A.; Hoffman, M.M. Machine Learning for Integrating Data in Biology and Medicine: Principles, Practice, and Opportunities. Inf. Fusion 2019, 50, 71–91. [Google Scholar] [CrossRef]
- Valletta, J.J.; Recker, M. Identification of immune signatures predictive of clinical protection from malaria. PLoS Comput. Biol. 2017, 13, e1005812. [Google Scholar] [CrossRef] [Green Version]
- James, G.; Witten, D.; Hastie, T.; Tibshirani, R. An Introduction to Statistical Learning: With Applications in R, 6th ed.; Springer: Berlin/Heidelberg, Germany, 2013; p. 426. [Google Scholar]
- Maglogiannis, I.G. Emerging Artificial Intelligence Applications in Computer Engineering: Real Word AI Systems with Applications in eHealth, HCI, Information Retrieval and Pervasive Technologies; IOS Press: Washington, DC, USA, 2007; p. 407. [Google Scholar]
- Bennett, K.P.; Campbell, C. Support vector machines. ACM SIGKDD Explor. Newsletter 2000, 2, 1–13. [Google Scholar] [CrossRef]
- Gordon, A.D.; Breiman, L.; Friedman, J.H.; Olshen, R.A.; Stone, C.J. Classification and Regression Trees. Biometrics 1984, 40, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Rathod, K.S.; Kapil, V.; Velmurugan, S.; Khambata, R.S.; Siddique, U.; Khan, S.; Van Eijl, S.; Gee, L.C.; Bansal, J.; Pitrola, K.; et al. Accelerated resolution of inflammation underlies sex differences in inflammatory responses in humans. J. Clin. Investig. 2017, 127, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Troisi, F.; Pace, S.; Jordan, P.M.; Meyer, K.P.L.; Bilancia, R.; Ialenti, A.; Borrelli, F.; Rossi, A.; Sautebin, L.; Serhan, C.N.; et al. Sex Hormone-Dependent Lipid Mediator Formation in Male and Female Mice During Peritonitis. Front. Pharmacol. 2021, 12, 818544. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
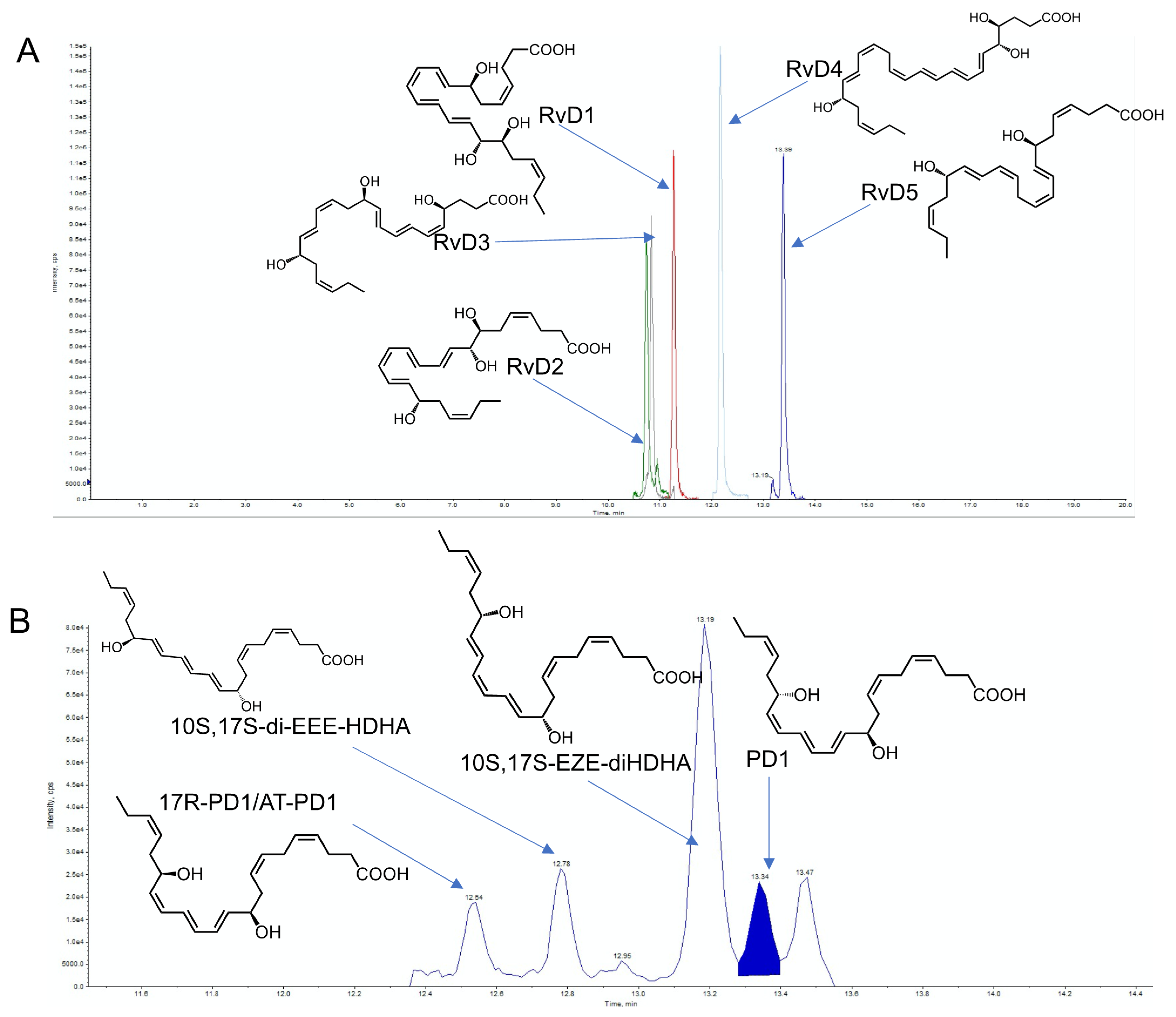
| Substrate | SPM Families | Abbreviation | Chemical Name | Lipid Maps LM ID | PubChem CID |
|---|---|---|---|---|---|
| DHA | D-series Resolvins | RvD1 | 7S,8R,17S-trihydroxy-4Z,9E,11E,13Z,15E,19Z-docosahexaenoic acid | LMFA04030011 | 44251266 |
| RvD2 | 7S,16R,17S-trihydroxy-4Z,8E,10Z,12E,14E,19Z-docosahexaenoic acid | LMFA04030001 | 11383310 | ||
| RvD3 | 4S,11R,17S-trihydroxy-5Z,7E,9E,13Z,15E,19Z-docosahexaenoic acid | LMFA04030012 | 71665428 | ||
| RvD4 | 4S,5R,17S-trihydroxydocosa-6E,8E,10Z,13Z,15E,19Z hexaenoic acid | LMFA04030002 | 16061138 | ||
| RvD5 | 7S,17S-dihydroxy-4Z,8E,10Z,13Z,15Z,19E-docosahexaenoic acid | LMFA04030003 | 16061139 | ||
| RvD6 | 4S,17S-dihydroxy-5E,7E,10Z,13Z,15E,19Z-docosahexaenoic acid | LMFA04030004 | 25073193 | ||
| 17R-RvD1 | 7S,8R,17R-trihydroxy-4Z,9E,11E,13Z,15E,19Z-docosahexaenoic acid | ||||
| 17R-RvD3 | 4S,11R,17R-trihydroxy-5Z,7E,9E,13Z,15E,19Z-docosahexaenoic acid | ||||
| Protectins | NPD1/PD1 | 10R,17S-dihydroxy-4Z,7Z,11E,13E,15Z,19Z- docosahexaenoic acid | LMFA04040001 | 16042541 | |
| PDX | 10S,17S-dihydroxy-4Z,7Z,11E,13Z,15E,19Z-docosahexaenoic acid | LMFA04040003 | 11667655 | ||
| 17R-PD1 | 10R,17R-dihydroxy-4Z,7Z,11E,13E,15Z,19Z-docosahexaenoic acid | 132282528 | |||
| 22-OH-PD1 | 10R,17S,22-trihydroxy-4Z,7Z,11E,13E,15Z,19Z-docosahexaenoic acid | 132472333 | |||
| cys-Protectins | PCTR1 | 16R-glutathionyl-17S-hydroxy-4Z,7Z,10Z,12E,14E,19Z-docosahexaenoic acid | LMFA04040004 | 132472316 | |
| PCTR2 | 16R-cysteinylglycinyl-17S-hydroxy-4Z,7Z,10Z,12E,14E,19Z-docosahexaenoic acid | LMFA04040005 | 132472317 | ||
| PCTR3 | 16R-cysteinyl-17S-hydroxy-4Z,7Z,10Z,12E,14E,19Z-docosahexaenoic acid | LMFA04040006 | 132472318 | ||
| Maresins | MaR1 | 7R,14S-dihydroxy-4Z,8E,10E,12Z,16Z,19Z- docosahexaenoic acid | LMFA04050001 | 60201795 | |
| MaR2 | 13R,14S-dihydroxy-4Z,7Z,9E,11E,16Z,19Z- docosahexaenoic acid | LMFA04050004 | 101894912 | ||
| cys-Maresins | MCTR1 | 13R-glutathionyl-14S-hydroxy-4Z,7Z,9E,11E,16Z,19Z-docosahexaenoic acid | LMFA04050005 | 122368871 | |
| MCTR2 | 13R-cysteinylglycinyl-14S-hydroxy-4Z,7Z,9E,11E,16Z,19Z-docosahexaenoic acid | LMFA04050006 | 122368872 | ||
| MCTR3 | 13R-cysteinyl-14S-hydroxy-4Z,7Z,9E,11E,16Z,19Z-docosahexaenoic acid | LMFA04050007 | 122368873 | ||
| n-3 DPA | 13-series Resolvins | RvT1 | 7S,13R,20S-trihydroxy-8E,10Z,14E,16Z,18E-docosapentaenoic acid | LMFA04000091 | 124202379 |
| RvT2 | 7,12,13-trihydroxy-8,10,14,16,19-docosapentaenoic acid | LMFA04000092 | 124202381 | ||
| RvT3 | 7,8,13-trihydroxy-9,11,14,16,19-docosapentaenoic acid | LMFA04000093 | 124202383 | ||
| RvT4 | 7S,13R-dihydroxy-8E,10Z,14E,16Z,19Z-docosapentaenoic acid | LMFA04000094 | 124202385 | ||
| D-series | RvD1n-3 DPA | 7S,8R,17S-trihydroxy-9E,11E,13Z,15E,19Z-docosapentaenoic acid | 132472356 | ||
| RvD2n-3 DPA | 7,16,17-trihydroxy-8,10,12,14,19-docosapentaenoic acid | 132472324 | |||
| RvD5n-3 DPA | 7S,17S-dihydroxy-8E,10Z,13Z,15Z,19E-docosapentaenoic acid | 132472358 | |||
| Protectins | PD1n-3 DPA | 10R,17S-dihydroxy-7Z,11E,13E,15Z,19Z-docosapentaenoic acid | LMFA04000096 | 132472351 | |
| PD2n-3 DPA | 16,17-dihydroxy-7Z,10,13,14,19-docosapentaenoic acid | LMFA04000097 | 132472319 | ||
| Maresins | MaR1n-3 DPA | 7R,14S-dihydroxy-8E,10E,12Z,16Z,19Z-docosapentaenoic acid | |||
| MaR2n-3 DPA | 13,14-dihydroxy-7,9,11,16,19-docosapentaenoic acid | ||||
| EPA | E-series Resolvin | RvE1 | 5S,12R,18R-trihydroxy-6Z,8E,10E,14Z,16E-eicosapentaenoic acid | LMFA03140003 | 10473088 |
| RvE2 | 5S,18R-dihydroxy-6E,8Z,11Z,14Z,16E-eicosapentaenoic acid | LMFA03140011 | 16061125 | ||
| RvE3 | 17R,18R-dihydroxy-5Z,8Z,11Z,13E,15E-eicosapentaenoic acid | LMFA03140006 | 60150429 | ||
| RvE4 | 5S,15S-dihydroxy-6E,8Z,11Z,13E,17Z-eicosapentaenoic acid | ||||
| AA | Lipoxins | LXA4 | 5S,6R,15S-trihydroxy-7E,9E,11Z,13E-eicosatetraenoic acid | LMFA03040001 | 5280914 |
| LXB4 | 5S,14R,15S-trihydroxy-6E,8Z,10E,12E-eicosatetraenoic acid | LMFA03040002 | 5280915 | ||
| 15-epi-LXA4 | 5S,6R,15R-trihydroxy-7E,9E,11Z,13E-eicosatetraenoic acid | LMFA03040010 | 9841438 | ||
| 15-epi-LXB4 | 5S,14R,15R-trihydroxy-6E,8Z,10E,12E-eicosatetraenoic acid | LMFA03040007 | 70678885 |
| Disease | SPM Identified | Tissue | Method | LC Solvent System | MS/MS Ionization Mode | References | |
|---|---|---|---|---|---|---|---|
| Neuronal Inflammation | Ischemic brain injury | LXA4 | Plasma | ELISA | N/A | N/A | [74] |
| Alzheimer’s disease | LXA4, RvD1, MaR1, 5,15-diHETE | CSF, Hippocampus | ELISA, LC-MS/MS | methanol/water/acetic acid | Negative | [75] | |
| RvD5, PD1, MaR1, | Entorhinal cortex tissue | LC-MS/MS | methanol/water/acetic acid | Negative | [76] | ||
| Cardiovascular diseases | Myocardial infarction | PD1, 10S,17S-diHDHA, AT-PD1, LXA4, AT-LXA4, RvD5n-3 DPA, PD2n-3 DPA, 10S,17S-diHDPA | Plasma | LC-MS/MS | methanol/water/acetic acid | Negative | [77] |
| PD1, RvE1 | Plasma | LC-MS/MS | Formic acid/water/acetonitrile | Negative | [78] | ||
| Peripheral Artery Disease | LXA4 | Plasma | LC-MS/MS | methanol/water/acetic acid | Negative | [79] | |
| 10,17-diHDHA | Plasma | LC-MS/MS | methanol/water/acetic acid | Negative | [80] | ||
| Atherosclerosis | RvD1, 7-epi,Δ12-trans-MaR1, | Human carotid atherosclerotic plaques | LC-MS/MS | methanol/water/acetic acid | Negative | [81] | |
| Infections | Infection, low-dose endotoxin | RvD1, RvE1 | Plasma and serum | LC-MS/MS | methanol/water/acetic acid and acetonitrile | Negative | [59] |
| Periodontal disease | PD1, MaR1, LXA4 | Saliva | ELISA | N/A | N/A | [82] | |
| Rhinosinusitis | RvD1, RvD2, LXA4 | Ethmoid sinus tissue | LC-MS/MS | Formic acid/water/acetonitrile | Negative | [83] | |
| Tuberculosis | RvD2, 17R-PD1, PCTR3, PD1n-3 DPA, RvE3, 15-oxo-LXA4 | Serum | LC-MS/MS | methanol/water/acetic acid | Positive | [84] | |
| Sepsis | RvD5, 17R-RvD1, PD1, 17R-PD1, RvE1, RvE2, RvE3, 5S,15S-diHETE, | Plasma | LC-MS/MS | methanol/water/acetic acid | Negative | [60] | |
| Metabolic disease | Obesity | RvD1, RvD2, 10S,17S-diHDHA, LXA4, LXA5 | Plasma | ELISA | N/A | N/A | [85] |
| RvE1 | Ex vivo neutrophil stimulation | LC-MS/MS | methanol/water/acetic acid | Negative | [86] | ||
| RvD2, RvD4, LXA4, RvE3 | Plasma | LC-MS/MS | methanol/water/acetic acid | Negative | [87] | ||
| RvD3, RvD4, PD1, MaR1 | Serum | LC-MS/MS | methanol/water/acetic acid | Negative | [88] | ||
| Diabetes | RvD1, RvE1 | Plasma | LC-MS/MS | Formic acid/water/acetonitrile | Positive | [89] | |
| RvD2, 17R-PD1, PCTR3, PD1n-3 DPA, RvE3, 15-oxo-LXA4 | Serum | LC-MS/MS | methanol/water/acetic acid | Positive | [84] | ||
| Autoimmune disease | Rheumatoid Arthritis | RvD1, 17R-RvD1, RvD2, PD1, 10S,17S-diHDHA, MaR1, RvE1, RvE2, RvE3, 18R-RvE3 | Plasma and synovial fluid | LC-MS/MS | ammonium acetate/methanol | Negative | [41] |
| RvD1, RvD2, RvD3, RvD4, 10S,17S-diHDHA, 17R-PD1, MaR1, 4S,14S-diHDHA, 10S,17S-diHDPA, MaR1n-3 DPA, 15R-LXA4, | Plasma | LC-MS/MS | methanol/water/acetic acid | Negative | [40] | ||
| PD1, LXA4, LXB4 | Synovial fluid | LC-MS/MS | acetonitrile/methanol/water | Negative | [90] | ||
| Osteoarthritis | PD1, LXA4, LXB4 | Synovial fluid | LC-MS/MS | acetonitrile/methanol/water | Negative | [90] | |
| Hashimoto’s Thyroiditis | RvD1 | Serum | ELISA | [91] | |||
| Omega-3 Supplementation | Healthy volunteers | RvD1, 17R-RvD1, RvD2, RvE1, RvE2, RvE3, 18R-RvE3 | Plasma | LC-MS/MS | methanol/water/acetic acid | Negative | [92] |
| Healthy volunteers and patients with periphery artery disease | LXA4 | Plasma | LC-MS/MS | methanol/water/acetic acid | Negative | [79] | |
| Periphery artery disease | PDX (10S,17S-diHDHA) | Plasma | LC-MS/MS | methanol/water/acetic acid | Negative | [80] | |
| Major depressive disorder and chronic inflammation | RvE2, RvE3, LXB4 | Plasma | LC-MS/MS | acetonitrile/methanol/water | Negative | [93] | |
| Chronic inflammation | RvD5n-3 DPA, MaR1n-3 DPA | Plasma | LC-MS/MS | acetonitrile/methanol/water | Negative | [94] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dalli, J.; Gomez, E.A.; Jouvene, C.C. Utility of the Specialized Pro-Resolving Mediators as Diagnostic and Prognostic Biomarkers in Disease. Biomolecules 2022, 12, 353. https://doi.org/10.3390/biom12030353
Dalli J, Gomez EA, Jouvene CC. Utility of the Specialized Pro-Resolving Mediators as Diagnostic and Prognostic Biomarkers in Disease. Biomolecules. 2022; 12(3):353. https://doi.org/10.3390/biom12030353
Chicago/Turabian StyleDalli, Jesmond, Esteban Alberto Gomez, and Charlotte Camille Jouvene. 2022. "Utility of the Specialized Pro-Resolving Mediators as Diagnostic and Prognostic Biomarkers in Disease" Biomolecules 12, no. 3: 353. https://doi.org/10.3390/biom12030353