
The Fibrinogen-like Domain of ANGPTL3 Facilitates Lipolysis in 3T3-L1 Cells by Activating the Intracellular Erk Pathway

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
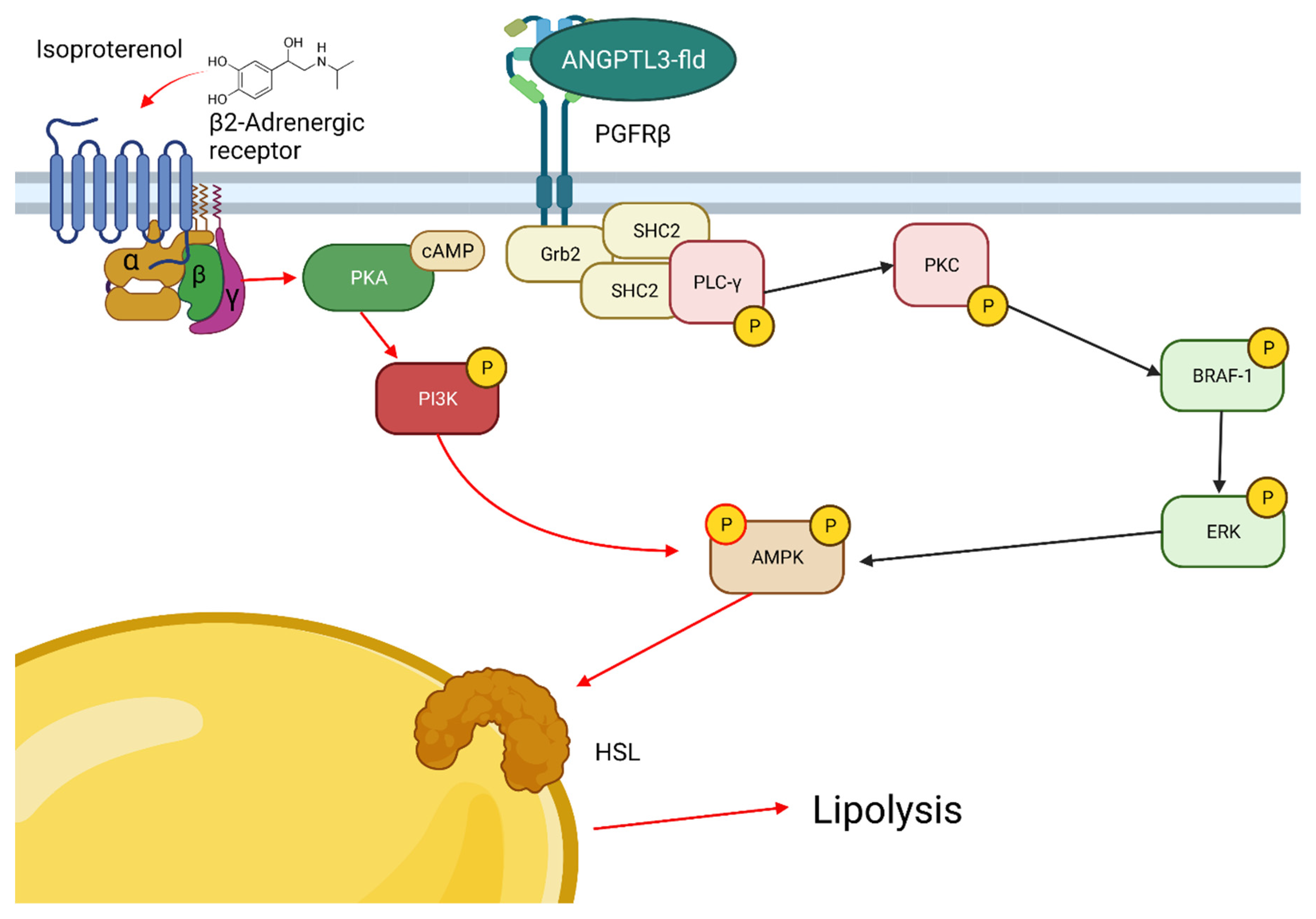
Abstract
:1. Introduction
2. Material and Methods
2.1. Cell Culture and Treatments
2.2. Quantification of Free Fatty Acid Concentrations
2.3. Western Blot Analysis
2.4. Human Phospho-Kinase Array
2.5. Statistical Analysis
3. Results
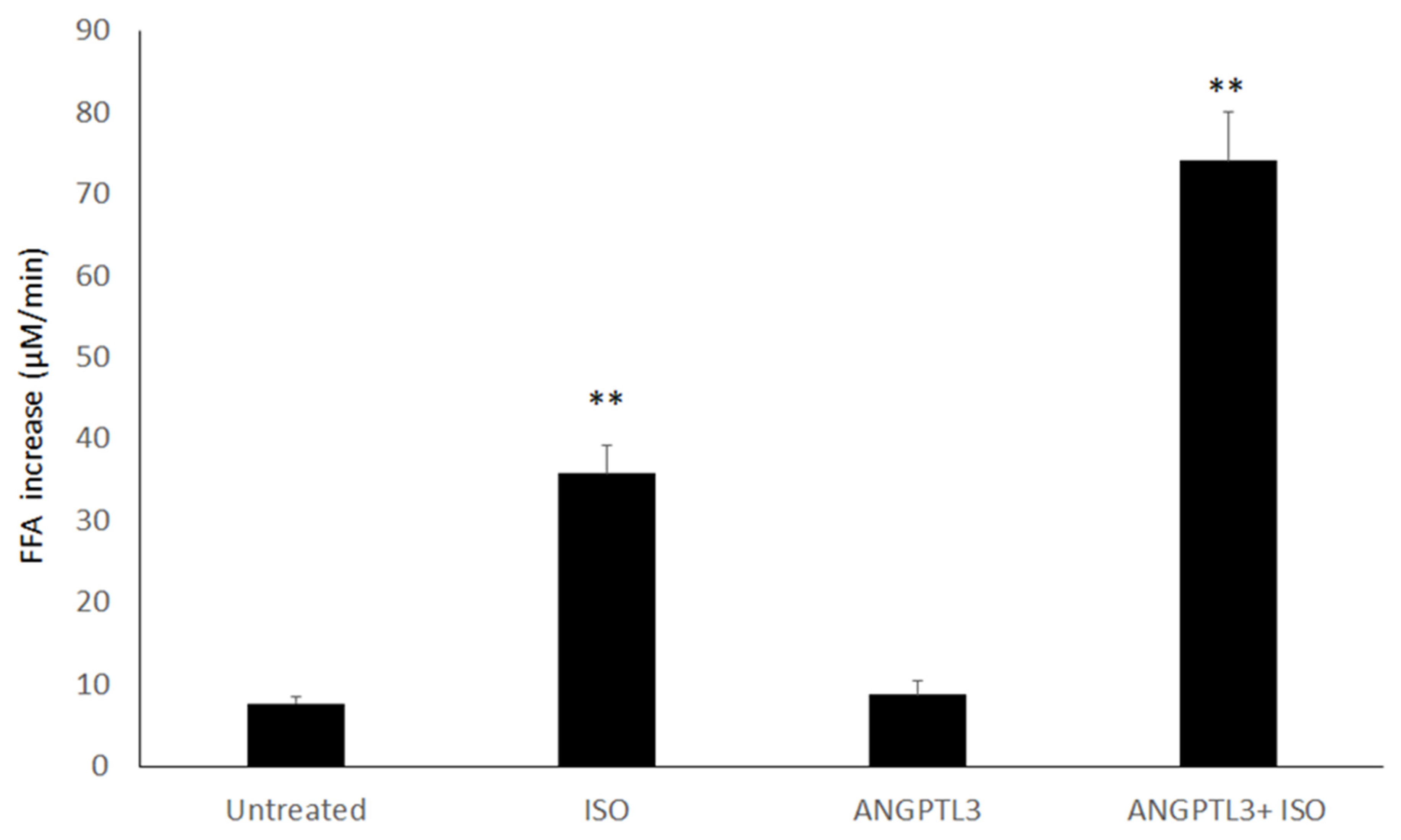
3.1. ANGPTL3-Fld Enhances the Lipolysis in 3T3-L1 Cells Treated with Isoproterenol
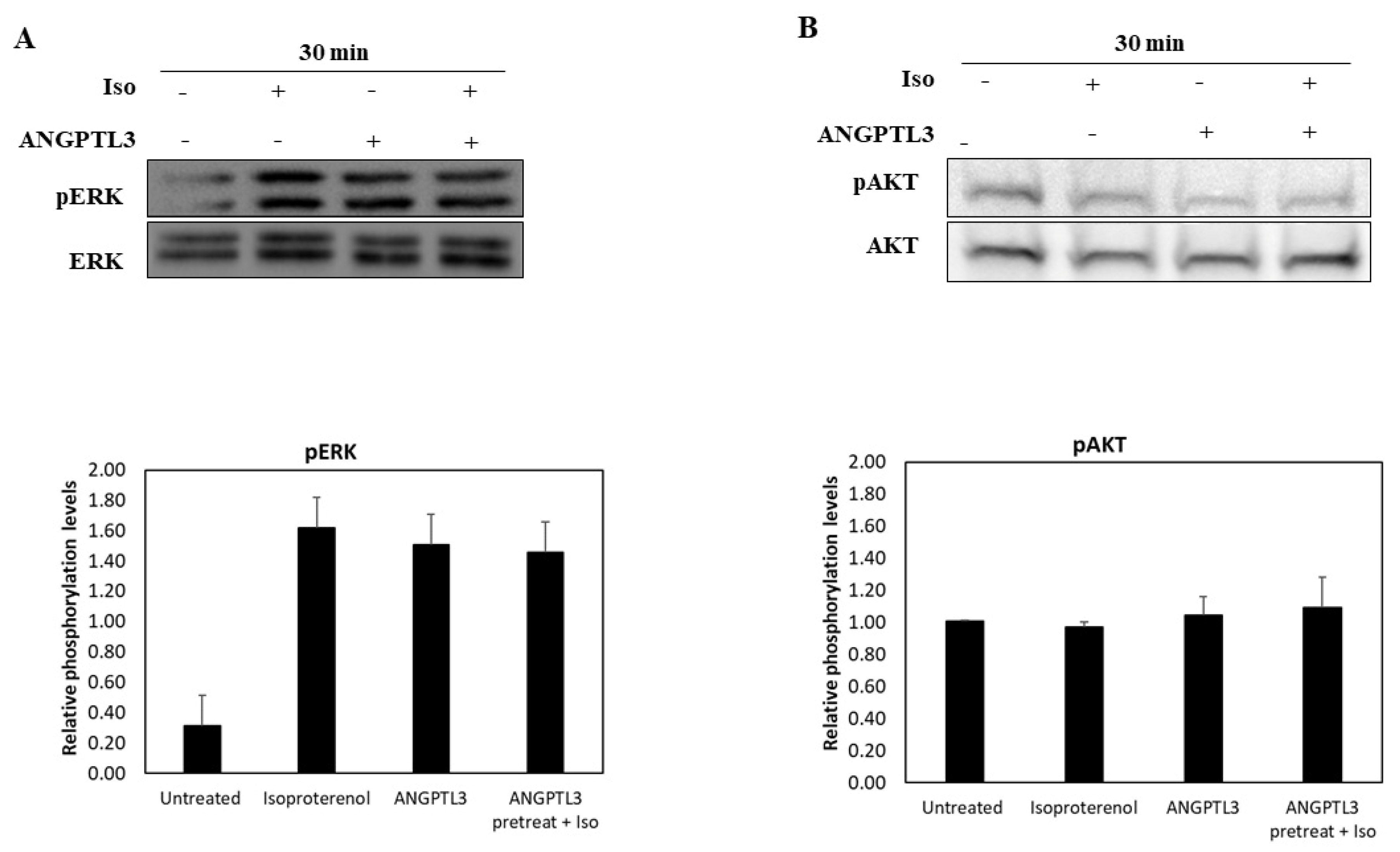
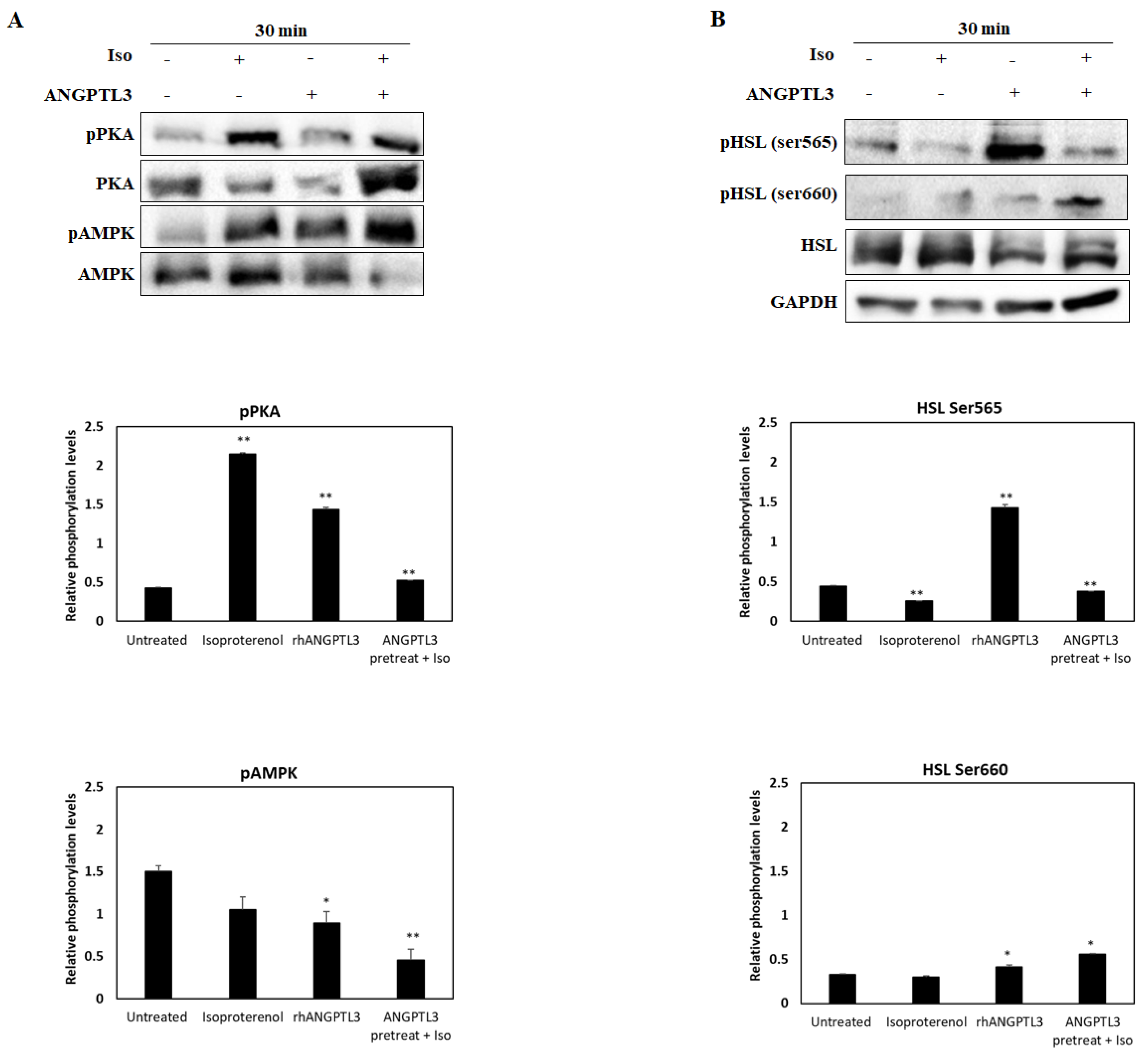
3.2. ANGPTL3-Fld Facilitates Lipolysis Mainly Activating the ERK Arm of the MAPK Pathway
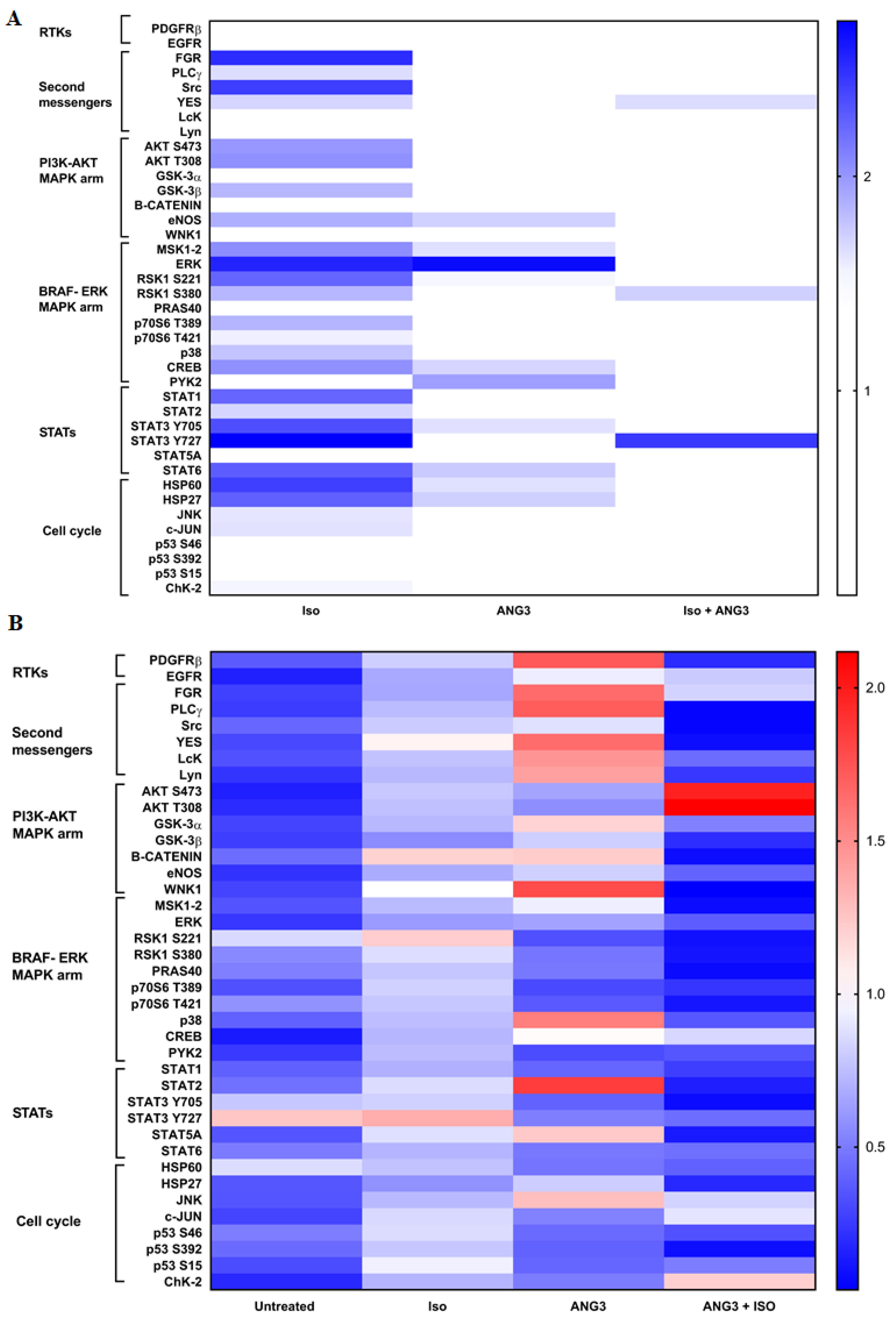
3.3. ANGPTL3-Fld Activates the PDGFRβ Receptor
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bini, S.; D’erasmo, L.; Di Costanzo, A.; Minicocci, I.; Pecce, V.; Arca, M. The Interplay between Angiopoietin-Like Proteins and Adipose Tissue: Another Piece of the Relationship between Adiposopathy and Cardiometabolic Diseases? Int. J. Mol. Sci. 2021, 22, 742. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, K. An updated ANGPTL3-4-8 model as a mechanism of triglyceride partitioning between fat and oxidative tissues. Prog. Lipid Res. 2021, 85, 101140. [Google Scholar] [CrossRef] [PubMed]
- Koishi, R.; Ando, Y.; Ono, M.; Shimamura, M.; Yasumo, H.; Fujiwara, T.; Horikoshi, H.; Furukawa, H. Angptl3 regulates lipid metabolism in mice. Nat. Genet. 2002, 30, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Arca, M.; D’Erasmo, L.; Minicocci, I. Familial combined hypolipidemia: Angiopoietin-like protein-3 deficiency. Curr. Opin. Lipidol. 2020, 31, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Robciuc, M.R.; Maranghi, M.; Lahikainen, A.; Rader, D.; Bensadoun, A.; Öörni, K.; Metso, J.; Minicocci, I.; Ciociola, E.; Ceci, F.; et al. Angptl3 Deficiency Is Associated With Increased Insulin Sensitivity, Lipoprotein Lipase Activity, and Decreased Serum Free Fatty Acids. Arter. Thromb. Vasc. Biol. 2013, 33, 1706–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, M.; Shimizugawa, T.; Shimamura, M.; Yoshida, K.; Noji-Sakikawa, C.; Ando, Y.; Koishi, R.; Furukawa, H. Protein Region Important for Regulation of Lipid Metabolism in Angiopoietin-like 3 (ANGPTL3): ANGPTL3 is cleaved and activated in vivo. J. Biol. Chem. 2003, 278, 41804–41809. [Google Scholar] [CrossRef] [Green Version]
- Tikkanen, E.; Minicocci, I.; Hällfors, J.; Di Costanzo, A.; D’Erasmo, L.; Poggiogalle, E.; Donini, L.M.; Würtz, P.; Jauhiainen, M.; Olkkonen, V.M.; et al. Metabolomic Signature of Angiopoietin-Like Protein 3 Deficiency in Fasting and Postprandial State. Arter. Thromb. Vasc. Biol. 2019, 39, 665–674. [Google Scholar] [CrossRef] [Green Version]
- Minicocci, I.; Tikka, A.; Poggiogalle, E.; Metso, J.; Montali, A.; Ceci, F.; Labbadia, G.; Fontana, M.; Di Costanzo, A.; Maranghi, M.; et al. Effects of angiopoietin-like protein 3 deficiency on postprandial lipid and lipoprotein metabolism. J. Lipid Res. 2016, 57, 1097–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camenisch, G.; Pisabarro, M.T.; Sherman, D.; Kowalski, J.; Nagel, M.; Hass, P.; Xie, M.-H.; Gurney, A.; Bodary, S.; Liang, X.H.; et al. ANGPTL3 Stimulates Endothelial Cell Adhesion and Migration via Integrin αvβ3 and Induces Blood Vessel Formation in Vivo. J. Biol. Chem. 2002, 277, 17281–17290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maik-Rachline, G.; Wortzel, I.; Seger, R. Alternative Splicing of MAPKs in the Regulation of Signaling Specificity. Cells 2021, 10, 3466. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Liu, H.T. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef]
- Watt, M.J.; Holmes, A.G.; Pinnamaneni, S.K.; Garnham, A.P.; Steinberg, G.; Kemp, B.; Febbraio, M.A. Regulation of HSL serine phosphorylation in skeletal muscle and adipose tissue. Am. J. Physiol. Metab. 2006, 290, E500–E508. [Google Scholar] [CrossRef] [PubMed]
- Vemurafenib Datasheet [Internet]. Available online: https://www.selleckchem.com/products/PLX-4032 (accessed on 13 April 2022).
- Brasaemle, D.L.; Levin, D.M.; Adler-Wailes, D.C.; Londos, C. The lipolytic stimulation of 3T3-L1 adipocytes promotes the translocation of hormone-sensitive lipase to the surfaces of lipid storage droplets. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2000, 1483, 251–262. [Google Scholar] [CrossRef]
- Pecce, V.; Verrienti, A.; Abballe, L.; Carletti, R.; Grani, G.; Falcone, R.; Ramundo, V.; Durante, C.; Di Gioia, C.; Russo, D.; et al. Loss of Function SETD2 Mutations in Poorly Differentiated Metastases from Two Hürthle Cell Carcinomas of the Thyroid. Cancers 2020, 12, 1892. [Google Scholar] [CrossRef] [PubMed]
- Berndt, S.; Liebscher, I. New Structural Perspectives in G Protein-Coupled Receptor-Mediated Src Family Kinase Activation. Int. J. Mol. Sci. 2021, 22, 6489. [Google Scholar] [CrossRef]
- Valle-Mendiola, A.; Soto-Cruz, I. Energy Metabolism in Cancer: The Roles of STAT3 and STAT5 in the Regulation of Metabolism-Related Genes. Cancers 2020, 12, 124. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, R.P.; Sheldon, R.D.; Rector, R.S. The Emerging Role of Hepatocellular eNOS in Non-alcoholic Fatty Liver Disease Development. Front. Physiol. 2020, 11, 767. [Google Scholar] [CrossRef]
- Evseeva, M.N.; Balashova, M.S.; Kulebyakin, K.Y.; Rubtsov, Y.P. Adipocyte Biology from the Perspective of In Vivo Research: Review of Key Transcription Factors. Int. J. Mol. Sci. 2021, 23, 322. [Google Scholar] [CrossRef] [PubMed]
- Jin, N.; Matter, W.F.; Michael, L.F.; Qian, Y.; Gheyi, T.; Cano, L.; Perez, C.; Lafuente, C.; Broughton, H.B.; Espada, A. The Angiopoietin-Like Protein 3 and 8 Complex Interacts with Lipoprotein Lipase and Induces LPL Cleavage. ACS Chem. Biol. 2021, 16, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Chi, X.; Britt, E.C.; Shows, H.W.; Hjelmaas, A.J.; Shetty, S.K.; Cushing, E.M.; Li, W.; Dou, A.; Zhang, R.; Davies, B.S. ANGPTL8 promotes the ability of ANGPTL3 to bind and inhibit lipoprotein lipase. Mol. Metab. 2017, 6, 1137–1149. [Google Scholar] [CrossRef] [PubMed]
- Tošić, I.; Frank, D.A. STAT3 as a mediator of oncogenic cellular metabolism: Pathogenic and therapeutic implications. Neoplasia 2021, 23, 1167–1178. [Google Scholar] [CrossRef] [PubMed]
- Inoue, H.; Ogawa, W.; Ozaki, M.; Haga, S.; Matsumoto, M.; Furukawa, K.; Hashimoto, N.; Kido, Y.; Mori, T.; Sakaue, H.; et al. Role of STAT-3 in regulation of hepatic gluconeogenic genes and carbohydrate metabolism in vivo. Nat. Med. 2004, 10, 168–174. [Google Scholar] [CrossRef]
- Gao, Z.; Daquinag, A.C.; Su, F.; Snyder, B.; Kolonin, M.G. PDGFRα / PDGFRβ signaling balance modulates progenitor cell differentiation into white and beige adipocytes. Development 2018, 145, dev155861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onogi, Y.; Wada, T.; Kamiya, C.; Inata, K.; Matsuzawa, T.; Inaba, Y.; Kimura, K.; Inoue, H.; Yamamoto, S.; Ishii, Y.; et al. PDGFRβ Regulates Adipose Tissue Expansion and Glucose Metabolism via Vascular Remodeling in Diet-Induced Obesity. Diabetes 2017, 66, 1008–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banfi, S.; Gusarova, V.; Gromada, J.; Cohen, J.C.; Hobbs, H.H. Increased thermogenesis by a noncanonical pathway in ANGPTL3/8-deficient mice. Proc. Natl. Acad. Sci. USA 2018, 115, E1249–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewey, F.E.; Gusarova, V.; Dunbar, R.; O’dushlaine, C.; Schurmann, C.; Gottesman, O.; McCarthy, S.; Van Hout, C.V.; Bruse, S.; Dansky, H.M.; et al. Genetic and Pharmacologic Inactivation of ANGPTL3 and Cardiovascular Disease. N. Engl. J. Med. 2017, 377, 211–221. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bini, S.; Pecce, V.; Di Costanzo, A.; Polito, L.; Ghadiri, A.; Minicocci, I.; Tambaro, F.; Covino, S.; Arca, M.; D’Erasmo, L. The Fibrinogen-like Domain of ANGPTL3 Facilitates Lipolysis in 3T3-L1 Cells by Activating the Intracellular Erk Pathway. Biomolecules 2022, 12, 585. https://doi.org/10.3390/biom12040585
Bini S, Pecce V, Di Costanzo A, Polito L, Ghadiri A, Minicocci I, Tambaro F, Covino S, Arca M, D’Erasmo L. The Fibrinogen-like Domain of ANGPTL3 Facilitates Lipolysis in 3T3-L1 Cells by Activating the Intracellular Erk Pathway. Biomolecules. 2022; 12(4):585. https://doi.org/10.3390/biom12040585
Chicago/Turabian StyleBini, Simone, Valeria Pecce, Alessia Di Costanzo, Luca Polito, Ameneh Ghadiri, Ilenia Minicocci, Federica Tambaro, Stella Covino, Marcello Arca, and Laura D’Erasmo. 2022. "The Fibrinogen-like Domain of ANGPTL3 Facilitates Lipolysis in 3T3-L1 Cells by Activating the Intracellular Erk Pathway" Biomolecules 12, no. 4: 585. https://doi.org/10.3390/biom12040585