
Expression of Beta-Catenin, Cadherins and P-Runx2 in Fibro-Osseous Lesions of the Jaw: Tissue Microarray Study

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Histological Classification of Samples
2.2. Microscopic and Histochemical Evaluations
- -
- Histologic pattern: (1), sclerotic-pagetoid; (2), Chinese alphabet; (3), sclerotic hyper-cellulated.
- -
- Cellularity: (1), low; (2), moderate; (3), high.
- -
- Hemorrhage: (0), negative; (1), positive.
- -
- Osteoids rimmed by fibroblast-like osteoblasts: (0), negative; (1), focal; (2), moderate; (3), high.
2.3. Tissue Microarray (TMA) Construction
2.4. TMA-Based Immunohistochemistry
2.5. Statistical Analysis
3. Results
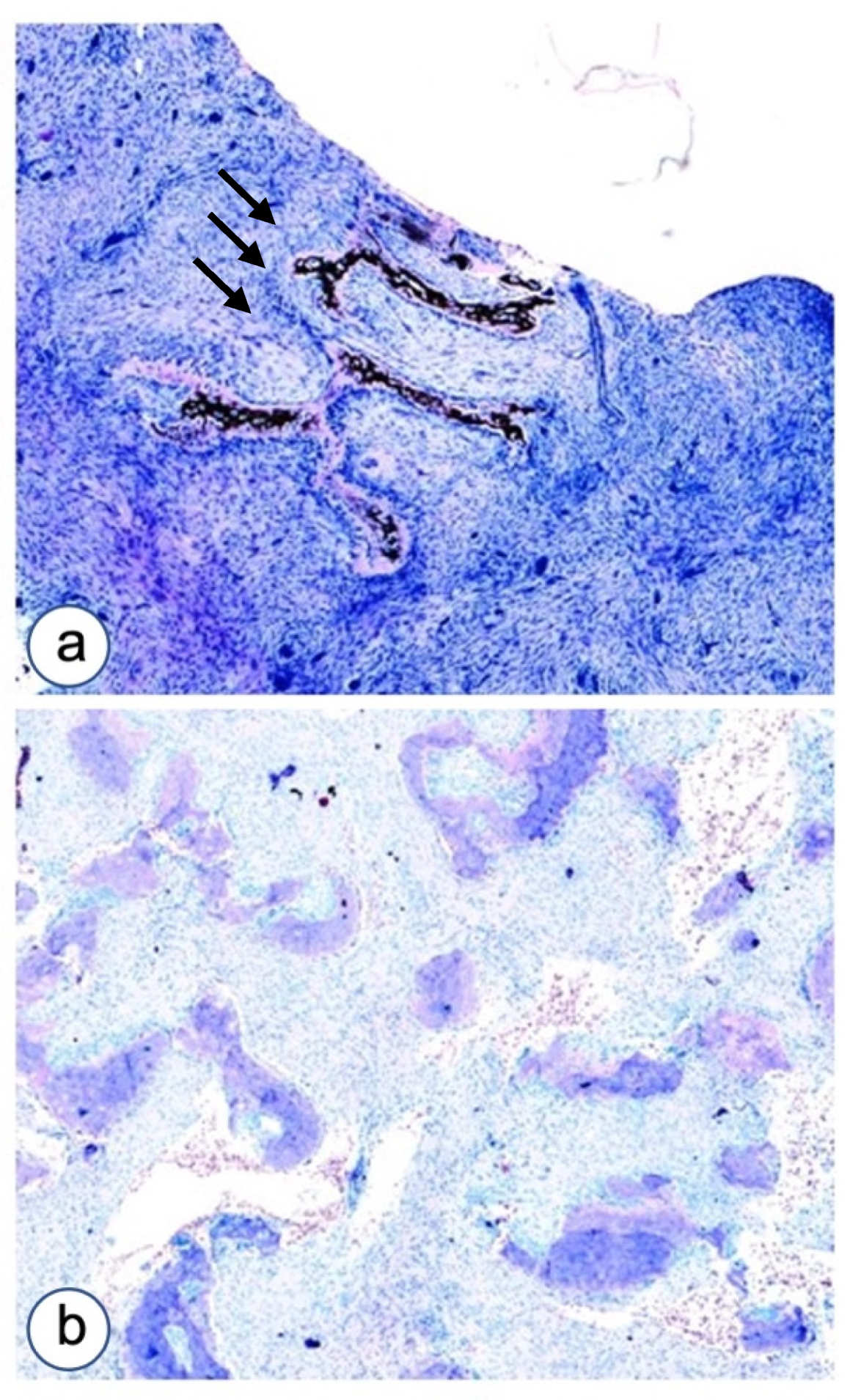
3.1. Histopathological Analysis
3.1.1. Fibrous Dysplasia
3.1.2. HPT-JT Syndrome
3.1.3. Controls
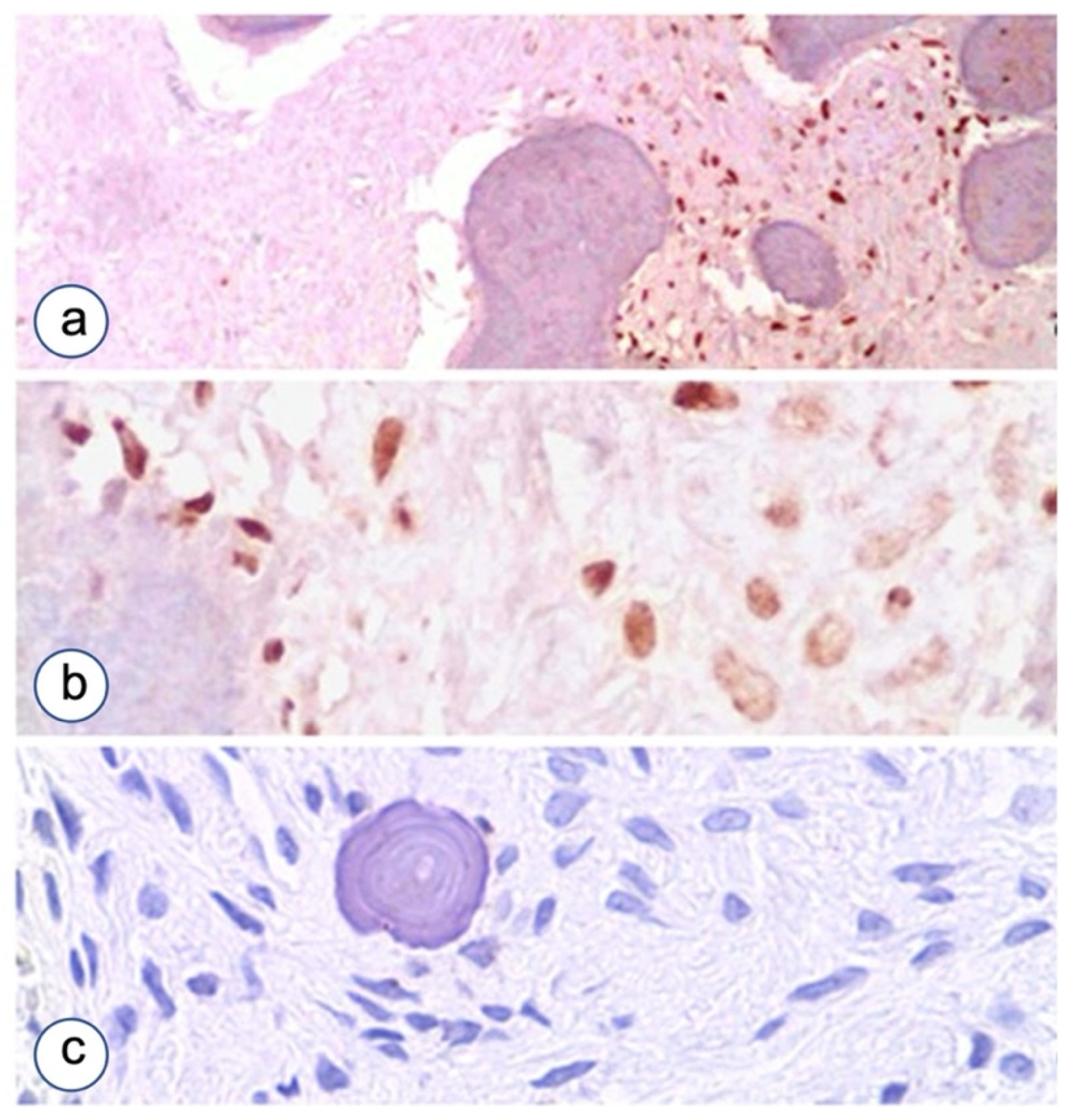
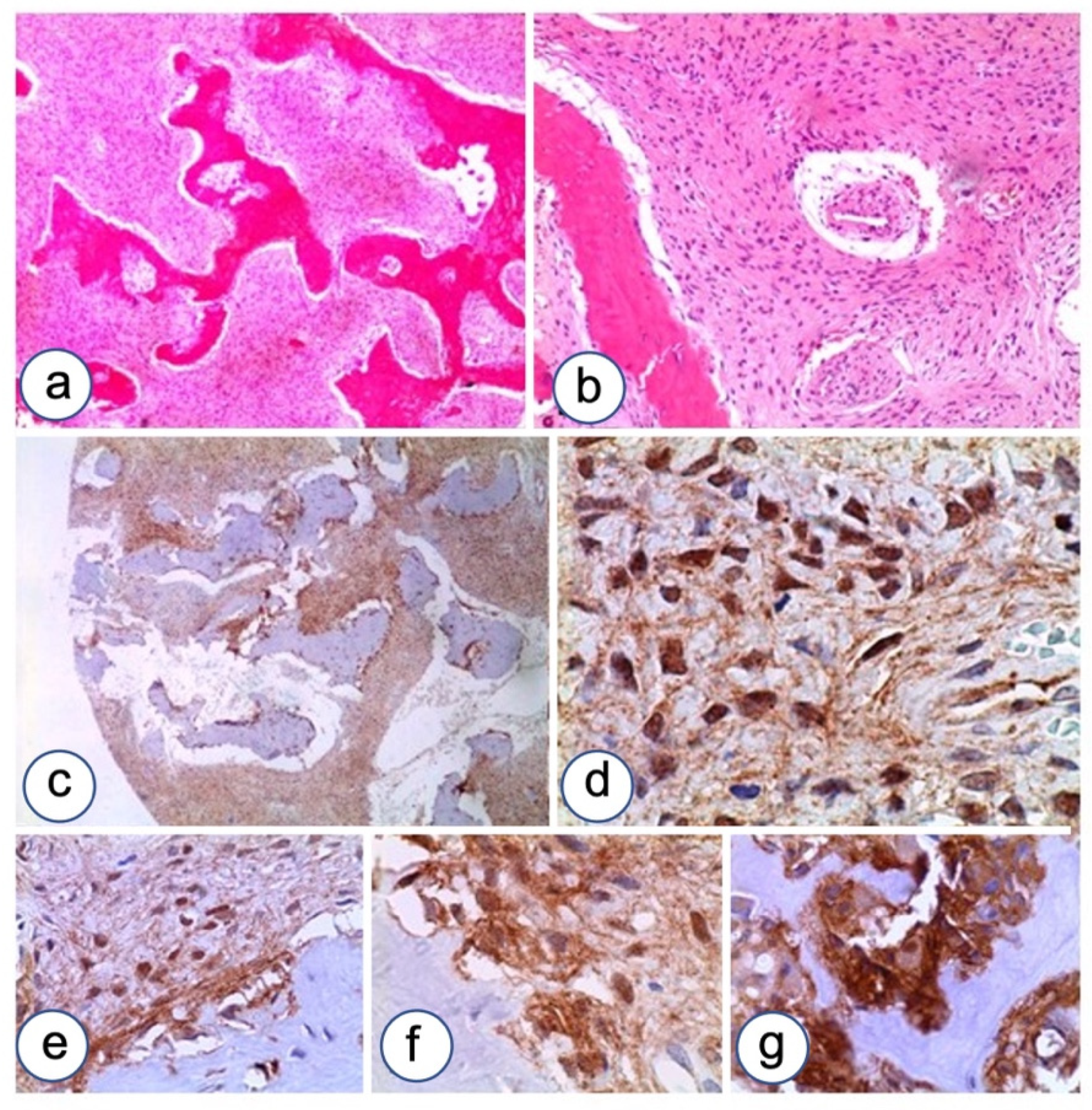
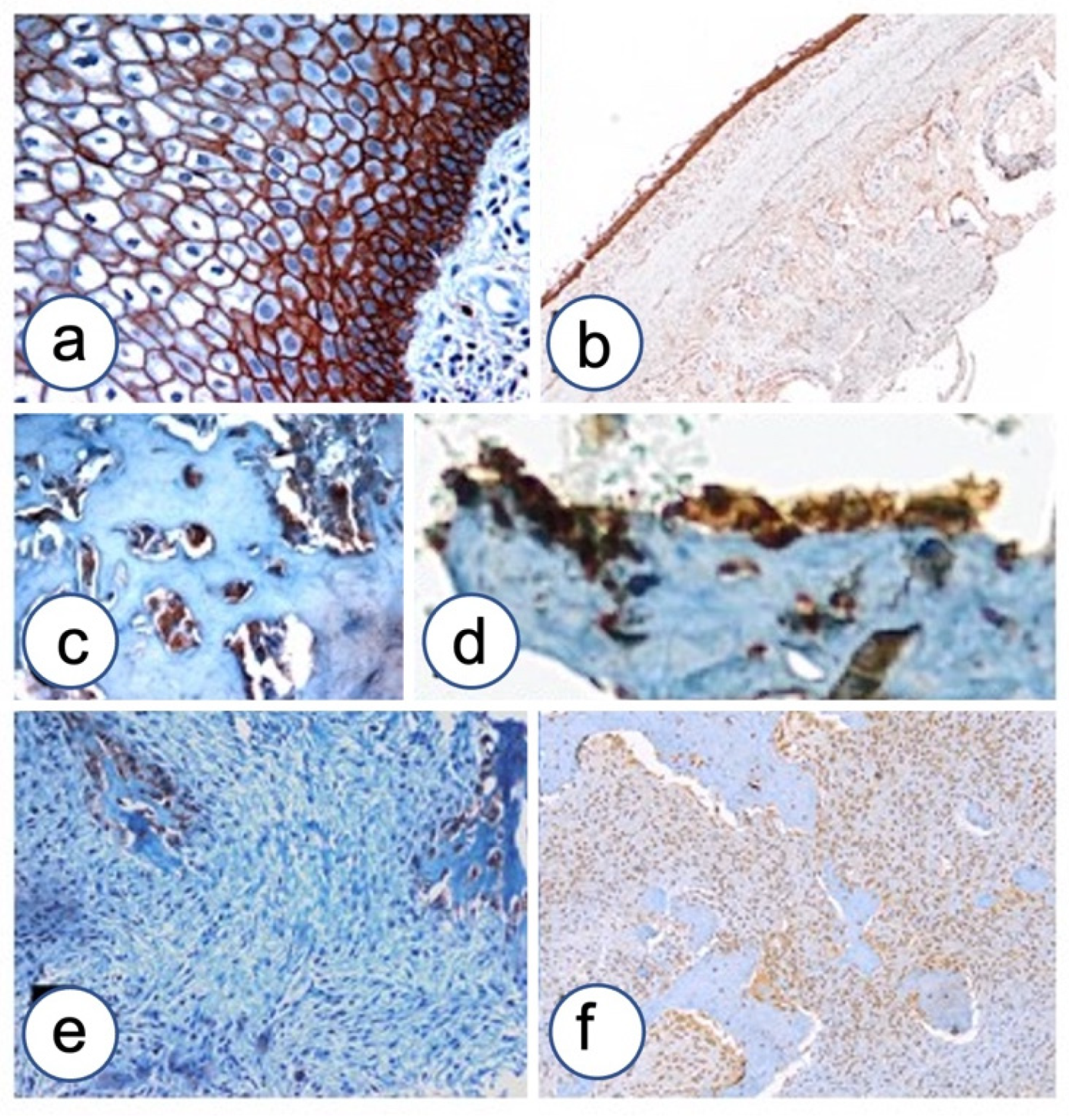
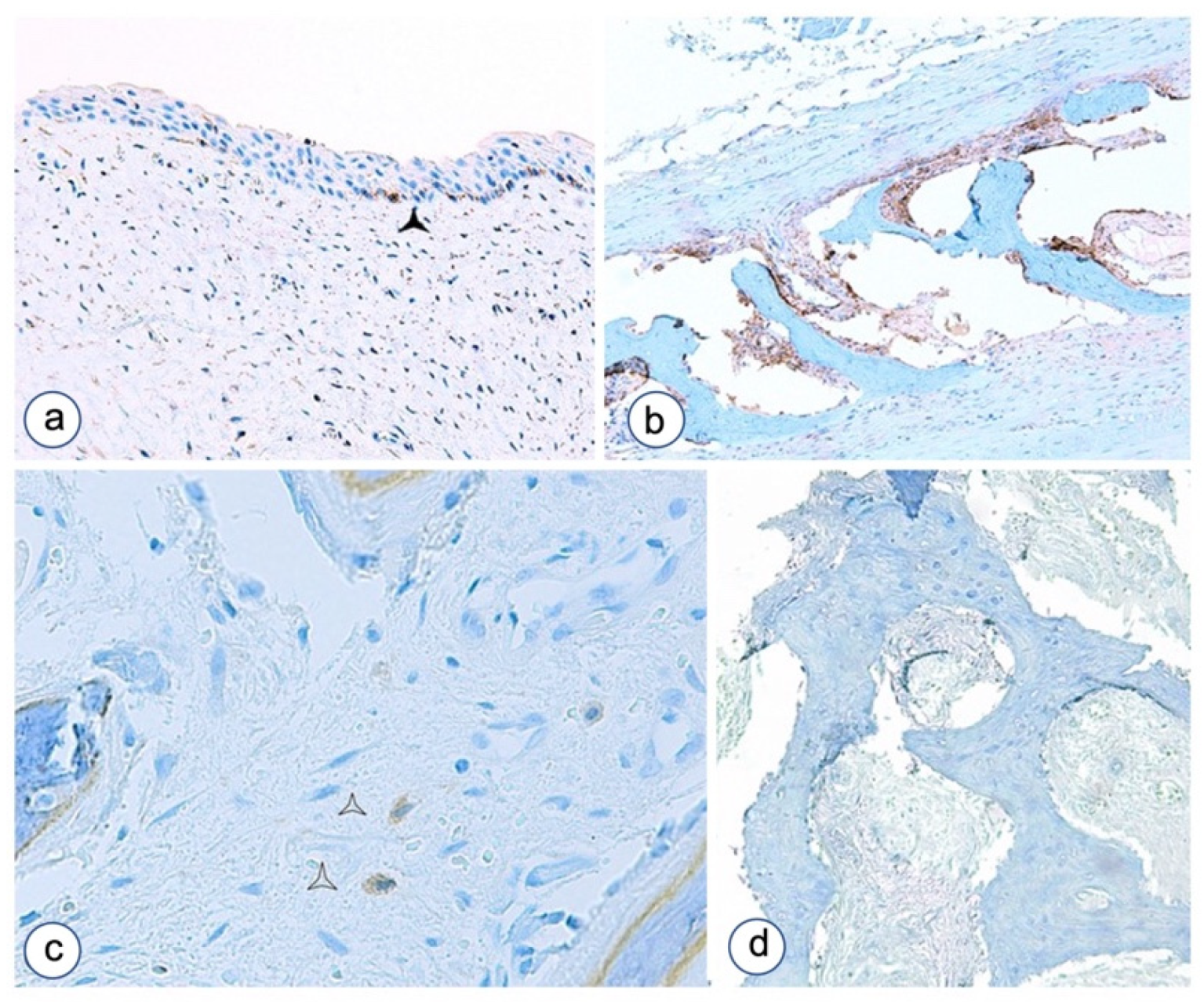
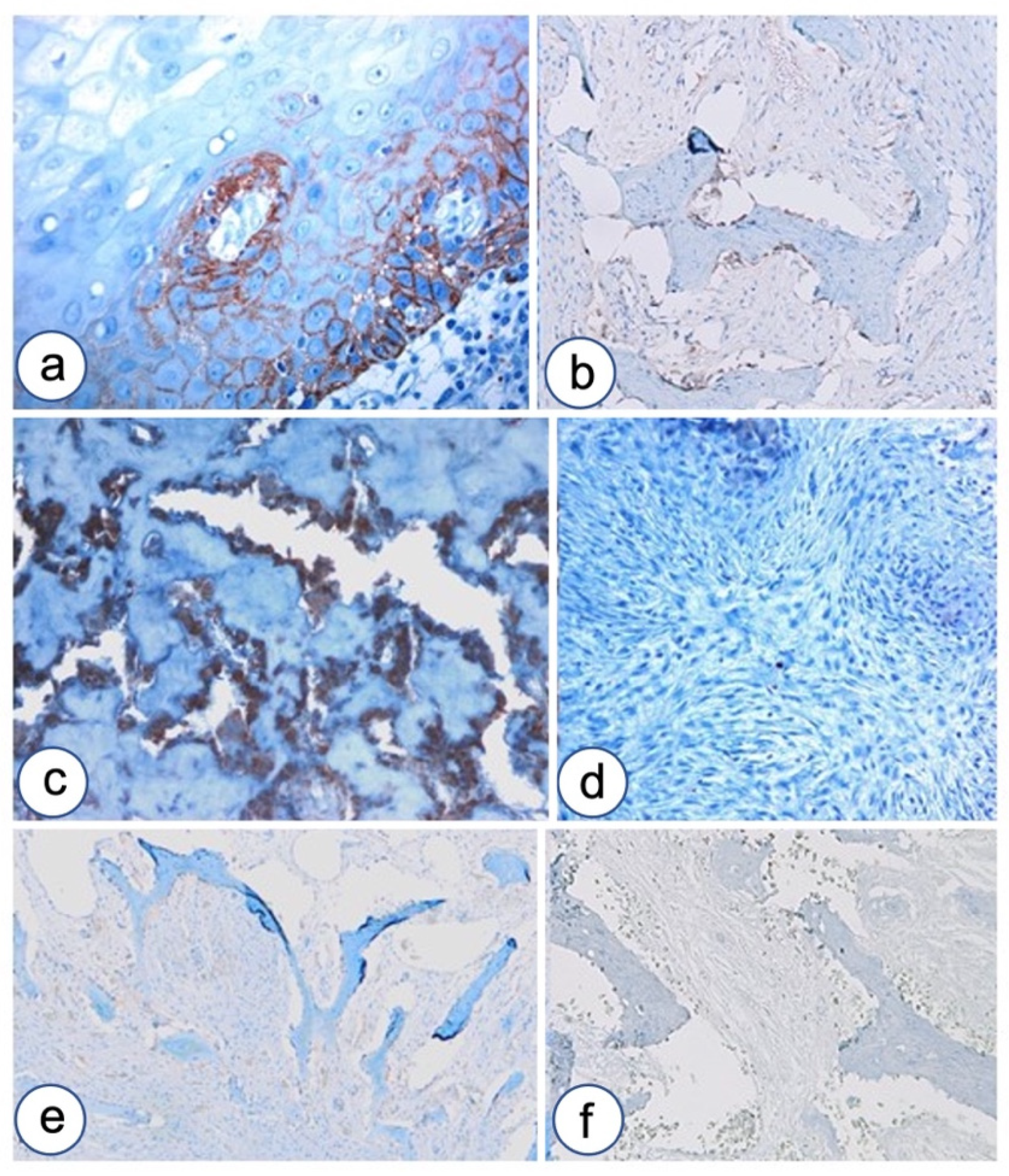
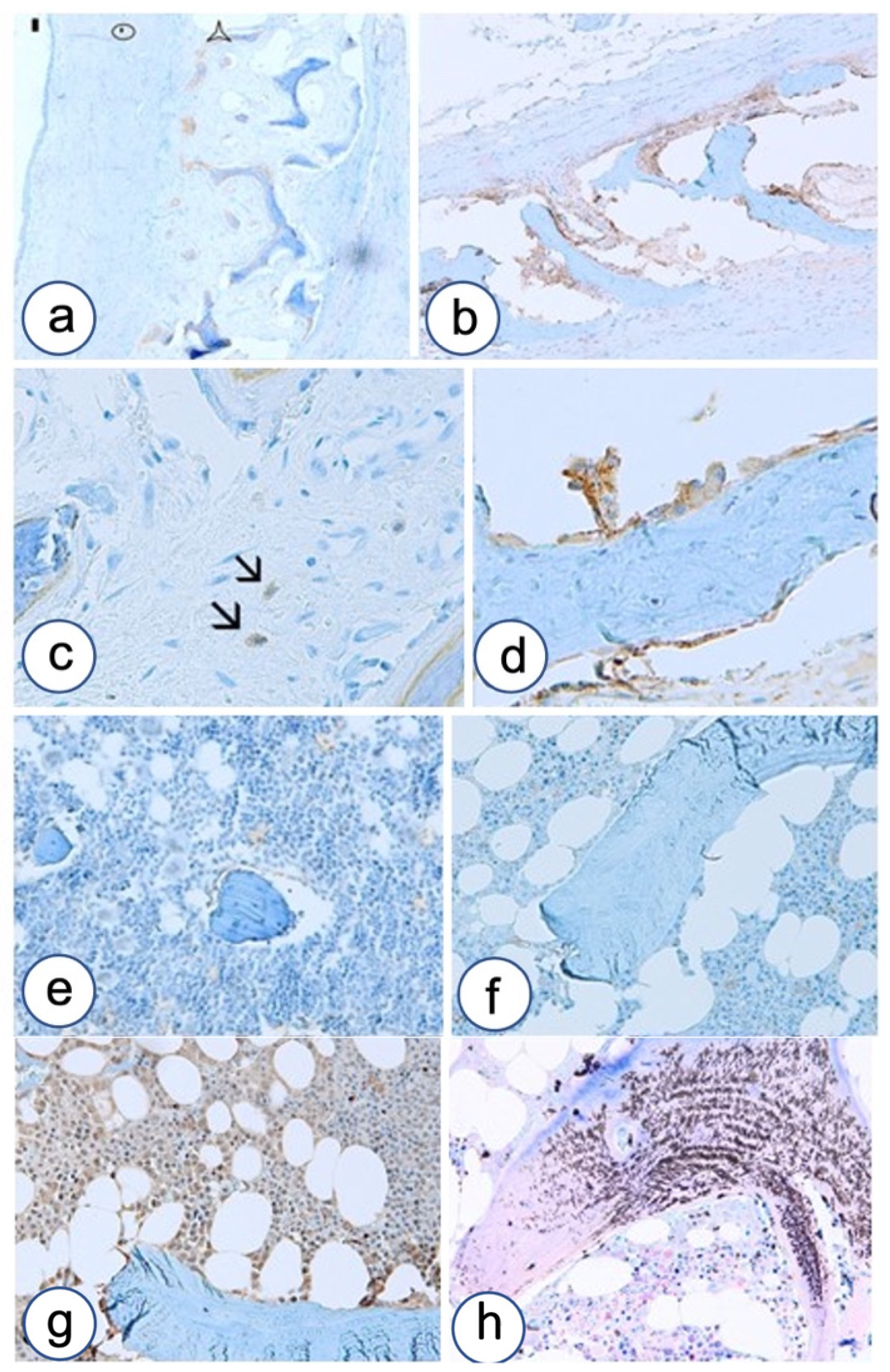
3.2. Immunohistochemistry
3.2.1. Fibrous Dysplasia
3.2.2. HPT-JT Syndrome
3.2.3. Controls
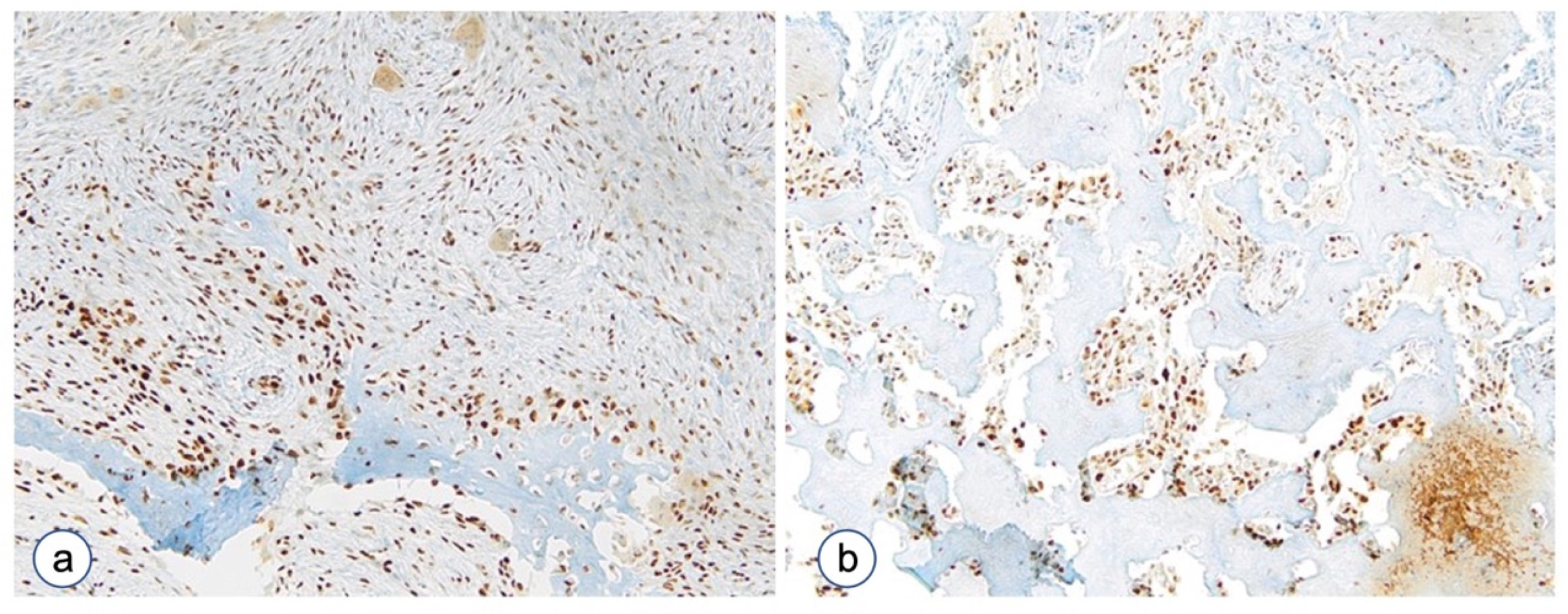
3.2.4. The Proliferative Index
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nelson, B.L.; Phillips, B.J. Benign Fibro-Osseous Lesions of the Head and Neck. Head Neck Pathol. 2019, 13, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Mainville, G.N.; Turgeon, D.P.; Kauzman, A. Diagnosis and management of benign fibro-osseous lesions of the jaws: A current review for the dental clinician. Oral Dis. 2017, 23, 440–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riminucci, M.; Robey, P.G.; Bianco, P. The pathology of fibrous dysplasia and the McCune-Albright syndrome. Pediatr. Endocrinol. Rev. 2007, 4 (Suppl. S4), 401–411. [Google Scholar] [PubMed]
- Carpten, J.D.; Robbins, C.M.; Villablanca, A.; Forsberg, L.; Presciuttini, S.; Bailey-Wilson, J.; Simonds, W.F.; Gillanders, E.M.; Kennedy, A.M.; Chen, J.D.; et al. HRPT2, encoding parafibromin, is mutated in hyperparathyroidism-jaw tumor syndrome. Nat. Genet. 2002, 32, 676–680. [Google Scholar] [CrossRef] [PubMed]
- DiCaprio, M.R.; Enneking, W.F. Fibrous dysplasia. Pathophysiology, evaluation, and treatment. J. Bone Joint Surg. Am. 2005, 87, 1848–1864. [Google Scholar] [CrossRef]
- Maki, M.; Saitoh, K.; Horiuchi, H.; Morohoshi, T.; Fukayama, M.; Machinami, R. Comparative study of fibrous dysplasia and osteofibrous dysplasia: Histopathological, immunohistochemical, argyrophilic nucleolar organizer region and DNA ploidy analysis. Pathol. Int. 2001, 51, 603–611. [Google Scholar] [CrossRef]
- Dumitrescu, C.E.; Collins, M.T. McCune-Albright syndrome. Orphanet J. Rare Dis. 2008, 3, 12. [Google Scholar] [CrossRef] [Green Version]
- El-Mofty, S.K. Odontogenic and maxillofacial bone tumours. In WHO Classification of Head and Neck Tumours, 4th ed.; El-Naggar, A.K., Chan, J.K.C., Grandis, J.R., Takata, T., Slootweg, P.J., Eds.; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Karadag, A.; Riminucci, M.; Bianco, P.; Cherman, N.; Kuznetsov, S.A.; Nguyen, N.; Collins, M.T.; Robey, P.G.; Fisher, L.W. A novel technique based on a PNA hybridization probe and FRET principle for quantification of mutant genotype in fibrous dysplasia/McCune-Albright syndrome. Nucleic Acids Res. 2004, 32, e63. [Google Scholar] [CrossRef] [Green Version]
- Slootweg, P.J.; El-Mofty, S.K. Ossifyng Fibroma. In Pathology & Genetics Head and Neck Tumors; Barnes, L., Eveson, J.W., Reichart, P., Sidransky, D., Eds.; IARC Press: Lyon, France, 2005; pp. 319–320. [Google Scholar]
- Burke, A.B.; Collins, M.T.; Boyce, A.M. Fibrous dysplasia of bone: Craniofacial and dental implications. Oral Dis. 2017, 23, 697–708. [Google Scholar] [CrossRef]
- Pereira, T.; Gomes, C.C.; Brennan, P.A.; Fonseca, F.P.; Gomez, R.S. Fibrous dysplasia of the jaws: Integrating molecular pathogenesis with clinical, radiological, and histopathological features. J. Oral Pathol. Med. 2019, 48, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Teh, B.T.; Farnebo, F.; Kristoffersson, U.; Sundelin, B.; Cardinal, J.; Axelson, R.; Yap, A.; Epstein, M.; Heath, H., 3rd; Cameron, D.; et al. Autosomal dominant primary hyperparathyroidism and jaw tumor syndrome associated with renal hamartomas and cystic kidney disease: Linkage to 1q21-q32 and loss of the wild type allele in renal hamartomas. J. Clin. Endocrinol. Metab. 1996, 81, 4204–4211. [Google Scholar] [CrossRef] [PubMed]
- Gill, A.J.; Clarkson, A.; Gimm, O.; Keil, J.; Dralle, H.; Howell, V.M.; Marsh, D.J. Loss of nuclear expression of parafibromin distinguishes parathyroid carcinomas and hyperparathyroidism-jaw tumor (HPT-JT) syndrome-related adenomas from sporadic parathyroid adenomas and hyperplasias. Am. J. Surg. Pathol. 2006, 30, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Kennett, S.; Pollick, H. Jaw lesions in familial hyperparathyroidism. Oral Surg. Oral Med. Oral Pathol. 1971, 31, 502–510. [Google Scholar] [CrossRef]
- Guerrouani, A.; Rzin, A.; El Khatib, K. Hyperparathyroidism-jaw tumour syndrome detected by aggressive generalized osteitis fibrosa cystica. Clin. Cases Miner. Bone Metab. 2013, 10, 65–67. [Google Scholar] [CrossRef]
- Horvai, A.E.; Jordan, R.C. Fibro-osseous lesions of the craniofacial bones: Beta-catenin immunohistochemical analysis and CTNNB1 and APC mutation analysis. Head Neck Pathol. 2014, 8, 291–297. [Google Scholar] [CrossRef] [Green Version]
- Kramer, I.; Halleux, C.; Keller, H.; Pegurri, M.; Gooi, J.H.; Weber, P.B.; Feng, J.Q.; Bonewald, L.F.; Kneissel, M. Osteocyte Wnt/beta-catenin signaling is required for normal bone homeostasis. Mol. Cell Biol. 2010, 30, 3071–3085. [Google Scholar] [CrossRef] [Green Version]
- Bennett, C.N.; Longo, K.A.; Wright, W.S.; Suva, L.J.; Lane, T.F.; Hankenson, K.D.; MacDougald, O.A. Regulation of osteoblastogenesis and bone mass by Wnt10b. Proc. Natl. Acad. Sci. USA 2005, 102, 3324–3329. [Google Scholar] [CrossRef] [Green Version]
- Colas-Algora, N.; Millan, J. How many cadherins do human endothelial cells express? Cell Mol. Life Sci. 2019, 76, 1299–1317. [Google Scholar] [CrossRef]
- Kaszak, I.; Witkowska-Pilaszewicz, O.; Niewiadomska, Z.; Dworecka-Kaszak, B.; Ngosa Toka, F.; Jurka, P. Role of Cadherins in Cancer-A Review. Int. J. Mol. Sci. 2020, 21, 7624. [Google Scholar] [CrossRef]
- Cao, Z.Q.; Wang, Z.; Leng, P. Aberrant N-cadherin expression in cancer. Biomed. Pharmacother. 2019, 118, 109320. [Google Scholar] [CrossRef]
- Eversole, L.R.; Sabes, W.R.; Rovin, S. Fibrous dysplasia: A nosologic problem in the diagnosis of fibro-osseous lesions of the jaws. J. Oral Pathol. 1972, 1, 189–220. [Google Scholar] [CrossRef] [PubMed]
- Riminucci, M.; Fisher, L.W.; Shenker, A.; Spiegel, A.M.; Bianco, P.; Gehron Robey, P. Fibrous dysplasia of bone in the McCune-Albright syndrome: Abnormalities in bone formation. Am. J. Pathol. 1997, 151, 1587–1600. [Google Scholar] [PubMed]
- Riminucci, M.; Liu, B.; Corsi, A.; Shenker, A.; Spiegel, A.M.; Robey, P.G.; Bianco, P. The histopathology of fibrous dysplasia of bone in patients with activating mutations of the Gs alpha gene: Site-specific patterns and recurrent histological hallmarks. J. Pathol. 1999, 187, 249–258. [Google Scholar] [CrossRef]
- Aquino, G.; Pannone, G.; Santoro, A.; Liguori, G.; Franco, R.; Serpico, R.; Florio, G.; De Rosa, A.; Mattoni, M.; Cozza, V.; et al. pEGFR-Tyr 845 expression as prognostic factors in oral squamous cell carcinoma: A tissue-microarray study with clinic-pathological correlations. Cancer Biol. Ther. 2012, 13, 967–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, C.; Yang, Q.; Zhao, G.; Yu, H.; Kirkwood, K.L.; Franceschi, R.T. Interactions between extracellular signal-regulated kinase 1/2 and p38 MAP kinase pathways in the control of RUNX2 phosphorylation and transcriptional activity. J. Bone Miner. Res. 2012, 27, 538–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Domenico, M.; Pierantoni, G.M.; Feola, A.; Esposito, F.; Laino, L.; De Rosa, A.; Rullo, R.; Mazzotta, M.; Martano, M.; Sanguedolce, F.; et al. Prognostic significance of N-Cadherin expression in oral squamous cell carcinoma. Anticancer Res. 2011, 31, 4211–4218. [Google Scholar] [PubMed]
- Santarelli, A.; Di Fede, O.; Carinci, F.; Rubini, C.; Ganzetti, G.; Perrone, D.; Simonetti, O.; Offidani, A.; Re, M.; Perrone, D.; et al. Reciprocal altered expression of E-cadherin and P-cadherin in mucous membrane pemphigoid. Int. J. Immunopathol. Pharmacol. 2011, 24, 103–106. [Google Scholar] [CrossRef]
- Papagerakis, P.; Pannone, G.; Shabana, A.H.; Depondt, J.; Santoro, A.; Ghirtis, K.; Berdal, A.; Papagerakis, S. Aberrant beta-catenin and LEF1 expression may predict the clinical outcome for patients with oropharyngeal cancer. Int. J. Immunopathol. Pharmacol. 2012, 25, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Santoro, A.; Pannone, G.; Errico, M.E.; Bifano, D.; Lastilla, G.; Bufo, P.; Loreto, C.; Donofrio, V. Role of beta-catenin expression in paediatric mesenchymal lesions: A tissue microarray-based immunohistochemical study. Eur. J. Histochem. 2012, 56, e25. [Google Scholar] [CrossRef] [Green Version]
- Santoro, A.; Pannone, G.; Papagerakis, S.; McGuff, H.S.; Cafarelli, B.; Lepore, S.; De Maria, S.; Rubini, C.; Mattoni, M.; Staibano, S.; et al. Beta-catenin and epithelial tumors: A study based on 374 oropharyngeal cancers. Biomed. Res. Int. 2014, 2014, 948264. [Google Scholar] [CrossRef]
- Pannone, G.; Santoro, A.; Feola, A.; Bufo, P.; Papagerakis, P.; Lo Muzio, L.; Staibano, S.; Ionna, F.; Longo, F.; Franco, R.; et al. The role of E-cadherin down-regulation in oral cancer: CDH1 gene expression and epigenetic blockage. Curr. Cancer Drug Targets 2014, 14, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Hamner, J.E., 3rd; Scofield, H.H.; Cornyn, J. Benign fibro-osseous jaw lesions of periodontal membrane origin. An analysis of 249 cases. Cancer 1968, 22, 861–878. [Google Scholar] [CrossRef]
- Nogueira, R.L.; Faria, M.H.; Osterne, R.L.; Cavalcante, R.B.; Ribeiro, R.A.; Nonaka, C.F.; Rabenhorst, S.H. Central giant cell lesion of the jaws: Study of CCND1 gene amplification and p16INK4a protein levels. J. Mol. Histol. 2013, 44, 527–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moskovszky, L.; Idowu, B.; Taylor, R.; Mertens, F.; Athanasou, N.; Flanagan, A. Analysis of giant cell tumour of bone cells for Noonan syndrome/cherubism-related mutations. J. Oral Pathol. Med. 2013, 42, 95–98. [Google Scholar] [CrossRef]
- Flucke, U.; Tops, B.B.; van Diest, P.J.; Slootweg, P.J. Desmoid-type fibromatosis of the head and neck region in the paediatric population: A clinicopathological and genetic study of seven cases. Histopathology 2014, 64, 769–776. [Google Scholar] [CrossRef]
- Tejpar, S.; Nollet, F.; Li, C.; Wunder, J.S.; Michils, G.; dal Cin, P.; Van Cutsem, E.; Bapat, B.; van Roy, F.; Cassiman, J.J.; et al. Predominance of beta-catenin mutations and beta-catenin dysregulation in sporadic aggressive fibromatosis (desmoid tumor). Oncogene 1999, 18, 6615–6620. [Google Scholar] [CrossRef] [Green Version]
- He, X. Unwinding a path to nuclear beta-catenin. Cell 2006, 127, 40–42. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Swope, D.; Raess, N.; Cheng, L.; Muller, E.J.; Radice, G.L. Cardiac tissue-restricted deletion of plakoglobin results in progressive cardiomyopathy and activation of {beta}-catenin signaling. Mol. Cell Biol. 2011, 31, 1134–1144. [Google Scholar] [CrossRef] [Green Version]
- Pannone, G.; Bufo, P.; Santoro, A.; Franco, R.; Aquino, G.; Longo, F.; Botti, G.; Serpico, R.; Cafarelli, B.; Abbruzzese, A.; et al. WNT pathway in oral cancer: Epigenetic inactivation of WNT-inhibitors. Oncol. Rep. 2010, 24, 1035–1041. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Xu, Y.; Fu, Q.; He, M. Parathyroid hormone regulates osteoblast differentiation in a Wnt/beta-catenin-dependent manner. Mol. Cell Biochem. 2011, 355, 211–216. [Google Scholar] [CrossRef]
- Regard, J.B.; Cherman, N.; Palmer, D.; Kuznetsov, S.A.; Celi, F.S.; Guettier, J.M.; Chen, M.; Bhattacharyya, N.; Wess, J.; Coughlin, S.R.; et al. Wnt/beta-catenin signaling is differentially regulated by Galpha proteins and contributes to fibrous dysplasia. Proc. Natl. Acad. Sci. USA 2011, 108, 20101–20106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, T.F.; Guo, X.; Garrett-Beal, L.; Yang, Y. Wnt/beta-catenin signaling in mesenchymal progenitors controls osteoblast and chondrocyte differentiation during vertebrate skeletogenesis. Dev. Cell 2005, 8, 739–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepore, S.; Milillo, L.; Trotta, T.; Castellani, S.; Porro, C.; Panaro, M.A.; Santarelli, A.; Bambini, F.; Lo Muzio, L.; Conese, M.; et al. Adhesion and growth of osteoblast-like cells on laser-engineered porous titanium surface: Expression and localization of N-cadherin and beta-catenin. J. Biol. Regul. Homeost Agents 2013, 27, 531–541. [Google Scholar] [PubMed]
- Vleminckx, K.; Kemler, R. Cadherins and tissue formation: Integrating adhesion and signaling. Bioessays 1999, 21, 211–220. [Google Scholar] [CrossRef]
- Kawaguchi, J.; Kii, I.; Sugiyama, Y.; Takeshita, S.; Kudo, A. The transition of cadherin expression in osteoblast differentiation from mesenchymal cells: Consistent expression of cadherin-11 in osteoblast lineage. J. Bone Miner. Res. 2001, 16, 260–269. [Google Scholar] [CrossRef]
- Choong, P.F. The molecular basis of skeletal metastases. Clin. Orthop. Relat. Res. 2003, S19–S31. [Google Scholar] [CrossRef]
- Marie, P.J. Signaling pathways affecting skeletal health. Curr. Osteoporos Rep. 2012, 10, 190–198. [Google Scholar] [CrossRef]
- Hay, E.; Lemonnier, J.; Modrowski, D.; Lomri, A.; Lasmoles, F.; Marie, P.J. N- and E-cadherin mediate early human calvaria osteoblast differentiation promoted by bone morphogenetic protein-2. J. Cell. Physiol. 2000, 183, 117–128. [Google Scholar] [CrossRef]
- Hay, E.; Laplantine, E.; Geoffroy, V.; Frain, M.; Kohler, T.; Muller, R.; Marie, P.J. N-cadherin interacts with axin and LRP5 to negatively regulate Wnt/beta-catenin signaling, osteoblast function, and bone formation. Mol. Cell Biol. 2009, 29, 953–964. [Google Scholar] [CrossRef] [Green Version]
- Simonneau, L.; Kitagawa, M.; Suzuki, S.; Thiery, J.P. Cadherin 11 expression marks the mesenchymal phenotype: Towards new functions for cadherins? Cell Adhes. Commun. 1995, 3, 115–130. [Google Scholar] [CrossRef]
- Kimura, Y.; Matsunami, H.; Takeichi, M. Expression of cadherin-11 delineates boundaries, neuromeres, and nuclei in the developing mouse brain. Dev. Dyn. 1996, 206, 455–462. [Google Scholar] [CrossRef]
- Patel, S.D.; Ciatto, C.; Chen, C.P.; Bahna, F.; Rajebhosale, M.; Arkus, N.; Schieren, I.; Jessell, T.M.; Honig, B.; Price, S.R.; et al. Type II cadherin ectodomain structures: Implications for classical cadherin specificity. Cell 2006, 124, 1255–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourne, S.; Polak, J.M.; Hughes, S.P.; Buttery, L.D. Osteogenic differentiation of mouse embryonic stem cells: Differential gene expression analysis by cDNA microarray and purification of osteoblasts by cadherin-11 magnetically activated cell sorting. Tissue Eng. 2004, 10, 796–806. [Google Scholar] [CrossRef] [PubMed]
- Komori, T.; Yagi, H.; Nomura, S.; Yamaguchi, A.; Sasaki, K.; Deguchi, K.; Shimizu, Y.; Bronson, R.T.; Gao, Y.H.; Inada, M.; et al. Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell 1997, 89, 755–764. [Google Scholar] [CrossRef] [Green Version]
- Cooper, S.C.; Flaitz, C.M.; Johnston, D.A.; Lee, B.; Hecht, J.T. A natural history of cleidocranial dysplasia. Am. J. Med. Genet. 2001, 104, 1–6. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Data | ||||||||
|---|---|---|---|---|---|---|---|---|
| Case N. | Age | Sex | Diagnosis | Anatomical Site | Bone/Osteoid Pattern: Pagetoid (P) Chinese Alphabet (CA) Hyper-Cellulated (HC) | ∇∇ Cellularity 1. Low 2. Moderate 3. High | Hemorrhage 0. Negative 1. Positive | Rimming Osteoblasts 0. Negative 1. Focal 2. Moderate 3. High |
| 1 | 49 | M | FD | Maxilla | P/HC | 3 | 0 | 2 |
| 2 | 44 | M | FD | Maxilla | CA | 2 | 0 | 1 |
| 3 | 37 | M | FD | Mandible | P | 1 | 1 | 1 |
| 4 | 31 | M | FD | Mandible | P | 1 | 1 | 1 |
| 5 | 25 | F | FD | Mandible | CA | 3 | 1 | 1 |
| 6 | 30 | F | FD | Mandible | CA | 3 | 0 | 1 |
| 7 | 35 | F | FD | Maxilla | P | 1 | 0 | 1 |
| 8 | FD | CA | 3 | 1 | ||||
| 9 | 37 | F | HPT-JT | Maxilla | P | 1 | 0 | 1 |
| Controls | Bone pattern | |||||||
| 10 | 26 | M | Reactive fibromyxoid | Mandible | Parallel bone | 2 | 0 | 1 |
| 11 | 60 | M | Mature reactive fibrous tissue | Maxilla | Bone absent | 1 | 0 | 0 |
| 12 | 23 | M | Reactive fibrous tissue | Mandible | Parallel bone | 2 | 0 | 1 |
| 13 | 27 | Reactive fibromyxoid | Maxilla | Bone absent | 2 | 0 | ||
| 14 | 60 | F | Mature reactive fibrous tissue | Maxilla | Bone absent | 1 | 0 | 0 |
| 15 | 40 | F | Normal bone with hematopoietic cells | Iliac crest | Trabecular bone | 2 | 0 | 0 |
| 16 | 21 | M | Remodeling bone surrounding developmental odontogenic cyst | Maxilla | Parallel trabeculae; focal fish-hook appearance | 2 | 0 | 2 |
| Immunohistochemical Findings | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cases | Ki-67 | CK AE1/AE3 | Beta-Catenin | E-Cadherin | N-Cadherin | OB-Cadherin | P-Cadherin | P-Runx2 | ||
| Cell Count/HPF ° | Score | Sub-Cellular Localization | Score | Nuclear Score | Score | Score | Score | Score | Score | |
| 1 | 6 | 0 | C | 3+Os | 0 | 0 | 3+Os Fbs | 0 | 3+ | 3+Os, Fbs |
| 2 | 0 | 0 | CM | 3+Os | 0 | 0 | 3+Os Octs | 1+ | 0 | 3+Os, Fbs |
| 3 | 4 | 0 | 0 | 0 | 0 | 3+ | 0 | 0 | 3+Os, Fbs | |
| 4 | 1 | 0 | C | 2+Os | 0 | 0 | 2+Os; 0 Fbs | 0 | 0 | 0 |
| 5 | 3 | 0 | C | 3+Os Fbs | 0 | 0 | 2+Os Fbs | 0 | 1+ | 3+Os, Fbs |
| 6 | 0 | - | 0 | 0 | 0 | 1+ | 0 | 0 | 2+Os, Fbs | |
| 7 | 0 | - | 0 | 0 | 0 | 3+ | 0 | 0 | 0 | |
| 8 | 5 | 0 | CN | 3+Os Fbs | 3+ | 0 | 3+ | 0 | 0 | 3+Os, Fbs |
| 9 | 0 | 1+ | C | 1+ | 0 | 0 | 1+ | 0 | 0 | 2+Os, Fbs |
| Controls | - | |||||||||
| 10 | 4 | 0 | M | 0 | 0 | 0 | 0 | 0 | 1+ | |
| 11 | 2 | 0 | - | 0 | 0 | 1+ | 0 | 0 | 0 | 1+ |
| 12 | 4 | 0 | M | 0 | 0 | 0 | 0 | 0 | 1+ | |
| 13 | 5 | 0 | M | 1+ | 0 | 1+ | NV | 0 | 0 | 1+ |
| 14 | 6 | 0 | - | 0 | 0 | 0 | 2+Fbs | 0 | 0 | 2+ |
| 15 | 0 | 0 | M | 1+ | 0 | 1+ | 1+ | 2+ | 1+ | 1+ |
| 16 | 4 | 0 | M | 1+ | 0 | 0 | 2+ | 3+ | 1+Os | 2+ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pannone, G.; Nocini, R.; Santoro, A.; Spirito, F.; Nocini, P.F.; Papagerakis, S.; Franceschi, R.T.; Di Domenico, M.; Di Carlo, A.; Danelia, N.; et al. Expression of Beta-Catenin, Cadherins and P-Runx2 in Fibro-Osseous Lesions of the Jaw: Tissue Microarray Study. Biomolecules 2022, 12, 587. https://doi.org/10.3390/biom12040587
Pannone G, Nocini R, Santoro A, Spirito F, Nocini PF, Papagerakis S, Franceschi RT, Di Domenico M, Di Carlo A, Danelia N, et al. Expression of Beta-Catenin, Cadherins and P-Runx2 in Fibro-Osseous Lesions of the Jaw: Tissue Microarray Study. Biomolecules. 2022; 12(4):587. https://doi.org/10.3390/biom12040587
Chicago/Turabian StylePannone, Giuseppe, Riccardo Nocini, Angela Santoro, Francesca Spirito, Pier Francesco Nocini, Silvana Papagerakis, Renny T. Franceschi, Marina Di Domenico, Angelina Di Carlo, Nana Danelia, and et al. 2022. "Expression of Beta-Catenin, Cadherins and P-Runx2 in Fibro-Osseous Lesions of the Jaw: Tissue Microarray Study" Biomolecules 12, no. 4: 587. https://doi.org/10.3390/biom12040587