
O-GlcNAc-Mediated Regulation of Galectin Expression and Secretion in Human Promyelocytic HL-60 Cells Undergoing Neutrophilic Differentiation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Solutions
2.2. Cell Culture and Treatments
2.3. RNA Isolation, cDNA Synthesis, and PCR Gene Expression Assays
2.4. Western and Immunodot Blotting
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Scopoletin Assay to Measure PMA-Induced H2O2 Production
2.7. Nuclear Staining, Fluorescence Microscopy, and Nuclear Morphology
2.8. Statistical Analysis
3. Results
3.1. Effects of ATRA and Inhibitors of O-GlcNAc Cycle Enzymes on Neutrophilic Differentiation of HL-60 Cells in Serum-Contained and Serum-Free Cell Culture Media
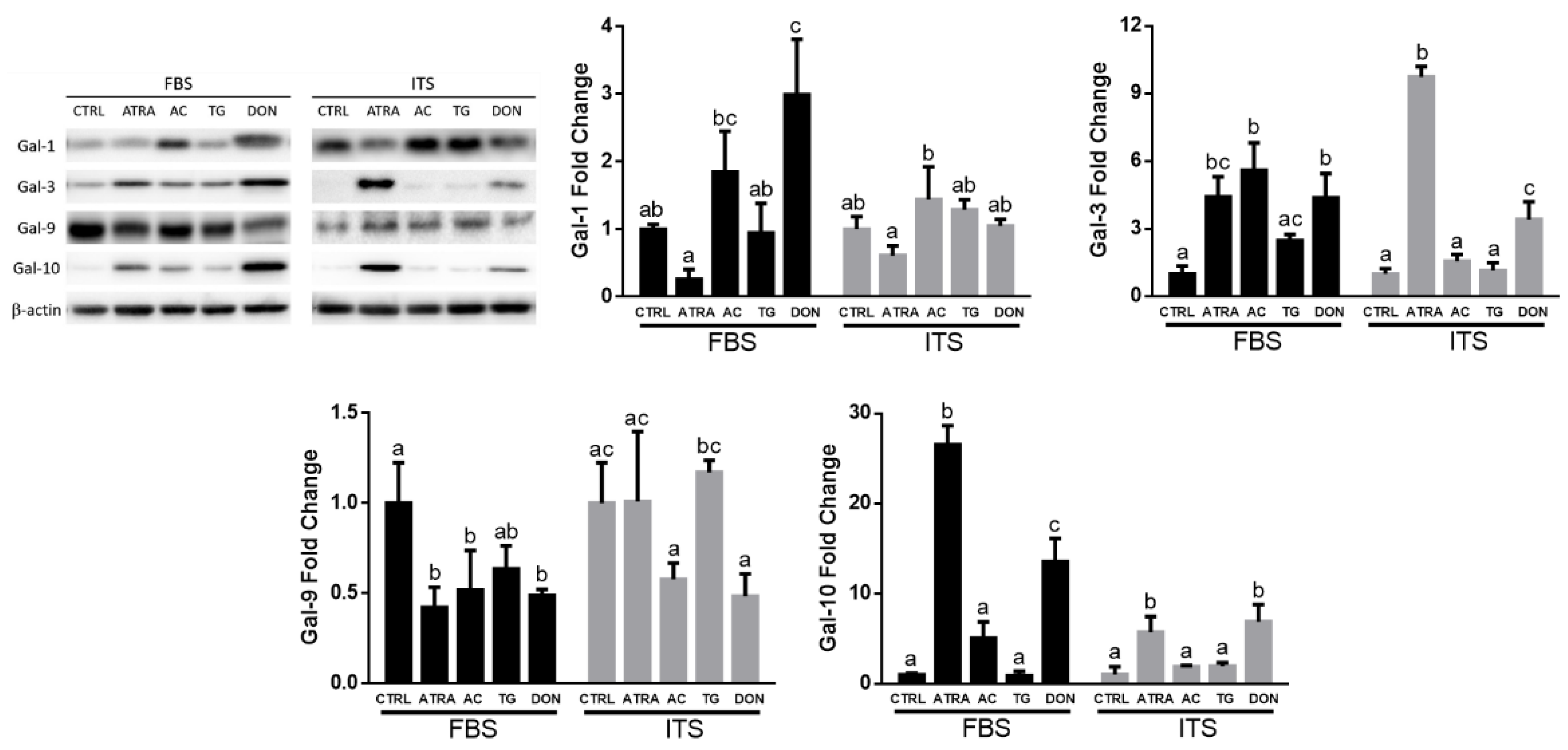
3.2. Galectin Expression Profiles Depend on O-GlcNAc Homeostatic Changes in HL-60 Cells
3.3. Effects of ATRA and O-GlcNAc Cycle Enzyme Inhibitors on the Secretion of Galectins from HL-60 Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence 5′-3′ | Size bp | 2 Step Cycling | Melting Peak Temperature | PMID Reference | |
|---|---|---|---|---|---|---|
| LGALS1 | F | CCTGGAGAGTGCCTTCGAGTG | 220 | 95 °C (5 s) 60 °C (25 s) | 86.5 °C | 23108139 |
| R | CTGCAACACTTCCAGGCTGG | |||||
| LGALS3 | F | CAGAATTGCTTTAGATTTCCAA | 108 | 95 °C (5 s) 60 °C (25 s) | 80 °C | 18202194 |
| R | TTATCCAGCTTTGTATTGCAA | |||||
| LGALS8 | F | TGGGGACGGGAAGAGATCAC | 172 | 95 °C (5 s) 62 °C (25 s) | 82.5 °C | 28365246 |
| R | TGCCATAAATGCCCAGAGTGTC | |||||
| LGALS9 | F | CTTTCATCACCACCATTCTG | 91 | 95 °C (5 s) 62 °C (25 s) | 84 °C | 18202194 |
| R | ATGTGGAACCTCTGAGCACTG | |||||
| LGALS10 (CLC) | F | GGATGGCCAAGAATTTGAACTG | 82 | 95 °C (5 s) 62 °C (25 s) | 80 °C | 28365246 |
| R | GGTGTAAGAGGATTGGCCATTG | |||||
| LGALS12 | F | TGTGAGCCTGAGGGACCA | 111 | 95 °C (5 s) 65 °C (25 s) | 87 °C | 18202194 |
| R | GCTGAGATCAGTTTCTTCTGC | |||||
| NCF1 | F | GTCAGATGAAAGCAAAGCGA | 93 | 95 °C (5 s) 60 °C (25 s) | 85.5 °C | 23147401 |
| R | CATAGTTGGGCTCAGGGTCT | |||||
| NCF1 * | F | ACCCAGCCAGCACTATGTGT | 767 | N/A | N/A | 10754283 |
| R | AGTAGCCTGTGACGTCGTCT | |||||
| NCF2 * | F | CGAGGGAACCAGCTGATAGA | 747 | N/A | N/A | 10754283 |
| R | CATGGGAACACTGAGCTTCA | |||||
| ACTB | F | TCAGCAAGCAGGAGTATGACGAG | 265 | 95 °C (5 s) 60 °C (25 s) | 84 °C | 21029043 |
| R | ACATTGTGAACTTTGGGGGATG |
| Antigen | Host | Type | Conjugates | Dilution | Source | Catalog # |
|---|---|---|---|---|---|---|
| Galectin-1 | Mouse | monoclonal | N/A | 1:800 | Santa Cruz Biotechnology | sc-166618 |
| Galectin-3 | Rabbit | polyclonal | N/A | 1:200 | Santa Cruz Biotechnology | sc-20157 |
| Galectin-9 | Rabbit | monoclonal | N/A | 1:1000 | Abcam | ab227046 |
| Galectin-10 | Rabbit | monoclonal | N/A | 1:10,000 | Abcam | ab157475 |
| O-GlcNAc (RL2) | Mouse | monoclonal | N/A | 1:1000 | Invitrogen | MA1-072 |
| Mouse IgG (H + L) | Goat | polyclonal | HRP | 1:10,000 | Invitrogen | A16066 |
| Rabbit IgG (H + L) | Goat | polyclonal | HRP | 1:10,000 | Invitrogen | A16096 |


References
- Timoshenko, A.V.; Lanteigne, J.; Kozak, K. Extracellular stress stimuli alter galectin expression profiles and adhesion characteristics of HL-60 cells. Mol. Cell. Biochem. 2016, 413, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Vinnai, J.R.; Cumming, R.C.; Thompson, G.J.; Timoshenko, A.V. The association between oxidative stress-induced galectins and differentiation of human promyelocytic HL-60 cells. Exp. Cell Res. 2017, 355, 113–123. [Google Scholar] [CrossRef]
- Sherazi, A.A.; Jariwala, K.A.; Cybulski, A.N.; Lewis, J.W.; Karagiannis, J.; Cumming, R.C.; Timoshenko, A.V. Effects of global O-GlcNAcylation on galectin gene-expression profiles in human cancer cell lines. Anticancer Res. 2018, 38, 6691–6697. [Google Scholar] [CrossRef] [PubMed]
- Johannes, L.; Jacob, R.; Leffler, H. Galectins at a glance. J. Cell Sci. 2018, 131, jcs208884. [Google Scholar] [CrossRef] [Green Version]
- Verkerke, H.; Dias-Baruffi, M.; Cummings, R.D.; Arthur, C.M.; Stowell, S.R. Galectins: An ancient family of carbohydrate binding proteins with modern functions. Methods Mol. Biol. 2022, 2442, 1–40. [Google Scholar] [CrossRef]
- Timoshenko, A.V. (Ed.) Cell Biology of Galectins; MDPI: Basel, Switzerland, 2022; Available online: https://www.mdpi.com/books/pdfview/book/5635 (accessed on 22 September 2022).
- Timoshenko, A.V. Towards molecular mechanisms regulating the expression of galectins in cancer cells under microenvironmental stress conditions. Cell. Mol. Life Sci. 2015, 72, 4327–4340. [Google Scholar] [CrossRef]
- Abedin, M.J.; Kashio, Y.; Seki, M.; Nakamura, K.; Hirashima, M. Potential roles of galectins in myeloid differentiation into three different lineages. J. Leukoc. Biol. 2003, 73, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Robinson, B.S.; Arthur, C.M.; Evavold, B.; Roback, E.; Kamili, N.A.; Stowell, C.S.; Vallecillo-Zúniga, M.L.; Van Ry, P.M.; Dias-Baruffi, M.; Cummings, R.D.; et al. The sweet-side of leukocytes: Galectins as master regulators of neutrophil function. Front. Immunol. 2019, 10, 1762. [Google Scholar] [CrossRef]
- Barrow, H.; Guo, X.; Wandall, H.H.; Pedersen, J.W.; Fu, B.; Zhao, Q.; Chen, C.; Rhodes, J.M.; Yu, L.G. Serum galectin-2, -4, and -8 are greatly increased in colon and breast cancer patients and promote cancer cell adhesion to blood vascular endothelium. Clin. Cancer Res. 2011, 17, 7035–7046. [Google Scholar] [CrossRef] [Green Version]
- He, X.W.; Li, W.L.; Li, C.; Liu, P.; Shen, Y.G.; Zhu, M.; Jin, X.P. Serum levels of galectin-1, galectin-3, and galectin-9 are associated with large artery atherosclerotic stroke. Sci. Rep. 2017, 7, 40994. [Google Scholar] [CrossRef]
- Asiamah, E.K.; Vailati-Riboni, M.; Zhou, Z.; Xu, T.; Loor, J.J.; Schimmel, K.; Worku, M. Rumen-protected methionine supplementation during the peripartal period alters the expression of galectin genes associated with inflammation in peripheral neutrophils and secretion in plasma of Holstein Cows. J. Dairy Res. 2019, 86, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Ni, C.; Wang, Y.; Zheng, G.; Zhang, J.; Xu, Y. Age-related trabecular bone loss is associated with a decline in serum galectin-1 level. BMC Musculoskelet. Disord. 2021, 22, 394. [Google Scholar] [CrossRef] [PubMed]
- Hughes, R.C. Secretion of the galectin family of mammalian carbohydrate-binding proteins. Biochim. Biophys. Acta 1999, 1473, 172–185. [Google Scholar] [CrossRef]
- Popa, S.J.; Stewart, S.E.; Moreau, K. Unconventional secretion of annexins and galectins. Semin. Cell Dev. Biol. 2018, 83, 42–50. [Google Scholar] [CrossRef]
- Bond, M.R.; Hanover, J.A. A little sugar goes a long way: The cell biology of O-GlcNAc. J. Cell Biol. 2015, 208, 869–880. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Qian, K. Protein O-GlcNAcylation: Emerging mechanisms and functions. Nat. Rev. Mol. Cell Biol. 2017, 18, 452–465. [Google Scholar] [CrossRef] [Green Version]
- Wulff-Fuentes, E.; Berendt, R.R.; Massman, L.; Danner, L.; Malard, F.; Vora, J.; Kahsay, R.; Olivier-Van Stichelen, S. The human O-GlcNAcome database and meta-analysis. Sci. Data 2021, 8, 25. [Google Scholar] [CrossRef]
- Zachara, N.E.; O'Donnell, N.; Cheung, W.D.; Mercer, J.J.; Marth, J.D.; Hart, G.W. Dynamic O-GlcNAc modification of nucleocytoplasmic proteins in response to stress. J. Biol. Chem. 2004, 279, 30133–30142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tazhitdinova, R.; Timoshenko, A.V. The emerging role of galectins and O-GlcNAc homeostasis in processes of cellular differentiation. Cells 2020, 9, 1792. [Google Scholar] [CrossRef]
- Hart, G.W. Nutrient regulation of signaling and transcription. J. Biol. Chem. 2019, 294, 2211–2231. [Google Scholar] [CrossRef]
- Liu, K.; Paterson, A.J.; Konrad, R.J.; Parlow, A.F.; Jimi, S.; Roh, M.; Chin, E.; Kudlow, J.E. Streptozotocin, an O-GlcNAcase inhibitor, blunts insulin and growth hormone secretion. Mol. Cell. Endocrinol. 2002, 194, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Akimoto, Y.; Hart, G.W.; Wells, L.; Vosseller, K.; Yamamoto, K.; Munetomo, E.; Ohara-Imaizumi, M.; Nishiwaki, C.; Nagamatsu, S.; Hirano, H.; et al. Elevation of the post-translational modification of proteins by O-linked N-acetylglucosamine leads to deterioration of the glucose-stimulated insulin secretion in the pancreas of diabetic Goto-Kakizaki rats. Glycobiology 2006, 17, 127–140. [Google Scholar] [CrossRef] [Green Version]
- Timoshenko, A.V. The role of galectins and O-GlcNAc in regulating promyelocytic cell stemness and differentiation. Mol. Biol. Cell 2019, 30, 3075. [Google Scholar]
- Mathew, M.P.; Abramowitz, L.K.; Donaldson, J.G.; Hanover, J.A. Nutrient-responsive O-GlcNAcylation dynamically modulates the secretion of glycan-binding protein galectin 3. J. Biol. Chem. 2022, 298, 101743. [Google Scholar] [CrossRef]
- Gatie, M.I.; Spice, D.M.; Garha, A.; McTague, A.; Ahmer, M.; Timoshenko, A.V.; Kelly, G.M. O-GlcNAcylation and regulation of galectin-3 in extraembryonic endoderm differentiation. Biomolecules 2022, 12, 623. [Google Scholar] [CrossRef] [PubMed]
- Olins, A.L.; Olins, D.E. Cytoskeletal influences on nuclear shape in granulocytic HL-60 cells. BMC Cell Biol. 2004, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Baxter, S.S.; Carlson, L.A.; Mayer, A.M.; Hall, M.L.; Fay, M.J. Granulocytic differentiation of HL-60 promyelocytic leukemia cells is associated with increased expression of CUL5. In Vitro Cell. Dev. Biol. Anim. 2009, 45, 264–274. [Google Scholar] [CrossRef]
- Levy, R.; Rotrosen, D.; Nagauke, O.; Leto, T.L.; Malech, H.L. Induction of the respiratory burst in HL-60 cells. Correlation of function and protein expression. J. Immunol. 1990, 145, 174–181. [Google Scholar]
- Gloster, T.M.; Zandberg, W.F.; Heinonen, J.E.; Shen, D.L.; Deng, L.; Vocadlo, D.J. Hijacking a biosynthetic pathway yields a glycosyltransferase inhibitor within cells. Nat. Chem. Biol. 2011, 7, 174–181. [Google Scholar] [CrossRef] [Green Version]
- Gupta, D.; Shah, H.P.; Malu, K.; Berliner, N.; Gaines, P. Differentiation and characterization of myeloid cells. Curr. Protoc. Immunol. 2014, 104, 22F.5.1–22F.5.28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asthana, A.; Ramakrishnan, P.; Vicioso, Y.; Zhang, K.; Parameswaran, R. Hexosamine biosynthetic pathway inhibition leads to AML cell differentiation and cell death. Mol. Cancer Ther. 2018, 17, 2226–2237. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Bradley, S.S.; Dick, M.F.; Guglielmo, C.G.; Timoshenko, A.V. Seasonal and flight-related variation of galectin expression in heart, liver and flight muscles of yellow-rumped warblers (Setophaga coronata). Glycoconj. J. 2017, 34, 603–611. [Google Scholar] [CrossRef]
- Jariwala, K.A.; Sherazi, A.A.; Tazhitdinova, R.; Shum, K.; Guevorguian, P.; Karagiannis, J.; Staples, J.F.; Timoshenko, A.V. The association between increasing levels of O-GlcNAc and galectins in the liver tissue of hibernating thirteen-lined ground squirrels (Ictidomys tridecemlineatus). Cell Tissue Res. 2020, 381, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Timoshenko, A.V.; Cherenkevich, S.N.; Gabius, H.J. Viscum album agglutinin-induced aggregation of blood cells and the lectin effects on neutrophil function. Biomed. Pharmacother. 1995, 49, 153–158. [Google Scholar] [CrossRef]
- Tazhitdinova, R.; Timoshenko, A.V. The Glycobiological Landscape of Breast Cancer: Insights into Galectins and O-GlcNAc Homeostasis-Related Genes. PREPRINT (Version 1) Available at Research Square. 2021. Available online: https://www.researchgate.net/publication/354590048_The_glycobiological_landscape_of_breast_cancer_insights_into_galectins_and_O-GlcNAc_homeostasis-related_genes (accessed on 19 October 2022). [CrossRef]
- Tasseff, R.; Jensen, H.A.; Congleton, J.; Dai, D.; Rogers, K.V.; Sagar, A.; Bunaciu, R.P.; Yen, A.; Varner, J.D. An effective model of the retinoic acid induced HL-60 differentiation program. Sci. Rep. 2017, 7, 14327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timoshenko, A.V.; Gorudko, I.V.; Maslakova, O.V.; André, S.; Kuwabara, I.; Liu, F.T.; Kaltner, H.; Gabius, H.J. Analysis of selected blood and immune cell responses to carbohydrate-dependent surface binding of proto- and chimera-type galectins. Mol. Cell. Biochem. 2003, 250, 139–149. [Google Scholar] [CrossRef]
- Loftus, G.R. On interpretation of interactions. Mem. Cogn. 1978, 6, 312–319. [Google Scholar] [CrossRef] [Green Version]
- Sharma, N.S.; Saluja, A.K.; Banerjee, S. “Nutrient-sensing” and self-renewal: O-GlcNAc in a new role. J. Bioenerg. Biomembr. 2017, 50, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, M.A.; Emerald, B.S.; Ansari, S.A. Stem cell fate determination through protein O-GlcNAcylation. J. Biol. Chem. 2021, 296, 100035. [Google Scholar] [CrossRef]
- Shimizu, M.; Shibuya, H.; Tanaka, N. Enhanced O-GlcNAc modification induced by the Ras/MAPK/Cdk1 pathway is required for Sox2 protein expression and generation of cancer stem cells. Sci. Rep. 2022, 12, 2910. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, K.; Takahashi, I.; Tsuchiya, Y.; Hasegawa, M.; Kamemura, K. Characteristic increase in nucleocytoplasmic protein glycosylation by O-GlcNAc in 3T3-L1 adipocyte differentiation. Biochem. Biophys. Res. Commun. 2010, 398, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Andrés-Bergós, J.; Tardio, L.; Larranaga-Vera, A.; Gómez, R.; Herrero-Beaumont, G.; Largo, R. The increase in O-linked N-acetylglucosamine protein modification stimulates chondrogenic differentiation both In Vitro and In Vivo. J. Biol. Chem. 2012, 287, 33615–33628. [Google Scholar] [CrossRef] [Green Version]
- Koyama, T.; Kamemura, K. Global increase in O-linked N-acetylglucosamine modification promotes osteoblast differentiation. Exp. Cell Res. 2015, 338, 194–202. [Google Scholar] [CrossRef]
- McColgan, N.M.; Feeley, M.N.; Woodward, A.M.; Guindolet, D.; Argüeso, P. The O-GlcNAc modification promotes terminal differentiation of human corneal epithelial cells. Glycobiology 2020, 30, 872–880. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Parker, M.P.; Graw, S.; Novikova, L.V.; Fedosyuk, H.; Fontes, J.D.; Koestler, D.C.; Peterson, K.R.; Slawson, C. O-GlcNAc homeostasis contributes to cell fate decisions during hematopoiesis. J. Biol. Chem. 2019, 294, 1363–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bänfer, S.; Jacob, R. Galectins in intra- and extracellular vesicles. Biomolecules 2020, 10, 1232. [Google Scholar] [CrossRef]
- Davuluri, G.V.; Chen, C.C.; Chiu, Y.C.; Tsai, H.W.; Chiu, H.C.; Chen, Y.L.; Tsai, P.J.; Kuo, W.T.; Tsao, N.; Lin, Y.S.; et al. Autophagy drives galectin-1 secretion from tumor-associated macrophages facilitating hepatocellular carcinoma progression. Front. Cell Dev. Biol. 2021, 9, 741820. [Google Scholar] [CrossRef]
- Thijssen, V.L.; Heusschen, R.; Caers, J.; Griffioen, A.W. Galectin expression in cancer diagnosis and prognosis: A systematic review. Biochim. Biophys. Acta 2015, 1855, 235–247. [Google Scholar] [CrossRef]
- Van der Hoeven, N.W.; Hollander, M.R.; Yıldırım, C.; Jansen, M.F.; Teunissen, P.F.; Horrevoets, A.J.; van der Pouw Kraan, T.C.; van Royen, N. The emerging role of galectins in cardiovascular disease. Vascul. Pharmacol. 2016, 81, 31–41. [Google Scholar] [CrossRef]
- Yang, R.Y.; Yu, L.; Graham, J.L.; Hsu, D.K.; Lloyd, K.C.; Havel, P.J.; Liu, F.-T. Ablation of a galectin preferentially expressed in adipocytes increases lipolysis, reduces adiposity, and improves insulin sensitivity in mice. Proc. Natl. Acad. Sci. USA 2011, 108, 18696–18701. [Google Scholar] [CrossRef] [PubMed]
- Hart, C.; Chase, L.G.; Hajivandi, M.; Agnew, B. Metabolic labeling and click chemistry detection of glycoprotein markers of mesenchymal stem cell differentiation. Methods Mol. Biol. 2011, 698, 459–484. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McTague, A.; Tazhitdinova, R.; Timoshenko, A.V. O-GlcNAc-Mediated Regulation of Galectin Expression and Secretion in Human Promyelocytic HL-60 Cells Undergoing Neutrophilic Differentiation. Biomolecules 2022, 12, 1763. https://doi.org/10.3390/biom12121763
McTague A, Tazhitdinova R, Timoshenko AV. O-GlcNAc-Mediated Regulation of Galectin Expression and Secretion in Human Promyelocytic HL-60 Cells Undergoing Neutrophilic Differentiation. Biomolecules. 2022; 12(12):1763. https://doi.org/10.3390/biom12121763
Chicago/Turabian StyleMcTague, Adam, Rada Tazhitdinova, and Alexander V. Timoshenko. 2022. "O-GlcNAc-Mediated Regulation of Galectin Expression and Secretion in Human Promyelocytic HL-60 Cells Undergoing Neutrophilic Differentiation" Biomolecules 12, no. 12: 1763. https://doi.org/10.3390/biom12121763