
An Evolutionary Perspective on the Origin, Conservation and Binding Partner Acquisition of Tankyrases

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Acquisition
2.2. Sequence Alignments and Construction of Phylogenetic Trees
2.3. Structure Visualization and Prediction
2.4. Position-Specific Sequence Conservation Analysis of Tankyrases
2.5. Identification of Potential TBMs in Orthologs of TNKS Binders
2.6. Molecular Cloning
2.7. Protein Expression
2.8. Protein Purification
2.9. Activity Analysis by Western Blot
2.10. Analytical Size-Exclusion Chromatography
2.11. Differential Scanning Fluorimetry
2.12. Determination of the Dissociation Constants by FRET
3. Results
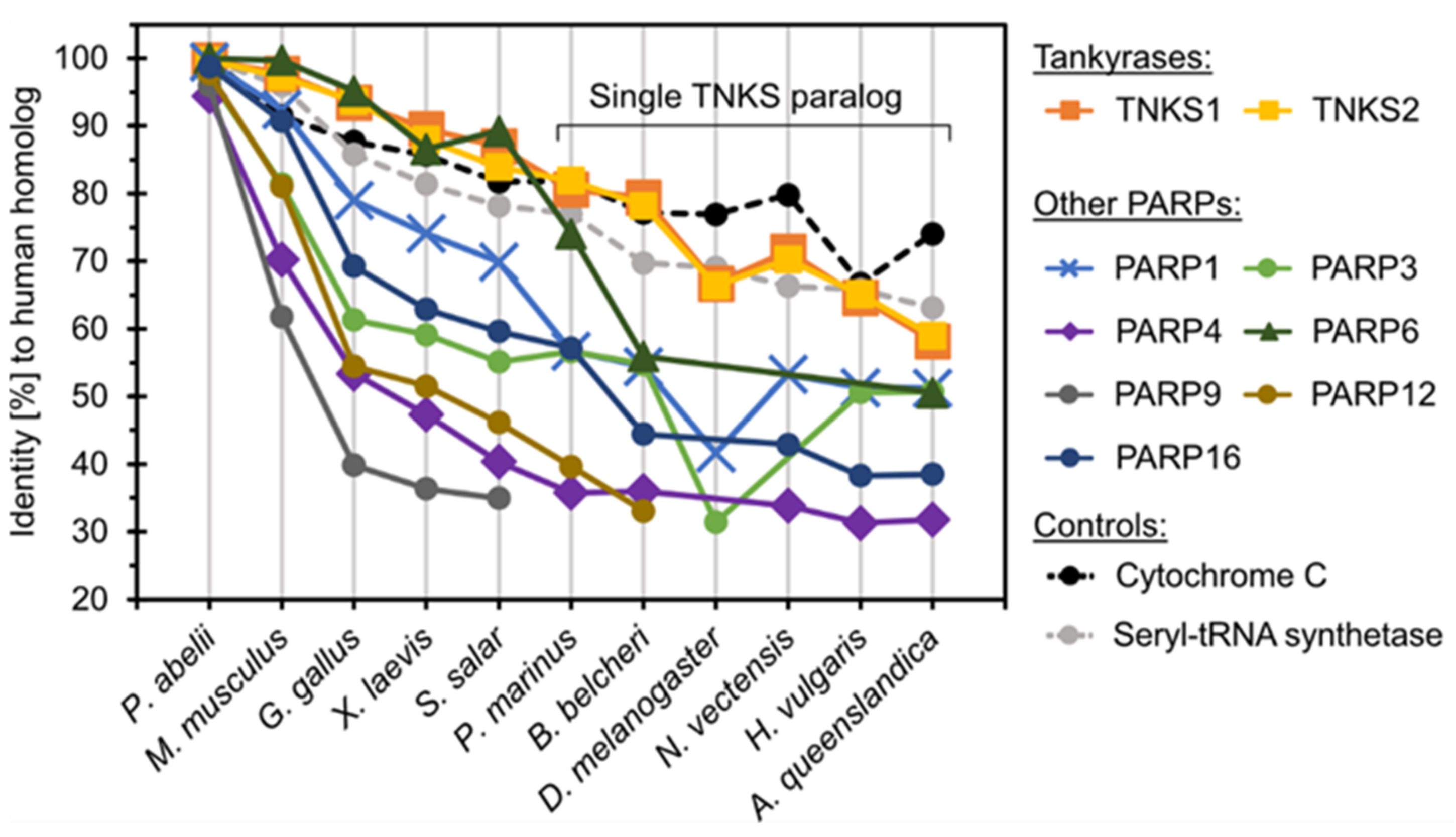
3.1. Distribution and Origin of Tankyrases
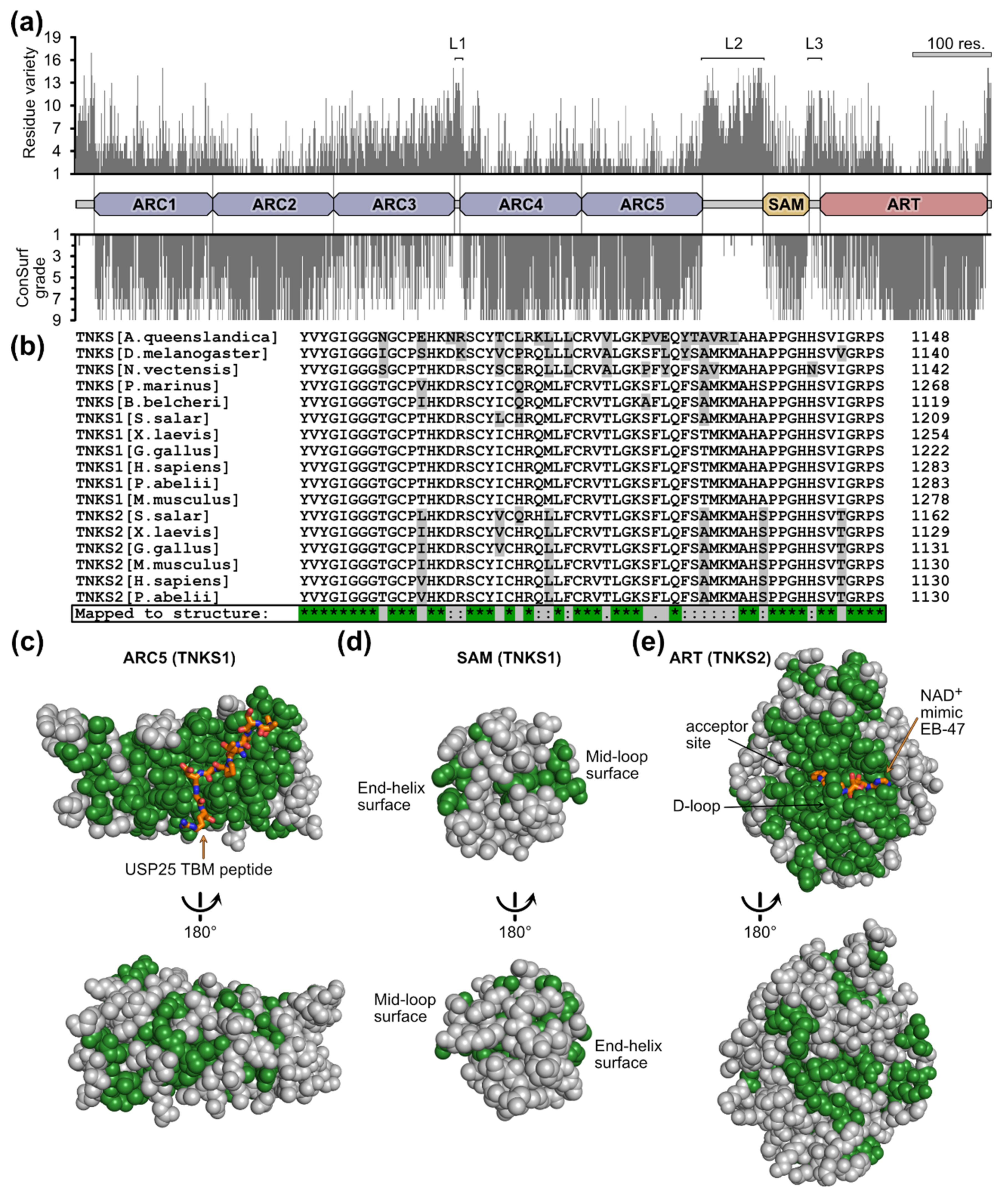
3.2. Conservation of Tankyrase Sequence and Structure
3.3. The Basic Molecular Characteristics of TNKSs Are Conserved
3.4. Acquisition of Tankyrase Binding Partners throughout Evolution

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TNKS Interaction Partner | Role or Cellular Function | TBM in Human Ortholog | Reference |
|---|---|---|---|
| AMOT | Hippo/YAP signaling | 77-RQEPQGQE | [11] |
| Arpin | Cell migration | 213-REQGDGAE | [84,85] |
| ATG9A | Pexophagy | 233-RLPGLGEA | [26] |
| Axin1 * | Wnt signaling | 22-RPPVPGEE | [10,14,16,31] |
| CASC3 | Pre-mRNA splicing | 146-RQSGDGQE | [15,82] |
| Dicer1 | RNA interference | 656-RELPDGTF | [86] |
| GMD | GDP-mannose 4,6 dehydratase | 12-RGSGDGEM | [24,87] |
| Golgin-45 | Golgi structure and protein maturation | 18-RGAGDGME | [15,82,83] |
| IRAP | Vesicle trafficking | 96-RQSPDGAC | [5,6,14,23] |
| Mcl-1 | Apoptosis regulation | 78-RPPPIGAE | [15,88] |
| MERIT40 | DNA damage repair | 28-RSNPEGAE 48-RSEGEGEA | [15,89] |
| NKD2 | Wnt signaling | 16-RESPEGDS | [86] |
| Notch2 | Notch signaling | 1726-RREPVGQD | [86] |
| NuMA | Mitotic spindle assembly | 1743-RTQPDGTS | [15,23,90] |
| PEX14 | Peroxisome homeostasis | 310-RMEVQGEE 350-RRGGDGQI | [26] |
| SH3BP2 | Osteoclast formation | 415-RSPPDGQS | [7,15] |
| SH3BP5 | Epithelial lumen formation | 269-RGCGVGAE 368-RSECSGAS | [91] |
| SOX9 | Chondrocyte differentiation | 257-RPLPEGGR 271-RDVDIGEL | [92] |
| Striatin | Dendritic Ca2+ signaling | 302-RSAGDGTD | [15,93] |
| TAB182 | Cytoskeletal maintenance | 1508-RPQPDGEA | [15,23] |
| TRF1 | Telomere maintenance | 13-RGCADGRD | [2,23,87] |
| USP25 | Ubiquitin protease | 1049-RTPADGR | [15,25] |
4. Discussion
4.1. Origin and Distribution of Tankyrases
4.2. Conservation of Tankyrases
4.3. Acquisition of Tankyrase Binding Partners
4.4. Implications for the Development of Therapeutics Targeting Tankyrases
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haikarainen, T.; Krauss, S.; Lehtiö, L. Tankyrases: Structure, Function and Therapeutic Implications in Cancer. Curr. Pharm. Des. 2014, 20, 6472–6488. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.; Giriat, I.; Schmitt, A.; de Lange, T. Tankyrase, a Poly(ADP-Ribose) Polymerase at Human Telomeres. Science 1998, 282, 1484–1487. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.; de Lange, T. Tankyrase Promotes Telomere Elongation in Human Cells. Curr. Biol. 2000, 10, 1299–1302. [Google Scholar] [CrossRef] [Green Version]
- Cook, B.D.; Dynek, J.N.; Chang, W.; Shostak, G.; Smith, S. Role for the Related Poly(ADP-Ribose) Polymerases Tankyrase 1 and 2 at Human Telomeres. Mol. Cell. Biol. 2002, 22, 332–342. [Google Scholar] [CrossRef] [Green Version]
- Yeh, T.-Y.J.; Sbodio, J.I.; Tsun, Z.-Y.; Luo, B.; Chi, N.-W. Insulin-Stimulated Exocytosis of GLUT4 Is Enhanced by IRAP and Its Partner Tankyrase. Biochem. J. 2007, 402, 279–290. [Google Scholar] [CrossRef] [Green Version]
- Chi, N.-W.; Lodish, H.F. Tankyrase Is a Golgi-Associated Mitogen-Activated Protein Kinase Substrate That Interacts with IRAP in GLUT4 Vesicles. J. Biol. Chem. 2000, 275, 38437–38444. [Google Scholar] [CrossRef] [Green Version]
- Levaot, N.; Voytyuk, O.; Dimitriou, I.; Sircoulomb, F.; Chandrakumar, A.; Deckert, M.; Krzyzanowski, P.M.; Scotter, A.; Gu, S.; Janmohamed, S.; et al. Loss of Tankyrase-Mediated Destruction of 3BP2 Is the Underlying Pathogenic Mechanism of Cherubism. Cell 2011, 147, 1324–1339. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.; Coughlin, M.; Mitchison, T.J. Tankyrase-1 Polymerization of Poly(ADP-Ribose) Is Required for Spindle Structure and Function. Nat. Cell Biol. 2005, 7, 1133–1139. [Google Scholar] [CrossRef]
- Chang, P.; Coughlin, M.; Mitchison, T.J. Interaction between Poly(ADP-Ribose) and NuMA Contributes to Mitotic Spindle Pole Assembly. Mol. Biol. Cell 2009, 20, 4575–4585. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.-M.A.; Mishina, Y.M.; Liu, S.; Cheung, A.; Stegmeier, F.; Michaud, G.A.; Charlat, O.; Wiellette, E.; Zhang, Y.; Wiessner, S.; et al. Tankyrase Inhibition Stabilizes Axin and Antagonizes Wnt Signalling. Nature 2009, 461, 614–620. [Google Scholar] [CrossRef]
- Wang, W.; Li, N.; Li, X.; Tran, M.K.; Han, X.; Chen, J. Tankyrase Inhibitors Target YAP by Stabilizing Angiomotin Family Proteins. Cell Rep. 2015, 13, 524–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, Y.J.; Hsiao, S.J.; Yver, D.; Cushman, S.W.; Tessarollo, L.; Smith, S.; Hodes, R.J. Tankyrase 1 and Tankyrase 2 Are Essential but Redundant for Mouse Embryonic Development. PLOS ONE 2008, 3, e2639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lüscher, B.; Ahel, I.; Altmeyer, M.; Ashworth, A.; Bai, P.; Chang, P.; Cohen, M.; Corda, D.; Dantzer, F.; Daugherty, M.D.; et al. ADP-ribosyltransferases, an Update on Function and Nomenclature. FEBS J. 2021. ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Eisemann, T.; McCauley, M.; Langelier, M.-F.; Gupta, K.; Roy, S.; Van Duyne, G.D.; Pascal, J.M. Tankyrase-1 Ankyrin Repeats Form an Adaptable Binding Platform for Targets of ADP-Ribose Modification. Structure 2016, 24, 1679–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guettler, S.; LaRose, J.; Petsalaki, E.; Gish, G.; Scotter, A.; Pawson, T.; Rottapel, R.; Sicheri, F. Structural Basis and Sequence Rules for Substrate Recognition by Tankyrase Explain the Basis for Cherubism Disease. Cell 2011, 147, 1340–1354. [Google Scholar] [CrossRef] [Green Version]
- Morrone, S.; Cheng, Z.; Moon, R.T.; Cong, F.; Xu, W. Crystal Structure of a Tankyrase-Axin Complex and Its Implications for Axin Turnover and Tankyrase Substrate Recruitment. Proc. Natl. Acad. Sci. USA 2012, 109, 1500–1505. [Google Scholar] [CrossRef] [Green Version]
- DaRosa, P.A.; Klevit, R.E.; Xu, W. Structural Basis for Tankyrase-RNF146 Interaction Reveals Noncanonical Tankyrase-Binding Motifs. Protein Sci. 2018, 27, 1057–1067. [Google Scholar] [CrossRef] [Green Version]
- DaRosa, P.A.; Ovchinnikov, S.; Xu, W.; Klevit, R.E. Structural Insights into SAM Domain-Mediated Tankyrase Oligomerization. Protein Sci. 2016, 25, 1744–1752. [Google Scholar] [CrossRef] [Green Version]
- Mariotti, L.; Templeton, C.M.; Ranes, M.; Paracuellos, P.; Cronin, N.; Beuron, F.; Morris, E.; Guettler, S. Tankyrase Requires SAM Domain-Dependent Polymerization to Support Wnt-β-Catenin Signaling. Mol. Cell 2016, 63, 498–513. [Google Scholar] [CrossRef] [Green Version]
- Riccio, A.A.; McCauley, M.; Langelier, M.-F.; Pascal, J.M. Tankyrase Sterile α Motif Domain Polymerization Is Required for Its Role in Wnt Signaling. Structure 2016, 24, 1573–1581. [Google Scholar] [CrossRef]
- Rycker, M.D.; Price, C.M. Tankyrase Polymerization Is Controlled by Its Sterile Alpha Motif and Poly(ADP-Ribose) Polymerase Domains. Mol. Cell. Biol. 2004, 24, 9802–9812. [Google Scholar] [CrossRef] [Green Version]
- Kaminker, P.G.; Kim, S.-H.; Taylor, R.D.; Zebarjadian, Y.; Funk, W.D.; Morin, G.B.; Yaswen, P.; Campisi, J. TANK2, a New TRF1-Associated Poly(ADP-Ribose) Polymerase, Causes Rapid Induction of Cell Death upon Overexpression. J. Biol. Chem. 2001, 276, 35891–35899. [Google Scholar] [CrossRef] [Green Version]
- Sbodio, J.I.; Chi, N.-W. Identification of a Tankyrase-Binding Motif Shared by IRAP, TAB182, and Human TRF1 but Not Mouse TRF1. J. Biol. Chem. 2002, 277, 31887–31892. [Google Scholar] [CrossRef] [Green Version]
- Bisht, K.K.; Dudognon, C.; Chang, W.G.; Sokol, E.S.; Ramirez, A.; Smith, S. GDP-Mannose-4,6-Dehydratase Is a Cytosolic Partner of Tankyrase 1 That Inhibits Its Poly(ADP-Ribose) Polymerase Activity. Mol. Cell. Biol. 2012, 32, 3044–3053. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Liu, J.; Fu, T.; Shan, B.; Qian, L.; Pan, L.; Yuan, J. USP25 Regulates Wnt Signaling by Controlling the Stability of Tankyrases. Genes Dev. 2017, 31, 1024–1035. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Han, H.; Zhou, M.-T.; Yang, B.; Ta, A.P.; Li, N.; Chen, J.; Wang, W. Proteomic Analysis of the Human Tankyrase Protein Interaction Network Reveals Its Role in Pexophagy. Cell Rep. 2017, 20, 737–749. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.; de Lange, T. Cell Cycle Dependent Localization of the Telomeric PARP, Tankyrase, to Nuclear Pore Complexes and Centrosomes. J. Cell Sci. 1999, 112, 3649–3656. [Google Scholar] [CrossRef]
- Muramatsu, Y.; Ohishi, T.; Sakamoto, M.; Tsuruo, T.; Seimiya, H. Cross-Species Difference in Telomeric Function of Tankyrase 1. Cancer Sci. 2007, 98, 850–857. [Google Scholar] [CrossRef]
- Qian, L.; Mahaffey, J.P.; Alcorn, H.L.; Anderson, K.V. Tissue-Specific Roles of Axin2 in the Inhibition and Activation of Wnt Signaling in the Mouse Embryo. Proc. Natl. Acad. Sci. USA 2011, 108, 8692–8697. [Google Scholar] [CrossRef] [Green Version]
- Kuusela, S.; Wang, H.; Wasik, A.A.; Suleiman, H.; Lehtonen, S. Tankyrase Inhibition Aggravates Kidney Injury in the Absence of CD2AP. Cell Death Dis. 2016, 7, e2302. [Google Scholar] [CrossRef]
- Feng, Y.; Li, X.; Ray, L.; Song, H.; Qu, J.; Lin, S.; Lin, X. The Drosophila Tankyrase Regulates Wg Signaling Depending on the Concentration of Daxin. Cell. Signal. 2014, 26, 1717–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, S.J.; Smith, S. Tankyrase Function at Telomeres, Spindle Poles, and Beyond. Biochimie 2008, 90, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K. Novel Insight into the Function of Tankyrase. Oncol. Lett. 2018, 16, 6895–6902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamudio-Martinez, E.; Herrera-Campos, A.B.; Muñoz, A.; Rodríguez-Vargas, J.M.; Oliver, F.J. Tankyrases as Modulators of Pro-Tumoral Functions: Molecular Insights and Therapeutic Opportunities. J. Exp. Clin. Cancer Res. CR 2021, 40, 144. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, Scalable Generation of High-quality Protein Multiple Sequence Alignments Using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The Rapid Generation of Mutation Data Matrices from Protein Sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Berman, H.M. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Baek, M.; DiMaio, F.; Anishchenko, I.; Dauparas, J.; Ovchinnikov, S.; Lee, G.R.; Wang, J.; Cong, Q.; Kinch, L.N.; Schaeffer, R.D.; et al. Accurate Prediction of Protein Structures and Interactions Using a Three-Track Neural Network. Science 2021, 373, 871–876. [Google Scholar] [CrossRef]
- Ashkenazy, H.; Abadi, S.; Martz, E.; Chay, O.; Mayrose, I.; Pupko, T.; Ben-Tal, N. ConSurf 2016: An Improved Methodology to Estimate and Visualize Evolutionary Conservation in Macromolecules. Nucleic Acids Res. 2016, 44, W344–W350. [Google Scholar] [CrossRef] [Green Version]
- Blum, M.; Chang, H.-Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro Protein Families and Domains Database: 20 Years On. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef]
- Sowa, S.T.; Vela-Rodríguez, C.; Galera-Prat, A.; Cázares-Olivera, M.; Prunskaite-Hyyryläinen, R.; Ignatev, A.; Lehtiö, L. A FRET-Based High-Throughput Screening Platform for the Discovery of Chemical Probes Targeting the Scaffolding Functions of Human Tankyrases. Sci. Rep. 2020, 10, 12357. [Google Scholar] [CrossRef]
- Sowa, S.T.; Lehtiö, L. The Zinc-Binding Motif in Tankyrases Is Required for the Structural Integrity of the Catalytic ADP-Ribosyltransferase Domain. Open Biol. 2022, 12, 210365. [Google Scholar] [CrossRef]
- Jeong, J.-Y.; Yim, H.-S.; Ryu, J.-Y.; Lee, H.S.; Lee, J.-H.; Seen, D.-S.; Kang, S.G. One-Step Sequence- and Ligation-Independent Cloning as a Rapid and Versatile Cloning Method for Functional Genomics Studies. Appl. Environ. Microbiol. 2012, 78, 5440–5443. [Google Scholar] [CrossRef] [Green Version]
- Hynes, M.F.; Quandt, J.; O’Connell, M.P.; Pühler, A. Direct Selection for Curing and Deletion of Rhizobium Plasmids Using Transposons Carrying the Bacillus Subtilis SacB Gene. Gene 1989, 78, 111–120. [Google Scholar] [CrossRef]
- Reyrat, J.-M.; Pelicic, V.; Gicquel, B.; Rappuoli, R. Counterselectable Markers: Untapped Tools for Bacterial Genetics and Pathogenesis. Infect. Immun. 1998, 66, 4011–4017. [Google Scholar] [CrossRef]
- Rial, D.V.; Ceccarelli, E.A. Removal of DnaK Contamination during Fusion Protein Purifications. Protein Expr. Purif. 2002, 25, 503–507. [Google Scholar] [CrossRef]
- Sowa, S.T.; Galera-Prat, A.; Wazir, S.; Alanen, H.I.; Maksimainen, M.M.; Lehtiö, L. A Molecular Toolbox for ADP-Ribosyl Binding Proteins. Cell Rep. Methods 2021, 1, 100121. [Google Scholar] [CrossRef]
- Brinch, S.A.; Amundsen-Isaksen, E.; Espada, S.; Hammarstrom, C.; Aizenshtadt, A.; Olsen, P.A.; Holmen, L.; Hoyem, M.; Scholz, H.; Grodeland, G.; et al. The Tankyrase Inhibitor OM-153 Demonstrates Anti-Tumor Efficacy and a Therapeutic Window in Mouse Models. Cancer Res. Commun. 2022, 2, 233–245. [Google Scholar] [CrossRef]
- Song, Y.; Rodgers, V.G.J.; Schultz, J.S.; Liao, J. Protein Interaction Affinity Determination by Quantitative FRET Technology. Biotechnol. Bioeng. 2012, 109, 2875–2883. [Google Scholar] [CrossRef] [PubMed]
- Citarelli, M.; Teotia, S.; Lamb, R.S. Evolutionary History of the Poly(ADP-Ribose) Polymerase Gene Family in Eukaryotes. BMC Evol. Biol. 2010, 10, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perina, D.; Mikoč, A.; Ahel, J.; Ćetković, H.; Žaja, R.; Ahel, I. Distribution of Protein Poly(ADP-Ribosyl)Ation Systems across All Domains of Life. DNA Repair 2014, 23, 4–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laumer, C.E.; Gruber-Vodicka, H.; Hadfield, M.G.; Pearse, V.B.; Riesgo, A.; Marioni, J.C.; Giribet, G. Support for a Clade of Placozoa and Cnidaria in Genes with Minimal Compositional Bias. eLife 2018, 7, e36278. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, M.; Begovic, E.; Chapman, J.; Putnam, N.H.; Hellsten, U.; Kawashima, T.; Kuo, A.; Mitros, T.; Salamov, A.; Carpenter, M.L.; et al. The Trichoplax Genome and the Nature of Placozoans. Nature 2008, 454, 955–960. [Google Scholar] [CrossRef] [Green Version]
- Miller, D.J.; Ball, E.E. Animal Evolution: Trichoplax, Trees, and Taxonomic Turmoil. Curr. Biol. 2008, 18, R1003–R1005. [Google Scholar] [CrossRef] [Green Version]
- Carr, M.; Leadbeater, B.S.C.; Hassan, R.; Nelson, M.; Baldauf, S.L. Molecular Phylogeny of Choanoflagellates, the Sister Group to Metazoa. Proc. Natl. Acad. Sci. USA 2008, 105, 16641–16646. [Google Scholar] [CrossRef] [Green Version]
- King, N.; Westbrook, M.J.; Young, S.L.; Kuo, A.; Abedin, M.; Chapman, J.; Fairclough, S.; Hellsten, U.; Isogai, Y.; Letunic, I.; et al. The Genome of the Choanoflagellate Monosiga Brevicollis and the Origin of Metazoans. Nature 2008, 451, 783–788. [Google Scholar] [CrossRef] [Green Version]
- Fairclough, S.R.; Chen, Z.; Kramer, E.; Zeng, Q.; Young, S.; Robertson, H.M.; Begovic, E.; Richter, D.J.; Russ, C.; Westbrook, M.J.; et al. Premetazoan Genome Evolution and the Regulation of Cell Differentiation in the Choanoflagellate Salpingoeca Rosetta. Genome Biol. 2013, 14, R15. [Google Scholar] [CrossRef] [Green Version]
- Richter, D.J.; Fozouni, P.; Eisen, M.B.; King, N. Gene Family Innovation, Conservation and Loss on the Animal Stem Lineage. eLife 2018, 7, e34226. [Google Scholar] [CrossRef]
- Nagy, A.; Patthy, L. Reassessing Domain Architecture Evolution of Metazoan Proteins: The Contribution of Different Evolutionary Mechanisms. Genes 2011, 2, 578–598. [Google Scholar] [CrossRef] [Green Version]
- Mosavi, L.K.; Cammett, T.J.; Desrosiers, D.C.; Peng, Z.-Y. The Ankyrin Repeat as Molecular Architecture for Protein Recognition. Protein Sci. Publ. Protein Soc. 2004, 13, 1435–1448. [Google Scholar] [CrossRef] [Green Version]
- Lehtiö, L.; Collins, R.; van den Berg, S.; Johansson, A.; Dahlgren, L.-G.; Hammarström, M.; Helleday, T.; Holmberg-Schiavone, L.; Karlberg, T.; Weigelt, J. Zinc Binding Catalytic Domain of Human Tankyrase 1. J. Mol. Biol. 2008, 379, 136–145. [Google Scholar] [CrossRef] [Green Version]
- Baba, M.L.; Darga, L.L.; Goodman, M.; Czelusniak, J. Evolution of Cytochromec Investigated by the Maximum Parsimony Method. J. Mol. Evol. 1981, 17, 197–213. [Google Scholar] [CrossRef]
- Härtlein, M.; Cusack, S. Structure, Function and Evolution of Seryl-TRNA Synthetases: Implications for the Evolution of Aminoacyl-TRNA Synthetases and the Genetic Code. J. Mol. Evol. 1995, 40, 519–530. [Google Scholar] [CrossRef]
- Daugherty, M.D.; Young, J.M.; Kerns, J.A.; Malik, H.S. Rapid Evolution of PARP Genes Suggests a Broad Role for ADP-Ribosylation in Host-Virus Conflicts. PLoS Genet. 2014, 10. [Google Scholar] [CrossRef] [Green Version]
- Pascal, J.M. The Comings and Goings of PARP-1 in Response to DNA Damage. DNA Repair 2018, 71, 177–182. [Google Scholar] [CrossRef]
- Haikarainen, T.; Narwal, M.; Joensuu, P.; Lehtiö, L. Evaluation and Structural Basis for the Inhibition of Tankyrases by PARP Inhibitors. ACS Med. Chem. Lett. 2014, 5, 18–22. [Google Scholar] [CrossRef] [Green Version]
- Seimiya, H.; Muramatsu, Y.; Smith, S.; Tsuruo, T. Functional Subdomain in the Ankyrin Domain of Tankyrase 1 Required for Poly(ADP-Ribosyl)Ation of TRF1 and Telomere Elongation. Mol. Cell. Biol. 2004, 24, 1944–1955. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.; Yarravarapu, N.; Chen, H.; Kulak, O.; Dasari, P.; Herbert, J.; Yamaguchi, K.; Lum, L.; Zhang, X. Regulation of Tankyrase Activity by a Catalytic Domain Dimer Interface. Biochem. Biophys. Res. Commun. 2018, 503, 1780–1785. [Google Scholar] [CrossRef]
- Cho-Park, P.F.; Steller, H. Proteasome Regulation by ADP-Ribosylation. Cell 2013, 153, 614–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Li, Z.; Lv, L.; Du, A.; Lin, Z.; Ye, X.; Lin, Y.; Lin, X. Tankyrase Regulates Apoptosis by Activating JNK Signaling in Drosophila. Biochem. Biophys. Res. Commun. 2018, 503, 2234–2239. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Tian, A.; Benchabane, H.; Tacchelly-Benites, O.; Yang, E.; Nojima, H.; Ahmed, Y. The ADP-Ribose Polymerase Tankyrase Regulates Adult Intestinal Stem Cell Proliferation during Homeostasis in Drosophila. Development 2016, 143, 1710–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bregman, H.; Chakka, N.; Guzman-Perez, A.; Gunaydin, H.; Gu, Y.; Huang, X.; Berry, V.; Liu, J.; Teffera, Y.; Huang, L.; et al. Discovery of Novel, Induced-Pocket Binding Oxazolidinones as Potent, Selective, and Orally Bioavailable Tankyrase Inhibitors. J. Med. Chem. 2013, 56, 4320–4342. [Google Scholar] [CrossRef]
- Chen, B.; Dodge, M.E.; Tang, W.; Lu, J.; Ma, Z.; Fan, C.-W.; Wei, S.; Hao, W.; Kilgore, J.; Williams, N.S.; et al. Small Molecule-Mediated Disruption of Wnt-Dependent Signaling in Tissue Regeneration and Cancer. Nat. Chem. Biol. 2009, 5, 100–107. [Google Scholar] [CrossRef] [Green Version]
- Leenders, R.G.G.; Brinch, S.A.; Sowa, S.T.; Amundsen-Isaksen, E.; Galera-Prat, A.; Murthy, S.; Aertssen, S.; Smits, J.N.; Nieczypor, P.; Damen, E.; et al. Development of a 1,2,4-Triazole-Based Lead Tankyrase Inhibitor: Part II. J. Med. Chem. 2021, 64, 17936–17949. [Google Scholar] [CrossRef]
- Voronkov, A.; Holsworth, D.D.; Waaler, J.; Wilson, S.R.; Ekblad, B.; Perdreau-Dahl, H.; Dinh, H.; Drewes, G.; Hopf, C.; Morth, J.P.; et al. Structural Basis and SAR for G007-LK, a Lead Stage 1,2,4-Triazole Based Specific Tankyrase 1/2 Inhibitor. J. Med. Chem. 2013, 56, 3012–3023. [Google Scholar] [CrossRef]
- Johannes, J.W.; Almeida, L.; Barlaam, B.; Boriack-Sjodin, P.A.; Casella, R.; Croft, R.A.; Dishington, A.P.; Gingipalli, L.; Gu, C.; Hawkins, J.L.; et al. Pyrimidinone Nicotinamide Mimetics as Selective Tankyrase and Wnt Pathway Inhibitors Suitable for in Vivo Pharmacology. ACS Med. Chem. Lett. 2015, 6, 254–259. [Google Scholar] [CrossRef] [Green Version]
- McGonigle, S.; Chen, Z.; Wu, J.; Chang, P.; Kolber-Simonds, D.; Ackermann, K.; Twine, N.C.; Shie, J.-L.; Miu, J.T.; Huang, K.-C.; et al. E7449: A Dual Inhibitor of PARP1/2 and Tankyrase1/2 Inhibits Growth of DNA Repair Deficient Tumors and Antagonizes Wnt Signaling. Oncotarget 2015, 6, 41307–41323. [Google Scholar] [CrossRef]
- Shultz, M.D.; Cheung, A.K.; Kirby, C.A.; Firestone, B.; Fan, J.; Chen, C.H.-T.; Chen, Z.; Chin, D.N.; DiPietro, L.; Fazal, A.; et al. Identification of NVP-TNKS656: The Use of Structure–Efficiency Relationships To Generate a Highly Potent, Selective, and Orally Active Tankyrase Inhibitor. J. Med. Chem. 2013, 56, 6495–6511. [Google Scholar] [CrossRef]
- Somoza, J.R.; Menon, S.; Schmidt, H.; Joseph-McCarthy, D.; Dessen, A.; Stahl, M.L.; Somers, W.S.; Sullivan, F.X. Structural and Kinetic Analysis of Escherichia Coli GDP-Mannose 4,6 Dehydratase Provides Insights into the Enzyme’s Catalytic Mechanism and Regulation by GDP-Fucose. Structure 2000, 8, 123–135. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, S.; Mickanin, C.; Feng, Y.; Charlat, O.; Michaud, G.A.; Schirle, M.; Shi, X.; Hild, M.; Bauer, A.; et al. RNF146 Is a Poly(ADP-Ribose)-Directed E3 Ligase That Regulates Axin Degradation and Wnt Signalling. Nat. Cell Biol. 2011, 13, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Yue, X.; Tiwari, N.; Zhu, L.; Ngo, H.D.T.; Lim, J.-M.; Gim, B.; Jing, S.; Wang, Y.; Qian, Y.; Lee, I. Tankyrase-1-Mediated Degradation of Golgin45 Regulates Glycosyltransferase Trafficking and Protein Glycosylation in Rab2-GTP-Dependent Manner. Commun. Biol. 2021, 4, 1370. [Google Scholar] [CrossRef] [PubMed]
- Fetics, S.; Thureau, A.; Campanacci, V.; Aumont-Nicaise, M.; Dang, I.; Gautreau, A.; Pérez, J.; Cherfils, J. Hybrid Structural Analysis of the Arp2/3 Regulator Arpin Identifies Its Acidic Tail as a Primary Binding Epitope. Structure 2016, 24, 252–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simanov, G.; Dang, I.; Fokin, A.I.; Oguievetskaia, K.; Campanacci, V.; Cherfils, J.; Gautreau, A.M. Arpin Regulates Migration Persistence by Interacting with Both Tankyrases and the Arp2/3 Complex. Int. J. Mol. Sci. 2021, 22, 4115. [Google Scholar] [CrossRef]
- Bhardwaj, A.; Yang, Y.; Ueberheide, B.; Smith, S. Whole Proteome Analysis of Human Tankyrase Knockout Cells Reveals Targets of Tankyrase-Mediated Degradation. Nat. Commun. 2017, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Eisemann, T.; Langelier, M.-F.; Pascal, J.M. Structural and Functional Analysis of Parameters Governing Tankyrase-1 Interaction with Telomeric Repeat-Binding Factor 1 and GDP-Mannose 4,6-Dehydratase. J. Biol. Chem. 2019, 294, 14574–14590. [Google Scholar] [CrossRef]
- Bae, J.; Donigian, J.R.; Hsueh, A.J.W. Tankyrase 1 Interacts with Mcl-1 Proteins and Inhibits Their Regulation of Apoptosis. J. Biol. Chem. 2003, 278, 5195–5204. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, K.; Ohishi, T.; Kuroiwa, M.; Iemura, S.; Natsume, T.; Seimiya, H. MERIT40-Dependent Recruitment of Tankyrase to Damaged DNA and Its Implication for Cell Sensitivity to DNA-Damaging Anticancer Drugs. Oncotarget 2018, 9, 35844–35855. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.; Dynek, J.N.; Smith, S. NuMA Is a Major Acceptor of Poly(ADP-Ribosyl)Ation by Tankyrase 1 in Mitosis. Biochem. J. 2005, 391, 177–184. [Google Scholar] [CrossRef]
- Chandrakumar, A.A.; Coyaud, É.; Marshall, C.B.; Ikura, M.; Raught, B.; Rottapel, R. Tankyrase Regulates Epithelial Lumen Formation via Suppression of Rab11 GEFs. J. Cell Biol. 2021, 220, e202008037. [Google Scholar] [CrossRef]
- Kim, S.; Han, S.; Kim, Y.; Kim, H.-S.; Gu, Y.-R.; Kang, D.; Cho, Y.; Kim, H.; Lee, J.; Seo, Y.; et al. Tankyrase Inhibition Preserves Osteoarthritic Cartilage by Coordinating Cartilage Matrix Anabolism via Effects on SOX9 PARylation. Nat. Commun. 2019, 10, 4898. [Google Scholar] [CrossRef] [Green Version]
- Lahav-Ariel, L.; Caspi, M.; Nadar-Ponniah, P.T.; Zelikson, N.; Hofmann, I.; Hanson, K.K.; Franke, W.W.; Sklan, E.H.; Avraham, K.B.; Rosin-Arbesfeld, R. Striatin Is a Novel Modulator of Cell Adhesion. FASEB J. 2019, 33, 4729–4740. [Google Scholar] [CrossRef]
- Zhou, Z.; Chan, C.H.; Xiao, Z.; Tan, E. Ring Finger Protein 146/Iduna Is a Poly(ADP-Ribose) Polymer Binding and PARsylation Dependent E3 Ubiquitin Ligase. Cell Adhes. Migr. 2011, 5, 463–471. [Google Scholar] [CrossRef]
- Callow, M.G.; Tran, H.; Phu, L.; Lau, T.; Lee, J.; Sandoval, W.N.; Liu, P.S.; Bheddah, S.; Tao, J.; Lill, J.R.; et al. Ubiquitin Ligase RNF146 Regulates Tankyrase and Axin to Promote Wnt Signaling. PloS ONE 2011, 6, e22595. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Michaud, G.A.; Cheng, Z.; Zhang, Y.; Hinds, T.R.; Fan, E.; Cong, F.; Xu, W. Recognition of the Iso-ADP-Ribose Moiety in Poly(ADP-Ribose) by WWE Domains Suggests a General Mechanism for Poly(ADP-Ribosyl)Ation-Dependent Ubiquitination. Genes Dev. 2012, 26, 235–240. [Google Scholar] [CrossRef] [Green Version]
- DaRosa, P.A.; Wang, Z.; Jiang, X.; Pruneda, J.N.; Cong, F.; Klevit, R.E.; Xu, W. Allosteric Activation of the RNF146 Ubiquitin Ligase by a Poly(ADP-Ribosyl)Ation Signal. Nature 2015, 517, 223–226. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Tacchelly-Benites, O.; Noble, G.P.; Johnson, M.K.; Gagné, J.-P.; Poirier, G.G.; Ahmed, Y. A Context-Dependent Role for the RNF146 Ubiquitin Ligase in Wingless/Wnt Signaling in Drosophila. Genetics 2019, 211, 913–923. [Google Scholar] [CrossRef]
- Paps, J.; Holland, P.W.H. Reconstruction of the Ancestral Metazoan Genome Reveals an Increase in Genomic Novelty. Nat. Commun. 2018, 9, 1730. [Google Scholar] [CrossRef] [Green Version]
- Ekman, D.; Björklund, Å.K.; Elofsson, A. Quantification of the Elevated Rate of Domain Rearrangements in Metazoa. J. Mol. Biol. 2007, 372, 1337–1348. [Google Scholar] [CrossRef]
- López-Escardó, D.; Grau-Bové, X.; Guillaumet-Adkins, A.; Gut, M.; Sieracki, M.E.; Ruiz-Trillo, I. Reconstruction of Protein Domain Evolution Using Single-Cell Amplified Genomes of Uncultured Choanoflagellates Sheds Light on the Origin of Animals. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20190088. [Google Scholar] [CrossRef] [PubMed]
- Tordai, H.; Nagy, A.; Farkas, K.; Bányai, L.; Patthy, L. Modules, Multidomain Proteins and Organismic Complexity: Mobile Domains. FEBS J. 2005, 272, 5064–5078. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, M.; Simakov, O.; Chapman, J.; Fahey, B.; Gauthier, M.E.A.; Mitros, T.; Richards, G.S.; Conaco, C.; Dacre, M.; Hellsten, U.; et al. The Amphimedon Queenslandica Genome and the Evolution of Animal Complexity. Nature 2010, 466, 720–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jespersen, N.; Barbar, E. Emerging Features of Linear Motif-Binding Hub Proteins. Trends Biochem. Sci. 2020, 45, 375–384. [Google Scholar] [CrossRef]
- Davey, N.E.; Van Roey, K.; Weatheritt, R.J.; Toedt, G.; Uyar, B.; Altenberg, B.; Budd, A.; Diella, F.; Dinkel, H.; Gibson, T.J. Attributes of Short Linear Motifs. Mol BioSyst 2012, 8, 268–281. [Google Scholar] [CrossRef]
- Van Roey, K.; Uyar, B.; Weatheritt, R.J.; Dinkel, H.; Seiler, M.; Budd, A.; Gibson, T.J.; Davey, N.E. Short Linear Motifs: Ubiquitous and Functionally Diverse Protein Interaction Modules Directing Cell Regulation. Chem. Rev. 2014, 114, 6733–6778. [Google Scholar] [CrossRef] [PubMed]
- Davey, N.E.; Cyert, M.S.; Moses, A.M. Short Linear Motifs - Ex Nihilo Evolution of Protein Regulation. Cell Commun. Signal. CCS 2015, 13, 43. [Google Scholar] [CrossRef] [Green Version]
- van der Lee, R.; Buljan, M.; Lang, B.; Weatheritt, R.J.; Daughdrill, G.W.; Dunker, A.K.; Fuxreiter, M.; Gough, J.; Gsponer, J.; Jones, D.T.; et al. Classification of Intrinsically Disordered Regions and Proteins. Chem. Rev. 2014, 114, 6589–6631. [Google Scholar] [CrossRef]
- Dynek, J.N.; Smith, S. Resolution of Sister Telomere Association Is Required for Progression Through Mitosis. Science 2004, 304, 97–100. [Google Scholar] [CrossRef]
- Ha, G.-H.; Kim, H.-S.; Go, H.; Lee, H.; Seimiya, H.; Chung, D.H.; Lee, C.-W. Tankyrase-1 Function at Telomeres and during Mitosis Is Regulated by Polo-like Kinase-1-Mediated Phosphorylation. Cell Death Differ. 2012, 19, 321–332. [Google Scholar] [CrossRef]
- Rippmann, J.F.; Damm, K.; Schnapp, A. Functional Characterization of the Poly(ADP-Ribose) Polymerase Activity of Tankyrase 1, a Potential Regulator of Telomere Length. J. Mol. Biol. 2002, 323, 217–224. [Google Scholar] [CrossRef]
- Donigian, J.R.; de Lange, T. The Role of the Poly(ADP-Ribose) Polymerase Tankyrase1 in Telomere Length Control by the TRF1 Component of the Shelterin Complex. J. Biol. Chem. 2007, 282, 22662–22667. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, S.J.; Poitras, M.F.; Cook, B.D.; Liu, Y.; Smith, S. Tankyrase 2 Poly(ADP-Ribose) Polymerase Domain-Deleted Mice Exhibit Growth Defects but Have Normal Telomere Length and Capping. Mol. Cell. Biol. 2006, 26, 2044–2054. [Google Scholar] [CrossRef] [Green Version]
- Seimiya, H.; Muramatsu, Y.; Ohishi, T.; Tsuruo, T. Tankyrase 1 as a Target for Telomere-Directed Molecular Cancer Therapeutics. Cancer Cell 2005, 7, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Thorvaldsen, T.E. Targeting Tankyrase to Fight WNT-Dependent Tumours. Basic Clin. Pharmacol. Toxicol. 2017, 121, 81–88. [Google Scholar] [CrossRef]






| Species | Common Name | Group | Notes |
|---|---|---|---|
| Homo sapiens | Modern human | Apes | Most studies of TNKS and its interaction partners were carried out in human model systems. |
| Pongo abelii | Sumatran orangutan | Apes | Extant ape species that is phylogenetically most distant from human. |
| Mus musculus | House mouse | Mammals | Most widely used mammalian model organism. |
| Gallus gallus | Red junglefowl | Birds | Includes the domesticated chicken. |
| Xenopus laevis | African clawed frog | Amphibians | Model amphibian. |
| Salmo salar | Atlantic salmon | Bony fishes | Representative of fishes with high-quality genome sequence data. |
| Petromyzon marinus | Sea lamprey | Jawless fishes | Model for vertebrate evolution. An early diverging vertebrate with only one TNKS instead of two paralogs (TNKS1/2). |
| Branchiostoma belcheri | Belcher’s lancelet | Lancelets | Closely related to vertebrates. Model organism for vertebrate evolution. |
| Drosophila melanogaster | Fruit fly | Insects | Popular model organism in genetics and developmental biology. |
| Caenorhabiditis elegans | C. elegans | Roundworms | Popular model organism for studying neuronal development. Gene-loss of TNKS and other PARPs was reported. |
| Nematostella vectensis | Starlet sea anemone | Cnidarians | Model cnidarian. |
| Hydra vulgaris | Fresh-water polyp | Cnidarians | Model organism in regenerative biology. |
| Amphimedon queenslandica | N/A | Sponges | Model for metazoan evolution and developmental biology. Functions from its TNKS ortholog were experimentally characterized in this work. |
| Salpingoeca kvevrii | N/A | Choanoflagellates | The most diverged TNKS orthologs identified in this work are from these choanoflagellates. |
| Salpingoeca macrocollata | |||
| Naegleria fowleri | Brain-eating amoeba | Percolozoa | Non-metazoan eukaryote that encodes an ART domain with high similarity to that of TNKSs. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sowa, S.T.; Bosetti, C.; Galera-Prat, A.; Johnson, M.S.; Lehtiö, L. An Evolutionary Perspective on the Origin, Conservation and Binding Partner Acquisition of Tankyrases. Biomolecules 2022, 12, 1688. https://doi.org/10.3390/biom12111688
Sowa ST, Bosetti C, Galera-Prat A, Johnson MS, Lehtiö L. An Evolutionary Perspective on the Origin, Conservation and Binding Partner Acquisition of Tankyrases. Biomolecules. 2022; 12(11):1688. https://doi.org/10.3390/biom12111688
Chicago/Turabian StyleSowa, Sven T., Chiara Bosetti, Albert Galera-Prat, Mark S. Johnson, and Lari Lehtiö. 2022. "An Evolutionary Perspective on the Origin, Conservation and Binding Partner Acquisition of Tankyrases" Biomolecules 12, no. 11: 1688. https://doi.org/10.3390/biom12111688