
Proteomic Profiling Reveals Increased Glycolysis, Decreased Oxidoreductase Activity and Fatty Acid Degradation in Skin Derived Fibroblasts from LHON Patients Bearing m.G11778A
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects and Fibroblasts Construction
2.2. Sample Preparation
2.3. LC-MS and MS Analysis
2.4. Data Processing
2.5. Bioinformatic Analysis
2.6. Data Availability
2.7. ROS Detection
2.8. Determination of Extra-Cellular Acidification Rate
3. Results
3.1. Identification of Different Expression Proteins (DEPs)
3.2. Annotation and Classification of Upregulated Proteins
3.3. Glycolysis Was Upregulated in LHON Fibroblasts (m.G11778A)
3.4. Pathway Interaction Network Analysis of Upregulated Proteins
3.5. Oxidoreductase Activity Was Downregulated in LHON Fibroblasts (m.G11778A)
3.6. Fatty Acid Metabolism Was Dysfunction in LHON Fibroblasts (m.G11778A)
3.7. Pathway Interaction Network Analysis of Downregulated Proteins
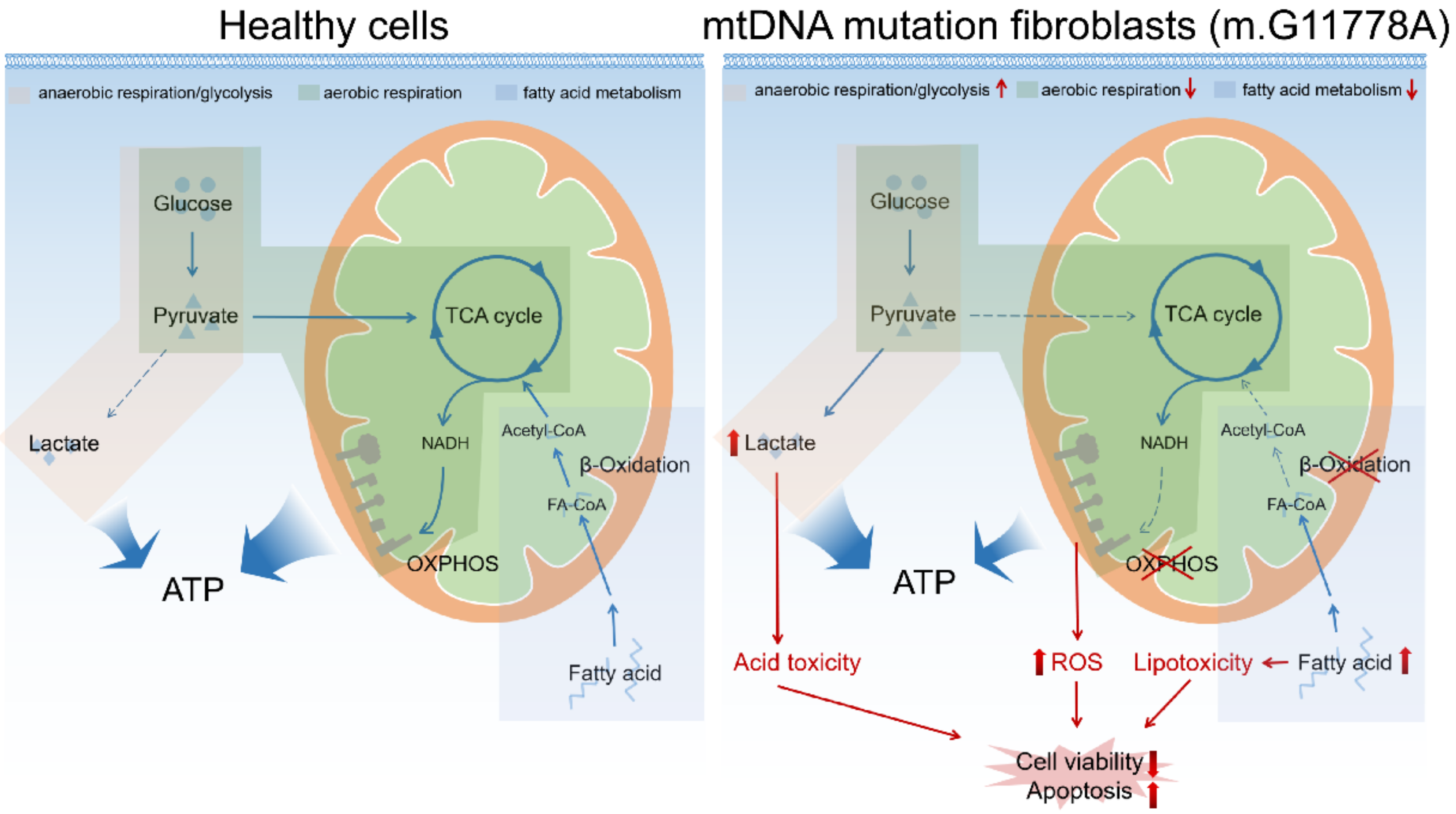
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu-Wai-Man, P.; Griffiths, P.G.; Howell, N.; Turnbull, D.M.; Chinnery, P.F. The Epidemiology of Leber Hereditary Optic Neuropathy in the North East of England. Am. J. Hum. Genet. 2016, 98, 1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hage, R.; Vignal-Clermont, C. Leber Hereditary Optic Neuropathy: Review of Treatment and Management. Front. Neurol. 2021, 12, 651639. [Google Scholar] [CrossRef] [PubMed]
- Sundaramurthy, S.; SelvaKumar, A.; Ching, J.; Dharani, V.; Sarangapani, S.; Yu-Wai-Man, P. Leber hereditary optic neuropathy-new insights and old challenges. Graefe’s Arch. Clin. Exp. Ophthalmol. 2021, 259, 2461–2472. [Google Scholar] [CrossRef] [PubMed]
- Yu-Wai-Man, P.; Chinnery, P.F. Leber Hereditary Optic Neuropathy. In GeneReviews((R)); Adam, M.P., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Gripp, K.W., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Wallace, D.C.; Singh, G.; Lott, M.T.; Hodge, J.A.; Schurr, T.G.; Lezza, A.M.; Elsas, L.J., 2nd; Nikoskelainen, E.K. Mitochondrial DNA mutation associated with Leber’s hereditary optic neuropathy. Science 1988, 242, 1427–1430. [Google Scholar] [CrossRef] [PubMed]
- Johns, D.R.; Neufeld, M.J.; Park, R.D. An ND-6 mitochondrial DNA mutation associated with Leber hereditary optic neuropathy. Biochem. Biophys. Res. Commun. 1992, 187, 1551–1557. [Google Scholar] [CrossRef]
- Huoponen, K.; Vilkki, J.; Aula, P.; Nikoskelainen, E.K.; Savontaus, M.L. A new mtDNA mutation associated with Leber hereditary optic neuroretinopathy. Am. J. Hum. Genet. 1991, 48, 1147–1153. [Google Scholar]
- Ma, Y.X.; Zhou, Y.G.; Zhang, J.P.; Zhang, Q.B.; Liu, W.L.; Ren, C.F.; Li, X.Y. Study on three common mitochondrial DNA mutations in Leber’s hereditary optic neuropathy. Chin. J. Med. Genet. 2012, 29, 519–523. [Google Scholar] [CrossRef]
- Ji, Y.; Zhang, J.; Yu, J.; Wang, Y.; Lu, Y.; Liang, M.; Li, Q.; Jin, X.; Wei, Y.; Meng, F.; et al. Contribution of mitochondrial ND1 3394T>C mutation to the phenotypic manifestation of Leber’s hereditary optic neuropathy. Hum. Mol. Genet. 2019, 28, 1515–1529. [Google Scholar] [CrossRef]
- Ji, Y.; Zhang, J.; Lu, Y.; Yi, Q.; Chen, M.; Xie, S.; Mao, X.; Xiao, Y.; Meng, F.; Zhang, M.; et al. Complex I mutations synergize to worsen the phenotypic expression of Leber’s hereditary optic neuropathy. J. Biol. Chem. 2020, 295, 13224–13238. [Google Scholar] [CrossRef]
- Xie, S.; Zhang, J.; Sun, J.; Zhang, M.; Zhao, F.; Wei, Q.P.; Tong, Y.; Liu, X.; Zhou, X.; Jiang, P.; et al. Mitochondrial haplogroup D4j specific variant m.11696G > a(MT-ND4) may increase the penetrance and expressivity of the LHON-associated m.11778G > a mutation in Chinese pedigrees. Mitochondrial DNA. Part A DNA Mapp. Seq. Anal. 2017, 28, 434–441. [Google Scholar] [CrossRef]
- Zhang, J.; Ji, Y.; Lu, Y.; Fu, R.; Xu, M.; Liu, X.; Guan, M.X. Leber’s hereditary optic neuropathy (LHON)-associated ND5 12338T > C mutation altered the assembly and function of complex I, apoptosis and mitophagy. Hum. Mol. Genet. 2018, 27, 1999–2011. [Google Scholar] [CrossRef]
- Jin, X.; Zhang, J.; Yi, Q.; Meng, F.; Yu, J.; Ji, Y.; Mo, J.Q.; Tong, Y.; Jiang, P.; Guan, M.X. Leber’s Hereditary Optic Neuropathy Arising from the Synergy Between ND1 3635G>A Mutation and Mitochondrial YARS2 Mutations. Investig. Ophthalmol. Vis. Sci. 2021, 62, 22. [Google Scholar] [CrossRef]
- Yu, J.; Liang, X.; Ji, Y.; Ai, C.; Liu, J.; Zhu, L.; Nie, Z.; Jin, X.; Wang, C.; Zhang, J.; et al. PRICKLE3 linked to ATPase biogenesis manifested Leber’s hereditary optic neuropathy. J. Clin. Investig. 2020, 130, 4935–4946. [Google Scholar] [CrossRef]
- Olesen, M.A.; Villavicencio-Tejo, F.; Quintanilla, R.A. The use of fibroblasts as a valuable strategy for studying mitochondrial impairment in neurological disorders. Transl. Neurodegener. 2022, 11, 36. [Google Scholar] [CrossRef]
- Yao, S.; Zhou, Q.; Yang, M.; Li, Y.; Jin, X.; Guo, Q.; Yang, L.; Qin, F.; Lei, B. Multi-mtDNA Variants May Be a Factor Contributing to Mitochondrial Function Variety in the Skin-Derived Fibroblasts of Leber’s Hereditary Optic Neuropathy Patients. Front. Mol. Neurosci. 2022, 15, 920221. [Google Scholar] [CrossRef]
- Tun, A.W.; Chaiyarit, S.; Kaewsutthi, S.; Katanyoo, W.; Chuenkongkaew, W.; Kuwano, M.; Tomonaga, T.; Peerapittayamongkol, C.; Thongboonkerd, V.; Lertrit, P. Profiling the mitochondrial proteome of Leber’s Hereditary Optic Neuropathy (LHON) in Thailand: Down-regulation of bioenergetics and mitochondrial protein quality control pathways in fibroblasts with the 11778G>A mutation. PLoS ONE 2014, 9, e106779. [Google Scholar] [CrossRef] [Green Version]
- Bertholet, A.M.; Delerue, T.; Millet, A.M.; Moulis, M.F.; David, C.; Daloyau, M.; Arnaune-Pelloquin, L.; Davezac, N.; Mils, V.; Miquel, M.C.; et al. Mitochondrial fusion/fission dynamics in neurodegeneration and neuronal plasticity. Neurobiol. Dis. 2016, 90, 3–19. [Google Scholar] [CrossRef]
- Mary, A.; Eysert, F.; Checler, F.; Chami, M. Mitophagy in Alzheimer’s disease: Molecular defects and therapeutic approaches. Mol. Psychiatry 2022, 1–15. [Google Scholar] [CrossRef]
- Wang, D.K.; Zheng, H.L.; Zhou, W.S.; Duan, Z.W.; Jiang, S.D.; Li, B.; Zheng, X.F.; Jiang, L.S. Mitochondrial Dysfunction in Oxidative Stress-Mediated Intervertebral Disc Degeneration. Orthop. Surg. 2022, 14, 1569–1582. [Google Scholar] [CrossRef]
- Jin, X.; Zhang, X.; Liu, J.; Wang, W.; Liu, M.; Yang, L.; Liu, G.; Qiu, R.; Yang, M.; Yao, S.; et al. Retinal Proteomic Alterations and Combined Transcriptomic-Proteomic Analysis in the Early Stages of Progression of a Mouse Model of X-Linked Retinoschisis. Cells 2022, 11, 2150. [Google Scholar] [CrossRef]
- Ma, J.; Chen, T.; Wu, S.; Yang, C.; Bai, M.; Shu, K.; Li, K.; Zhang, G.; Jin, Z.; He, F.; et al. iProX: An integrated proteome resource. Nucleic Acids Res. 2019, 47, D1211–D1217. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Liu, J.; Wang, W.; Li, J.; Liu, G.; Qiu, R.; Yang, M.; Liu, M.; Yang, L.; Du, X.; et al. Identification of age-associated proteins and functional alterations in human retinal pigment epithelium. Genom. Proteom. Bioinform. 2022. online ahead of print. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Rigoulet, M.; Bouchez, C.L.; Paumard, P.; Ransac, S.; Cuvellier, S.; Duvezin-Caubet, S.; Mazat, J.P.; Devin, A. Cell energy metabolism: An update. Biochim. Et Biophys. Acta. Bioenerg. 2020, 1861, 148276. [Google Scholar] [CrossRef] [PubMed]
- De Moura, M.B.; Van Houten, B. Bioenergetic analysis of intact mammalian cells using the Seahorse XF24 Extracellular Flux analyzer and a luciferase ATP assay. Methods Mol. Biol. 2014, 1105, 589–602. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.; de Moura, M.B.; Lin, Y.; Vincent, G.; Thorne, S.; Duncan, L.M.; Hui-Min, L.; Kirkwood, J.M.; Becker, D.; Van Houten, B.; et al. Importance of glycolysis and oxidative phosphorylation in advanced melanoma. Mol. Cancer 2012, 11, 76. [Google Scholar] [CrossRef] [Green Version]
- Geldon, S.; Fernandez-Vizarra, E.; Tokatlidis, K. Redox-Mediated Regulation of Mitochondrial Biogenesis, Dynamics, and Respiratory Chain Assembly in Yeast and Human Cells. Front. Cell Dev. Biol. 2021, 9, 720656. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, L.F.; Cui, P.F.; Qi, L.Y.; Xing, L.; Jiang, H.L. Pathologically Responsive Mitochondrial Gene Therapy in an Allotopic Expression-Independent Manner Cures Leber’s Hereditary Optic Neuropathy. Adv. Mater. 2021, 33, e2103307. [Google Scholar] [CrossRef]
- Lin, C.S.; Sharpley, M.S.; Fan, W.; Waymire, K.G.; Sadun, A.A.; Carelli, V.; Ross-Cisneros, F.N.; Baciu, P.; Sung, E.; McManus, M.J.; et al. Mouse mtDNA mutant model of Leber hereditary optic neuropathy. Proc. Natl. Acad. Sci. USA 2012, 109, 20065–20070. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, G.; Cortopassi, G. Oxidative stress in inherited mitochondrial diseases. Free Radic. Biol. Med. 2015, 88, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Li, M.X.; Dewson, G. Mitochondria and apoptosis: Emerging concepts. F1000Prime Rep. 2015, 7, 42. [Google Scholar] [CrossRef]
- Martinou, J.C.; Youle, R.J. Mitochondria in apoptosis: Bcl-2 family members and mitochondrial dynamics. Dev. Cell 2011, 21, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Jiao, L.; Du, X.; Li, Y.; Jiao, Q.; Jiang, H. Role of Mitophagy in neurodegenerative Diseases and potential tagarts for Therapy. Mol. Biol. Rep. 2022. online ahead of print. [Google Scholar] [CrossRef]
- Galloway, C.A.; Yoon, Y. Mitochondrial morphology in metabolic diseases. Antioxid. Redox Signal. 2013, 19, 415–430. [Google Scholar] [CrossRef] [Green Version]
- Vujic, A.; Koo, A.N.M.; Prag, H.A.; Krieg, T. Mitochondrial redox and TCA cycle metabolite signaling in the heart. Free Radic. Biol. Med. 2021, 166, 287–296. [Google Scholar] [CrossRef]
- Miwa, S.; Kashyap, S.; Chini, E.; von Zglinicki, T. Mitochondrial dysfunction in cell senescence and aging. J. Clin. Investig. 2022, 132, e158447. [Google Scholar] [CrossRef]
- Hill, S.; Van Remmen, H. Mitochondrial stress signaling in longevity: A new role for mitochondrial function in aging. Redox Biol. 2014, 2, 936–944. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; An, P.; Gu, Z.; Luo, Y.; Luo, J. Mitochondrial Metal Ion Transport in Cell Metabolism and Disease. Int. J. Mol. Sci. 2021, 22, 7525. [Google Scholar] [CrossRef]
- Pathak, T.; Trebak, M. Mitochondrial Ca(2+) signaling. Pharmacol. Ther. 2018, 192, 112–123. [Google Scholar] [CrossRef]
- Xue, C.; Dong, N.; Shan, A. Putative role of STING-mitochondria associated membrane crosstalk in immunity. Trends Immunol. 2022, 43, 513–522. [Google Scholar] [CrossRef]
- Peng, J.; Ghosh, D.; Pang, J.; Zhang, L.; Yin, S.; Jiang, Y. Intertwined Relation between the Endoplasmic Reticulum and Mitochondria in Ischemic Stroke. Oxidative Med. Cell. Longev. 2022, 2022, 3335887. [Google Scholar] [CrossRef] [PubMed]
- Kishita, Y.; Ishikawa, K.; Nakada, K.; Hayashi, J.I.; Fushimi, T.; Shimura, M.; Kohda, M.; Ohtake, A.; Murayama, K.; Okazaki, Y. A high mutation load of m.14597A>G in MT-ND6 causes Leigh syndrome. Sci. Rep. 2021, 11, 11123. [Google Scholar] [CrossRef] [PubMed]
- Morvan, D.; Demidem, A. NMR metabolomics of fibroblasts with inherited mitochondrial Complex I mutation reveals treatment-reversible lipid and amino acid metabolism alterations. Metab. Off. J. Metab. Soc. 2018, 14, 55. [Google Scholar] [CrossRef] [PubMed]
- Magini, A.; Urbanelli, L.; Ciccarone, V.; Tancini, B.; Polidoro, M.; Timperio, A.M.; Zolla, L.; Tedde, A.; Sorbi, S.; Emiliani, C. Fibroblasts from PS1 mutated pre-symptomatic subjects and Alzheimer’s disease patients share a unique protein levels profile. J. Alzheimer’s Dis.: JAD 2010, 21, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Drabik, K.; Malinska, D.; Piecyk, K.; Debska-Vielhaber, G.; Vielhaber, S.; Duszynski, J.; Szczepanowska, J. Effect of Chronic Stress Present in Fibroblasts Derived from Patients with a Sporadic Form of AD on Mitochondrial Function and Mitochondrial Turnover. Antioxidants 2021, 10, 938. [Google Scholar] [CrossRef]
- Teves, J.M.Y.; Bhargava, V.; Kirwan, K.R.; Corenblum, M.J.; Justiniano, R.; Wondrak, G.T.; Anandhan, A.; Flores, A.J.; Schipper, D.A.; Khalpey, Z.; et al. Parkinson’s Disease Skin Fibroblasts Display Signature Alterations in Growth, Redox Homeostasis, Mitochondrial Function, and Autophagy. Front. Neurosci. 2017, 11, 737. [Google Scholar] [CrossRef] [Green Version]
- Yakhine-Diop, S.M.S.; Niso-Santano, M.; Rodriguez-Arribas, M.; Gomez-Sanchez, R.; Martinez-Chacon, G.; Uribe-Carretero, E.; Navarro-Garcia, J.A.; Ruiz-Hurtado, G.; Aiastui, A.; Cooper, J.M.; et al. Impaired Mitophagy and Protein Acetylation Levels in Fibroblasts from Parkinson’s Disease Patients. Mol. Neurobiol. 2019, 56, 2466–2481. [Google Scholar] [CrossRef]
- Jedrak, P.; Mozolewski, P.; Wegrzyn, G.; Wieckowski, M.R. Mitochondrial alterations accompanied by oxidative stress conditions in skin fibroblasts of Huntington’s disease patients. Metab. Brain Dis. 2018, 33, 2005–2017. [Google Scholar] [CrossRef] [Green Version]
- Del Hoyo, P.; Garcia-Redondo, A.; de Bustos, F.; Molina, J.A.; Sayed, Y.; Alonso-Navarro, H.; Caballero, L.; Arenas, J.; Jimenez-Jimenez, F.J. Oxidative stress in skin fibroblasts cultures of patients with Huntington’s disease. Neurochem. Res. 2006, 31, 1103–1109. [Google Scholar] [CrossRef]
- Onesto, E.; Colombrita, C.; Gumina, V.; Borghi, M.O.; Dusi, S.; Doretti, A.; Fagiolari, G.; Invernizzi, F.; Moggio, M.; Tiranti, V.; et al. Gene-specific mitochondria dysfunctions in human TARDBP and C9ORF72 fibroblasts. Acta Neuropathol. Commun. 2016, 4, 47. [Google Scholar] [CrossRef] [Green Version]
- Rizzardini, M.; Mangolini, A.; Lupi, M.; Ubezio, P.; Bendotti, C.; Cantoni, L. Low levels of ALS-linked Cu/Zn superoxide dismutase increase the production of reactive oxygen species and cause mitochondrial damage and death in motor neuron-like cells. J. Neurol. Sci. 2005, 232, 95–103. [Google Scholar] [CrossRef]
- Diaz-Ruiz, R.; Rigoulet, M.; Devin, A. The Warburg and Crabtree effects: On the origin of cancer cell energy metabolism and of yeast glucose repression. Biochim. Et Biophys. Acta 2011, 1807, 568–576. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Moncada, I.; Ruminot, I.; Robles-Maldonado, D.; Alegria, K.; Deitmer, J.W.; Barros, L.F. Neuronal control of astrocytic respiration through a variant of the Crabtree effect. Proc. Natl. Acad. Sci. USA 2018, 115, 1623–1628. [Google Scholar] [CrossRef] [Green Version]
- Denzel, M.S.; Storm, N.J.; Gutschmidt, A.; Baddi, R.; Hinze, Y.; Jarosch, E.; Sommer, T.; Hoppe, T.; Antebi, A. Hexosamine pathway metabolites enhance protein quality control and prolong life. Cell 2014, 156, 1167–1178. [Google Scholar] [CrossRef] [Green Version]
- Wong, A.; Cavelier, L.; Collins-Schramm, H.E.; Seldin, M.F.; McGrogan, M.; Savontaus, M.L.; Cortopassi, G.A. Differentiation-specific effects of LHON mutations introduced into neuronal NT2 cells. Hum. Mol. Genet. 2002, 11, 431–438. [Google Scholar] [CrossRef] [Green Version]
- Worth, A.J.; Basu, S.S.; Snyder, N.W.; Mesaros, C.; Blair, I.A. Inhibition of neuronal cell mitochondrial complex I with rotenone increases lipid beta-oxidation, supporting acetyl-coenzyme A levels. J. Biol. Chem. 2014, 289, 26895–26903. [Google Scholar] [CrossRef] [Green Version]
- Leong, D.W.; Komen, J.C.; Hewitt, C.A.; Arnaud, E.; McKenzie, M.; Phipson, B.; Bahlo, M.; Laskowski, A.; Kinkel, S.A.; Davey, G.M.; et al. Proteomic and metabolomic analyses of mitochondrial complex I-deficient mouse model generated by spontaneous B2 short interspersed nuclear element (SINE) insertion into NADH dehydrogenase (ubiquinone) Fe-S protein 4 (Ndufs4) gene. J. Biol. Chem. 2012, 287, 20652–20663. [Google Scholar] [CrossRef] [Green Version]
- Schonfeld, P.; Wojtczak, L. Fatty acids as modulators of the cellular production of reactive oxygen species. Free Radic. Biol. Med. 2008, 45, 231–241. [Google Scholar] [CrossRef]
- Rial, E.; Rodriguez-Sanchez, L.; Gallardo-Vara, E.; Zaragoza, P.; Moyano, E.; Gonzalez-Barroso, M.M. Lipotoxicity, fatty acid uncoupling and mitochondrial carrier function. Biochim. Biophys. Acta 2010, 1797, 800–806. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, S.; Zhang, X.; Jin, X.; Yang, M.; Li, Y.; Yang, L.; Xu, J.; Lei, B. Proteomic Profiling Reveals Increased Glycolysis, Decreased Oxidoreductase Activity and Fatty Acid Degradation in Skin Derived Fibroblasts from LHON Patients Bearing m.G11778A. Biomolecules 2022, 12, 1568. https://doi.org/10.3390/biom12111568
Yao S, Zhang X, Jin X, Yang M, Li Y, Yang L, Xu J, Lei B. Proteomic Profiling Reveals Increased Glycolysis, Decreased Oxidoreductase Activity and Fatty Acid Degradation in Skin Derived Fibroblasts from LHON Patients Bearing m.G11778A. Biomolecules. 2022; 12(11):1568. https://doi.org/10.3390/biom12111568
Chicago/Turabian StyleYao, Shun, Xiaoli Zhang, Xiuxiu Jin, Mingzhu Yang, Ya Li, Lin Yang, Jin Xu, and Bo Lei. 2022. "Proteomic Profiling Reveals Increased Glycolysis, Decreased Oxidoreductase Activity and Fatty Acid Degradation in Skin Derived Fibroblasts from LHON Patients Bearing m.G11778A" Biomolecules 12, no. 11: 1568. https://doi.org/10.3390/biom12111568