
Heat Shock Protein Upregulation Supplemental to Complex mRNA Alterations in Autoimmune Glaucoma
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
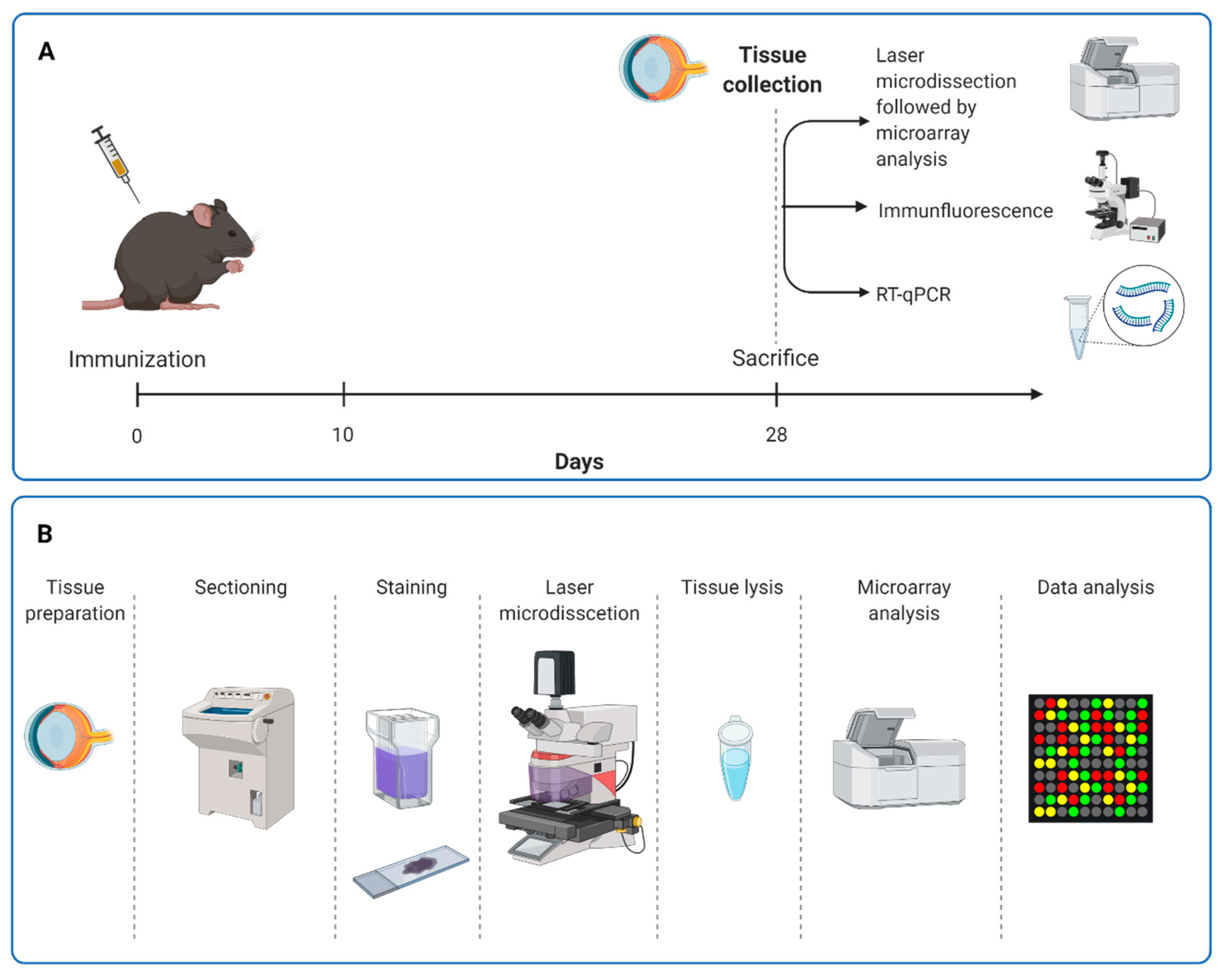
2.1. Animals and Immunizations
2.2. Electroretinography
2.3. Quantitative Real-Time PCR
2.4. Immunohistology of Retinal Sections
2.5. Evaluation of Immunohistological Stainings
2.6. Sample Preparation for mRNA Microarrays
2.7. Microarray Analyses
2.8. Statistics
3. Results
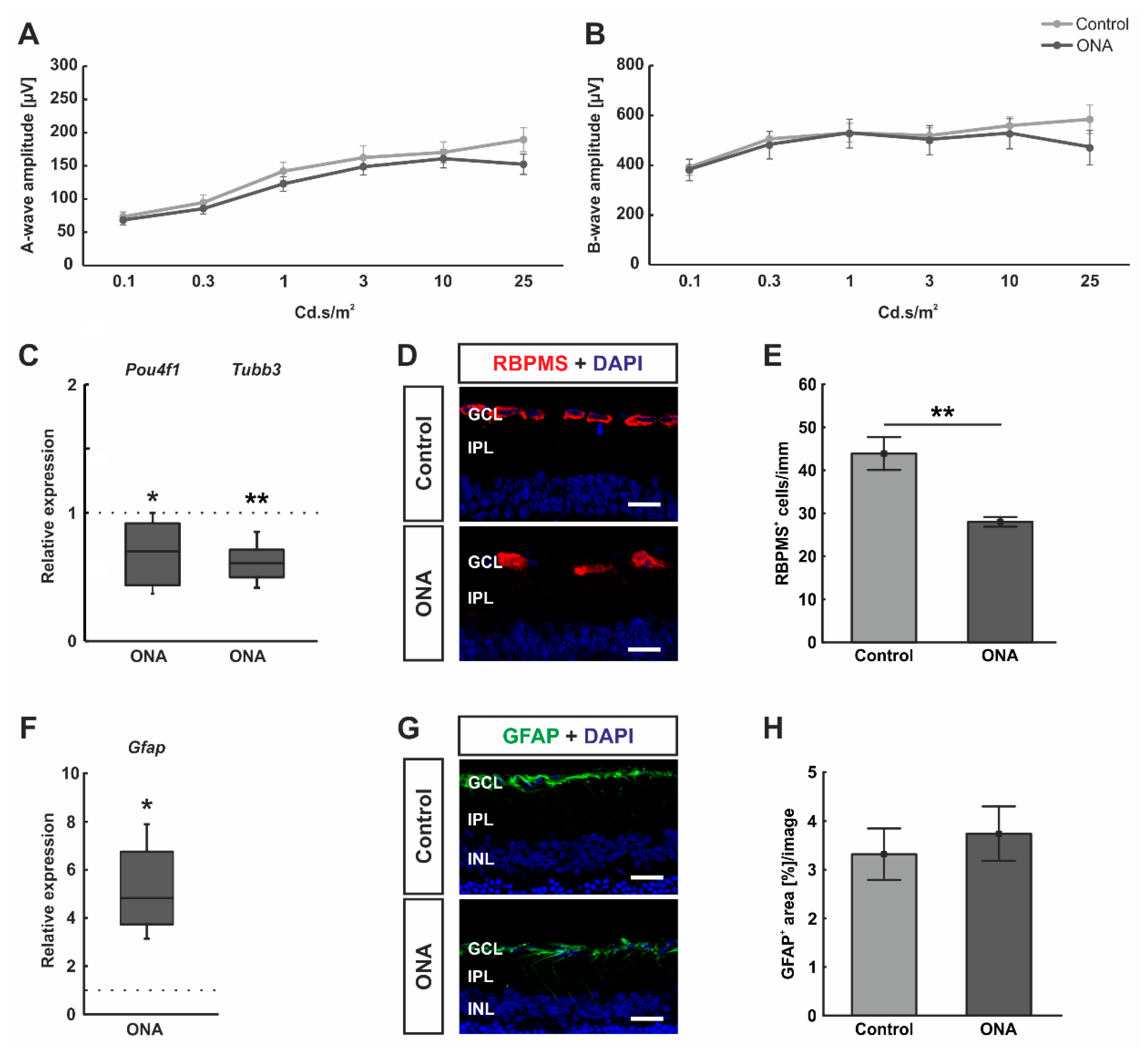
3.1. No Functional Alterations after Immunization, but Loss of Retinal Ganglion Cells
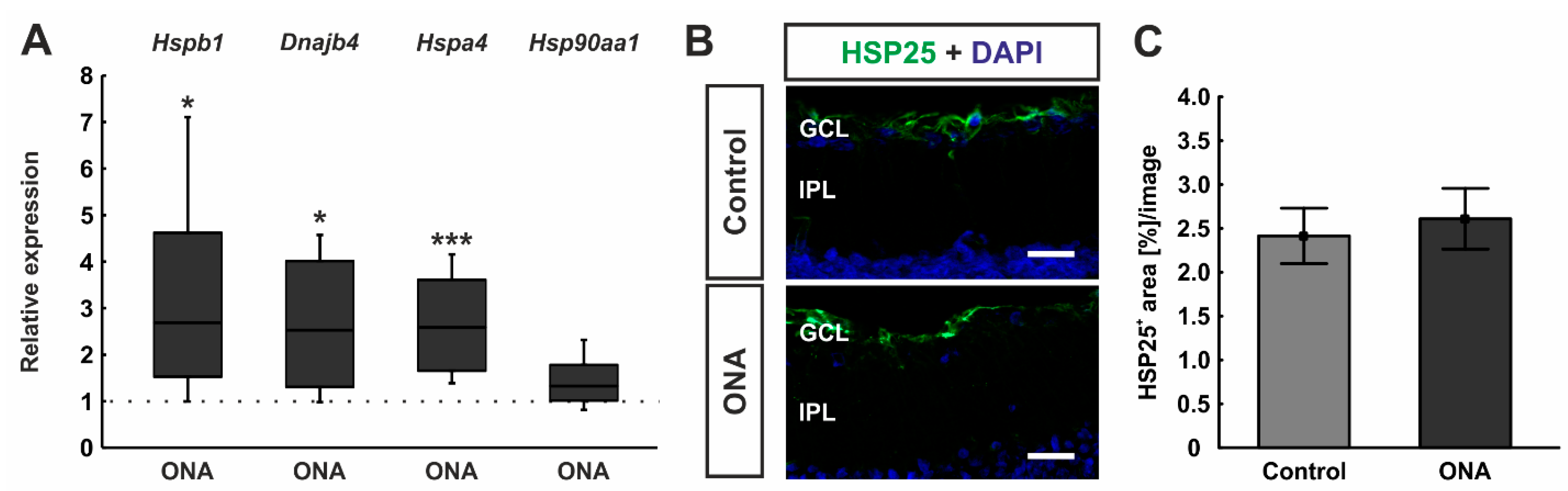
3.2. Regulation of HSPs after ONA Immunization
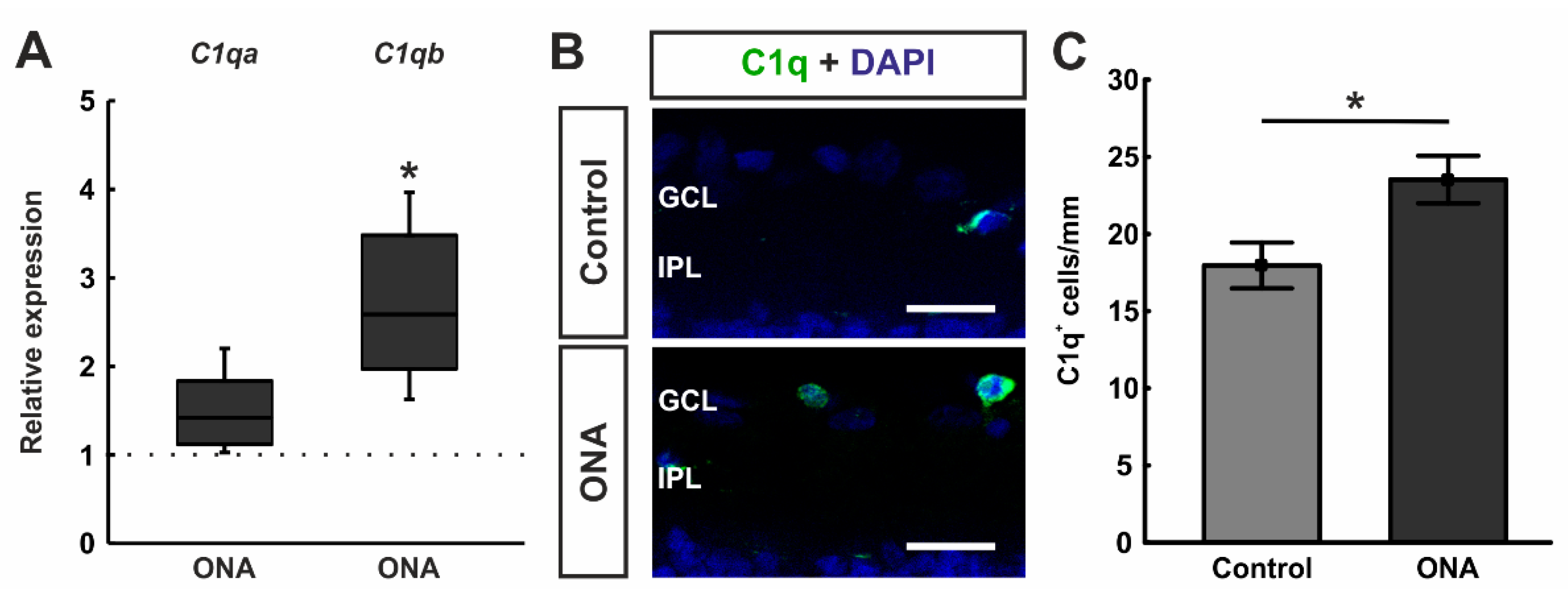
3.3. Involvement of Classical Complement Pathway
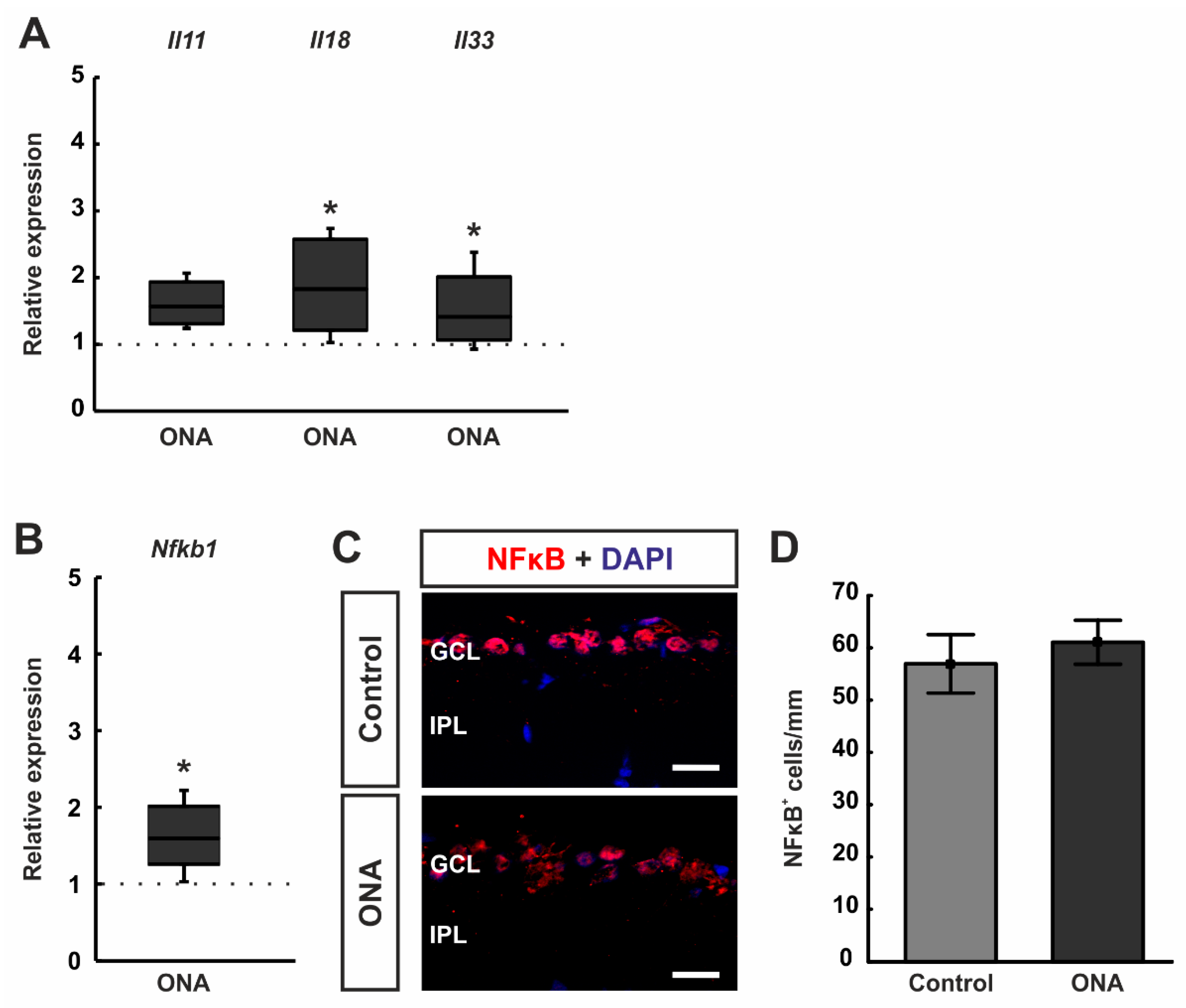
3.4. Increased Inflammatory Cytokines
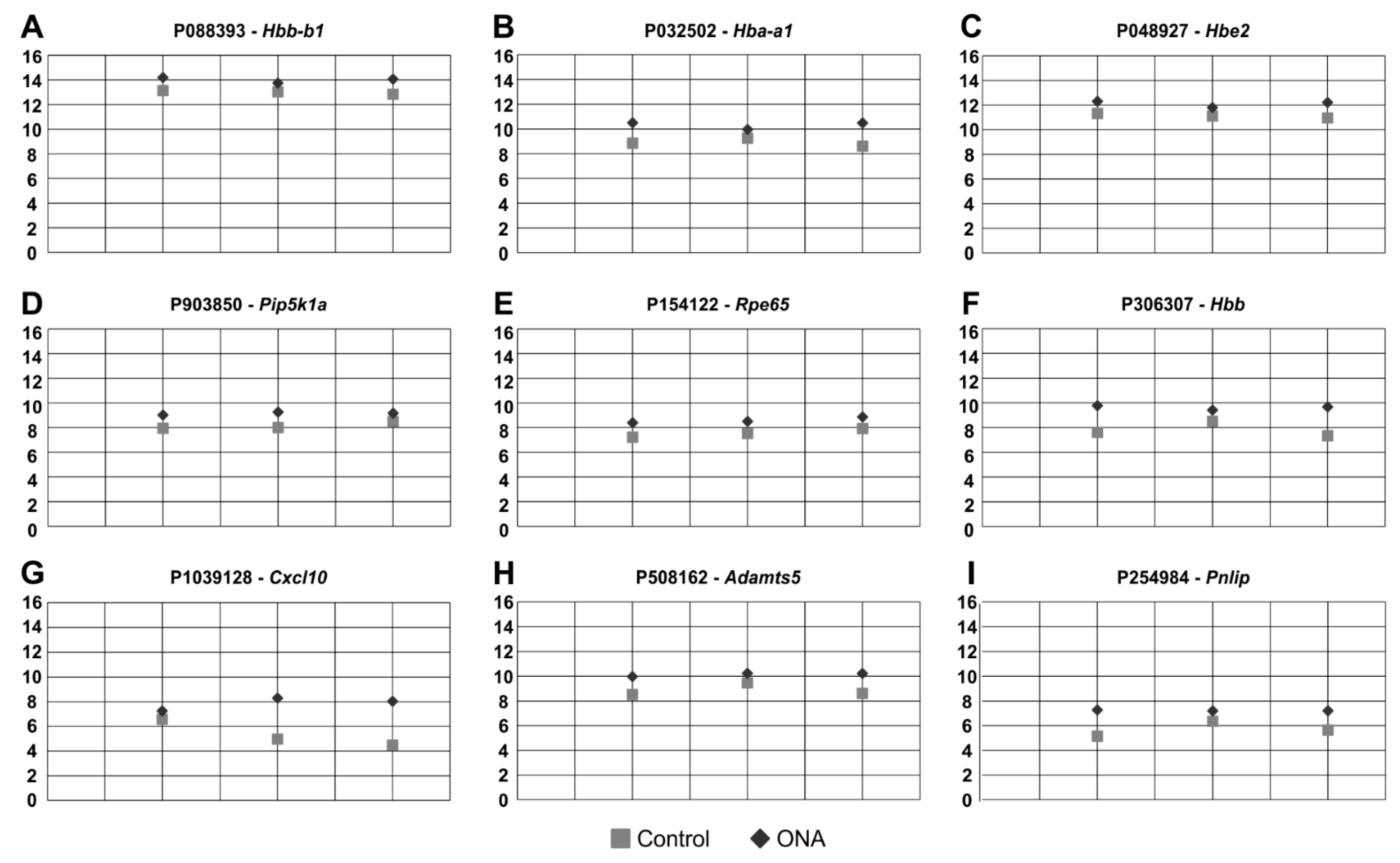
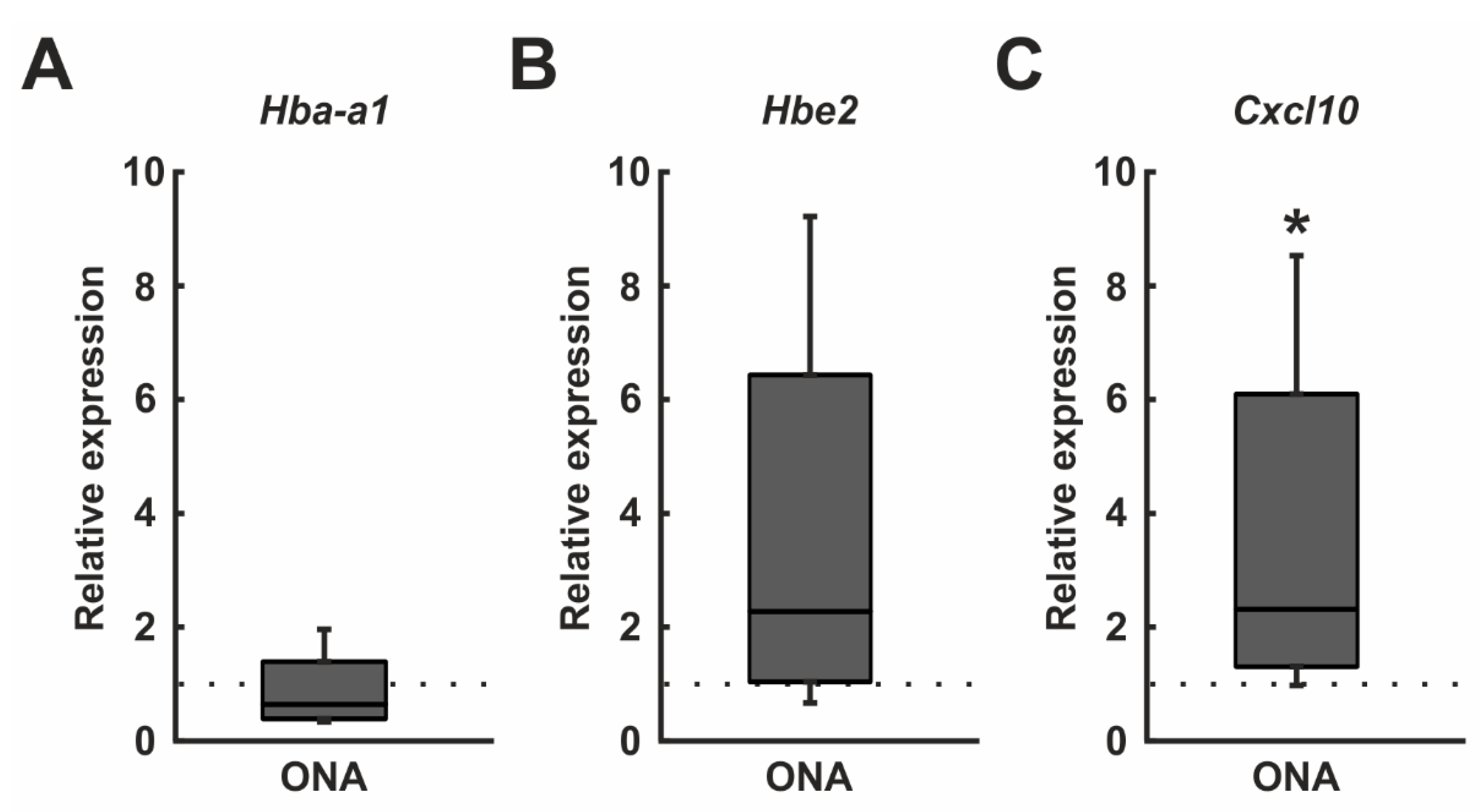
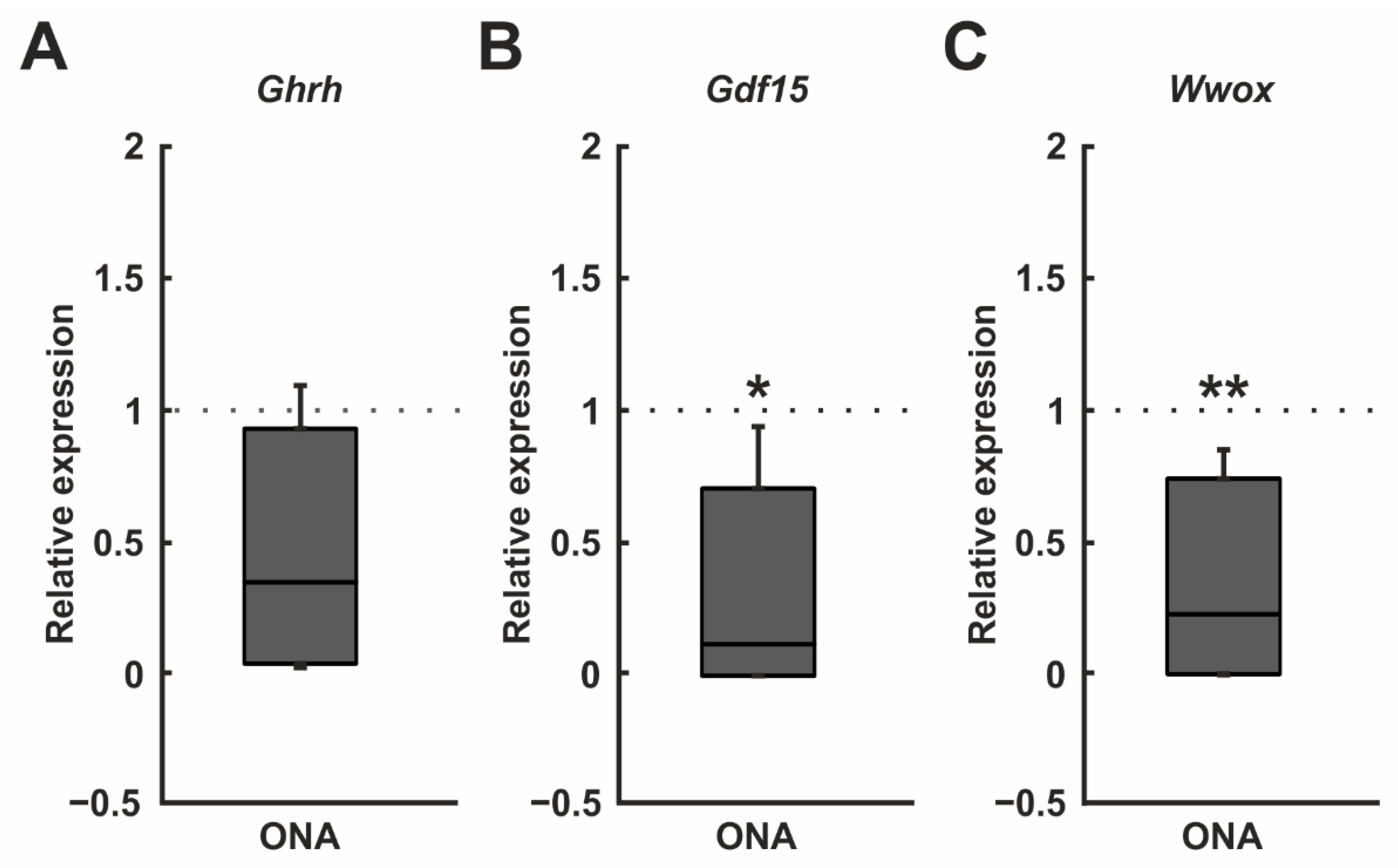
3.5. Regulated Genes in Microarray Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tham, Y.C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.Y. Global prevalence of glaucomaand projections of glaucoma burdenthrough 2040: A systematic review and meta-analysis. Ophthalmology 2014, 121, 2081–2090. [Google Scholar] [CrossRef]
- Burton, M.J.; Ramke, J.; Marques, A.P.; Bourne, R.R.A.; Congdon, N.; Jones, I.; Ah Tong, B.A.M.; Arunga, S.; Bachani, D.; Bascaran, C.; et al. The Lancet Global Health Commission on Global Eye Health: Vision beyond 2020. Lancet Global Health 2021, 9, e489–e551. [Google Scholar] [CrossRef]
- European Glaucoma Society Terminology and Guidelines for Glaucoma, 5th Edition. Br. J. Ophthalmol. 2021, 105, 1–169. [CrossRef]
- Sommer, A.; Tielsch, J.M.; Katz, J.; Quigley, H.A.; Gottsch, J.D.; Javitt, J.; Singh, K. Relationship between intraocular pressure and primary open angle glaucoma among white and black Americans. The Baltimore Eye Survey. Arch. Ophthalmol. 1991, 109, 1090–1095. [Google Scholar] [CrossRef] [PubMed]
- Klein, B.E.; Klein, R.; Sponsel, W.E.; Franke, T.; Cantor, L.B.; Martone, J.; Menage, M.J. Prevalence of glaucoma. The Beaver Dam Eye Study. Ophthalmology 1992, 99, 1499–1504. [Google Scholar] [CrossRef]
- Tezel, G.; Yang, X.; Luo, C.; Kain, A.D.; Powell, D.W.; Kuehn, M.H.; Kaplan, H.J. Oxidative stress and the regulation of complement activation in human glaucoma. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5071–5082. [Google Scholar] [CrossRef] [PubMed]
- Almasieh, M.; Wilson, A.M.; Morquette, B.; Cueva Vargas, J.L.; Di Polo, A. The molecular basis of retinal ganglion cell death in glaucoma. Prog. Retin. Eye Res. 2012, 31, 152–181. [Google Scholar] [CrossRef] [PubMed]
- Palmhof, M.; Frank, V.; Rappard, P.; Kortenhorn, E.; Demuth, J.; Biert, N.; Stute, G.; Dick, H.B.; Joachim, S.C. From Ganglion Cell to Photoreceptor Layer: Timeline of Deterioration in a Rat Ischemia/Reperfusion Model. Front. Cell Neurosci. 2019, 13, 174. [Google Scholar] [CrossRef] [Green Version]
- Kuehn, S.; Rodust, C.; Stute, G.; Grotegut, P.; Meissner, W.; Reinehr, S.; Dick, H.B.; Joachim, S.C. Concentration-Dependent Inner Retina Layer Damage and Optic Nerve Degeneration in a NMDA Model. J. Mol. Neurosci. 2017, 63, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Lambuk, L.; Jafri, A.J.A.; Iezhitsa, I.; Agarwal, R.; Bakar, N.S.; Agarwal, P.; Abdullah, A.; Ismail, N.M. Dose-dependent effects of NMDA on retinal and optic nerve morphology in rats. Int. J. Ophthalmol. 2019, 12, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Bell, K.; Und Hohenstein-Blaul, N.V.T.; Teister, J.; Grus, F. Modulation of the Immune System for the Treatment of Glaucoma. Curr. Neuropharmacol. 2018, 16, 942–958. [Google Scholar] [CrossRef] [PubMed]
- Grus, F.H.; Joachim, S.C.; Hoffmann, E.M.; Pfeiffer, N. Complex autoantibody repertoires in patients with glaucoma. Mol. Vis. 2004, 10, 132–137. [Google Scholar] [PubMed]
- Joachim, S.C.; Bruns, K.; Lackner, K.J.; Pfeiffer, N.; Grus, F.H. Antibodies to alpha B-crystallin, vimentin, and heat shock protein 70 in aqueous humor of patients with normal tension glaucoma and IgG antibody patterns against retinal antigen in aqueous humor. Curr. Eye Res. 2007, 32, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Wax, M.B.; Tezel, G.; Saito, I.; Gupta, R.S.; Harley, J.B.; Li, Z.; Romano, C. Anti-Ro/SS-A positivity and heat shock protein antibodies in patients with normal-pressure glaucoma. Am. J. Ophthalmol. 1998, 125, 145–157. [Google Scholar] [CrossRef]
- Tezel, G.; Hernandez, R.; Wax, M.B. Immunostaining of heat shock proteins in the retina and optic nerve head of normal and glaucomatous eyes. Arch. Ophthalmol. 2000, 118, 511–518. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, P. Roles of heat-shock proteins in innate and adaptive immunity. Nat. Rev. Immunol. 2002, 2, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Binder, R.J. Functions of heat shock proteins in pathways of the innate and adaptive immune system. J. Immunol. 2014, 193, 5765–5771. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Anders, F.; Funke, S.; Mercieca, K.; Grus, F.; Prokosch, V. Proteome alterations in aqueous humour of primary open angle glaucoma patients. Int. J. Ophthalmol. 2020, 13, 176–179. [Google Scholar] [CrossRef]
- Hubens, W.H.G.; Beckers, H.J.M.; Gorgels, T.; Webers, C.A.B. Increased ratios of complement factors C3a to C3 in aqueous humor and serum mark glaucoma progression. Exp. Eye Res. 2021, 204, 108460. [Google Scholar] [CrossRef] [PubMed]
- Boehm, N.; Beck, S.; Lossbrand, U.; Pfeiffer, N.; Grus, F.H. Analysis of Complement Proteins in Retina and Sera of Glaucoma Patients. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5221. [Google Scholar]
- Noristani, R.; Kuehn, S.; Stute, G.; Reinehr, S.; Stellbogen, M.; Dick, H.B.; Joachim, S.C. Retinal and Optic Nerve Damage is Associated with Early Glial Responses in an Experimental Autoimmune Glaucoma Model. J. Mol. Neurosci. 2016, 58, 470–482. [Google Scholar] [CrossRef] [PubMed]
- Reinehr, S.; Reinhard, J.; Wiemann, S.; Hesse, K.; Voss, C.; Gandej, M.; Dick, H.B.; Faissner, A.; Joachim, S.C. Transfer of the Experimental Autoimmune Glaucoma Model from Rats to Mice-New Options to Study Glaucoma Disease. Int. J. Mol. Sci. 2019, 20, 2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wax, M.B.; Tezel, G.; Yang, J.; Peng, G.; Patil, R.V.; Agarwal, N.; Sappington, R.M.; Calkins, D.J. Induced autoimmunity to heat shock proteins elicits glaucomatous loss of retinal ganglion cell neurons via activated T-cell-derived fas-ligand. J. Neurosci. 2008, 28, 12085–12096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laspas, P.; Gramlich, O.W.; Muller, H.D.; Cuny, C.S.; Gottschling, P.F.; Pfeiffer, N.; Dick, H.B.; Joachim, S.C.; Grus, F.H. Autoreactive antibodies and loss of retinal ganglion cells in rats induced by immunization with ocular antigens. Investig. Ophthalmol. Vis. Sci. 2011, 52, 8835–8848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinehr, S.; Reinhard, J.; Gandej, M.; Kuehn, S.; Noristani, R.; Faissner, A.; Burkhard Dick, H.; Joachim, S.C. Simultaneous Complement Response via Lectin Pathway in Retina and Optic Nerve in an Experimental Autoimmune Glaucoma Model. Front. Cell Neurosci. 2016, 10, 140. [Google Scholar] [CrossRef] [Green Version]
- Reinehr, S.; Reinhard, J.; Wiemann, S.; Stute, G.; Kuehn, S.; Woestmann, J.; Dick, H.B.; Faissner, A.; Joachim, S.C. Early remodelling of the extracellular matrix proteins tenascin-C and phosphacan in retina and optic nerve of an experimental autoimmune glaucoma model. J. Cell Mol. Med. 2016, 20, 2122–2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joachim, S.C.; Reinehr, S.; Kuehn, S.; Laspas, P.; Gramlich, O.W.; Kuehn, M.; Tischoff, I.; von Pein, H.D.; Dick, H.B.; Grus, F.H. Immune response against ocular tissues after immunization with optic nerve antigens in a model of autoimmune glaucoma. Mol. Vis. 2013, 19, 1804–1814. [Google Scholar] [PubMed]
- Reinehr, S.; Gomes, S.C.; Gassel, C.J.; Asaad, M.A.; Stute, G.; Schargus, M.; Dick, H.B.; Joachim, S.C. Intravitreal Therapy Against the Complement Factor C5 Prevents Retinal Degeneration in an Experimental Autoimmune Glaucoma Model. Front. Pharmacol. 2019, 10, 1381. [Google Scholar] [CrossRef] [Green Version]
- Benning, L.; Reinehr, S.; Grotegut, P.; Kuehn, S.; Stute, G.; Dick, H.B.; Joachim, S.C. Synapse and Receptor Alterations in Two Different S100B-Induced Glaucoma-Like Models. Int. J. Mol. Sci. 2020, 21, 6998. [Google Scholar] [CrossRef]
- Reinehr, S.; Kuehn, S.; Casola, C.; Koch, D.; Stute, G.; Grotegut, P.; Dick, H.B.; Joachim, S.C. HSP27 immunization reinforces AII amacrine cell and synapse damage induced by S100 in an autoimmune glaucoma model. Cell Tissue Res. 2018, 371, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Aring, L.; Steinbach, S.; Marcus, K.; May, C. Isolation of Distinct Types of Neurons from Fresh Brain Tissue Using Laser Microdissection in Combination with High-Performance Liquid Chromatography-Mass Spectrometry. Methods Mol. Biol. 2018, 1723, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Plum, S.; Steinbach, S.; Attems, J.; Keers, S.; Riederer, P.; Gerlach, M.; May, C.; Marcus, K. Proteomic characterization of neuromelanin granules isolated from human substantia nigra by laser-microdissection. Sci. Rep. 2016, 6, 37139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puscz, F.; Dadras, M.; Dermietzel, A.; Jacobsen, F.; Lehnhardt, M.; Behr, B.; Hirsch, T.; Kueckelhaus, M. A chronic rejection model and potential biomarkers for vascularized composite allotransplantation. PLoS ONE 2020, 15, e0235266. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, L.; Digiuni, M.; Montesano, G.; Centofanti, M.; Fea, A.M.; Iester, M.; Frezzotti, P.; Figus, M.; Ferreras, A.; Oddone, F.; et al. Blindness and Glaucoma: A Multicenter Data Review from 7 Academic Eye Clinics. PLoS ONE 2015, 10, e0136632. [Google Scholar] [CrossRef]
- Wolfram, C.; Schuster, A.K.; Elflein, H.M.; Nickels, S.; Schulz, A.; Wild, P.S.; Beutel, M.E.; Blettner, M.; Munzel, T.; Lackner, K.J.; et al. The Prevalence of Visual Impairment in the Adult Population. Dtsch. Arztebl Int. 2019, 116, 289–295. [Google Scholar] [CrossRef]
- Gupta, D.; Chen, P.P. Glaucoma. Am. Fam. Physician 2016, 93, 668–674. [Google Scholar]
- Reinehr, S.; Koch, D.; Weiss, M.; Froemel, F.; Voss, C.; Dick, H.B.; Fuchshofer, R.; Joachim, S.C. Loss of retinal ganglion cells in a new genetic mouse model for primary open-angle glaucoma. J. Cell Mol. Med. 2019, 23, 5497–5507. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.; Vecino, E. Role of Muller glia in neuroprotection and regeneration in the retina. Histol. Histopathol. 2003, 18, 1205–1218. [Google Scholar] [CrossRef]
- Bringmann, A.; Pannicke, T.; Grosche, J.; Francke, M.; Wiedemann, P.; Skatchkov, S.N.; Osborne, N.N.; Reichenbach, A. Muller cells in the healthy and diseased retina. Prog. Retin. Eye Res. 2006, 25, 397–424. [Google Scholar] [CrossRef]
- Galan, A.; Dergham, P.; Escoll, P.; de-la-Hera, A.; D’Onofrio, P.M.; Magharious, M.M.; Koeberle, P.D.; Frade, J.M.; Saragovi, H.U. Neuronal injury external to the retina rapidly activates retinal glia, followed by elevation of markers for cell cycle re-entry and death in retinal ganglion cells. PLoS ONE 2014, 9, e101349. [Google Scholar] [CrossRef]
- Casola, C.; Schiwek, J.E.; Reinehr, S.; Kuehn, S.; Grus, F.H.; Kramer, M.; Dick, H.B.; Joachim, S.C. S100 Alone Has the Same Destructive Effect on Retinal Ganglion Cells as in Combination with HSP 27 in an Autoimmune Glaucoma Model. J. Mol. Neurosci. 2015, 56, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, S.; Craig, E.A. The heat-shock proteins. Annu. Rev. Genet. 1988, 22, 631–677. [Google Scholar] [CrossRef]
- Michel, G.P.; Starka, J. Effect of ethanol and heat stresses on the protein pattern of Zymomonas mobilis. J. Bacteriol. 1986, 165, 1040–1042. [Google Scholar] [CrossRef] [Green Version]
- Kampinga, H.H.; Hageman, J.; Vos, M.J.; Kubota, H.; Tanguay, R.M.; Bruford, E.A.; Cheetham, M.E.; Chen, B.; Hightower, L.E. Guidelines for the nomenclature of the human heat shock proteins. Cell Stress Chaperones 2009, 14, 105–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kampinga, H.H.; Craig, E.A. The HSP70 chaperone machinery: J proteins as drivers of functional specificity. Nat. Rev. Mol. Cell Biol. 2010, 11, 579–592. [Google Scholar] [CrossRef] [Green Version]
- Jakob, U.; Gaestel, M.; Engel, K.; Buchner, J. Small heat shock proteins are molecular chaperones. J. Biol. Chem. 1993, 268, 1517–1520. [Google Scholar] [CrossRef]
- Bruey, J.M.; Ducasse, C.; Bonniaud, P.; Ravagnan, L.; Susin, S.A.; Diaz-Latoud, C.; Gurbuxani, S.; Arrigo, A.P.; Kroemer, G.; Solary, E.; et al. Hsp27 negatively regulates cell death by interacting with cytochrome c. Nat. Cell Biol. 2000, 2, 645–652. [Google Scholar] [CrossRef]
- Jin, C.; Cleveland, J.C.; Ao, L.; Li, J.; Zeng, Q.; Fullerton, D.A.; Meng, X. Human myocardium releases heat shock protein 27 (HSP27) after global ischemia: The proinflammatory effect of extracellular HSP27 through toll-like receptor (TLR)-2 and TLR4. Mol. Med. 2014, 20, 280–289. [Google Scholar] [CrossRef]
- Tsai, T.; Grotegut, P.; Reinehr, S.; Joachim, S.C. Role of Heat Shock Proteins in Glaucoma. Int. J. Mol. Sci. 2019, 20, 5160. [Google Scholar] [CrossRef] [Green Version]
- Wax, M.B.; Tezel, G.; Edward, P.D. Clinical and ocular histopathological findings in a patient with normal-pressure glaucoma. Arch. Ophthalmol. 1998, 116, 993–1001. [Google Scholar] [CrossRef] [Green Version]
- Tezel, G.; Seigel, G.M.; Wax, M.B. Autoantibodies to small heat shock proteins in glaucoma. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2277–2287. [Google Scholar]
- Joachim, S.C.; Grus, F.H.; Kraft, D.; White-Farrar, K.; Barnes, G.; Barbeck, M.; Ghanaati, S.; Cao, S.; Li, B.; Wax, M.B. Complex antibody profile changes in an experimental autoimmune glaucoma animal model. Investig. Ophthalmol. Vis. Sci. 2009, 50, 4734–4742. [Google Scholar] [CrossRef]
- Grotegut, P.; Kuehn, S.; Dick, H.B.; Joachim, S.C. Destructive Effect of Intravitreal Heat Shock Protein 27 Application on Retinal Ganglion Cells and Neurofilament. Int. J. Mol. Sci. 2020, 21, 549. [Google Scholar] [CrossRef] [Green Version]
- Boehm, N.; Wolters, D.; Thiel, U.; Lossbrand, U.; Wiegel, N.; Pfeiffer, N.; Grus, F.H. New insights into autoantibody profiles from immune privileged sites in the eye: A glaucoma study. Brain Behav Immun. 2012, 26, 96–102. [Google Scholar] [CrossRef]
- Prohaszka, Z.; Singh, M.; Nagy, K.; Kiss, E.; Lakos, G.; Duba, J.; Fust, G. Heat shock protein 70 is a potent activator of the human complement system. Cell Stress Chaperones 2002, 7, 17–22. [Google Scholar] [CrossRef]
- Reinehr, S.; Reinhard, J.; Gandej, M.; Gottschalk, I.; Stute, G.; Faissner, A.; Dick, H.B.; Joachim, S.C. S100B immunization triggers NFκB and complement activation in an autoimmune glaucoma model. Sci. Rep. 2018, 8, 9821. [Google Scholar] [CrossRef] [Green Version]
- Prakken, B.J.; Samodal, R.; Le, T.D.; Giannoni, F.; Yung, G.P.; Scavulli, J.; Amox, D.; Roord, S.; de Kleer, I.; Bonnin, D.; et al. Epitope-specific immunotherapy induces immune deviation of proinflammatory T cells in rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 2004, 101, 4228–4233. [Google Scholar] [CrossRef] [Green Version]
- Reinehr, S.; Doerner, J.D.; Mueller-Buehl, A.M.; Koch, D.; Fuchshofer, R.; Dick, H.B.; Joachim, S.C. Cytokine and Complement Response in the Glaucomatous betaB1-CTGF Mouse Model. Front. Cell Neurosci. 2021, 15, 718087. [Google Scholar] [CrossRef]
- Kuehn, M.H.; Kim, C.Y.; Ostojic, J.; Bellin, M.; Alward, W.L.; Stone, E.M.; Sakaguchi, D.S.; Grozdanic, S.D.; Kwon, Y.H. Retinal synthesis and deposition of complement components induced by ocular hypertension. Exp. Eye Res. 2006, 83, 620–628. [Google Scholar] [CrossRef]
- Krishnan, A.; Kocab, A.J.; Zacks, D.N.; Marshak-Rothstein, A.; Gregory-Ksander, M. A small peptide antagonist of the Fas receptor inhibits neuroinflammation and prevents axon degeneration and retinal ganglion cell death in an inducible mouse model of glaucoma. J. Neuroinflamm. 2019, 16, 184. [Google Scholar] [CrossRef] [Green Version]
- Markiewski, M.M.; Lambris, J.D. The role of complement in inflammatory diseases from behind the scenes into the spotlight. Am. J. Pathol. 2007, 171, 715–727. [Google Scholar] [CrossRef] [Green Version]
- Matarazzo, L.; Hernandez Santana, Y.E.; Walsh, P.T.; Fallon, P.G. The IL-1 cytokine family as custodians of barrier immunity. Cytokine 2022, 154, 155890. [Google Scholar] [CrossRef]
- Levkovitch-Verbin, H.; Waserzoog, Y.; Vander, S.; Makarovsky, D.; Piven, I. Minocycline upregulates pro-survival genes and downregulates pro-apoptotic genes in experimental glaucoma. Graefes Arch. Clin Exp. Ophthalmol. 2014, 252, 761–772. [Google Scholar] [CrossRef]
- Zhou, X.; Li, F.; Kong, L.; Chodosh, J.; Cao, W. Anti-inflammatory effect of pigment epithelium-derived factor in DBA/2J mice. Mol. Vis. 2009, 15, 438–450. [Google Scholar]
- Liew, F.Y.; Girard, J.P.; Turnquist, H.R. Interleukin-33 in health and disease. Nat. Rev. Immunol. 2016, 16, 676–689. [Google Scholar] [CrossRef]
- Barbour, M.; Allan, D.; Xu, H.; Pei, C.; Chen, M.; Niedbala, W.; Fukada, S.Y.; Besnard, A.G.; Alves-Filho, J.C.; Tong, X.; et al. IL-33 attenuates the development of experimental autoimmune uveitis. Eur. J. Immunol. 2014, 44, 3320–3329. [Google Scholar] [CrossRef] [Green Version]
- Scott, L.M.; Vincent, E.E.; Hudson, N.; Neal, C.; Jones, N.; Lavelle, E.C.; Campbell, M.; Halestrap, A.P.; Dick, A.D.; Theodoropoulou, S. Interleukin-33 regulates metabolic reprogramming of the retinal pigment epithelium in response to immune stressors. JCI insight 2021, 6, e129429. [Google Scholar] [CrossRef]
- Blank, T.; Goldmann, T.; Koch, M.; Amann, L.; Schon, C.; Bonin, M.; Pang, S.; Prinz, M.; Burnet, M.; Wagner, J.E.; et al. Early Microglia Activation Precedes Photoreceptor Degeneration in a Mouse Model of CNGB1-Linked Retinitis Pigmentosa. Front. Immunol. 2017, 8, 1930. [Google Scholar] [CrossRef]
- Lacotte, S.; Brun, S.; Muller, S.; Dumortier, H. CXCR3, inflammation, and autoimmune diseases. Ann. N. Y. Acad. Sci. 2009, 1173, 310–317. [Google Scholar] [CrossRef]
- Ha, Y.; Liu, H.; Zhu, S.; Yi, P.; Liu, W.; Nathanson, J.; Kayed, R.; Loucas, B.; Sun, J.; Frishman, L.J.; et al. Critical Role of the CXCL10/C-X-C Chemokine Receptor 3 Axis in Promoting Leukocyte Recruitment and Neuronal Injury during Traumatic Optic Neuropathy Induced by Optic Nerve Crush. Am. J. Pathol. 2017, 187, 352–365. [Google Scholar] [CrossRef] [Green Version]
- Ban, N.; Siegfried, C.J.; Lin, J.B.; Shui, Y.B.; Sein, J.; Pita-Thomas, W.; Sene, A.; Santeford, A.; Gordon, M.; Lamb, R.; et al. GDF15 is elevated in mice following retinal ganglion cell death and in glaucoma patients. JCI Insight 2017, 2, e91455. [Google Scholar] [CrossRef] [Green Version]
- Iwata, Y.; Inagaki, S.; Morozumi, W.; Nakamura, S.; Hara, H.; Shimazawa, M. Treatment with GDF15, a TGFbeta superfamily protein, induces protective effect on retinal ganglion cells. Exp. Eye Res. 2021, 202, 108338. [Google Scholar] [CrossRef]
- Cheng, Y.Y.; Chou, Y.T.; Lai, F.J.; Jan, M.S.; Chang, T.H.; Jou, I.M.; Chen, P.S.; Lo, J.Y.; Huang, S.S.; Chang, N.S.; et al. Wwox deficiency leads to neurodevelopmental and degenerative neuropathies and glycogen synthase kinase 3beta-mediated epileptic seizure activity in mice. Acta Neuropathol. Commun. 2020, 8, 6. [Google Scholar] [CrossRef]
- Iacomino, M.; Baldassari, S.; Tochigi, Y.; Kosla, K.; Buffelli, F.; Torella, A.; Severino, M.; Paladini, D.; Mandara, L.; Riva, A.; et al. Loss of Wwox Perturbs Neuronal Migration and Impairs Early Cortical Development. Front. Neurosci. 2020, 14, 644. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (F) and Reverse (R) Oligonucleotides | GenBank Accession Number | Amplicon Size |
|---|---|---|---|
| Actb-F Actb-R | cccgcgagtacaaccttct cgtcatccatggcgaact | NM_031144.3 | 72 bp |
| C1qa-F C1qa-R | cgggtctcaaaggagagagag ccagattcccccatgtctc | XM_032888144.1 | 88 bp |
| C1qb-F C1qb-R | gcactccagggataaaagga ccctttctctcctaactcacca | NM_019262.2 | 74 bp |
| Cxcl10-F Cxcl10-R | caagtgctgctgtcgttctc atctcaacatgcggacagga | NM_139089.2 | 178 bp |
| Dnajb1-F Dnajb1-R | attttcgaccgctatggaga cattagcaccaccactgctc | NM_001108441.1 | 73 bp |
| Gdf15-F Gdf15-R | tcagctgaggttcctgctgttc gctcgtccgggttgagttg | NM_019216.2 | 128 bp |
| Gfap-F Gfap-R | tttctccaacctccagatcc gaggtggccttctgacacag | NM_017009.2 | 64 bp |
| Ghrh-F Ghrh-R | tttgtgctcctcaccctcac aattggcccaggattctccg | NM_031577.1 | 122 bp |
| Hba-a1-F Hba-a1-R | ccctggagatttcacacccg aacggtacttggaggtcagc | NM_013096.2 | 86 bp |
| Hbe2-F Hbe2-R | cggccatcatgggtaatccc agaagaaagaacaatcaccagcac | NM_001024805.1 | 194 bp |
| Hsp90aa1-F Hsp90aa1-R | gggagctcatttccaactcc gggttcggtcttgcttgtt | NM_175761.2 | 129 bp |
| Hspa4-F Hspa4-R | catatccaatatctttgaggtgga tggggaagacttcacagtca | NM_153629.1 | 69 bp |
| Hspb1-F Hspb1-R | gaggagctcacagttaagaccaa ttcatcctgcctttcttcg | NM_031970.4 | 72 bp |
| Il11-F Il11-R | gctggtccttccctaaagactc aaggctaggcgagacatcaa | NM_133519.4 | 104 bp |
| Il18-F Il18-R | gcctgatatcgaccgaaca ccttccatccttcacagatagg | NM_019165.1 | 112 bp |
| Il33-F Il33-R | gcaaagtgcgacagcaca ctttggtcttctgttgggatct | NM_001014166.1 | 76 bp |
| Nfkb1-F Nfkb1-R | ctggcagctcttctcaaagc ccaggtcatagagaggctcaa | NM_001276711.1 | 70 bp |
| Pou4f1-F Pou4f1-R | ctggccaacctcaagatcc cgtgagcgactcgaacct | XM_008770931.2 | 72 bp |
| Ppid-F Ppid-R | tgctggaccaaacacaaatg cttcccaaagaccacatgct | M19553.1 | 88 bp |
| Tubb3-F Tubb3-R | tccagctcactcactcactg ggtctcatccgtgttctcca | NM_139254.2 | 200 bp |
| Wwox-F Wwox-R | cagagatacgacgggagcac gctccagtaaccaggaccac | NM_001106188.1 | 83 bp |
| Primary Antibodies | Secondary Antibodies | ||||
|---|---|---|---|---|---|
| Antibody | Company | Dilution | Antibody | Company | Dilution |
| Anti-C1q | Abcam | 1:400 | Donkey anti-rabbit Alexa Fluor 488 | Jackson Immuno Research | 1:500 |
| Anti-GFAP | Millipore | 1:2000 | Donkey anti-chicken Alexa Fluor 488 | Jackson Immuno Research | 1:500 |
| Anti-HSP25 | Enzo Life Science | 1:100 | Goat anti-rabbit Alexa Fluor 488 | Invitrogen | 1:500 |
| Anti-NFκB | Santa Cruz | 1:300 | Donkey anti-mouse Alexa Flour 555 | Abcam | 1:500 |
| Anti-RBPMS | Millipore | 1:500 | Donkey anti-rabbit Alexa Fluor 555 | Invitrogen | 1:500 |
| Probe Name | Gene Symbol | Description | p-Value | Fold Change ONA vs. Control |
|---|---|---|---|---|
| A_64_P088393 | Hbb-b1 | Rattus norvegicus hemoglobin, beta adult major chain (Hbb-b1), mRNA [NM_198776] | 0.006 | 1.077 (↑) |
| A_64_P032502 | Hba-a1 | Rattus norvegicus hemoglobin, alpha 1 (Hba1), mRNA [NM_013096.2] | 0.006 | 1.159 (↑) |
| A_64_P048927 | Hbe2 | Rattus norvegicus hemoglobin, epsilon 2 (Hbe2), mRNA [NM_001024805] | 0.009 | 1.088 (↑) |
| A_44_P903850 | Pip5k1a | Rattus norvegicus phosphatidylinositol-4-phosphate 5-kinase, type 1, alpha (Pip5k1a), mRNA [NM_001042621] | 0.017 | 1.123 (↑) |
| A_64_P154122 | Rpe65 | Rattus norvegicus RPE65, retinoid isomerohydrolase (Rpe65), mRNA [NM_053562] | 0.018 | 1.136 (↑) |
| A_44_P306307 | Hbb | Rattus norvegicus hemoglobin subunit beta (Hbb), mRNA [NM_033234] | 0.027 | 1.231 (↑) |
| A_44_P1039128 | Cxcl10 | Rattus norvegicus C-X-C motif chemokine ligand 10 (Cxcl10), mRNA [NM_139089] | 0.039 | 1.470 (↑) |
| A_44_P508162 | Adamts5 | Rattus norvegicus ADAM metallopeptidase with thrombospondin type 1 motif, 5 (Adamts5), mRNA [NM_198761] | 0.042 | 1.144 (↑) |
| A_44_P254984 | Pnlip | Rattus norvegicus pancreatic lipase (Pnlip), mRNA [NM_013161] | 0.049 | 1.264 (↑) |
| Probe Name | Gene Symbol | Description | p-Value | Fold Change ONA vs. Control |
|---|---|---|---|---|
| A_64_P062965 | Ghrh | Rattus norvegicus growth hormone releasing hormone (Ghrh), mRNA [NM_031577] | 0.002 | 0.866 (↓) |
| A_64_P011489 | Gdf15 | Rattus norvegicus growth differentiation factor 15 (Gdf15), mRNA [NM_019216] | 0.011 | 0.849 (↓) |
| A_64_P128359 | Marf1 | meiosis arrest female protein 1-like [Source:RGD Symbol;Acc:9219913] [ENSRNOT00000003193] | 0.019 | 0.912 (↓) |
| A_64_P112599 | Marf1 | Rattus norvegicus meiosis arrest female 1 (Marf1), mRNA [NM_133421] | 0.020 | 0.912 (↓) |
| A_44_P727274 | Chtf8 | Rattus norvegicus chromosome transmission fidelity factor 8 (Chtf8), mRNA [NM_001194951] | 0.021 | 0.877 (↓) |
| A_64_P124893 | Kctd16 | Rattus norvegicus potassium channel tetramerization domain containing 16 (Kctd16), mRNA [NM_001172155] | 0.022 | 0.890 (↓) |
| A_64_P067366 | Cacna1h | Rattus norvegicus calcium voltage-gated channel subunit alpha1 H (Cacna1h), mRNA [NM_153814] | 0.028 | 0.900 (↓) |
| A_64_P028011 | Wwox | Rattus norvegicus WW domain-containing oxidoreductase (Wwox), mRNA [NM_001106188] | 0.030 | 0.864 (↓) |
| A_64_P105486 | Tafa2 | PREDICTED: Rattus rattus TAFA chemokine like family member 2 (Tafa2), transcript variant X2, mRNA [XM_032913314.1] | 0.036 | 0.826 (↓) |
| A_64_P119502 | Pikfyve | phosphoinositide kinase, FYVE-type zinc finger containing [Source:RGD Symbol;Acc:1592067] [ENSRNOT00000020447] | 0.040 | 0.816 (↓) |
| A_64_P089410 | Complete sequence | Rattus norvegicus Y Chr BAC RNAEX-239M11 (Amplicon Express Rat SHR-Akr BAC library) complete sequence [AC244815.7] | 0.043 | 0.881 (↓) |
| A_64_P115182 | LOC690617 | Rattus norvegicus hypothetical protein LOC690617 (LOC690617), mRNA [NM_001109605] | 0.044 | 0.909 (↓) |
| A_64_P074316 | Complete sequence | Rattus norvegicus, 25 clones, strain BN/SsNHsdMCW RNOR03324481, whole genome shotgun sequence [AABR03128056] | 0.044 | 0.885 (↓) |
| A_64_P017771 | Galntl6 | Rattus norvegicus polypeptide N-acetylgalactosaminyltransferase-like 6 (Galntl6), mRNA [NM_001135756] | 0.046 | 0.851 (↓) |
| A_44_P177807 | Epas | Rattus norvegicus endothelial PAS domain protein 1 (Epas1), mRNA [NM_023090] | 0.047 | 0.844 (↓) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reinehr, S.; Safaei, A.; Grotegut, P.; Guntermann, A.; Tsai, T.; Hahn, S.A.; Kösters, S.; Theiss, C.; Marcus, K.; Dick, H.B.; et al. Heat Shock Protein Upregulation Supplemental to Complex mRNA Alterations in Autoimmune Glaucoma. Biomolecules 2022, 12, 1538. https://doi.org/10.3390/biom12101538
Reinehr S, Safaei A, Grotegut P, Guntermann A, Tsai T, Hahn SA, Kösters S, Theiss C, Marcus K, Dick HB, et al. Heat Shock Protein Upregulation Supplemental to Complex mRNA Alterations in Autoimmune Glaucoma. Biomolecules. 2022; 12(10):1538. https://doi.org/10.3390/biom12101538
Chicago/Turabian StyleReinehr, Sabrina, Armin Safaei, Pia Grotegut, Annika Guntermann, Teresa Tsai, Stephan A. Hahn, Steffen Kösters, Carsten Theiss, Katrin Marcus, H. Burkhard Dick, and et al. 2022. "Heat Shock Protein Upregulation Supplemental to Complex mRNA Alterations in Autoimmune Glaucoma" Biomolecules 12, no. 10: 1538. https://doi.org/10.3390/biom12101538