
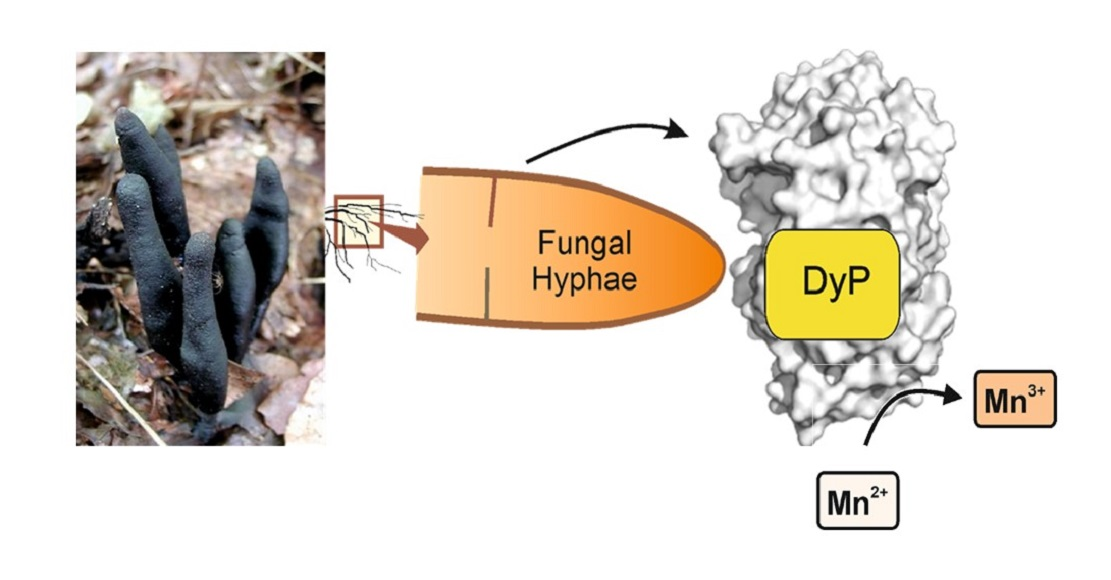
First Dye-Decolorizing Peroxidase from an Ascomycetous Fungus Secreted by Xylaria grammica

 , ,
, ,  , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
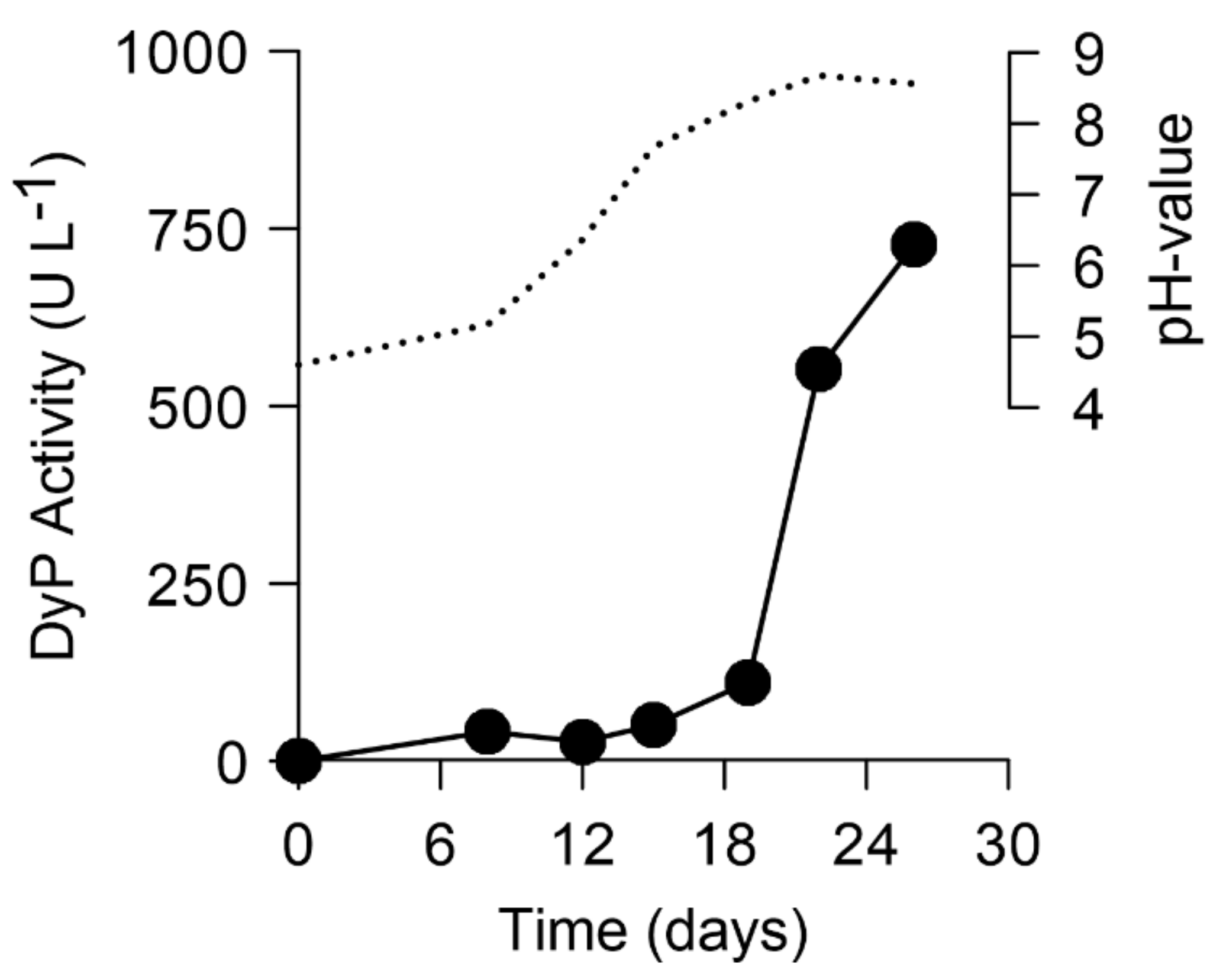
2.1. Screening and Production of a DyP-Type Peroxidase by X. grammica
2.2. Enzyme Assays
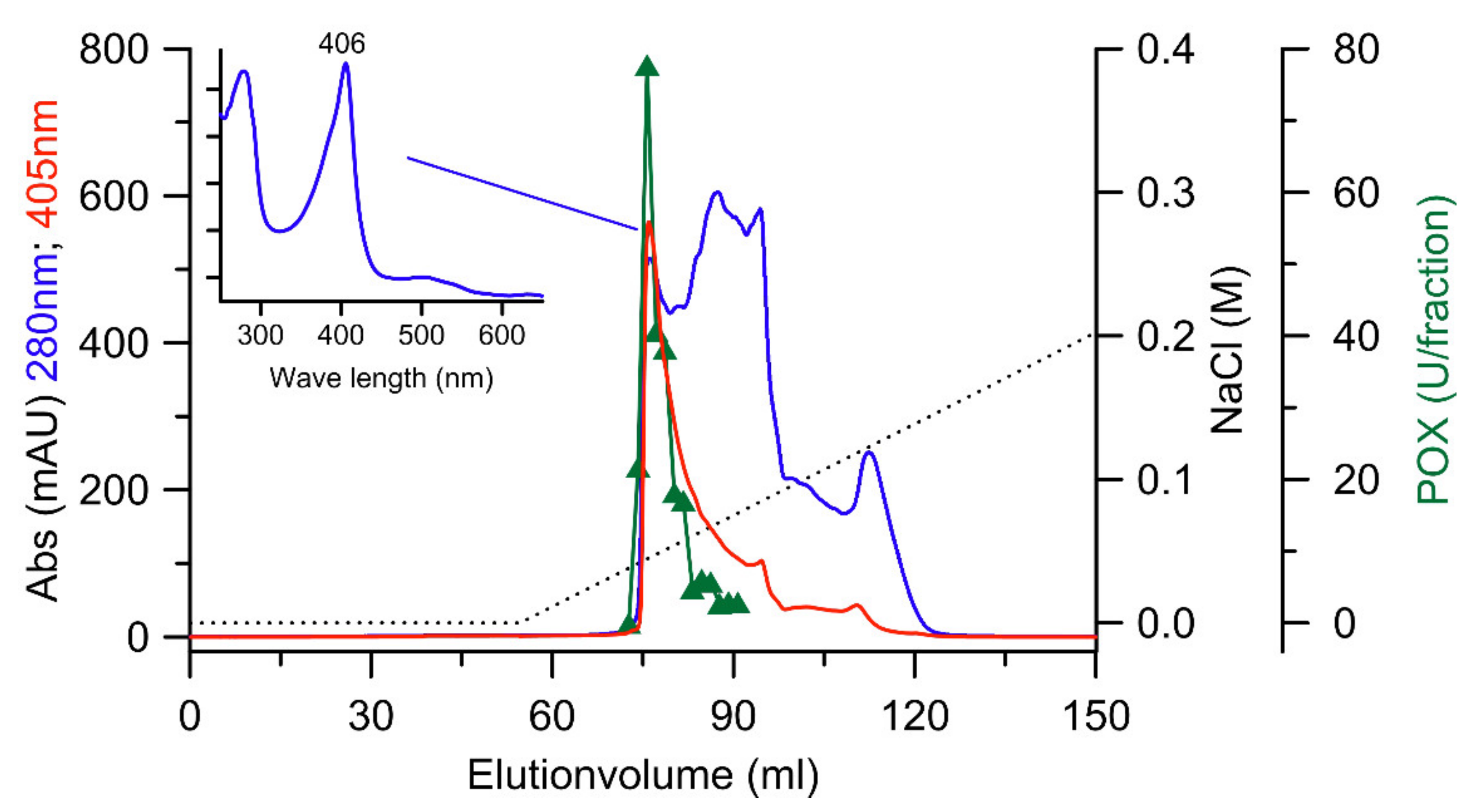
2.3. Purification of the DyP from X. grammica (XgrDyP)
2.4. Protein Determination
2.5. Protein Electrophoresis
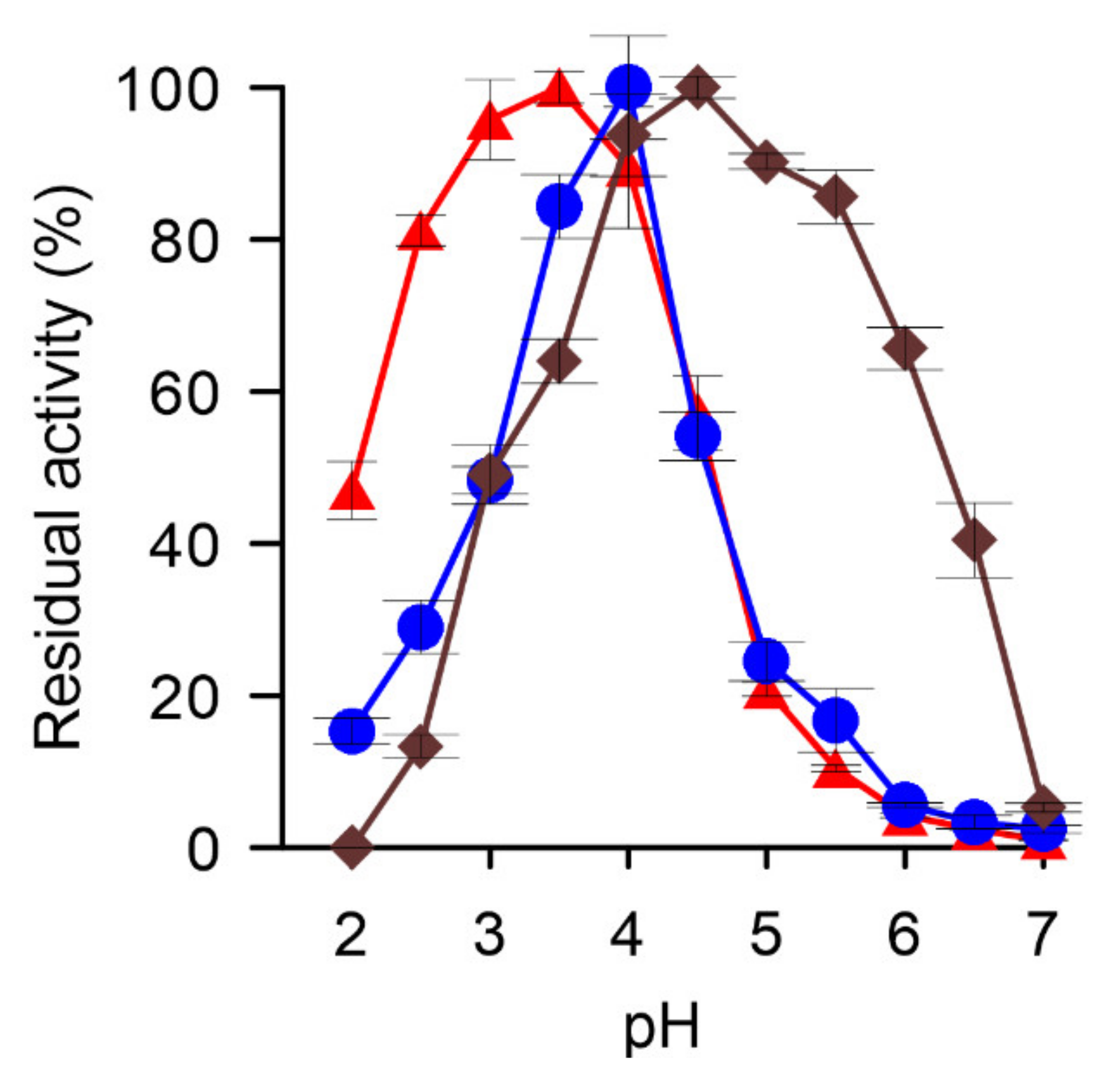
2.6. Determination of pH-Optima and Stability
2.7. Kinetic Parameters
2.8. Peptide Sequencing
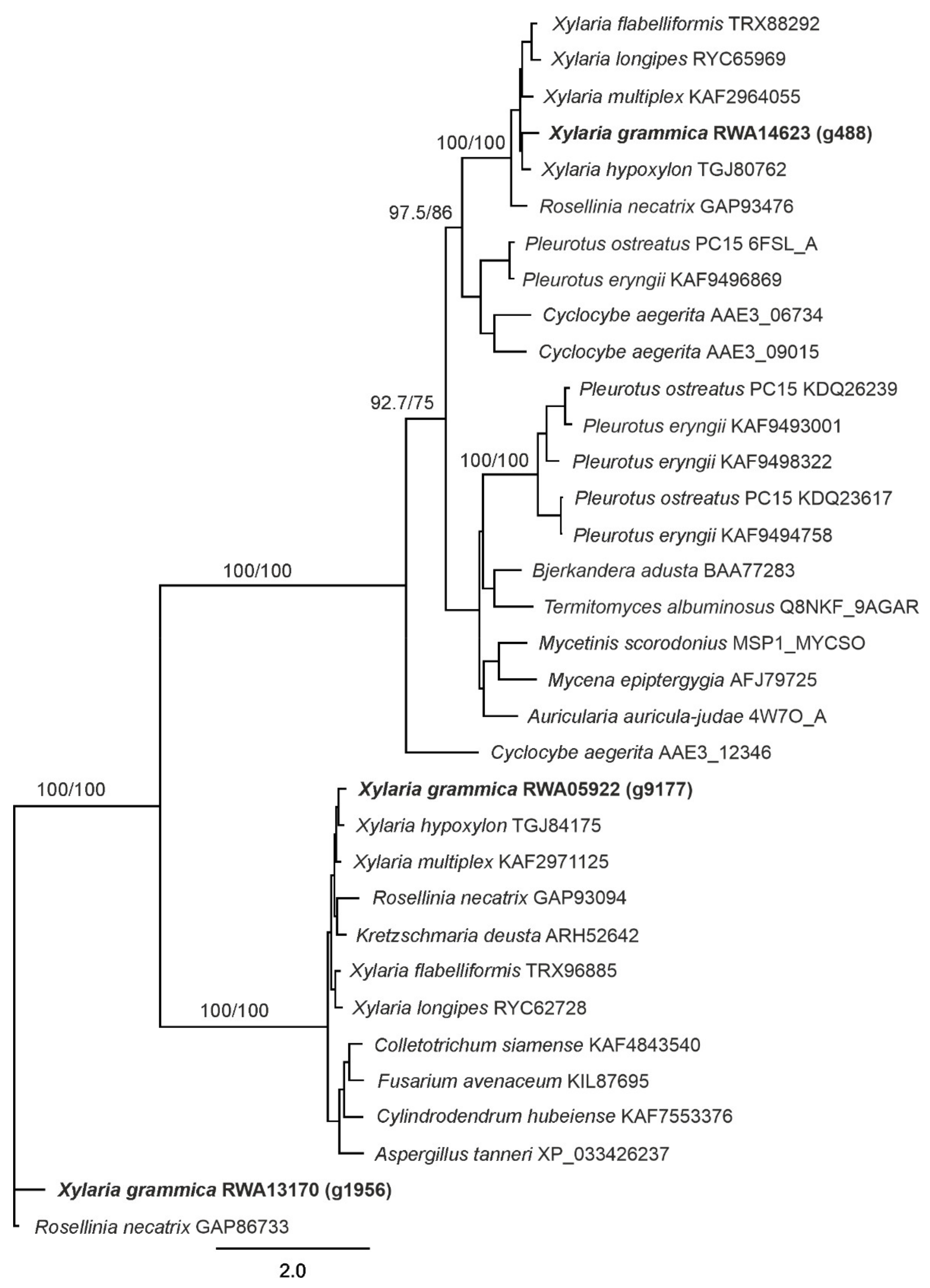
2.9. Phylogenetic Analysis
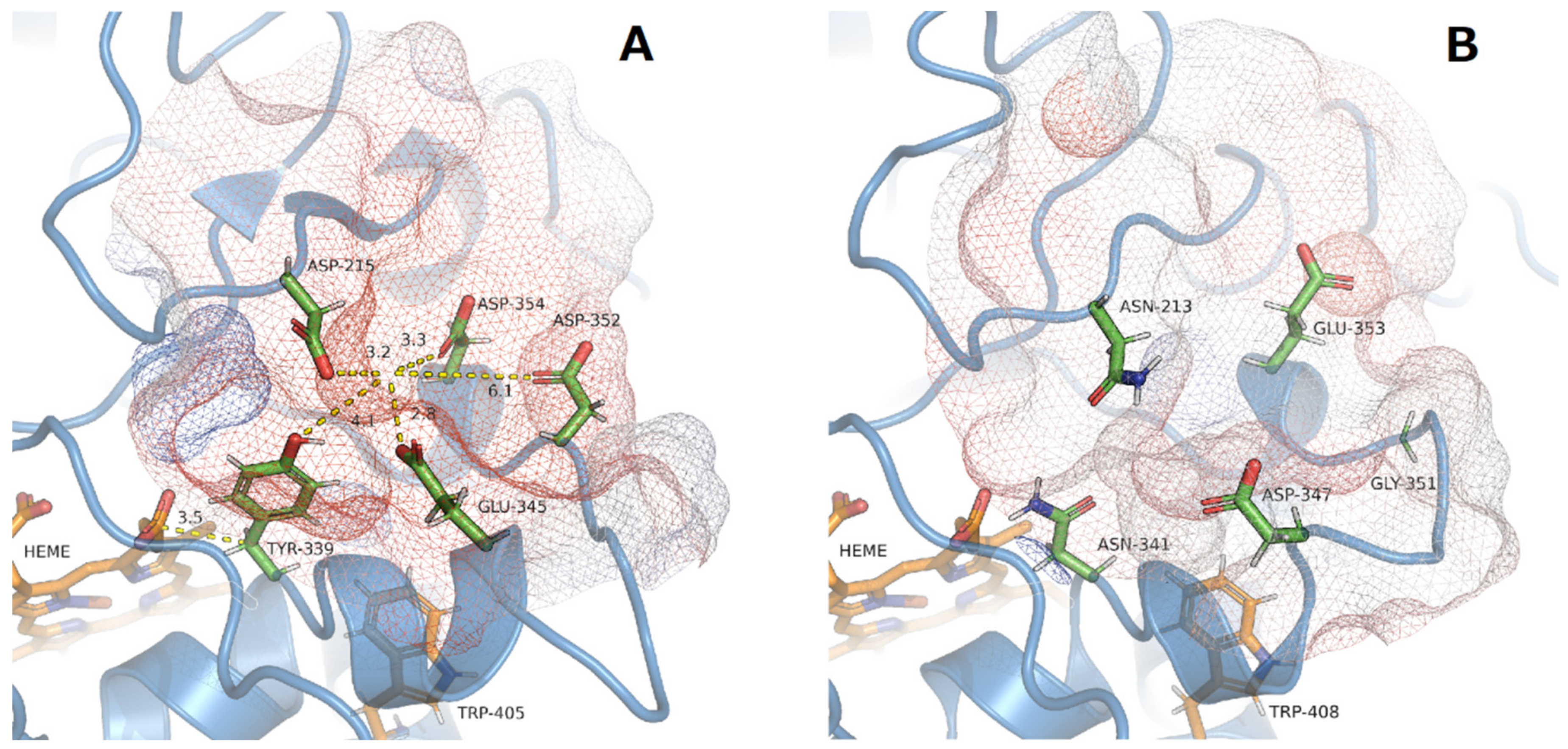
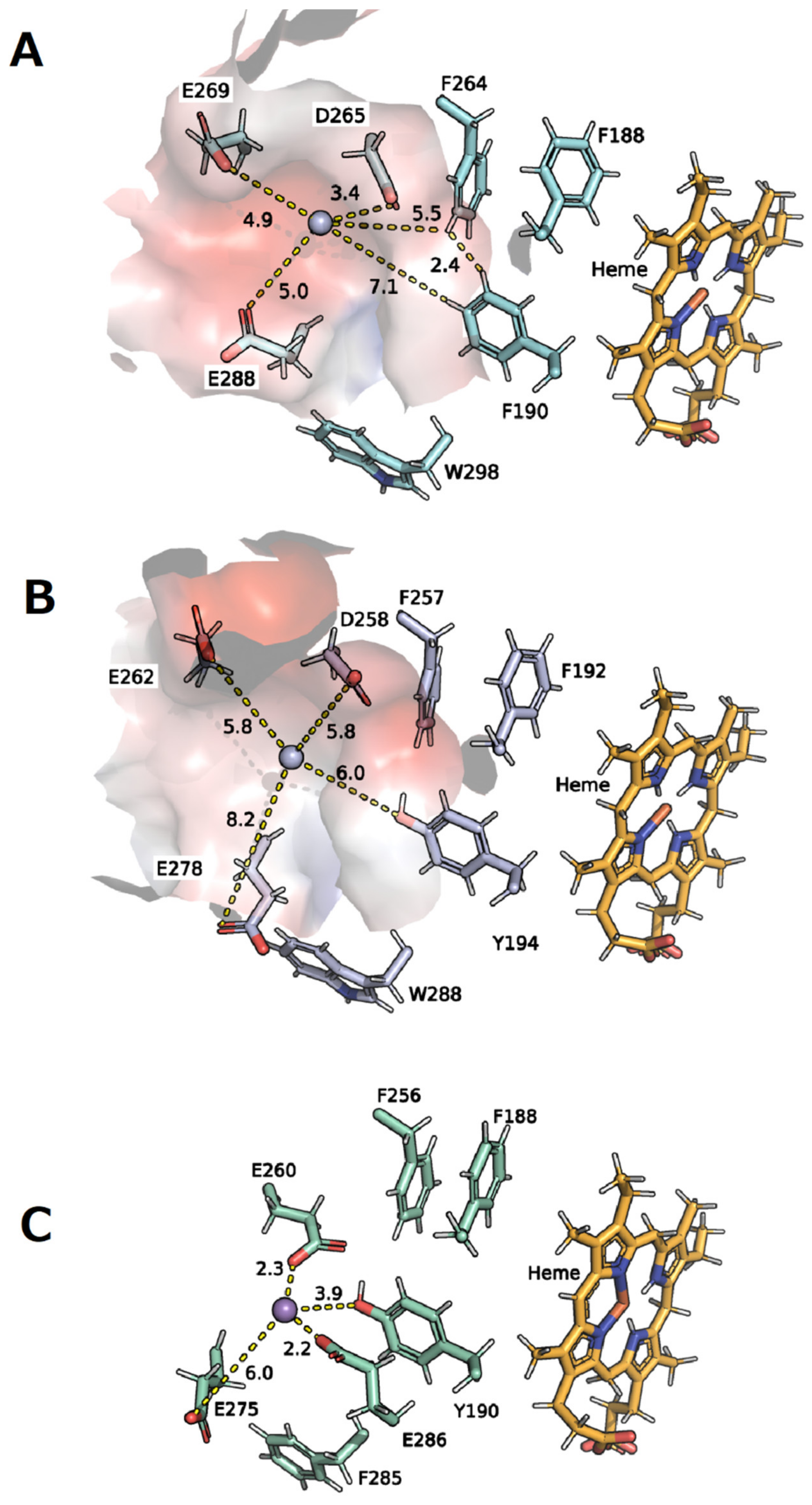
2.10. Homology Modeling
3. Results
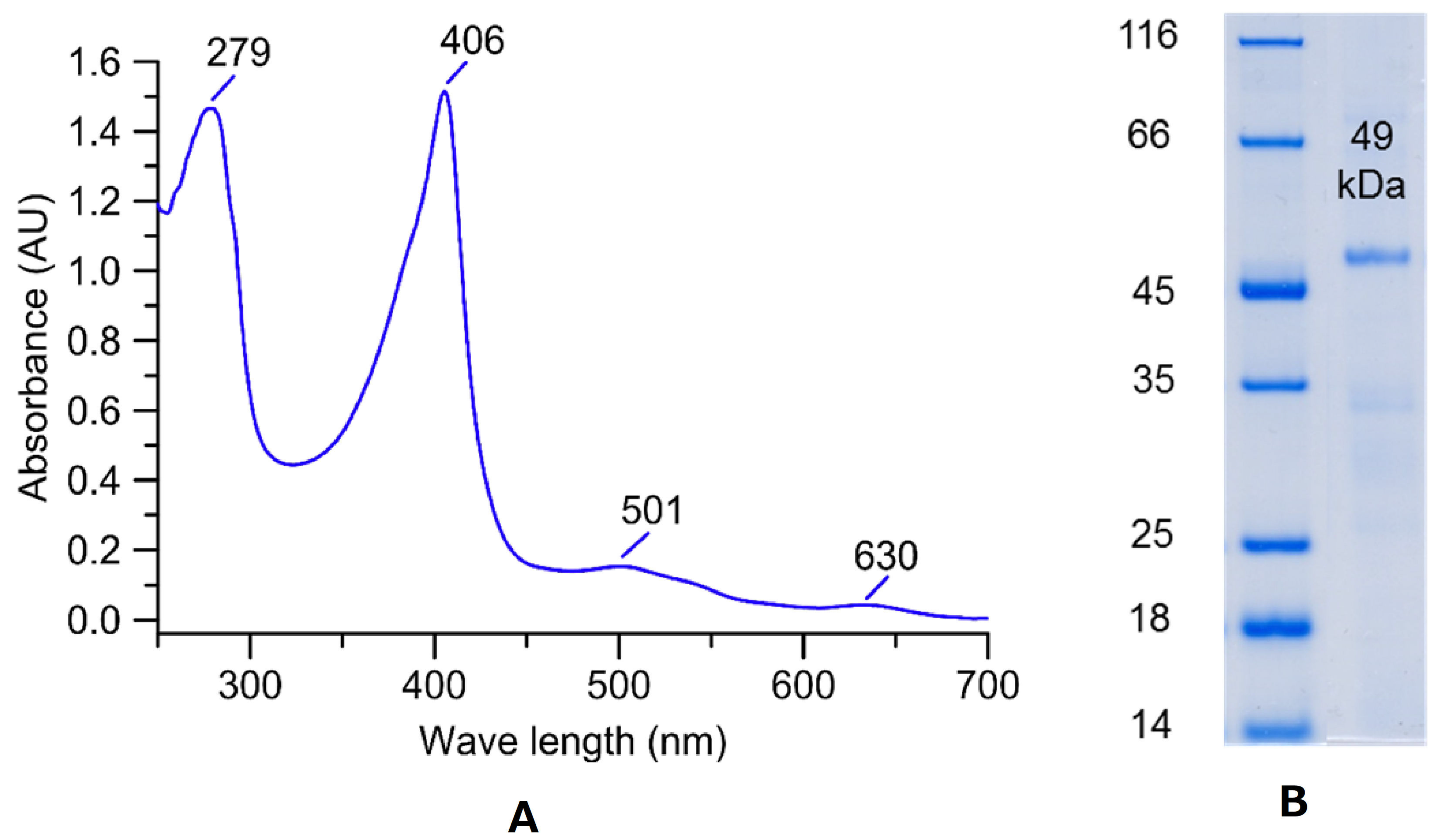
3.1. Production and Purification of a DyP from X. grammica
3.2. Characterization of the X. grammica DyP
3.3. Kinetic Parameters of the Purified X. grammica DyP
3.4. XgrDyP Encoding Genes and Their Deduced Protein Sequences
4. Discussion
4.1. Production of a New DyP-Type Peroxidase from the Ascomycete X. grammica
4.2. Comparison of XgrDyP with Other Fungal DyP-Type Peroxidases
4.3. Catalytic Properties of X. grammica DyP
4.4. Structural Aspects
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, S.J.; Ishikawa, K.; Hirai, M.; Shoda, M. Characteristics of a newly isolated fungus, Geotrichum candidum Dec 1, which decolorizes various dyes. J. Ferment. Bioeng. 1995, 79, 601–607. [Google Scholar] [CrossRef]
- Kim, S.J.; Shoda, M. Purification and Characterization of a Novel Peroxidase from Geotrichum candidum Dec 1 Involved in Decolorization of Dyes. Appl. Environ. Microbiol. 1999, 65, 1029–1035. [Google Scholar] [CrossRef] [Green Version]
- Sugano, Y.; Sasaki, K.; Shoda, M. cDNA cloning and genetic analysis of a novel decolorizing enzyme, peroxidase gene dyp from Geotrichum candidum Dec 1. J. Biosci. Bioeng. 1999, 87, 411–417. [Google Scholar] [CrossRef]
- Sugano, Y.; Matsushima, Y.; Tsuchiya, K.; Aoki, H.; Hirai, M.; Shoda, M. Degradation pathway of an anthraquinone dye catalyzed by a unique peroxidase DyP from Thanatephorus cucumeris Dec 1. Biodegradation 2009, 20, 433–440. [Google Scholar] [CrossRef]
- Hofrichter, M.; Ullrich, R.; Pecyna, M.J.; Liers, C.; Lundell, T. New and classic families of secreted fungal heme peroxidases. Appl. Microbiol. Biotechnol. 2010, 87, 871–897. [Google Scholar] [CrossRef] [PubMed]
- Strittmatter, E.; Wachter, S.; Liers, C.; Ullrich, R.; Hofrichter, M.; Plattner, D.A.; Piontek, K. Radical formation on a conserved tyrosine residue is crucial for DyP activity. Arch. Biochem. Biophys. 2013, 537, 161–167. [Google Scholar] [CrossRef]
- Scheibner, M.; Hülsdau, B.; Zelena, K.; Nimtz, M.; De Boer, L.; Berger, R.G.; Zorn, H. Novel peroxidases of Marasmius scorodonius degrade β-carotene. Appl. Microbiol. Biotechnol. 2008, 77, 1241–1250. [Google Scholar] [CrossRef] [Green Version]
- Liers, C.; Bobeth, C.; Pecyna, M.; Ullrich, R.; Hofrichter, M. DyP-like peroxidases of the jelly fungus Auricularia auricula-judae oxidize nonphenolic lignin model compounds and high-redox potential dyes. Appl. Microbiol. Biotechnol. 2010, 85, 1869–1879. [Google Scholar] [CrossRef] [PubMed]
- Liers, C.; Pecyna, M.J.; Kellner, H.; Worrich, A.; Zorn, H.; Steffen, K.T.; Hofrichter, M.; Ullrich, R. Substrate oxidation by dye-decolorizing peroxidases (DyPs) from wood- and litter-degrading agaricomycetes compared to other fungal and plant heme-peroxidases. Appl. Microbiol. Biotechnol. 2012, 97, 5839–5849. [Google Scholar] [CrossRef]
- Rahmanpour, R.; Rea, D.; Jamshidi, S.; Fülöp, V.; Bugg, T.D. Structure of Thermobifida fusca DyP-type peroxidase and activity towards Kraft lignin and lignin model compounds. Arch. Biochem. Biophys. 2016, 594, 54–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, J.N.; Singh, R.; Grigg, J.C.; Murphy, M.E.P.; Bugg, T.D.H.; Eltis, L.D. Characterization of Dye-Decolorizing Peroxidases from Rhodococcus jostii RHA1. Biochemistry 2011, 50, 5108–5119. [Google Scholar] [CrossRef]
- Fernández-Fueyo, E.; Linde, D.; Almendral, D.; López-Lucendo, M.F.; Ruiz-Dueñas, F.J.; Martínez, A.T. Description of the first fungal dye-decolorizing peroxidase oxidizing manganese(II). Appl. Microbiol. Biotechnol. 2015, 99, 8927–8942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugano, Y.; Muramatsu, R.; Ichiyanagi, A.; Sata, T.; Shoda, M. DyP, a unique dye-decolorizing peroxidase, represents a novel heme peroxidase family: ASP171 replaces the distal histidine of classical peroxidases. J. Biol. Chem. 2007, 282, 36652–36658. [Google Scholar] [CrossRef] [Green Version]
- Linde, D.; Coscolín, C.; Liers, C.; Hofrichter, M.; Martínez, A.T.; Ruiz-Dueñas, F.J. Heterologous expression and physico-chemical characterization of a fungal dye-decolorizing peroxidase from Auricularia auricula-judae. Protein Expr. Purif. 2014, 103, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Linde, D.; Pogni, R.; Cañellas, M.; Lucas, F.; Guallar, V.; Baratto, M.C.; Sinicropi, A.; Sáez-Jiménez, V.; Coscolín, C.; Romero, A.; et al. Catalytic surface radical in dye-decolorizing peroxidase: A computational, spectroscopic and site-directed mutagenesis study. Biochem. J. 2015, 466, 253–262. [Google Scholar] [CrossRef]
- Yoshida, T.; Tsuge, H.; Konno, H.; Hisabori, T.; Sugano, Y. The catalytic mechanism of dye-decolorizing peroxidase DyP may require the swinging movement of an aspartic acid residue. FEBS J. 2011, 278, 2387–2394. [Google Scholar] [CrossRef]
- Yoshida, T.; Tsuge, H.; Hisabori, T.; Sugano, Y. Crystal structures of dye-decolorizing peroxidase with ascorbic acid and 2,6-dimethoxyphenol. FEBS Lett. 2012, 586, 4351–4356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strittmatter, E.; Liers, C.; Ullrich, R.; Wachter, S.; Hofrichter, M.; Plattner, D.A.; Piontek, K. First crystal structure of a fungal high-redox potential dye-decolorizing peroxidase substrate interaction sites and long-range electron transfer. J. Biol. Chem. 2013, 288, 4095–4102. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, A.; Chang, H.Y.; Daugherty, L.; Fraser, M.; Hunter, S.; López, R.; McAnulla, C.; McMenamin, C.; Nuka, G.; Pesseat, S.; et al. The InterPro protein families database: The classification resource after 15 years. Nucleic Acids Res. 2015, 43, D213–D221. [Google Scholar] [CrossRef]
- Van Bloois, E.; Pazmiño, D.E.T.; Winter, R.T.; Fraaije, M.W. A robust and extracellular heme-containing peroxidase from Thermobifida fusca as prototype of a bacterial peroxidase superfamily. Appl. Microbiol. Biotechnol. 2009, 86, 1419–1430. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Sugano, Y. A structural and functional perspective of DyP-type peroxidase family. Arch. Biochem. Biophys. 2015, 574, 49–55. [Google Scholar] [CrossRef]
- Zámocký, M.; Obinger, C. Molecular phylogeny of heme peroxidases. In Biocatalysis Based on Heme Peroxidases, 1st ed.; Torres, E., Ayala, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 7–35. [Google Scholar]
- Salvachúa, D.; Martínez, A.T.; Tien, M.; López-Lucendo, M.F.; García, F.; Ríos, V.D.L.; Martínez, M.J.; Prieto, A. Differential proteomic analysis of the secretome of Irpex lacteus and other white-rot fungi during wheat straw pretreatment. Biotechnol. Biofuels 2013, 6, 115. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, K.; Igeta, E.; Amano, Y.; Hyuga, M.; Sugano, Y. Degradation of antifungal anthraquinone compounds is a probable physiological role of DyP secreted by Bjerkandera adusta. AMB Express 2019, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kellner, H.; Luis, P.; Pecyna, M.J.; Barbi, F.; Kapturska, D.; Krüger, D.; Zak, D.R.; Marmeisse, R.; Vandenbol, M.; Hofrichter, M. Widespread Occurrence of Expressed Fungal Secretory Peroxidases in Forest Soils. PLoS ONE 2014, 9, e95557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Büttner, E.; Kimani, V.W.; Kellner, H.; Hofrichter, M.; Liers, C. Genome Sequence of the Versatile Deadwood Decomposer Xylaria grammica IHIA82. Microbiol. Resour. Announc. 2021, 10, 1306–1320. [Google Scholar] [CrossRef]
- Gröbe, G.; Ullrich, R.; Pecyna, M.J.; Kapturska, D.; Friedrich, S.; Hofrichter, M.; Scheibner, K. High-yield production of aromatic peroxygenase by the agaric fungus Marasmius rotula. AMB Express 2011, 1, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiebist, J.; Schmidtke, K.-U.; Zimmermann, J.; Kellner, H.; Jehmlich, N.; Ullrich, R.; Zänder, D.; Hofrichter, M.; Scheibner, K. A Peroxygenase from Chaetomium globosum Catalyzes the Selective Oxygenation of Testosterone. ChemBioChem 2017, 18, 563–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfenden, B.S.; Willson, R.L. Radical-cations as reference chromogens in kinetic studies of one-electron transfer reactions: Pulse radiolysis studies of 2,2′-azinobis-(3-ethylbenzthiazoline-6-sulphonate). J. Chem. Soc. Perkin. Trans. 1982, 2, 805–812. [Google Scholar] [CrossRef]
- Wariishi, H.; Valli, K.; Gold, M. Manganese(II) oxidation by manganese peroxidase from the basidiomycete Phanerochaete chrysosporium. Kinetic mechanism and role of chelators. J. Biol. Chem. 1992, 267, 23688–23695. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Bollag, J.-M.; Sjoblad, R.D.; Liu, S.-Y. Characterization of an enzyme from Rhizoctonia praticola which polymerizes phenolic compounds. Can. J. Microbiol. 1979, 25, 229–233. [Google Scholar] [CrossRef]
- Shimokawa, T.; Hirai, M.; Shoda, M.; Sugano, Y. Efficient Dye Decolorization and Production of Dye Decolorizing Enzymes by the Basidiomycete Thanatephorus cucumeris Dec 1 in a Liquid and Solid Hybrid Culture. J. Biosci. Bioeng. 2008, 106, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Zhang, C.; Li, Y.; Pearce, R.; Bell, E.W.; Zhang, Y. Folding non-homologous proteins by coupling deep-learning contact maps with I-TASSER assembly simulations. Cell Rep. Methods 2021, 1, 100014. [Google Scholar] [CrossRef] [PubMed]
- Pecyna, M.J. Molekularbiologische Charakterisierung von Häm-Thiolat-und DyP-Type-Peroxidasen Ausgewählter Basidiomyceten. Ph.D. Thesis, Technical University Dresden, Dresden, Germany, 2015. [Google Scholar]
- Hofrichter, M.; Kellner, H.; Pecyna, M.J.; Ullrich, R. Fungal Unspecific Peroxygenases: Heme-Thiolate Proteins That Combine Peroxidase and Cytochrome P450 Properties. In Advances in Experimental Medicine and Biology; Springer: Berlin/Heidelberg, Germany, 2015; Volume 851, pp. 341–368. [Google Scholar]
- Liers, C.; Arnstadt, T.; Ullrich, R.; Hofrichter, M. Patterns of lignin degradation and oxidative enzyme secretion by different wood- and litter-colonizing basidiomycetes and ascomycetes grown on beech-wood. FEMS Microbiol. Ecol. 2011, 78, 91–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, M.E.; Barros, T.; Chang, M.C.Y. Identification and characterization of a multifunctional dye peroxidase from a lig-nin-reactive bacterium. ACS Chem. Biol. 2012, 7, 2074–2081. [Google Scholar] [CrossRef]
- Rahmanpour, R.; Bugg, T.D. Characterisation of Dyp-type peroxidases from Pseudomonas fluorescens Pf-5: Oxidation of Mn(II) and polymeric lignin by Dyp1B. Arch. Biochem. Biophys. 2015, 574, 93–98. [Google Scholar] [CrossRef]
- Pease, E.A.; Tien, M. Heterogeneity and regulation of manganese peroxidases from Phanerochaete chrysosporium. J. Bacteriol. 1992, 174, 3532–3540. [Google Scholar] [CrossRef] [Green Version]
- Steffen, K.T.; Hatakka, A.; Hofrichter, M. Degradation of humic acids by the litter-decomposing basidiomycete Collybia dry-ophila. Appl. Environ. Microbiol. 2002, 68, 3442–3448. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.; Martínez, A.T.; Martínez, M.J. Biochemical and molecular characterization of a manganese peroxidase isoenzyme from Pleurotus ostreatus. Biochim. Biophys. Acta Protein Struct. Mol. Enzym. 1997, 1339, 23–30. [Google Scholar] [CrossRef]
- Martinez, M.J.; Ruiz-Duenas, F.J.; Guillen, F.; Martínez, A.T. Purification and Catalytic Properties of Two Manganese Peroxidase Isoenzymes from Pleurotus eryngii. JBIC J. Biol. Inorg. Chem. 1996, 237, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Vazquez-Duhalt, R.; Pickard, M.A. Purification, characterization, and chemical modification of manganese perox-idase from Bjerkandera adusta UAMH 8258. Curr. Microbiol. 2002, 45, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Giardina, P.; Palmieri, G.; Fontanella, B.; Rivieccio, V.; Sannia, G. Manganese Peroxidase Isoenzymes Produced by Pleurotus ostreatus Grown on Wood Sawdust. Arch. Biochem. Biophys. 2000, 376, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Dueñas, F.J.; Martínez, Á.T. Microbial degradation of lignin: How a bulky recalcitrant polymer is efficiently recycled in nature and how we can take advantage of this. Microb. Biotechnol. 2009, 2, 164–177. [Google Scholar] [CrossRef] [Green Version]
- Hofrichter, M. Review: Lignin conversion by manganese peroxidase (MnP). Enzym. Microb. Technol. 2002, 30, 454–466. [Google Scholar] [CrossRef]
- Arnstadt, T.; Hoppe, B.; Kahl, T.; Kellner, H.; Krüger, D.; Bässler, C.; Bauhus, J.; Hofrichter, M. Patterns of laccase and peroxidases in coarse woody debris of Fagus sylvatica, Picea abies and Pinus sylvestris and their relation to different wood parameters. Eur. J. For. Res. 2015, 135, 109–124. [Google Scholar] [CrossRef] [Green Version]
- Noll, L.; Leonhardt, S.; Arnstadt, T.; Hoppe, B.; Poll, C.; Matzner, E.; Hofrichter, M.; Kellner, H. Fungal biomass and extra-cellular enzyme activities in coarse woody debris of 13 tree species in the early phase of decomposition. For. Ecol. Manag. 2016, 378, 181–192. [Google Scholar] [CrossRef]
- Büttner, E.; Gebauer, A.M.; Hofrichter, M.; Liers, C.; Kellner, H. Draft Genome Sequence of the Wood-Degrading Ascomycete Kretzschmaria deusta DSM 104547. Genome Announc. 2017, 5, e01076-17. [Google Scholar] [CrossRef] [Green Version]
- Büttner, E.; Gebauer, A.M.; Hofrichter, M.; Liers, C.; Kellner, H. Draft Genome Sequence of Scytalidium lignicola DSM 105466, a Ubiquitous Saprotrophic Fungus. Microbiol. Resour. Announc. 2018, 7, e01208-18. [Google Scholar] [CrossRef] [Green Version]
- Büttner, E.; Liers, C.; Gebauer, A.M.; Collemare, J.; Navarro, J.; Hofrichter, M.; Kellner, H. Draft Genome Sequence of the Wood-Staining Ascomycete Chlorociboria aeruginascens DSM 107184. Microbiol. Resour. Announc. 2019, 8, e00249-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liers, C.; Ullrich, R.; Steffen, K.T.; Hatakka, A.; Hofrichter, M. Mineralization of 14C-labelled synthetic lignin and extracellular enzyme activities of the wood-colonizing ascomycetes Xylaria hypoxylon and Xylaria polymorpha. Appl. Microbiol. Biotechnol. 2005, 69, 573–579. [Google Scholar] [CrossRef]
- Fernández-Fueyo, E.; Davó-Siguero, I.; Almendral, D.; Linde, D.; Baratto, M.C.; Pogni, R.; Romero, A.; Guallar, V.; Martínez, A.T. Description of a Non-Canonical Mn(II)-Oxidation Site in Peroxidases. ACS Catal. 2018, 8, 8386–8395. [Google Scholar] [CrossRef]
- Sundaramoorthy, M.; Kishi, K.; Gold, M.H.; Poulos, T.L. The crystal structure of manganese peroxidase from Phanerochaete chrysosporium at 2.06-A resolution. J. Biol. Chem. 1994, 269, 32759–32767. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purifications Step | Activity (U) | Specific Activity (U mg−1) | Protein Amount (mg) | Yield (%) | Purification (x-Fold) |
|---|---|---|---|---|---|
| Culture filtrate | 2028 | 3.4 | 595.7 | 100 | - |
| Ultrafiltrate 10 kDa | 1923 | 3.3 | 584.0 | 95 | 1.0 |
| Q-Sepharose_I | 1175 | 4.4 | 265.2 | 58 | 1.3 |
| Q-Sepharose_II | 1160 | 4.6 | 254.4 | 57 | 1.4 |
| SEC | 1301 | 10.9 | 119.9 | 64 | 3.2 |
| MonoQ_I | 568 | 13.8 | 41.1 | 28 | 4.1 |
| MonoQ_II | 120 | 51.0 | 2.4 | 6 | 15.0 |
| Substrate | kcat (s−1) | KM (µM) | kcat/KM (s−1 mM−1) |
|---|---|---|---|
| ABTS | 12 | 41 | 287 |
| 2,6-DMP | 29 | 12 | 2499 |
| RBlue5 | 20 | 41 | 495 |
| Mn2+ | 0.4 | 49 | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kimani, V.; Ullrich, R.; Büttner, E.; Herzog, R.; Kellner, H.; Jehmlich, N.; Hofrichter, M.; Liers, C. First Dye-Decolorizing Peroxidase from an Ascomycetous Fungus Secreted by Xylaria grammica. Biomolecules 2021, 11, 1391. https://doi.org/10.3390/biom11091391
Kimani V, Ullrich R, Büttner E, Herzog R, Kellner H, Jehmlich N, Hofrichter M, Liers C. First Dye-Decolorizing Peroxidase from an Ascomycetous Fungus Secreted by Xylaria grammica. Biomolecules. 2021; 11(9):1391. https://doi.org/10.3390/biom11091391
Chicago/Turabian StyleKimani, Virginia, René Ullrich, Enrico Büttner, Robert Herzog, Harald Kellner, Nico Jehmlich, Martin Hofrichter, and Christiane Liers. 2021. "First Dye-Decolorizing Peroxidase from an Ascomycetous Fungus Secreted by Xylaria grammica" Biomolecules 11, no. 9: 1391. https://doi.org/10.3390/biom11091391