
The Surprising Story of Fusicoccin: A Wilt-Inducing Phytotoxin, a Tool in Plant Physiology and a 14-3-3-Targeted Drug †


Abstract
:1. FC: A Wilt-Inducing Phytotoxin
1.1. Discovery
1.2. Structure Determination
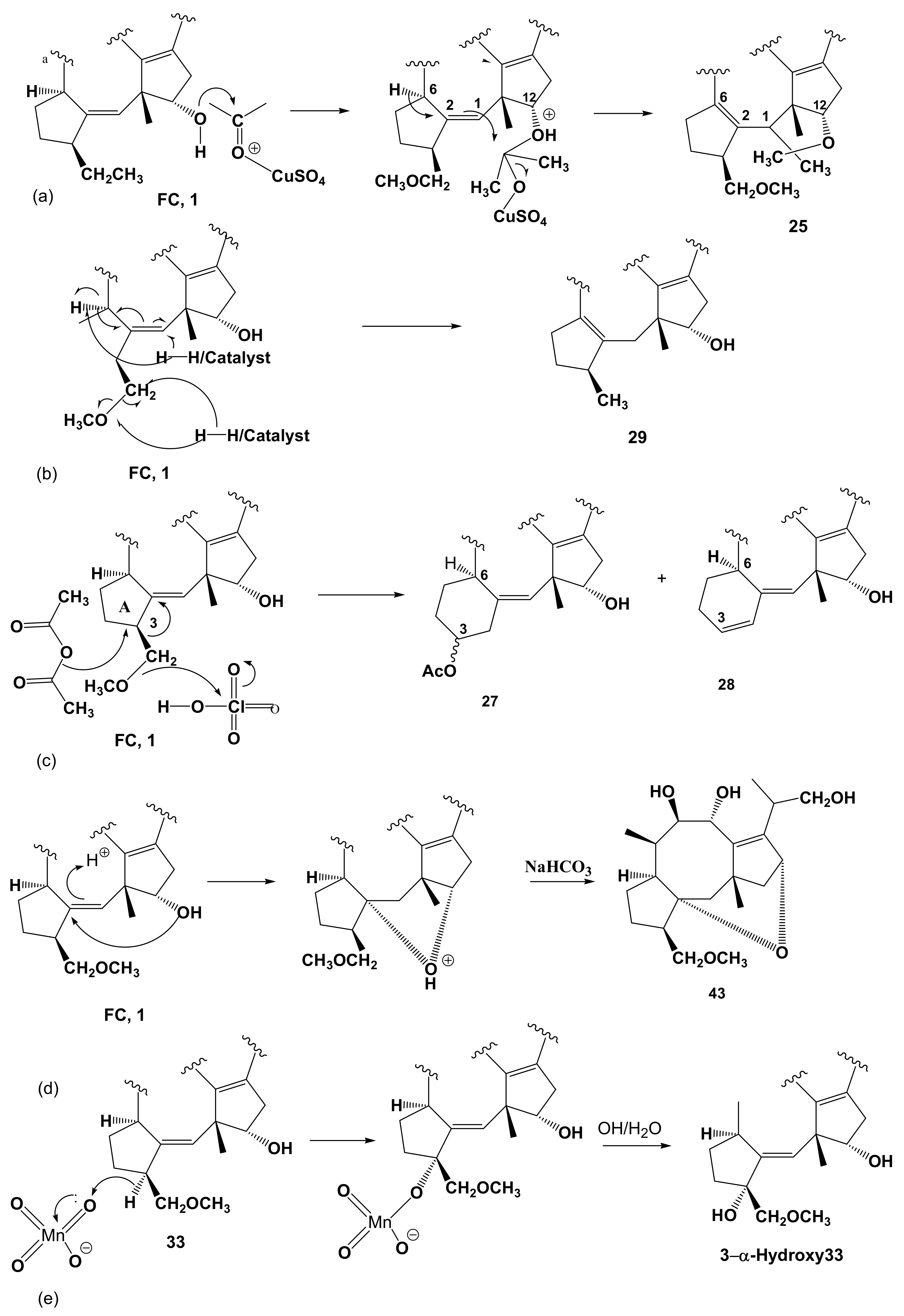
1.3. Minor Diterpenoid Metabolites of Phomopsis amygdali
1.4. Biosynthesis
2. FC: A Tool in Plant Physiology
2.1. Biological Activity
2.2. Structure-Activity Relationships
2.3. Mode of Action in Plants
3. FC: A 14-3-3-Targeted Drug
3.1. Stabilization of 14-3-3/Mode III-Client Interactions
3.2. FC Derivatives for Pharmacological Applications
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Graniti, A. Phytotoxic action of Fusicoccum amygdali Del. on almond (Prunus amygdali St.). Phytopathol. Mediterr. 1962, 1, 182–185. [Google Scholar]
- Graniti, A. Some phytotoxicity data of fusicoccin A, a toxin produced in vitro by Fusicoccum amygdali Del. Phytopathol. Mediterr. 1964, 3, 125–128. [Google Scholar]
- Ballio, A.; Chain, E.B.; De Leo, P.; Erlanger, B.F.; Mauri, M.; Tonolo, A. Fusicoccin: A New Wilting Toxin produced by Fusicoccum amygdali Del. Nature 1964, 203, 297. [Google Scholar] [CrossRef]
- Turner, N.; Graniti, A. Fusicoccin: A Fungal Toxin that opens Stomata. Nature 1969, 223, 1070–1071. [Google Scholar] [CrossRef]
- Turner, N.C.; Graniti, A. Stomatal response of two almond cultivars to fusicoccin. Physiol. Plant Pathol. 1976, 9, 175–182. [Google Scholar] [CrossRef]
- Ballio, A.; D’Alessio, V.; Randazzo, G.; Bottalico, A.; Graniti, A.; Sparapano, L.; Bosnar, B.; Casinovi, C.; Gribanovski-Sassu, O. Occurrence of fusicoccin in plant tissues infected by Fusicoccum amygdali del. Physiol. Plant Pathol. 1976, 8, 163–169. [Google Scholar] [CrossRef]
- Ballio, A.; Brufani, M.; Casinovi, C.G.; Cerrini, S.; Fedeli, W.; Pellicciari, R.; Santurbano, B.; Vaciago, A. The structure of fusicoccin A. Cell. Mol. Life Sci. 1968, 24, 631–635. [Google Scholar] [CrossRef]
- Barrow, K.D.; Barton, D.H.R.; Chain, E.B.; Ohnsorge, U.F.W.; Thomas, R. The constitution of fusicoccin. Chem. Commun. 1968, 1198–1200. [Google Scholar] [CrossRef]
- Ballio, A.; Casinovi, C.G.; Randazzo, G.; Rossi, C. Characterization of by-products of fusicoccin in culture filtrates of Fusicoccum amygdali Del. Cell. Mol. Life Sci. 1970, 26, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Ballio, A.; Bottalico, A.; Framondino, M.; Graniti, A.; Randazzo, G. Fusicoccin: Structure-phytotoxicity relationships. Phyto-pathol. Mediterr. 1971, 10, 26–32. [Google Scholar]
- Ballio, A.; Pocchiari, F.; Russi, S.; Silano, V. Effects of fusicoccin and some related compounds on etiolated pea tissues. Physiol. Plant Pathol. 1971, 1, 95–103. [Google Scholar] [CrossRef]
- Ballio, A.; Bottalico, A.; Framondino, M.; Graniti, A.; Randazzo, G. Phytotoxicity of minor metabolites of Fusicoccum amygdali Del. and related products. Phytopathol. Mediterr. 1973, 12, 22–29. [Google Scholar]
- Sparapano, L.; Graniti, A.; Feldman, A.W. Effects of fusicoccin and its analogues on the abscission of Citrus leaf explants. Phytopathol. Mediterr. 1971, 10, 33–36. [Google Scholar]
- Feldman, A.W.; Graniti, A.; Sparapano, L. Effect of fusicoccin on abscission, cellulase activity and ethylene production in citrus leaf explants. Physiol. Plant Pathol. 1971, 1, 115–122. [Google Scholar] [CrossRef]
- De Michelis, M.I.; Pugliarello, M.C.; Olivari, C.; Rasi-Caldogno, F. Fusicoccin binding to its plasma membrane receptor and the activation of the plasma membrane H+-ATPase. II. Stimulation of the H+-ATPase in a plasma membrane fraction purified by phase-partitioning. Bot. Acta 1991, 104, 265–271. [Google Scholar] [CrossRef]
- Lanfermeijer, F.C.; Prins, H. Modulation of H+-ATPase activity by fusicoccin in plasma membrane vesicles from oat (Avena sativa L.) roots (A comparison of modulation by fusicoccin, trypsin, and lysophosphatidylcholine). Plant Physiol. 1994, 104, 1277–1285. [Google Scholar] [CrossRef] [Green Version]
- Ballio, A.; Michelis, M.I.; Lado, P.; Randazzo, G. Fusicoccin structure-activity relationships: Stimulation of growth by cell enlargement and promotion of seed germination. Physiol. Plant. 1981, 52, 471–475. [Google Scholar] [CrossRef]
- Yoneyama, K.; Takeuchi, Y.; Ogasawara, M.; Konnai, M.; Sugimoto, Y.; Sassa, T. Cotylenins and fusicoccins stimulate seed germination of Striga hermonthica(Del.) Benth and Orobanche minor Smith. J. Agric. Food Chem. 1998, 46, 1583–1586. [Google Scholar] [CrossRef]
- Evidente, A.; Andolfi, A.; Fiore, M.; Boari, A.; Vurro, M. Stimulation of Orobanche ramosa seed germination by fusicoccin derivatives: A structure–activity relationship study. Phytochemistry 2006, 67, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Ballio, A.; Casinovi, C.G.; Framondino, M.; Grandolini, G.; Menichini, F.; Randazzo, G.; Rossi, C. The structures of isofusicoccin and allofusicoccin. Cell. Mol. Life Sci. 1972, 28, 126–127. [Google Scholar] [CrossRef] [PubMed]
- Ballio, A.; Casinovi, C.G.; Framondino, M.; Grandolini, G.; Randazzo, G.; Rossi, C. The structure of three isomers of monodeacetylfusicoccin. Cell. Mol. Life Sci. 1972, 28, 1150–1151. [Google Scholar] [CrossRef]
- Ballio, A.; Casinovi, C.G.; D’Alessio, V.; Grandolini, G.; Randazzo, G.; Rossi, C. 19-Deoxydideacetylfusicoccin, a minor metabolite of Fusicoccum amygdali Del. Cell. Mol. Life Sci. 1974, 30, 844–845. [Google Scholar] [CrossRef] [PubMed]
- Barrow, K.D.; Barton, D.H.R.; Chain, E.; Ohnsorge, U.F.W.; Sharma, R.P. Fusicoccin. Part III. The structure of fusicoccin H. J. Chem. Soc. Perkin Trans. 1 1973, 1590–1599. [Google Scholar] [CrossRef]
- Ballio, A.; Casinovi, G.C.; Grandolini, G.; Marta, M.; Randazzo, G. 19-Deoxy-3α-hydroxydideacetyl-fusicoccin. Gazz. Chim. Ital. 1975, 105, 1325–1328. [Google Scholar]
- Randazzo, G.; Evidente, A.; Capasso, R.; Chiosi, S.; Casinovi, G.C.; Ballio, A. A new 3α-Hydroxyfusicoccin from culture filtrates of Fusicoccum amygdali Del. Phytopathol. Mediterr. 1980, 19, 181–182. [Google Scholar]
- Randazzo, G.; Evidente, A.; Chiosi, S.; Casinovi, C.G. Regiospecific and streoselective 3-alpha-hydroxylation of fusicoccin derivatives. Gazz. Chim. Ital. 1981, 111, 81–84. [Google Scholar]
- Ballio, A.; Casinovi, G.C.; Grandolini, G.; Pomponi, M.; Randazzo, G.; Rossi, C. 16-O-Demethyl-19-deoxydideacetyl-3-epifusicoccin J. Gazz. Chim. Ital. 1975, 105, 645–650. [Google Scholar]
- Cerrini, S.W.; Fedeli, W.; Gavuzzo, E. Crystal structure of the aglycone of 16-O-demethyl-19-deoxydideadecyl-3-epifusicoccin. Gazz. Chim. Ital. 1975, 105, 651–655. [Google Scholar]
- Barrow, K.D.; Barton, D.H.R.; Chain, E.; Conlay, C.; Smale, T.C.; Thomas, R.; Waight, E.S. Fusicoccin. Part I. The nature of the substituent groups. J. Chem. Soc. C 1971, 1259–1264. [Google Scholar] [CrossRef]
- Capasso, R.; Evidente, A.; Lasaponara, M.; Randazzo, G. Una insolita reazione dell’acetone sulla fusicoccina. Rend. Accad. Sci. Fis. Mat. Napoli 1977, 44, 1–5. [Google Scholar]
- Chiosi, S.; Evidente, A.; Randazzo, G.; Segre, A.L.; Ballio, A. The reaction of fusicoccin with Fritz and Schenk reagent. Gazz. Chim. Ital. 1983, 113, 717–720. [Google Scholar]
- Canonica, L.; Fiecchi, A.; Kienle, M.; Ranzi, B.; Scala, A.; Salvatori, T.; Pella, E. The biosynthesis-of ophiobolins. Tetrahedron Lett. 1967, 8, 3371–3376. [Google Scholar] [CrossRef]
- Banerji, A.; Hunter, R.; Mellows, G.; Sim, K.-Y.; Barton, D.H.R. Use of deuterium as a tracer with 13C nuclear magnetic resonance spectroscopy in following deuteride migration in terpenoid biosynthesis: Mechanism of geranylgeranyl pyrophosphate cyclisation in fusicoccin biosynthesis. J. Chem. Soc. Chem. Commun. 1978, 843–845. [Google Scholar] [CrossRef]
- Barrow, K.D.; Barton, D.H.R.; Chain, E.; Ohnsorge, U.F.W.; Thomas, R. Fusicoccin. Part II. The constitution of fusicoccin. J. Chem. Soc. C 1971, 1265–1274. [Google Scholar] [CrossRef]
- Randazzo, G.; Evidente, A.; Capasso, R.; Colantuoni, F.; Tuttobello, L.; Ballio, A. Further experiments on the biosynthesis of fusicoccin. Gazz. Chim. Ital. 1979, 109, 101–104. [Google Scholar]
- Sassa, T.; Negoro, T.; Ueki, H. Production and characterization of a new fungal metabolite, cotylenol. Agric. Biol. Chem. 1972, 36, 2281–2285. [Google Scholar] [CrossRef]
- Sassa, T.; Togashi, M.; Kitaguchi, T. The structures of cotylenins A, B, C, D and E. Agric. Biol. Chem. 1975, 39, 1735–1744. [Google Scholar] [CrossRef]
- Sassa, T. The mass spectra of cotylenol and cotylenins. Agric. Biol. Chem. 1974, 38, 2041–2043. [Google Scholar]
- Evidente, A.; Randazzo, G. The biosynthetic origin of fusicoccin hydroxyisopropyl group. Tetrahedron 1982, 38, 3169–3172. [Google Scholar] [CrossRef]
- Casinovi, C.G.; Santurbano, B.; Conti, G.; Malorni, A.; Randazzo, G. The structure and spectroscopic properties of an isomer of fusicoccin deacetylaglycone and some derivatives. Gazz. Chim. Ital. 1974, 104, 679–691. [Google Scholar]
- Au, T.; Chick, W.S.; Leung, P. The biology of ophiobolins. Life Sci. 2000, 67, 733–742. [Google Scholar] [CrossRef]
- Nozoe, S.; Morisaki, M.; Tsuda, K.; Iitaka, Y.; Takahashi, N.; Tamura, S.; Ishibashi, K.; Shirasaka, M. The structure of ophiobolin, a C25 terpenoid having a novel skeleton. J. Am. Chem. Soc. 1965, 87, 4968–4970. [Google Scholar] [CrossRef] [PubMed]
- Canonica, L.; Friecchi, A.; Kienle, U.; Scala, A. Isolation and constitution of cochliobolin b. Tetrahedron Lett. 1966, 7, 1329–1333. [Google Scholar] [CrossRef]
- Ohkawa, H.; Tamura, T. Studies on the Metabolites of Cochliobolus miyabeanus. Agric. Biol. Chem. 1966, 30, 285–291. [Google Scholar] [CrossRef]
- Sassa, T. Cotylenins, leaf growth substances produced by a fungus. Agric. Biol. Chem. 1971, 35, 1415–1418. [Google Scholar] [CrossRef]
- Sugawara, F.; Strobel, G.; Strange, R.N.; Siedow, J.N.; Van Duyne, G.D.; Clardy, J. Phytotoxins from the pathogenic fungi Drechslera maydis and Drechslera sorghicola. Proc. Natl. Acad. Sci. USA 1987, 84, 3081–3085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sassa, T.; Ooi, T.; Nukina, M.; Ikeda, M.; Kato, N. Structural confirmation of cotylenin A, a novel fusicoccane-diterpene glycoside with potent plant growth-regulating activity from Cladosporium fungus sp. 501-7W. Biosci. Biotechnol. Biochem. 1998, 62, 1815–1818. [Google Scholar] [CrossRef] [Green Version]
- Masi, M.; Dasari, R.; Evidente, A.; Mathieu, V.; Kornienko, A. Chemistry and biology of ophiobolin A and its congeners. Bioorganic Med. Chem. Lett. 2019, 29, 859–869. [Google Scholar] [CrossRef]
- Barrow, K.D.; Barton, D.H.R.; Chain, E.B.; Smale, T.V.; Waight, E.S.; Conlay, C.; Thomas, R. Fusicoccin: Characterisation of the oxygen substituents. Chem. Commun. 1968, 1195–1197. [Google Scholar] [CrossRef]
- Ballio, A.; Carilli, A.; Santurbano, B.; Tuttobello, L. Produzione di fusicoccina in scala pilota. Ann. Ist. Sup. Sanità Roma 1968, 4, 317–332. [Google Scholar]
- Evidente, A.; Capasso, R.; Randazzo, G.; Ballio, A.; Tuttobello, L. The biosynthesis of fusicoccin. Phytopathol. Mediterr. 1980, 19, 170–180. [Google Scholar]
- Fritz, J.S.; Schenk, G.H. Acid-catalyzed acetylation of organic hydroxyl groups. Anal. Chem. 1959, 31, 1808–1812. [Google Scholar] [CrossRef]
- Canonica, L.; Fiecchi, A.; Kienle, H.; Ranzi, B.; Scala, A. The streochemical, course of the 1,5-shift of hydrogen in the biosynthesis of ophiobolins. Tetrahedron Lett. 1967, 8, 4657–4659. [Google Scholar] [CrossRef]
- Randazzo, G.; Evidente, A.; Capasso, R.; Lasaponara, M.; Tuttobello, L.; Ballio, A. On the biosynthesis of fusicoccin. Gazz. Chim. Ital. 1978, 108, 139–142. [Google Scholar]
- Barrow, K.D.; Jones, R.B.; Pemberton, P.W.; Phillips, L. Fusicoccin. Part V. The biosynthesis of fusicoccin from [1-13C]- and [2-13C]-acetate. J. Chem. Soc. Perkin Trans. 1 1975, 1405–1410. [Google Scholar] [CrossRef]
- Banerji, A.; Jones, R.B.; Mellows, G.; Phillips, L.; Sim, K.-Y. Fusicoccin. Part 6. Biosynthesis of fusicoccin from [3-13C]- and (4R)-[4-3H1]-mevalonic acid. J. Chem. Soc. Perkin Trans. 1 1976, 2221–2228. [Google Scholar] [CrossRef]
- Evidente, A.; Randazzo, G.; Capasso, R.; Lasaponara, M. Catalytic hydrogenation of fusicoccin. Gazz. Chim. Ital. 1984, 114, 61–64. [Google Scholar]
- Bowden, K.; Heilbron, I.M.; Jones, E.R.H.; Weedon, B.C.L. 13. Researches on acetylenic compounds. Part I. The preparation of acetylenic ketones by oxidation of acetylenic carbinols and glycols. J. Chem. Soc. 1946, 39–45. [Google Scholar] [CrossRef]
- Toyomasu, T.; Tsukahara, M.; Kaneko, A.; Niida, R.; Mitsuhashi, W.; Dairi, T.; Kato, N.; Sassa, T. Fusicoccins are biosynthesized by an unusual chimera diterpene synthase in fungi. Proc. Natl. Acad. Sci. USA 2007, 104, 3084–3088. [Google Scholar] [CrossRef] [Green Version]
- Marrè, E.; Lado, P.; Rasi-Caldogno, F.; Colombo, R. Fusicoccin as a tool for the analysis of auxin action. Rend. Accad. Naz. Lincei 1971, 50, 45–49. [Google Scholar]
- Marrè, E. Fusicoccin: A tool in plant physiology. Annu. Rev. Plant Physiol. 1979, 30, 273–288. [Google Scholar] [CrossRef]
- Jahn, T.; Fuglsang, A.T.; Olsson, A.; Brüntrup, I.M.; Collinge, D.B.; Volkmann, D.; Sommarin, M.; Palmgren, M.; Larsson, C. The 14-3-3 protein interacts directly with the C-terminal region of the plant plasma membrane H(+)-ATPase. Plant Cell 1997, 9, 1805–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baunsgaard, L.; Fuglsang, A.T.; Jahn, T.; Korthout, H.A.A.J.; De Boer, A.H.; Palmgren, M. Summary. Plant J. 1998, 13, 661–671. [Google Scholar] [CrossRef]
- Piotrowski, M.; Morsomme, P.; Boutry, M.; Oecking, C. Complementation of the Saccharomyces cerevisiae plasma membrane H+-ATPase by a plant H+-ATPase generates a highly abundant fusicoccin binding site. J. Biol. Chem. 1998, 273, 30018–30023. [Google Scholar] [CrossRef] [Green Version]
- Ballio, A.; Castellano, S.; Cerrini, S.; Evidente, A.; Randazzo, G.; Segre, A. 1H NMR conformational study of fusicoccin and related compounds: Molecular conformation and biological activity. Phytochemistry 1991, 30, 137–146. [Google Scholar] [CrossRef]
- Ballio, A. Fusicoccin: Structure-activity relationships. In Regulation of Cell Membrane Activities in Plants; Marrè, E., Ciferi, O., Eds.; Elsevier: Amsterdam, The Netherlands, 1977; pp. 217–223. [Google Scholar]
- Ballio, A. Chemistry and plant growth regulating activity of fusicoccin derivatives and analogues. In Advances in Pesticide Science; GeissbuÈhler, H., Ed.; Pergamon: Oxford, UK, 1979; pp. 366–372. [Google Scholar]
- Dohrmann, U.; Hertel, R.; Pesci, P.; Cocucci, S.; Marrè, E.; Randazzo, G.; Ballio, A. Localization of “in vitro” binding of the fungal toxin fusicoccin to plasma-membrane-rich fractions from corn coleoptiles. Plant Sci. Lett. 1977, 9, 291–299. [Google Scholar] [CrossRef]
- Ballio, A.; Federico, R.; Pessi, A.; Scalorbi, D. Fusicoccin binding sites in subcellular preparations of spinach leaves. Plant Sci. Lett. 1980, 18, 39–44. [Google Scholar] [CrossRef]
- Feyerabend, M.; Weiler, E.W. Characterization and localization of fusicoccin-binding sites in leaf tissues of Vicia faba L. probed with a novel radioligand. Planta 1988, 174, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Abramycheva, N.; Babakov, A.; Bilushi, S.; Danilina, E.; Shevchenko, V. Comparison of the biological activity of fusicoccin in higher plants with its binding to plasma membranes. Planta 1991, 183, 315–320. [Google Scholar] [CrossRef] [PubMed]
- De Michelis, M.I.; Pugliarello, M.C.; Rasi-Caldogno, F. Fusicoccin binding to its plasma membrane receptor and the activation of the plasma membrane H+-ATPase. Plant Physiol. 1989, 90, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Meyer, C.; Feyerabend, M.; Weiler, E.W. Fusicoccin-Binding Proteins in Arabidopsis thaliana (L.) Heynh. Plant Physiol. 1989, 89, 692–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aducci, P.; Ballio, A.; Marra, M. Incubation of corn coleoptiles with auxin enhances in-vitro fusicoccin binding. Planta 1986, 167, 129–132. [Google Scholar] [CrossRef]
- Stout, R.G. Fusicoccin activity and binding in Arabidopsis thaliana. Plant Physiol. 1988, 88, 999–1001. [Google Scholar] [CrossRef] [Green Version]
- Oecking, C.; Weiler, E.W. Characterization and purification of the fusicoccin-binding complex from plasma membranes of Commelina communis. JBIC J. Biol. Inorg. Chem. 1991, 199, 685–689. [Google Scholar] [CrossRef] [PubMed]
- Ballio, A.; Federico, R.; Scalorbi, D. Fusicoccin structure-activity relationships: In vitro binding to microsomal preparations of maize coleoptiles. Physiol. Plant. 1981, 52, 476–481. [Google Scholar] [CrossRef]
- Marra, M.; Fullone, M.R.; Fogliano, V.; Pen, J.; Mattei, M.; Masi, S.; Aducci, P. The 30-kilodalton protein present in purified fusicoccin receptor preparations is a 14-3-3-like protein. Plant Physiol. 1994, 106, 1497–1501. [Google Scholar] [CrossRef] [Green Version]
- Korthout, H.A.; de Boer, A.H. A fusicoccin binding protein belongs to the family of 14-3-3 brain protein homologs. Plant Cell 1994, 6, 1681–1692. [Google Scholar] [CrossRef] [Green Version]
- Oecking, C.; Eckerskorn, C.; Weiler, E.W. The fusicoccin receptor of plants is a member of the 14-3-3 superfamily of eukaryotic regulatory proteins. FEBS Lett. 1994, 352, 163–166. [Google Scholar] [CrossRef]
- Aitken, A.; Collinge, D.; van Heusden, B.; Isobe, T.; Roseboom, P.; Rosenfeld, G.; Soll, J. 14-3-3 proteins: A highly conserved, widespread family of eukaryotic proteins. Trends Biochem. Sci. 1992, 17, 498–501. [Google Scholar] [CrossRef] [Green Version]
- Fu, H.; Subramanian, R.; Masters, S.C. 14-3-3proteins: Structure, function, and regulation. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 617–647. [Google Scholar] [CrossRef] [PubMed]
- Aducci, P.; Camoni, L.; Marra, M.; Visconti, S. From cytosol to organelles: 14-3-3 proteins as multifunctional regulators of plant cell. IUBMB Life 2002, 53, 49–55. [Google Scholar] [CrossRef]
- Hermeking, H. The 14-3-3 cancer connection. Nat. Rev. Cancer 2003, 3, 931–943. [Google Scholar] [CrossRef] [PubMed]
- Freeman, A.K.; Morrison, D.K. 14-3-3 Proteins: Diverse functions in cell proliferation and cancer progression. Semin. Cell Dev. Biol. 2011, 22, 681–687. [Google Scholar] [CrossRef] [Green Version]
- Huber, S.C.; MacKintosh, C.; Kaiser, W.M. Metabolic enzymes as targets for 14-3-3 proteins. Plant Mol. Biol. 2002, 50, 1053–1063. [Google Scholar] [CrossRef]
- Bunney, T.; De Boer, A.H.; Levin, M. Fusicoccin signaling reveals 14-3-3 protein function as a novel step in left-right patterning during amphibian embryogenesis. Development 2003, 130, 4847–4858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sottocornola, B.; Visconti, S.; Orsi, S.; Gazzarrini, S.; Giacometti, S.; Olivari, C.; Camoni, L.; Aducci, P.; Marra, M.; Abenavoli, A.; et al. The potassium channel KAT1 is activated by plant and animal 14-3-3 proteins. J. Biol. Chem. 2006, 281, 35735–35741. [Google Scholar] [CrossRef] [Green Version]
- Camoni, L.; Marra, M.; Garufi, A.; Visconti, S.; Aducci, P. The maize root plasma membrane H+-ATPase is regulated by a sugar-induced transduction pathway. Plant Cell Physiol. 2006, 47, 743–747. [Google Scholar] [CrossRef] [Green Version]
- Taoka, K.-I.; Ohki, I.; Tsuji, H.; Furuita, K.; Hayashi, K.; Yanase, T.; Yamaguchi, M.; Nakashima, C.; Purwestri, Y.A.; Tamaki, S.; et al. 14-3-3 proteins act as intracellular receptors for rice Hd3a florigen. Nature 2011, 476, 332–335. [Google Scholar] [CrossRef] [PubMed]
- Camoni, L.; Visconti, S.; Aducci, P.; Marra, M. 14-3-3 Proteins in plant hormone signaling: Doing several things at once. Front. Plant Sci. 2018, 9, 297. [Google Scholar] [CrossRef] [PubMed]
- Visconti, S.; D’Ambrosio, C.; Fiorillo, A.; Arena, S.; Muzi, C.; Zottini, M.; Aducci, P.; Marra, M.; Scaloni, A.; Camoni, L. Overexpression of 14-3-3 proteins enhances cold tolerance and increases levels of stress-responsive proteins of Arabidopsis plants. Plant Sci. 2019, 289, 110215. [Google Scholar] [CrossRef]
- Marra, M.; Fogliano, V.; Zambardi, A.; Fullone, M.R.; Nasta, D.; Aducci, P. The H+-ATPase purified from maize root plasma membranes retains fusicoccin in vivo activation. FEBS Lett. 1996, 382, 293–296. [Google Scholar] [CrossRef] [Green Version]
- Oecking, C.; Piotrowski, M.; Hagemeier, J.; Hagemann, K. Topology and target interaction of the fusicoccin-binding 14-3-3 homologs of Commelina communis. Plant J. 1997, 12, 441–453. [Google Scholar] [CrossRef]
- Olivari, C.; Meanti, C.; De Michelis, M.I.; Rasi-Caldogno, F. Fusicoccin Binding to its plasma membrane receptor and the activation of the plasma membrane H+-ATPase1. Plant Physiol. 1998, 116, 529–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camoni, L.; Iori, V.; Marra, M.; Aducci, P. Phosphorylation-dependent interaction between plant plasma membrane H+-ATPase and 14-3-3 proteins. J. Biol. Chem. 2000, 275, 9919–9923. [Google Scholar] [CrossRef] [Green Version]
- Pallucca, R.; Visconti, S.; Camoni, L.; Cesareni, G.; Melino, S.M.; Panni, S.; Torreri, P.; Aducci, P. Specificity of ε and non-ε isoforms of Arabidopsis 14-3-3 proteins towards the H+-ATPase and other targets. PLoS ONE 2014, 9, e90764. [Google Scholar] [CrossRef] [Green Version]
- Fullone, M.R.; Visconti, S.; Marra, M.; Fogliano, V.; Aducci, P. Fusicoccin effect on the in vitro interaction between plant 14-3-3 proteins and plasma membrane H+-ATPase. J. Biol. Chem. 1998, 273, 7698–7702. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Bienkowska, J.; Petosa, C.; Collier, R.J.; Fu, H.; Liddington, R.C. Crystal structure of the zeta isoform of the 14-3-3 protein. Nature 1995, 376, 191–194. [Google Scholar] [CrossRef]
- Xiao, B.; Smerdon, S.; Jones, D.H.; Dodson, G.G.; Soneji, Y.; Aitken, A.; Gamblin, S. Structure of a 14-3-3 protein and implications for coordination of multiple signalling pathways. Nature 1995, 376, 188–191. [Google Scholar] [CrossRef]
- Oecking, C.; Jaspert, N. Plant 14-3-3 proteins catch up with their mammalian orthologs. Curr. Opin. Plant Biol. 2009, 12, 760–765. [Google Scholar] [CrossRef]
- Yaffe, M.B.; Rittinger, K.; Volinia, S.; Caron, P.R.; Aitken, A.; Leffers, H.; Gamblin, S.; Smerdon, S.; Cantley, L. The Structural Basis for 14-3-3:Phosphopeptide Binding Specificity. Cell 1997, 91, 961–971. [Google Scholar] [CrossRef] [Green Version]
- Ottmann, C.; Marco, S.; Jaspert, N.; Marcon, C.; Schauer, N.; Weyand, M.; Vandermeeren, C.; Duby, G.; Boutry, M.; Wittinghofer, A.; et al. Structure of a 14-3-3 coordinated hexamer of the plant plasma membrane H+-ATPase by combining X-ray crystallography and electron cryomicroscopy. Mol. Cell 2007, 25, 427–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuglsang, A.T.; Visconti, S.; Drumm, K.; Jahn, T.; Stensballe, A.; Mattei, B.; Jensen, O.N.; Aducci, P.; Palmgren, M. Binding of 14-3-3 protein to the plasma membrane H+-ATPase AHA2 involves the three C-terminal residues Tyr946-Thr-Val and requires phosphorylation of Thr947. J. Biol. Chem. 1999, 274, 36774–36780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svennelid, F.; Olsson, A.; Piotrowski, M.; Rosenquist, M.; Ottman, C.; Larsson, C.; Oecking, C.; Sommarin, M. Phosphorylation of Thr-948 at the C terminus of the plasma membrane H+-ATPase creates a binding site for the regulatory 14-3-3 protein. Plant Cell 1999, 11, 2379–2391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Würtele, M.; Jelich-Ottmann, C.; Wittinghofer, A.; Oecking, C. Structural view of a fungal toxin acting on a 14-3-3 regulatory complex. EMBO J. 2003, 22, 987–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T. UCSF Chimera? A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Coblitz, B.; Wu, M.; Shikano, S.; Li, M. C-terminal binding: An expanded repertoire and function of 14-3-3 proteins. FEBS Lett. 2006, 580, 1531–1535. [Google Scholar] [CrossRef] [Green Version]
- Camoni, L.; Di Lucente, C.; Visconti, S.; Aducci, P. The phytotoxin fusicoccin promotes platelet aggregation via 14-3-3–glycoprotein Ib-IX-V interaction1. Biochem. J. 2011, 436, 429–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camoni, L.; Visconti, S.; Aducci, P. The phytotoxin fusicoccin, a selective stabilizer of 14-3-3 interactions? IUBMB Life 2013, 65, 513–517. [Google Scholar] [CrossRef]
- Paiardini, A.; Aducci, P.; Cervoni, L.; Cutruzzolà, F.; Di Lucente, C.; Janson, G.; Pascarella, S.; Rinaldo, S.; Visconti, S.; Camoni, L. The phytotoxin fusicoccin differently regulates 14-3-3 proteins association to mode III targets. IUBMB Life 2014, 66, 52–62. [Google Scholar] [CrossRef]
- Meyer, C.; Waldkötter, K.; Sprenger, A.; Schlösset, U.G.; Luther, M.; Weiler, E.W. Survey of the taxonomic and tissue distribution of microsomal binding sites for the non-host selective fungal phytotoxin, fusicoccin. Z. Nat. C 1993, 48, 595–602. [Google Scholar] [CrossRef]
- Giordanetto, F.; Schäfer, A.; Ottmann, C. Stabilization of protein–protein interactions by small molecules. Drug Discov. Today 2014, 19, 1812–1821. [Google Scholar] [CrossRef] [Green Version]
- Camoni, L.; Visconti, S.; Aducci, P.; Marra, M. From plant physiology to pharmacology: Fusicoccin leaves the leaves. Planta 2018, 249, 49–57. [Google Scholar] [CrossRef]
- Fan, X.; Cui, L.; Zeng, Y.; Song, W.; Gaur, U.; Yang, M. 14-3-3 Proteins are on the crossroads of cancer, aging, and age-related neurodegenerative disease. Int. J. Mol. Sci. 2019, 20, 3518. [Google Scholar] [CrossRef] [Green Version]
- Saponaro, A.; Porro, A.; Chaves-Sanjuan, A.; Nardini, M.; Rauh, O.; Thiel, G.; Moroni, A. Fusicoccin activates KAT1 channels by stabilizing their interaction with 14-3-3 proteins. Plant Cell 2017, 29, 2570–2580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salles, I.I.; Feys, H.B.; Iserbyt, B.F.; De Meyer, S.F.; Vanhoorelbeke, K.; Deckmyn, H. Inherited traits affecting platelet function. Blood Rev. 2008, 22, 155–172. [Google Scholar] [CrossRef] [PubMed]
- Kent, C.B.; Shimada, T.; Ferraro, G.B.; Ritter, B.; Yam, P.T.; McPherson, P.S.; Charron, F.; Kennedy, T.E.; Fournier, A.E. 14-3-3 Proteins regulate protein kinase A activity to modulate growth cone turning responses. J. Neurosci. 2010, 30, 14059–14067. [Google Scholar] [CrossRef] [Green Version]
- Yam, P.; Kent, C.B.; Morin, S.; Farmer, W.T.; Alchini, R.; Lepelletier, L.; Colman, D.R.; Tessier-Lavigne, M.; Fournier, A.E.; Charron, F. 14-3-3 proteins regulate a cell-intrinsic switch from sonic hedgehog-mediated commissural axon attraction to repulsion after midline crossing. Neuron 2012, 76, 735–749. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, A.; Morquette, B.; Kroner, A.; Leong, S.; Madwar, C.; Sanz, R.; Banerjee, S.L.; Antel, J.; Bisson, N.; David, S.; et al. Small-molecule stabilization of 14-3-3 protein-protein interactions stimulates axon regeneration. Neuron 2017, 93, 1082–1093.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, A.; Ottmann, C.; Fournier, A.E. 14-3-3 adaptor protein-protein interactions as therapeutic targets for CNS diseases. Pharmacol. Res. 2017, 125, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, A.; Andrei, S.A.; Altena, A.V.R.; Simas, T.; Banerjee, S.L.; Kato, N.; Bisson, N.; Higuchi, Y.; Ottmann, C.; Fournier, A.E. Polypharmacological Perturbation of the 14-3-3 adaptor protein interactome stimulates neurite outgrowth. Cell Chem. Biol. 2020, 27, 657–667. [Google Scholar] [CrossRef]
- Honma, Y. Cotylenin A—A plant growth regulator as a differentiation-inducing agent against myeloid leukemia. Leuk. Lymphoma 2002, 43, 1169–1178. [Google Scholar] [CrossRef]
- Bury, M.; Girault, A.; Mégalizzi, V.; Spiegl-Kreinecker, S.; Mathieu, V.; Berger, W.; Evidente, A.; Kornienko, A.; Gailly, P.; Vandier, C.; et al. Ophiobolin A induces paraptosis-like cell death in human glioblastoma cells by decreasing BKCa channel activity. Cell Death Dis. 2013, 4, e561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodolfo, C.; Rocco, M.; Cattaneo, L.; Tartaglia, M.; Sassi, M.; Aducci, P.; Scaloni, A.; Camoni, L.; Marra, M. Ophiobolin A induces autophagy and activates the mitochondrial pathway of apoptosis in human melanoma cells. PLoS ONE 2016, 11, e0167672. [Google Scholar] [CrossRef] [PubMed]
- Leeuwen, I.J.D.V.-V.; Kortekaas-Thijssen, C.; Mandouckou, J.A.N.; Kas, S.; Evidente, A.; de Boer, A.H. Fusicoccin-A selectively induces apoptosis in tumor cells after interferon-α priming. Cancer Lett. 2010, 293, 198–206. [Google Scholar] [CrossRef]
- Leeuwen, I.J.D.V.-V.; Pereira, D.D.C.; Flach, K.D.; Piersma, S.R.; Haase, C.; Bier, D.; Yalcin, Z.; Michalides, R.; Feenstra, A.; Jimenez, C.R.; et al. Interaction of 14-3-3 proteins with the estrogen receptor alpha F domain provides a drug target interface. Proc. Natl. Acad. Sci. USA 2013, 110, 8894–8899. [Google Scholar] [CrossRef] [Green Version]
- Bury, M.; Andolfi, A.; Rogister, B.; Cimmino, A.; Mégalizzi, V.; Mathieu, V.; Feron, O.; Evidente, A.; Kiss, R. Fusicoccin A, a phytotoxic carbotricyclic diterpene glucoside of fungal origin, reduces proliferation and invasion of glioblastoma cells by targeting multiple tyrosine kinases. Transl. Oncol. 2013, 6, 112–123. [Google Scholar] [CrossRef] [Green Version]
- Stevers, L.M.; Lam, C.V.; Leysen, S.F.R.; Meijer, F.; van Scheppingen, D.S.; de Vries, R.; Carlile, G.W.; Milroy, L.G.; Thomas, D.; Brunsveld, L.; et al. Characterization and small-molecule stabilization of the multisite tandem binding between 14-3-3 and the R domain of CFTR. Proc. Natl. Acad. Sci. USA 2016, 113, E1152–E1161. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Honma, Y.; Miyake, T.; Adachi, K.; Takami, S.; Okada, M.; Kumanomidou, S.; Ikejiri, F.; Jo, Y.; Onishi, C.; et al. Synergistic combination therapy with cotylenin A and vincristine in multiple myeloma models. Int. J. Oncol. 2015, 46, 1801–1809. [Google Scholar] [CrossRef] [PubMed]
- Kasukabe, T.; Honma, Y.; Okabe-Kado, J. Cotylenin A, a new differentiation inducer, and rapamycin cooperatively inhibit growth of cancer cells through induction of cyclin G2. Cancer Sci. 2008, 99, 1693–1698. [Google Scholar] [CrossRef]
- Kashiwagi, Y.; Kato, N.; Sassa, T.; Nishitsuka, K.; Yamamoto, T.; Takamura, H.; Yamashita, H. Cotylenin A inhibits cell proliferation and induces apoptosis and PAX6 mRNA transcripts in retinoblastoma cell lines. Mol. Vis. 2010, 16, 970–982. [Google Scholar] [PubMed]
- Kasukabe, T.; Okabe-Kado, J.; Kato, N.; Honma, Y.; Kumakura, S. Cotylenin A and arsenic trioxide cooperatively suppress cell proliferation and cell invasion activity in human breast cancer cells. Int. J. Oncol. 2014, 46, 841–848. [Google Scholar] [CrossRef] [Green Version]
- Ikejiri, F.; Honma, Y.; Okada, T.; Urano, T.; Suzumiya, J. Cotylenin A and tyrosine kinase inhibitors synergistically inhibit the growth of chronic myeloid leukemia cells. Int. J. Oncol. 2018, 52, 2061–2068. [Google Scholar] [CrossRef] [Green Version]
- Ottmann, C.; Weyand, M.; Sassa, T.; Inoue, T.; Kato, N.; Wittinghofer, A.; Oecking, C. A Structural rationale for selective stabilization of anti-tumor interactions of 14-3-3 proteins by cotylenin A. J. Mol. Biol. 2009, 386, 913–919. [Google Scholar] [CrossRef]
- Milroy, L.-G.; Brunsveld, L.; Ottmann, C. Stabilization and inhibition of protein–protein interactions: The 14-3-3 case study. ACS Chem. Biol. 2012, 8, 27–35. [Google Scholar] [CrossRef]
- Molzan, M.; Kasper, S.; Röglin, L.; Skwarczynska, M.; Sassa, T.; Inoue, T.; Breitenbuecher, F.; Ohkanda, J.; Kato, N.; Schuler, M.; et al. Stabilization of physical RAF/14-3-3 interaction by cotylenin A as treatment strategy for RAS mutant cancers. ACS Chem. Biol. 2013, 8, 1869–1875. [Google Scholar] [CrossRef] [PubMed]
- Ohkanda, J.; Kusumoto, A.; Punzalan, L.; Masuda, R.; Wang, C.; Parvatkar, P.; Akase, D.; Aida, M.; Uesugi, M.; Higuchi, Y.; et al. Structural effects of fusicoccin upon upregulation of 14-3-3-phospholigand interaction and cytotoxic activity. Chem. A Eur. J. 2018, 24, 16066–16071. [Google Scholar] [CrossRef]
- Kawakami, K.; Hattori, M.; Inoue, T.; Maruyama, Y.; Ohkanda, J.; Kato, N.; Tongu, M.; Yamada, T.; Akimoto, M.; Takenaga, K.; et al. A novel fusicoccin derivative preferentially targets hypoxic tumor cells and inhibits tumor growth in xenografts. Anti-Cancer Agents Med. Chem. 2012, 12, 791–800. [Google Scholar] [CrossRef]
- Miyake, T.; Honma, Y.; Urano, T.; Kato, N.; Suzumiya, J. Combined treatment with tamoxifen and a fusicoccin derivative (ISIR-042) to overcome resistance to therapy and to enhance the antitumor activity of 5-fluorouracil and gemcitabine in pancreatic cancer cells. Int. J. Oncol. 2015, 47, 315–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, C.; Higuchi, Y.; Koschinsky, K.; Bartel, M.; Schumacher, B.; Thiel, P.; Nitta, H.; Preisig-Müller, R.; Schlichthörl, G.; Renigunta, V.; et al. A Semisynthetic fusicoccane stabilizes a protein-protein interaction and enhances the expression of K+ channels at the cell surface. Chem. Biol. 2013, 20, 583–593. [Google Scholar] [CrossRef] [Green Version]
- Stevers, L.M.; Sijbesma, E.; Botta, M.; MacKintosh, C.; Obsil, T.; Landrieu, I.; Cau, Y.; Wilson, A.J.; Karawajczyk, A.; Eickhoff, J.; et al. Modulators of 14-3-3 Protein–protein interactions. J. Med. Chem. 2017, 61, 3755–3778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maki, T.; Kawamura, A.; Kato, N.; Ohkanda, J. Chemical ligation of epoxide-containing fusicoccins and peptide fragments guided by 14-3-3 protein. Mol. BioSyst. 2012, 9, 940–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parvatkar, P.; Kato, N.; Uesugi, M.; Sato, S.-I.; Ohkanda, J. Intracellular generation of a diterpene-peptide conjugate that inhibits 14-3-3-mediated interactions. J. Am. Chem. Soc. 2015, 137, 15624–15627. [Google Scholar] [CrossRef]
- Masuda, R.; Kawasaki, Y.; Igawa, K.; Manabe, Y.; Fujii, H.; Kato, N.; Tomooka, K.; Ohkanda, J. Copper-Free Huisgen Cycloaddition for the 14-3-3-templated synthesis of fusicoccin-peptide conjugates. Chem. Asian J. 2020, 15, 742–747. [Google Scholar] [CrossRef] [Green Version]
- Bayer, K.U.; Schulman, H. CaM Kinase: Still inspiring at 40. Neuron 2019, 103, 380–394. [Google Scholar] [CrossRef]
- Russell, F.M.; Hardie, D.G. AMP-Activated Protein Kinase: Do we need activators or inhibitors to treat or prevent cancer? Int. J. Mol. Sci. 2020, 22, 186. [Google Scholar] [CrossRef] [PubMed]
- Santo, D.L.; Petrvalska, O.; Obsilova, V.; Ottmann, C.; Obsil, T. Stabilization of protein–protein interactions between CaMKK2 and 14–3–3 by fusicoccins. ACS Chem. Biol. 2020, 15, 3060–3071. [Google Scholar] [CrossRef]
- Sengupta, A.; Liriano, J.; Bienkiewicz, E.A.; Miller, B.G.; Frederich, J.H. Probing the 14-3-3 isoform-specificity profile of protein–protein interactions stabilized by fusicoccin A. ACS Omega 2020, 5, 25029–25035. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fusicoccanes | Relation to FC | Biological Activity | References |
|---|---|---|---|
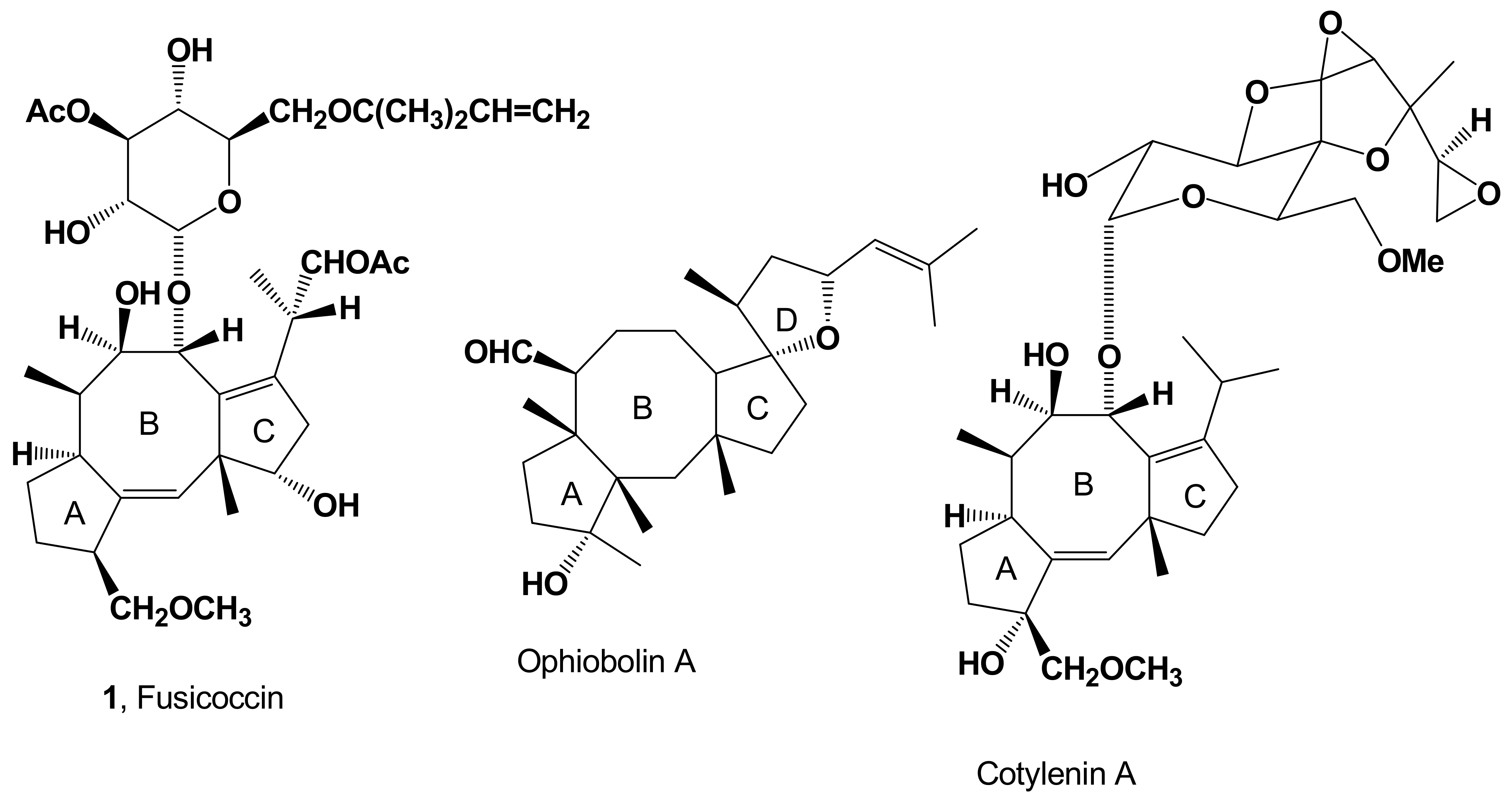
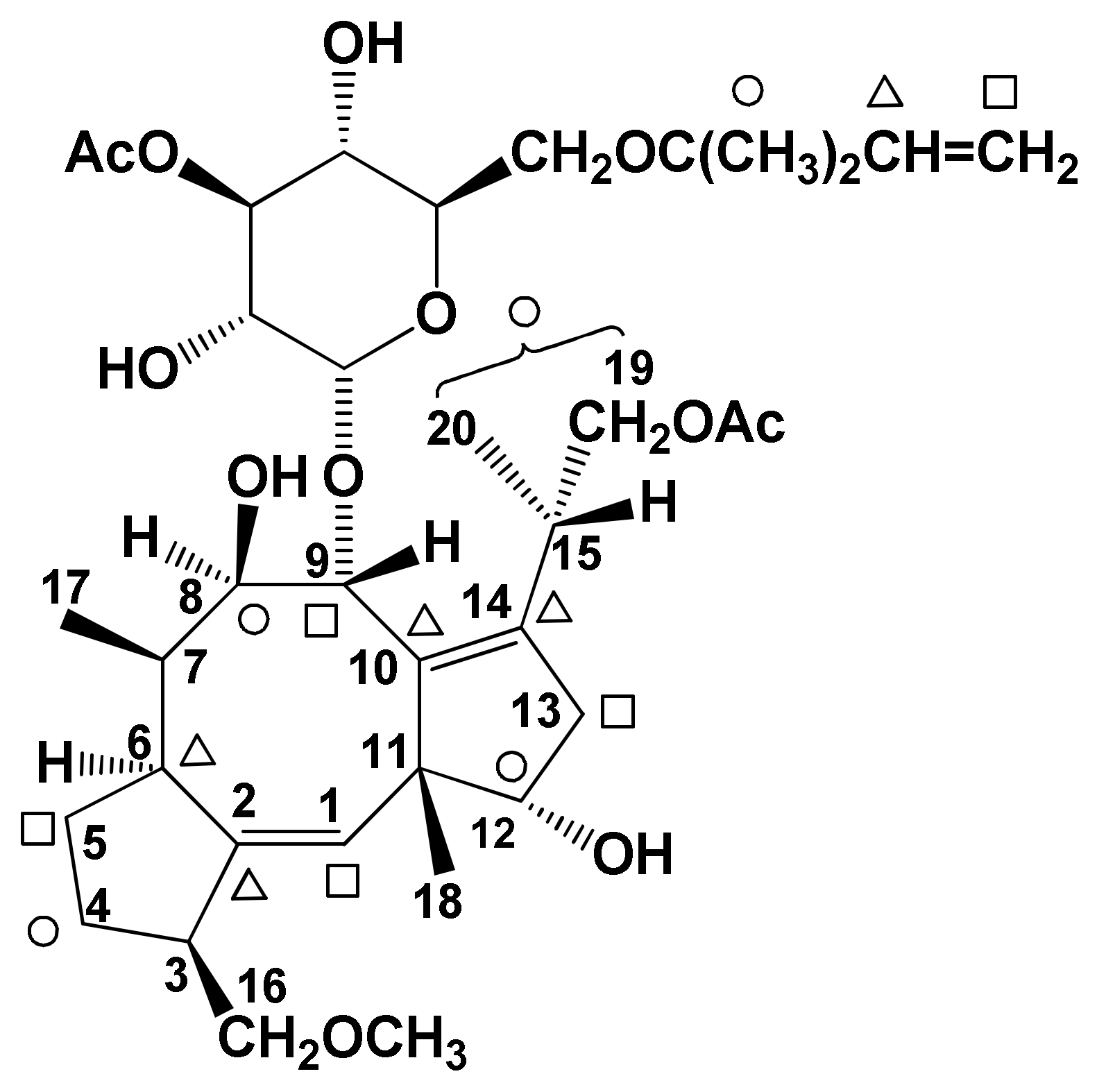
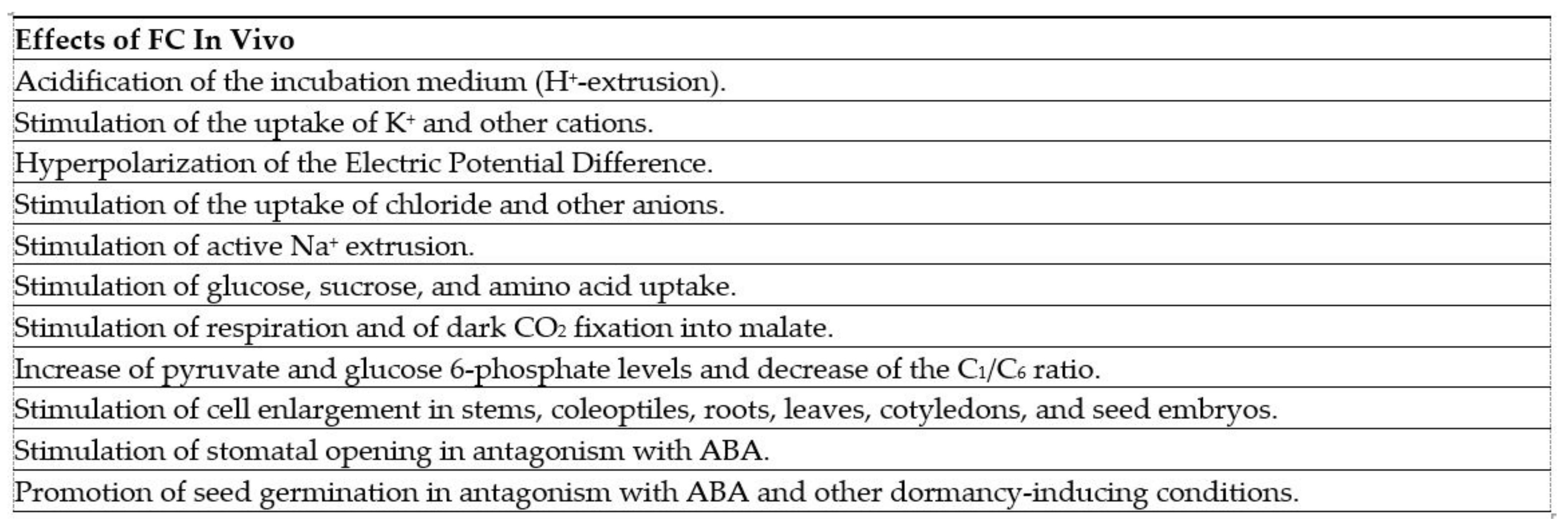
| FC (1, Figure 1) | Phytotoxicity | [7,8,9,10,11,12] | |
| Induction of abscission | [13,14] | ||
| Proton extrusion, potassium uptake and stomatal opening | [15,16] | ||
| Cell enlargement, proton extrusion, cotyledon growth and seed germination | [17] | ||
| Stimulation of seed germination | [18] | ||
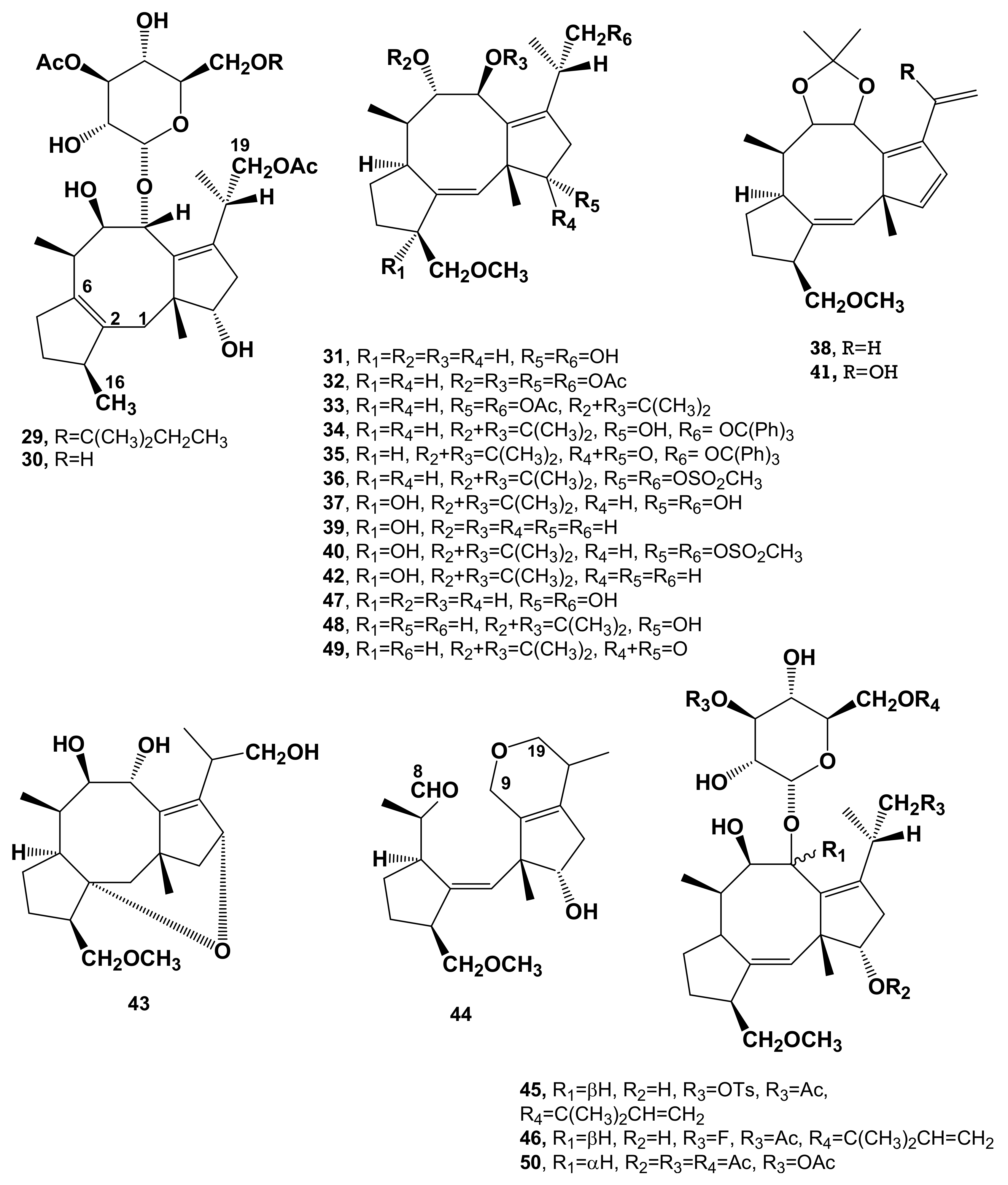
| MonodeacetylFC (2, Figure 2) | MFRM | Reduced phytotoxicity | [2,9,10] |
| Induction of abscission | [13,14] | ||
| DideacetylFC (3, Figure 2) | MFRM | Reduced Phytotoxicity | [2,9,10] |
| Cell enlargement, proton extrusion, cotyledon growth and seed germination | [17] | ||
| Induction of abscission | [13,14] | ||
| Stimulation of O. ramosa seed germination | [19] | ||
| IsoFC (4, Figure 2) | MFRM | NT | [2,9,20] |
| Induction of abscission | [13,14] | ||
| AlloFC (5, Figure 2) | MFRM | NT | [2,9,20] |
| MonodeacetylalloFC (6, Figure 2) | MFRM | NT | [21] |
| MonodeacetylisoFC (7, Figure 2) | MFRM | NT | [21] |
| 12-AcetyldideacetylFC (8, Figure 2) | MFRM | NT | [21] |
| Stimulation of O. ramosa seed germination | [19] | ||
| 19-DeoxydideacetylFC (9, Figure 2) | MFRM | Cell enlargement, proton extrusion, cotyledon growth and seed germination | [17,22,23] |
| Stimulation of O. ramosa seed germination | [19] | ||
| 3-α-Hydroxy-19-deoxydideacetylFC (10, Figure 2) | MFRM | Cell enlargement, proton extrusion, cotyledon growth and seed germination | [17,24] |
| Stimulation of O. ramosa seed germination | [19] | ||
| 3-α−HydroxydideacetylFC (11, Figure 2) | MFRM | Cell enlargement, proton extrusion, cotyledon growth and seed germination | [17,25,26] |
| Stimulation of O. ramosa seed germination | [19] | ||
| 16-O-demethyl-19-deoxydideacetyl-3-epi-FC (12, Figure 2) | MFRM | Reduced phytotoxicity | [10,27,28] |
| Cell enlargement, proton extrusion, cotyledon growth and seed germination | [17] | ||
| FC-H (13, Figure 2) | MFRM | Reduced phytotoxicity | [23] |
| Cell enlargement, proton extrusion, cotyledon growth and seed germination | [10] | ||
| Stimulation of O. ramosa seed germination | [19] | ||
| DihydroFC (14, Figure 2) | SFD | Phytotoxicity | [10,12,29] |
| Induction of abscission | [12,13] | ||
| TriacetylFC (15, Figure 2) | SFD | NA | [10,29] |
| De-t-pentenyltetracetylFC (16, Figure 2) | SFD | Reduced phytotoxicity | [10,12,29] |
| Stimulation of O. ramosa seed germination | [19] | ||
| De-t-pentenylFC, (17, Figure 2) | SFD | Reduced phytotoxicity | [10,12,29] |
| Induction of abscission | [13,14] | ||
| Stimulation of O. ramosa seed germination | [19] | ||
| IsodihydroFC (18, Figure 2) | SFD | NA | [10,12,29] |
| MonodeacetyldihydroFC (19, Figure 2) | SFD | Reduced phytoxicity | [10,29] |
| Induction of abscission | [11,12] | ||
| DihydrodidacetylFC (20, Figure 2) | SFD | Phytotoxicity | [10,29] |
| Induction of abscission | [13,14] | ||
| Stimulation of O. ramosa seed germination | [19] | ||
| TriacetyldihydroFC (21, Figure 2) | SFD | No phytotoxicity | [10,29] |
| Dideacetyl-de-t-pentenylFC (22, Figure 2) | SFD | Reduced phytotoxicity | [10,29] |
| Triacetyl-de-t-pentenylFC (23, Figure 2) | SFD | NA | [10,29] |
| 8-Oxo-triacetylFC (24, Figure 2) | SFD | NT | [13,29] |
| PseudoacetonideFC (25 Figure 2) | SFD | NT | [29,30] |
| Stimulation of O. ramosa seed germination | [19] | ||
| 8-oxo-9-epi-dideacetylFC (26, Figure 2) | SFD | NT | [17] |
| Derivative 27 (Figure 2) | SFD | NT | [10,31] |
| Stimulation of O. ramosa seed germination | [19] | ||
| Derivative 28 (Figure 2) | SFD | NT | [10,32] |
| Stimulation of O. ramosa seed germination | [19] | ||
| PerhydroFC (29, Figure 3) | SFD | NT | [33] |
| Stimulation of O. ramosa seed germination | [19] | ||
| De-t-pentenylperhydroFC (30, Figure 3) | SFD | NT | [33] |
| DeacetylaglyconeFC (32, Figure 3) | SFD | Reduced phytotoxicity | [7,10] |
| Stimulation of seed germination | [18] | ||
| Stimulation of O. ramosa seed germination | [19] | ||
| TetracetyldeacetylaglyconeFC (32, Figure 3) | SFD | No phytotoxicity | [7,10,34] |
| Cell enlargement, proton extrusion, cotyledon growth and seed germination | [13] | ||
| 8,9-IsopropylidenedeacetylaglyconeFC= 8,9-acetonideacetylglyconeFC (33, Figure 3) | SFD | NT | [7,34] |
| No phytotoxicity | [10] | ||
| Stimulation of O. ramosa seed germination | [19] | ||
| 19-Trytil-8,9-cetonidedeacetylaglyconeFC (34, Figure 3) | SFD | No phytotoxicity | [10] |
| Stimulation of O. ramosa seed germination | [19] | ||
| 12-Oxo-19-trytil-8,9-actonidedeacetylaglyconeFC (35, Figure 3) | SFD | NT | [7,34] |
| 12,19-Dimesyla-8,9-cetonidedeacetylagllyconeFC (36, Figure 3) | SFD | NT | [7,34,35] |
| 3-α-Hydroxy-8,9-acetonidedacetylaglycone FC (37, Figure 3) | SFD | NT | [26] |
| Tetraene of 8,9-acetonidedeacetylaglyconeFC (38, Figure 3) | SFD | NT | [35] |
| Cotylenol (39, Figure 3) | CM | Cell enlargement, proton extrusion, cotyledon growth and seed germination | [17,36,37,38] |
| Stimulation of seed germination | [18] | ||
| 3α-Hydroxy-12,19-dimesyl-8,9-acetonidedeacetylaglyconeFC (40, Figure 3) | SFD | NT | [39] |
| 20-Hydroxytetrahene (41, Figure 3) | SFD | NT | [39] |
| 8,9-Acetonide cotylenol (42, Figure 3) | SFD | NT | [39] |
| Isomer of deacetylaglycone of FC (43, Figure 3) | SFD | NT | [40] |
| Stimulation of O. ramosa seed germination | [19] | ||
| MonoaldehydedihydropyrandeacetylaglyconeFC (44, Figure 3) | SFD | NT | [34] |
| 19-TosyldideactylFC (45, Figure 3) | SFD | NT | [19] |
| 19-Fluoro-19-dehydroxydideacetylFC (46, Figure 3) | SFD | Stimulation of O. ramosa seed germination | [19] |
| AglyconeFC (47, Figure 3) | SFD | Stimulation of O. ramosa seed germination | [19] |
| 19-Deoxy-12-epi-8,9-acetonide of deacetylaglycone FC (48, Figure 3) | SFD | Stimulation of O. ramosa seed germination | [19] |
| 19-Deoxy-12-oxo-8,9-acetonide of deacetylaglycone FC (49, Figure 3) | SFD | Stimulation of O. ramosa seed germination | [19] |
| De-t-pentenyl-9-epi-tetracetylFC (50, Figure 3) | SFD | No phytotoxicity | [10] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marra, M.; Camoni, L.; Visconti, S.; Fiorillo, A.; Evidente, A. The Surprising Story of Fusicoccin: A Wilt-Inducing Phytotoxin, a Tool in Plant Physiology and a 14-3-3-Targeted Drug. Biomolecules 2021, 11, 1393. https://doi.org/10.3390/biom11091393
Marra M, Camoni L, Visconti S, Fiorillo A, Evidente A. The Surprising Story of Fusicoccin: A Wilt-Inducing Phytotoxin, a Tool in Plant Physiology and a 14-3-3-Targeted Drug. Biomolecules. 2021; 11(9):1393. https://doi.org/10.3390/biom11091393
Chicago/Turabian StyleMarra, Mauro, Lorenzo Camoni, Sabina Visconti, Anna Fiorillo, and Antonio Evidente. 2021. "The Surprising Story of Fusicoccin: A Wilt-Inducing Phytotoxin, a Tool in Plant Physiology and a 14-3-3-Targeted Drug" Biomolecules 11, no. 9: 1393. https://doi.org/10.3390/biom11091393