
The Carbon Source Controls the Secretion and Yield of Polysaccharide-Hydrolyzing Enzymes of Basidiomycetes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Organisms and Inoculum Preparation
2.2. Cultivation Conditions
2.3. Cultivation in a Fermenter
2.4. Wheat Straw Saccharification
2.5. Analytical Methods
2.6. Statistical Analysis
3. Results and Discussion
3.1. Effect of the Polymeric Carbon Sources on Basidiomycetes Enzyme Activity
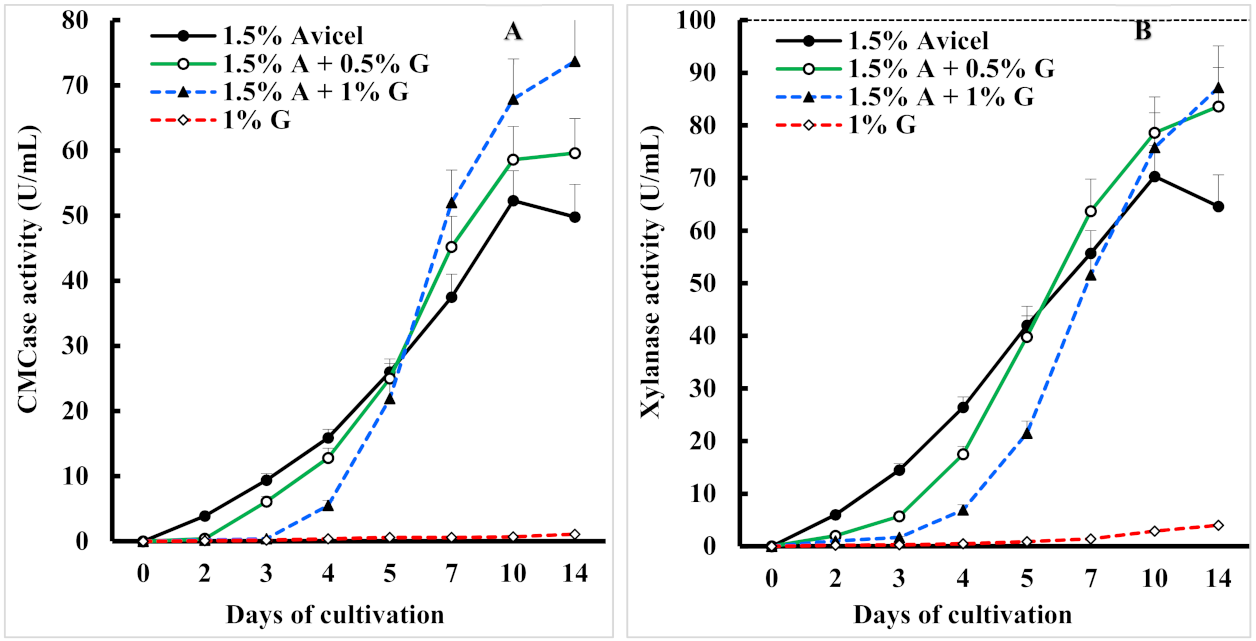
3.2. Effect of Glycerol as an Additional Easily Metabolizable Carbon Source
3.3. Effect of the Type of Carbon Source in the Inoculum Preparation Medium
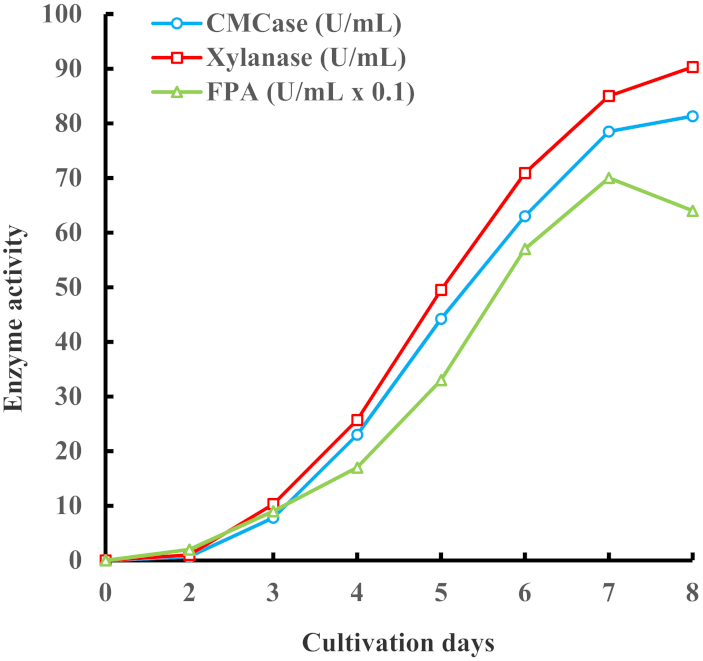
3.4. Scaled Up Enzyme Production in I. lacteus Cultivation in a Fermenter
3.5. Saccharification of Pretreated Wheat Straw with an Enzyme Preparation from I. lacteus
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Phitsuwan, P.; Laohakunjit, N.; Kerdchoechuen, O.; Kyu, K.L.; Ratanakhanokchai, K. Present and potential applications of cellulases in agriculture, biotechnology, and bioenergy. Folia Microbiol. 2013, 58, 163–176. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Microbial cellulases: Engineering, production and applications. Renew. Sustain. Energy Rev. 2014, 33, 188–203. [Google Scholar] [CrossRef]
- Liu, L.; Huang, W.C.; Liu, Y.; Li, M. Diversity of cellulolytic microorganisms and microbial cellulases. Int. Biodeterior. Biodegrad. 2021, 163. [Google Scholar] [CrossRef]
- Rytioja, J.; Hildén, K.; Yuzon, J.; Hatakka, A.; de Vries, R.P.; Mäkelä, M.R. Plant-polysaccharide-degrading enzymes from basidiomycetes. Microbiol. Mol. Biol. Rev. 2014, 7, 614–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentil, J.A. Biocatalytic potential of basidiomycetes: Relevance, challenges and research interventions in industrial processes. Sci. Afr. 2021, 11, e00717. [Google Scholar] [CrossRef]
- Agnihotri, S.; Dutt, D.; Tyagi, C.H.; Kumar, A.; Upadhyaya, J.S. Production and biochemical characterization of a novel cellulase-poor alkali-thermo-tolerant xylanase from Coprinellus disseminatus SW-1 NTCC 1165. World J. Microbiol. Biotechnol. 2010, 26, 1349–1359. [Google Scholar] [CrossRef]
- Jagtap, S.S.; Dhiman, S.S.; Kim, T.-S.; Kim, I.-W.; Lee, J.-K. Characterization of a novel endo-β-1,4-glucanase from Armillaria gemina and its application in biomass hydrolysis. Appl. Microbiol. Biotechnol. 2014, 98, 661–669. [Google Scholar] [CrossRef]
- Jagtap, S.S.; Dhiman, S.S.; Kim, T.-S.; Li, L.; Kang, Y.C.; Lee, J.-K. Characterization of a β-1,4-glucosidase from a newly isolated strain of Pholiota adiposa and its application to the hydrolysis of biomass. Biomass Bioenergy. 2013, 54, 181–190. [Google Scholar] [CrossRef]
- Boiko, S.M. Cellulases of basidiomycetes for the development of bioconversion technologies. Ukr. Bot. J. 2020, 77, 378–385. [Google Scholar] [CrossRef]
- Metreveli, E.; Kachlishvili, E.; Singer, S.W.; Elisashvili, V. Alteration of white-rot basidiomycetes cellulase and xylanase activities in the submerged co-cultivation and optimization of enzyme production by Irpex lacteus and Schizophyllum commune. Bioresour. Technol. 2017, 241, 652–660. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Ghose, T.K. Measurement of cellulase activities. Pure Appl. Chem. 1987, 59, 257–268. [Google Scholar] [CrossRef]
- Bailey, M.J.; Biely, P.; Poutanen, K. Interlaboratory testing of methods for assay of xylanase activity. J. Biotechnol. 1992, 23, 257–270. [Google Scholar] [CrossRef]
- Poutanen, K.; Pulls, J. Characteristics of Trichoderma reesei β-xylosidase and its use in hydrolysis of solubilized xylans. Appl. Microbiol. Biotechnol. 1988, 28, 425–432. [Google Scholar] [CrossRef]
- Salmon, D.N.X.; Spier, M.R.; Soccol, C.R.; Vandenberghe, L.P.S.; Weingartner Montibeller, V.; Bier, M.C.J.; Faraco, V. Analysis of inducers of xylanase and cellulase activities production by Ganoderma applanatum LPB MR-56. Fungal Biol. 2014, 118, 655–662. [Google Scholar] [CrossRef] [Green Version]
- Kobakhidze, A.; Asatiani, M.; Kachlishvili, E.; Elisashvili, V. Induction and catabolite repression of cellulase and xylanases synthesis in the selected white-rot Basidiomycetes. Ann. Agric. Sci. 2016, 14, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Sornlake, W.; Rattanaphanjak, P.; Champreda, V.; Eurwilaichitr, L.; Kittisenachai, S.; Roytrakul, S.; Fujii, T.; Inoue, H. Characterization of cellulolytic enzyme system of Schizophyllum commune mutant and evaluation of its efficiency on biomass hydrolysis. Biosci. Biotechnol. Biochem. 2017, 81, 1289–1299. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.; Su, X.; Luo, H.; Ma, R.; Yao, B.; Ma, F. Deciphering lignocellulose deconstruction by the white rot fungus Irpex lacteus based on genomic and transcriptomic analyses. Biotechnol. Biofuels 2018, 11, 58. [Google Scholar] [CrossRef] [Green Version]
- Mezule, L.; Civzele, A. Bioprospecting white-rot basidiomycete Irpex lacteus for improved extraction of lignocellulose-degrading enzymes and their further application. J. Fungi 2020, 6, 256. [Google Scholar] [CrossRef]
- Kachlishvili, E.; Kobakhidze, A.; Rusitashvili, M.; Tsokilauri, A.; Elisashvili, V. Elucidation of the higher basidiomycetes’ enzyme activity dependence on the mushroom inoculum form, pre-cultivation medium, age, and size. Int. J. Med. Mushrooms 2020, 22, 1099–1108. [Google Scholar] [CrossRef]
- Manavalan, T.; Manavalan, A.; Thangavelu, K.P.; Heese, K. Characterization of a novel endoglucanase from Ganoderma lucidum. J. Basic Microbiol. 2015, 54, 761–771. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Growth Substrate | Final pH | CMCase (U/mL) | Xylanase (U/mL) | FPA (U/mL) | β-Glucosidase (U/mL) | β-Xylosidase (U/mL) |
|---|---|---|---|---|---|---|
| P. coccineus | ||||||
| 1.5% Avicel | 5.9 ± 0.1 | 63 ± 6.011 * | 31 ± 3.814 * | 4.6 ± 0.514 * | 3.6 ± 0.311 * | 0.06 ± 0.0111 * |
| 4% mandarin squeeze (MS) | 6.7 ± 0.1 | 19 ± 2.111 | 14 ± 1.18 | 2.2 ± 0.28 | 2.2 ± 0.28 | 0.05 ± 06 |
| 4% wheat straw (WS) | 6.2 ± 0.1 | 22 ± 1.411 | 18 ± 1.211 | 1.9 ± 0.28 | 1.5 ± 0.26 | 0.04 ± 06 |
| 1.5% Avicel + 4% MS | 5.6 ± 0.1 | 82 ± 8.314 | 65 ± 5.811 | 5.7 ± 0.611 | 3.3 ± 0.38 | 0.12 ± 0.0111 |
| 4% WS +4% MS | 5.9 ± 0.1 | 27 ± 2.411 | 19 ± 1.711 | 2.2 ± 0.311 | 1.6 ± 0.26 | 0.05 ± 0.016 |
| S. commune | ||||||
| 1.5% Avicel | 6.1 ± 0.1 | 39 ± 4.314 | 626 ± 7814 | 2.1 ± 0.311 | 10.7 ± 0.814 | 0.12 ± 0.0114 |
| 4% MS | 6.5 ± 0.1 | 23 ± 3.011 | 531 ± 968 | 3.0 ± 0.311 | 10.1 ± 0.914 | 0.15 ± 0.0214 |
| 4% WS | 6.8 ± 0.1 | 8 ± 1.014 | 120 ± 1014 | 1.3 ± 0.114 | 3.8 ± 0.314 | 0.07 ± 0.0114 |
| 1.5% Avicel + 4% MS | 6.3 ± 0.1 | 39 ± 2.911 | 740 ± 8014 | 4.2 ± 0.414 | 18.6 ± 2.014 | 0.31 ± 0.0314 |
| 4% WS + 4% MS | 5.9 ± 0.1 | 14 ± 1.311 | 528 ± 596 | 2.2 ± 0.214 | 5.9 ± 0.414 | 0.07 ± 0.0111 |
| I. lacteus | ||||||
| 1.5% Avicel | 5.4 ± 0.1 | 54 ± 6.611 | 67 ± 8.111 | 5.1 ± 0.411 | 1.4 ± 0.1211 | 0.02 ± 06 |
| 4% MS | 6.9 ± 0.1 | 18 ± 1.48 | 20 ± 1.711 | 1.9 ± 0.28 | 1.1 ± 0.128 | 0.02 ± 06 |
| 4% WS | 6.2 ± 0 | 23 ± 2.011 | 29 ± 1.48 | 2.3 ± 0.211 | 1.0 ± 0.118 | 0.05 ± 0.018 |
| 1.5% Avicel + 4% MS | 5.9 ± 0.1 | 76 ± 9.011 | 106 ± 8.614 | 6.8 ± 0.511 | 2.3 ± 0.278 | 0.08 ± 0.018 |
| 4% WS + 4% MS | 6.3 ± 0.2 | 40 ± 4.78 | 34 ± 2.18 | 2.5 ± 0.38 | 1.6 ± 0.198 | 0.05 ± 0.016 |
| Growth Substrate | Final pH | CMCase (U/mL) | Xylanase (U/mL) | β-Glucosidase (U/mL) |
|---|---|---|---|---|
| The inoculum was grown in the presence of glucose | ||||
| Avicel | 5.4 ± 0.1 | 52.6 ± 6.111 * | 71.0 ± 6.111 * | 1.65 ± 0.208 * |
| Mandarin squeeze | 6.9 ± 0.1 | 11.0 ± 0.98 | 23.8 ± 2.214 | 0.87 ± 0.068 |
| Wheat bran | 5.5 ± 0.1 | 17.4 ± 1.611 | 45.2 ± 5.611 | 1.18 ± 0.168 |
| Wheat straw | 5.7 ± 0.1 | 29.4 ± 3.911 | 39.1 ± 3.411 | 1.34 ± 0.148 |
| Beech sawdust | 5.7 ± 0.1 | 3.8 ± 0.211 | 10.3 ± 1.214 | 0.21 ± 0.0311 |
| The inoculum was grown in the presence of Avicel | ||||
| Avicel | 5.7 ± 0.1 | 72.6 ± 8.06 | 106.5 ± 9.411 | 2.84 ± 0.218 |
| Mandarin squeeze | 7.0 ± 0.1 | 14.2 ± 1.16 | 24.5 ± 1.98 | 0.82 ± 0.0611 |
| Wheat bran | 6.0 ± 0.1 | 24.5 ± 2.011 | 67.2 ± 6.011 | 1.46 ± 0.1611 |
| Wheat straw | 5.5 ± 0.1 | 46.5 ± 3.46 | 63.2 ± 7.18 | 2.08 ± 0.178 |
| Beech sawdust | 6.1 ± 0.1 | 4.3 ± 0.46 | 8.2 ± 1.18 | 0.38 ± 0.0311 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Metreveli, E.; Khardziani, T.; Elisashvili, V. The Carbon Source Controls the Secretion and Yield of Polysaccharide-Hydrolyzing Enzymes of Basidiomycetes. Biomolecules 2021, 11, 1341. https://doi.org/10.3390/biom11091341
Metreveli E, Khardziani T, Elisashvili V. The Carbon Source Controls the Secretion and Yield of Polysaccharide-Hydrolyzing Enzymes of Basidiomycetes. Biomolecules. 2021; 11(9):1341. https://doi.org/10.3390/biom11091341
Chicago/Turabian StyleMetreveli, Eka, Tamar Khardziani, and Vladimir Elisashvili. 2021. "The Carbon Source Controls the Secretion and Yield of Polysaccharide-Hydrolyzing Enzymes of Basidiomycetes" Biomolecules 11, no. 9: 1341. https://doi.org/10.3390/biom11091341