
Transport and Secretion of the Wnt3 Ligand by Motor Neuron-like Cells and Developing Motor Neurons

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chick Embryos
2.2. Plasmids, Injection, and In Ovo Electroporation
2.3. Cell Culture
2.4. Transient Transfection
2.5. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
2.6. Western Blot Analysis
2.7. Immunohistochemistry, Immunofluorescence Microscopy, and Image Analyses
2.8. Statistical Analyses
3. Results
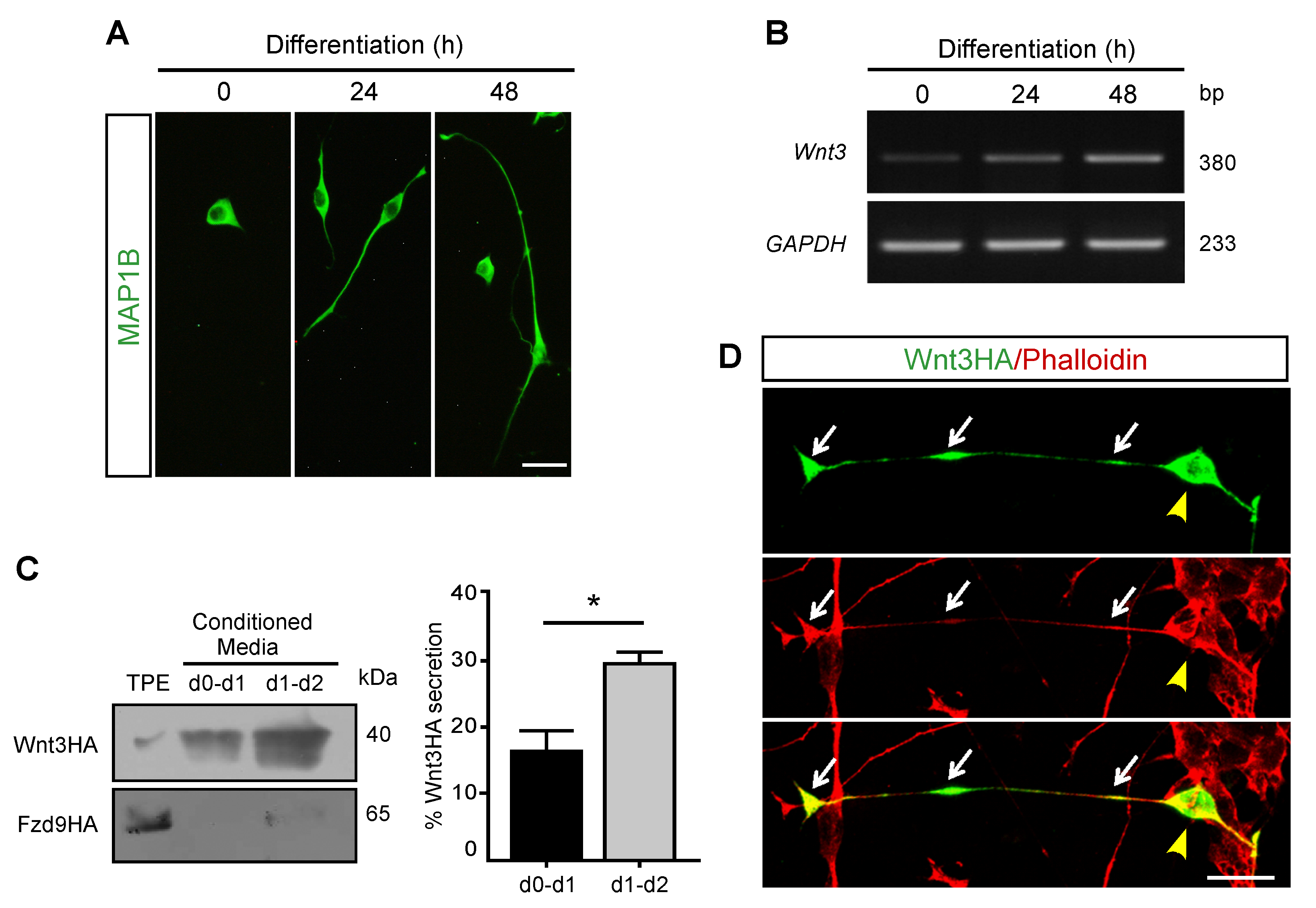
3.1. Wnt3 Is Expressed and Secreted by Differentiating Motor Neuron-like Cells
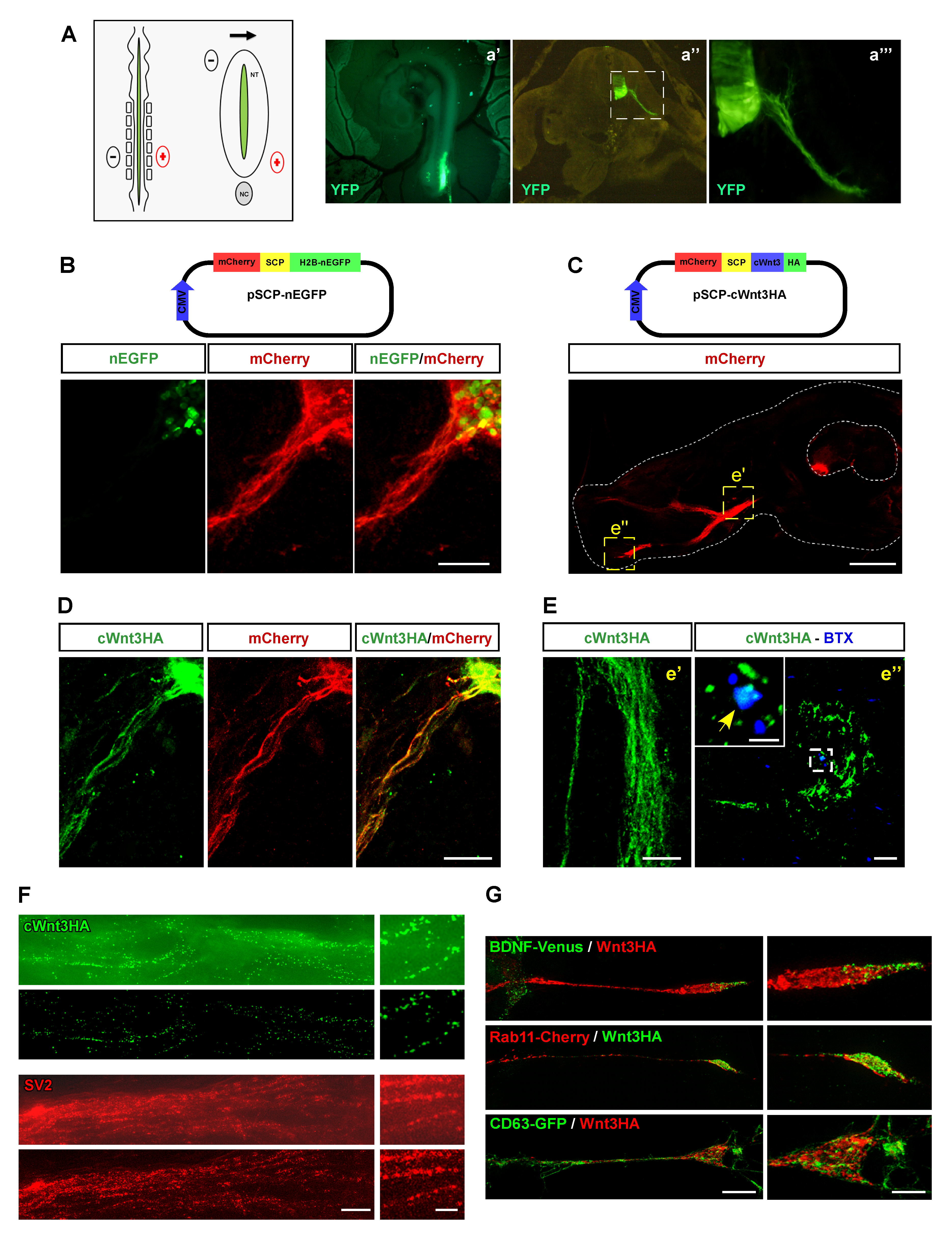
3.2. Overexpressed Wnt3 Is Transported by Motor Neurons in a Vesicular-like Pattern
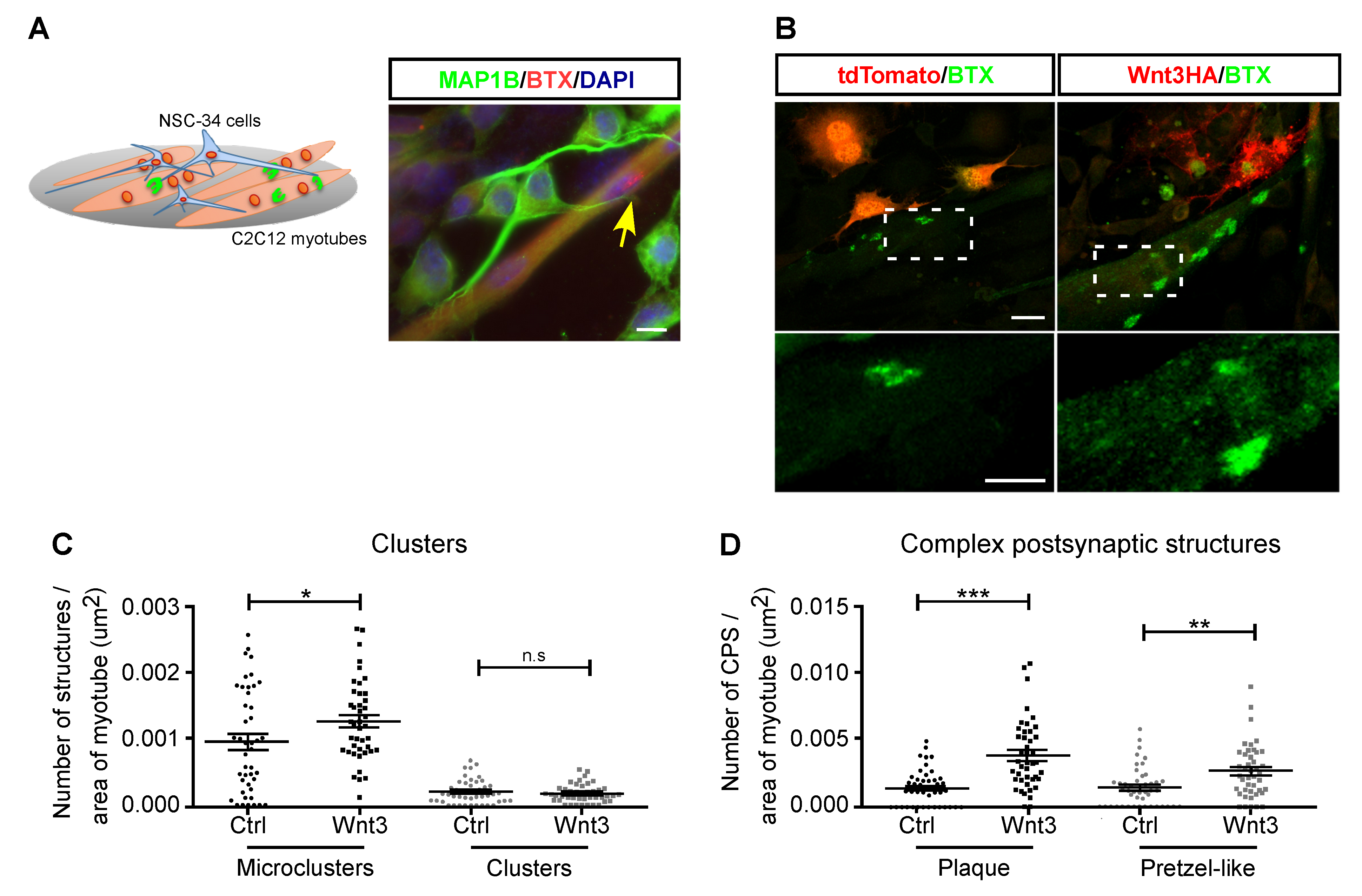
3.3. Motor Neuron-Secreted Wnt3 Induces Postsynaptic Assembly
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanes, J.R.; Lichtman, J.W. Induction, assembly, maturation and maintenance of a postsynaptic apparatus. Nat. Rev. Neurosci. 2001, 2, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Tintignac, L.A.; Brenner, H.R.; Ruegg, M.A. Mechanisms Regulating Neuromuscular Junction Development and Function and Causes of Muscle Wasting. Physiol. Rev. 2015, 95, 809–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeChiara, T.M.; Bowen, D.C.; Valenzuela, D.M.; Simmons, M.V.; Poueymirou, W.T.; Thomas, S.; Kinetz, E.; Compton, D.L.; Rojas, E.; Park, J.S.; et al. The receptor tyrosine kinase MuSK is required for neuromuscular junction formation in vivo. Cell 1996, 85, 501–512. [Google Scholar] [CrossRef] [Green Version]
- Nusse, R.; Clevers, H. Wnt/β-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef]
- Jing, L.; Lefebvre, J.L.; Gordon, L.R.; Granato, M. Wnt signals organize synaptic prepattern and axon guidance through the zebrafish unplugged/MuSK receptor. Neuron 2009, 61, 721–733. [Google Scholar] [CrossRef] [Green Version]
- Gordon, L.R.; Gribble, K.D.; Syrett, C.M.; Granato, M. Initiation of synapse formation by Wnt-induced MuSK endocytosis. Development 2012, 139, 1023–1033. [Google Scholar] [CrossRef] [Green Version]
- Henriquez, J.P.; Webb, A.; Bence, M.; Bildsoe, H.; Sahores, M.; Hughes, S.M.; Salinas, P.C. Wnt signaling promotes AChR aggregation at the neuromuscular synapse in collaboration with agrin. Proc. Natl. Acad. Sci. USA 2008, 105, 18812–18817. [Google Scholar] [CrossRef] [Green Version]
- Messeant, J.; Ezan, J.; Delers, P.; Glebov, K.; Marchiol, C.; Lager, F.; Renault, G.; Tissir, F.; Montcouquiol, M.; Sans, N.; et al. Wnt proteins contribute to neuromuscular junction formation through distinct signaling pathways. Development 2017, 144, 1712–1724. [Google Scholar]
- Shen, C.; Li, L.; Zhao, K.; Bai, L.; Wang, A.; Shu, X.; Xiao, Y.; Zhang, J.; Zhang, K.; Hui, T.; et al. Motoneuron Wnts regulate neuromuscular junction development. Elife 2018, 7, e34625. [Google Scholar] [CrossRef]
- Strochlic, L.; Falk, J.; Goillot, E.; Sigoillot, S.; Bourgeois, F.; Delers, P.; Rouviere, J.; Swain, A.; Castellani, V.; Schaeffer, L.; et al. Wnt4 participates in the formation of vertebrate neuromuscular junction. PLoS ONE 2012, 7, e29976. [Google Scholar] [CrossRef]
- Wang, J.; Ruan, N.J.; Qian, L.; Lei, W.L.; Chen, F.; Luo, Z.G. Wnt/beta-catenin signaling suppresses Rapsyn expression and inhibits acetylcholine receptor clustering at the neuromuscular junction. J. Biol. Chem. 2008, 283, 21668–21675. [Google Scholar] [CrossRef] [Green Version]
- Krylova, O.; Herreros, J.; Cleverley, K.E.; Ehler, E.; Henriquez, J.P.; Hughes, S.M.; Salinas, P.C. WNT-3, expressed by motoneurons, regulates terminal arborization of neurotrophin-3-responsive spinal sensory neurons. Neuron 2002, 35, 1043–1056. [Google Scholar] [CrossRef] [Green Version]
- Hamburger, V.; Hamilton, H.L. A series of normal stages in the development of the chick embryo. 1951. Dev. Dyn. 1992, 195, 231–272. [Google Scholar] [CrossRef]
- Szymczak, A.L.; Workman, C.J.; Wang, Y.; Vignali, K.M.; Dilioglou, S.; Vanin, E.F.; Vignali, D.A. Correction of multi-gene deficiency in vivo using a single ’self-cleaving’ 2A peptide-based retroviral vector. Nat. Biotechnol. 2004, 22, 589–594. [Google Scholar] [CrossRef]
- Escudero, C.A.; Lazo, O.M.; Galleguillos, C.; Parraguez, J.I.; Lopez-Verrilli, M.A.; Cabeza, C.; Leon, L.; Saeed, U.; Retamal, C.; Gonzalez, A.; et al. The p75 neurotrophin receptor evades the endolysosomal route in neuronal cells, favouring multivesicular bodies specialised for exosomal release. J. Cell Sci. 2014, 127, 1966–1979. [Google Scholar] [CrossRef] [Green Version]
- Krull, C.E. A primer on using in ovo electroporation to analyze gene function. Dev. Dyn. 2004, 229, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Cashman, N.R.; Durham, H.D.; Blusztajn, J.K.; Oda, K.; Tabira, T.; Shaw, I.T.; Dahrouge, S.; Antel, J.P. Neuroblastoma x spinal cord (NSC) hybrid cell lines resemble developing motor neurons. Dev. Dyn. 1992, 194, 209–221. [Google Scholar] [CrossRef]
- Kummer, T.T.; Misgeld, T.; Lichtman, J.W.; Sanes, J.R. Nerve-independent formation of a topologically complex postsynaptic apparatus. J. Cell Biol. 2004, 164, 1077–1087. [Google Scholar] [CrossRef] [Green Version]
- Benavente, F.; Pinto, C.; Parada, M.; Henriquez, J.P.; Osses, N. Bone morphogenetic protein 2 inhibits neurite outgrowth of motor neuron-like NSC-34 cells and up-regulates its type II receptor. J. Neurochem. 2012, 122, 594–604. [Google Scholar] [CrossRef]
- Pinto, C.; Cardenas, P.; Osses, N.; Henriquez, J.P. Characterization of Wnt/beta-catenin and BMP/Smad signaling pathways in an in vitro model of amyotrophic lateral sclerosis. Front. Cell. Neurosci. 2013, 7, 239. [Google Scholar] [CrossRef] [Green Version]
- Aviles, E.C.; Pinto, C.; Hanna, P.; Ojeda, J.; Perez, V.; De Ferrari, G.V.; Zamorano, P.; Albistur, M.; Sandoval, D.; Henriquez, J.P. Frizzled-9 impairs acetylcholine receptor clustering in skeletal muscle cells. Front. Cell. Neurosci. 2014, 8, 110. [Google Scholar] [PubMed] [Green Version]
- Winning, R.S.; Krull, C.E. Knockdown of ephrin-A5 expression by 40% does not affect motor axon growth or migration into the chick hindlimb. Int. J. Mol. Sci. 2011, 12, 8362–8371. [Google Scholar] [CrossRef] [PubMed]
- Dieni, S.; Matsumoto, T.; Dekkers, M.; Rauskolb, S.; Ionescu, M.S.; Deogracias, R.; Gundelfinger, E.D.; Kojima, M.; Nestel, S.; Frotscher, M.; et al. BDNF and its pro-peptide are stored in presynaptic dense core vesicles in brain neurons. J. Cell Biol. 2012, 196, 775–788. [Google Scholar] [CrossRef] [Green Version]
- Shimojo, M.; Courchet, J.; Pieraut, S.; Torabi-Rander, N.; Sando, R., 3rd; Polleux, F.; Maximov, A. SNAREs Controlling Vesicular Release of BDNF and Development of Callosal Axons. Cell Rep. 2015, 11, 1054–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ascano, M.; Richmond, A.; Borden, P.; Kuruvilla, R. Axonal targeting of Trk receptors via transcytosis regulates sensitivity to neurotrophin responses. J. Neurosci. 2009, 29, 11674–11685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koseki, H.; Donega, M.; Lam, B.Y.; Petrova, V.; van Erp, S.; Yeo, G.S.; Kwok, J.C.; Ffrench-Constant, C.; Eva, R.; Fawcett, J.W. Selective rab11 transport and the intrinsic regenerative ability of CNS axons. Elife 2017, 6, e26956. [Google Scholar] [CrossRef] [PubMed]
- Koles, K.; Budnik, V. Exosomes go with the Wnt. Cell. Logist. 2012, 2, 169–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korkut, C.; Ataman, B.; Ramachandran, P.; Ashley, J.; Barria, R.; Gherbesi, N.; Budnik, V. Trans-synaptic transmission of vesicular Wnt signals through Evi/Wntless. Cell 2009, 139, 393–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salinas, P.C. Wnt signaling in the vertebrate central nervous system: From axon guidance to synaptic function. Cold Spring Harb. Perspect. Biol. 2012, 4, a008003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inestrosa, N.C.; Varela-Nallar, L. Wnt signalling in neuronal differentiation and development. Cell Tissue Res. 2015, 359, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Castro-Piedras, I.; Simmons, G.E., Jr.; Pruitt, K. Dishevelled: A masterful conductor of complex Wnt signals. Cell. Signal. 2018, 47, 52–64. [Google Scholar] [CrossRef]
- Mella, J.; Perez, V.; Zelada, D.; Moreno, N.; Ionescu, A.; Perlson, E.; Henriquez, J.P. Efficient gene transfer into primary muscle cells to analyze nerve-independent postsynaptic organization in vitro. Neuromuscul. Disord. 2019, 29, 533–542. [Google Scholar] [CrossRef]
- Neuhuber, B.; Daniels, M.P. Targeting of recombinant agrin to axonal growth cones. Mol. Cell. Neurosci. 2003, 24, 1180–1196. [Google Scholar] [CrossRef] [PubMed]
- Varela-Nallar, L.; Alfaro, I.E.; Serrano, F.G.; Parodi, J.; Inestrosa, N.C. Wingless-type family member 5A (Wnt-5a) stimulates synaptic differentiation and function of glutamatergic synapses. Proc. Natl. Acad. Sci. USA 2010, 107, 21164–21169. [Google Scholar] [CrossRef] [Green Version]
- Buckley, K.; Kelly, R.B. Identification of a transmembrane glycoprotein specific for secretory vesicles of neural and endocrine cells. J. Cell Biol. 1985, 100, 1284–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arimura, N.; Kimura, T.; Nakamuta, S.; Taya, S.; Funahashi, Y.; Hattori, A.; Shimada, A.; Menager, C.; Kawabata, S.; Fujii, K.; et al. Anterograde transport of TrkB in axons is mediated by direct interaction with Slp1 and Rab27. Dev. Cell 2009, 16, 675–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niwa, S.; Tanaka, Y.; Hirokawa, N. KIF1Bbeta- and KIF1A-mediated axonal transport of presynaptic regulator Rab3 occurs in a GTP-dependent manner through DENN/MADD. Nat. Cell Biol. 2008, 10, 1269–1279. [Google Scholar] [CrossRef]
- Lund, V.K.; Lycas, M.D.; Schack, A.; Andersen, R.C.; Gether, U.; Kjaerulff, O. Rab2 drives axonal transport of dense core vesicles and lysosomal organelles. Cell Rep. 2021, 35, 108973. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, J.B.; Nagy, G.; Varoqueaux, F.; Nehring, R.B.; Brose, N.; Wilson, M.C.; Neher, E. Differential control of the releasable vesicle pools by SNAP-25 splice variants and SNAP-23. Cell 2003, 114, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Sudhof, T.C.; Rothman, J.E. Membrane fusion: Grappling with SNARE and SM proteins. Science 2009, 323, 474–477. [Google Scholar] [CrossRef] [Green Version]
- Torres, V.I.; Barrera, D.P.; Varas-Godoy, M.; Arancibia, D.; Inestrosa, N.C. Selective Surface and Intraluminal Localization of Wnt Ligands on Small Extracellular Vesicles Released by HT-22 Hippocampal Neurons. Front. Cell Dev. Biol. 2021, 9, 735888. [Google Scholar] [CrossRef]
- David, M.D.; Canti, C.; Herreros, J. Wnt-3a and Wnt-3 differently stimulate proliferation and neurogenesis of spinal neural precursors and promote neurite outgrowth by canonical signaling. J. Neurosci. Res. 2010, 88, 3011–3023. [Google Scholar] [CrossRef] [Green Version]
- Anderson, K.N.; Potter, A.C.; Piccenna, L.G.; Quah, A.K.; Davies, K.E.; Cheema, S.S. Isolation and culture of motor neurons from the newborn mouse spinal cord. Brain Res. Protoc. 2004, 12, 132–136. [Google Scholar] [CrossRef]
- Pinto, C.; Medinas, D.B.; Fuentes-Villalobos, F.; Maripillan, J.; Castro, A.F.; Martinez, A.D.; Osses, N.; Hetz, C.; Henriquez, J.P. beta-catenin aggregation in models of ALS motor neurons: GSK3beta inhibition effect and neuronal differentiation. Neurobiol. Dis. 2019, 130, 104497. [Google Scholar] [CrossRef]
- Ciani, L.; Salinas, P.C. c-Jun N-terminal kinase (JNK) cooperates with Gsk3beta to regulate Dishevelled-mediated microtubule stability. BMC Cell Biol. 2007, 8, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosso, S.B.; Sussman, D.; Wynshaw-Boris, A.; Salinas, P.C. Wnt signaling through Dishevelled, Rac and JNK regulates dendritic development. Nat. Neurosci. 2005, 8, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Stamatakou, E.; Hoyos-Flight, M.; Salinas, P.C. Wnt Signalling Promotes Actin Dynamics during Axon Remodelling through the Actin-Binding Protein Eps8. PLoS ONE 2015, 10, e0134976. [Google Scholar]
- Gonzalez-Fernandez, C.; Gonzalez, P.; Rodriguez, F.J. New insights into Wnt signaling alterations in amyotrophic lateral sclerosis: A potential therapeutic target? Neural Regen. Res. 2020, 15, 1580–1589. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Guan, Y.; Zhao, Z.; Meng, F.; Wang, X.; Gao, X.; Liu, J.; Chen, Y.; Zhou, F.; Zhou, S.; et al. Potential Roles of the WNT Signaling Pathway in Amyotrophic Lateral Sclerosis. Cells 2021, 10, 839. [Google Scholar] [CrossRef]
- Jing, H.; Chen, P.; Hui, T.; Yu, Z.; Zhou, J.; Fei, E.; Wang, S.; Ren, D.; Lai, X.; Li, B. Synapse-specific Lrp4 mRNA enrichment requires Lrp4/MuSK signaling, muscle activity and Wnt non-canonical pathway. Cell Biosci. 2021, 11, 105. [Google Scholar] [CrossRef] [PubMed]
- Barik, A.; Lu, Y.; Sathyamurthy, A.; Bowman, A.; Shen, C.; Li, L.; Xiong, W.C.; Mei, L. LRP4 is critical for neuromuscular junction maintenance. J. Neurosci. 2014, 34, 13892–13905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohkawara, B.; Cabrera-Serrano, M.; Nakata, T.; Milone, M.; Asai, N.; Ito, K.; Ito, M.; Masuda, A.; Ito, Y.; Engel, A.G.; et al. LRP4 third beta-propeller domain mutations cause novel congenital myasthenia by compromising agrin-mediated MuSK signaling in a position-specific manner. Hum. Mol. Genet. 2014, 23, 1856–1868. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Lu, Y.; Zhang, B.; Figueiredo, D.; Bean, J.; Jung, J.; Wu, H.; Barik, A.; Yin, D.M.; Xiong, W.C.; et al. Antibodies against low-density lipoprotein receptor-related protein 4 induce myasthenia gravis. J. Clin. Investig. 2013, 123, 5190–5202. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinto, C.; Pérez, V.; Mella, J.; Albistur, M.; Caprile, T.; Bronfman, F.C.; Henríquez, J.P. Transport and Secretion of the Wnt3 Ligand by Motor Neuron-like Cells and Developing Motor Neurons. Biomolecules 2021, 11, 1898. https://doi.org/10.3390/biom11121898
Pinto C, Pérez V, Mella J, Albistur M, Caprile T, Bronfman FC, Henríquez JP. Transport and Secretion of the Wnt3 Ligand by Motor Neuron-like Cells and Developing Motor Neurons. Biomolecules. 2021; 11(12):1898. https://doi.org/10.3390/biom11121898
Chicago/Turabian StylePinto, Cristina, Viviana Pérez, Jessica Mella, Miguel Albistur, Teresa Caprile, Francisca C. Bronfman, and Juan Pablo Henríquez. 2021. "Transport and Secretion of the Wnt3 Ligand by Motor Neuron-like Cells and Developing Motor Neurons" Biomolecules 11, no. 12: 1898. https://doi.org/10.3390/biom11121898