
Allelic Variation Analysis at the Vernalization Response and Photoperiod Genes in Russian Wheat Varieties Identified Two Novel Alleles of Vrn-B3


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. PCR Amplification
- FTpr-F//R: 94 °C for 4 min to denature the DNA, 35 cycles at 94 °C for 10–40 s, 61 °C for 30 s, and 68 °C for 2.5 min, followed by a final extension at 72 °C for 10 min. The reaction was performed with 2X LR HS-PCR-Color (Biolabmix, Novosibirsk, Russia);
- VRN-B3d-F//R: 95 °C for 5 min; 35 cycles of 95 °C for 45 s, 62 °C for 1 min, 72 °C for 1 min, and a final extension at 72 °C for 10 min;
- VRN-B3e-F//R: 95 °C for 5 min, 35 cycles of 95 °C for 15 s, 59 °C for 30 s, 72 °C for 30 s, and a final extension at 72 °C for 5 min.
2.3. DNA Sequencing and Analysis
2.4. Gene Expression Analysis
3. Results
3.1. Allelic Composition of Vrn-1, Vrn-3, and Ppd-D1 Alleles in Common Russian Wheat Varieties
3.2. Sequence Analysis of the Two Novel Vrn-B3 Alleles
3.3. Development of Specific Markers for Detecting Novel Vrn-B3 Alleles
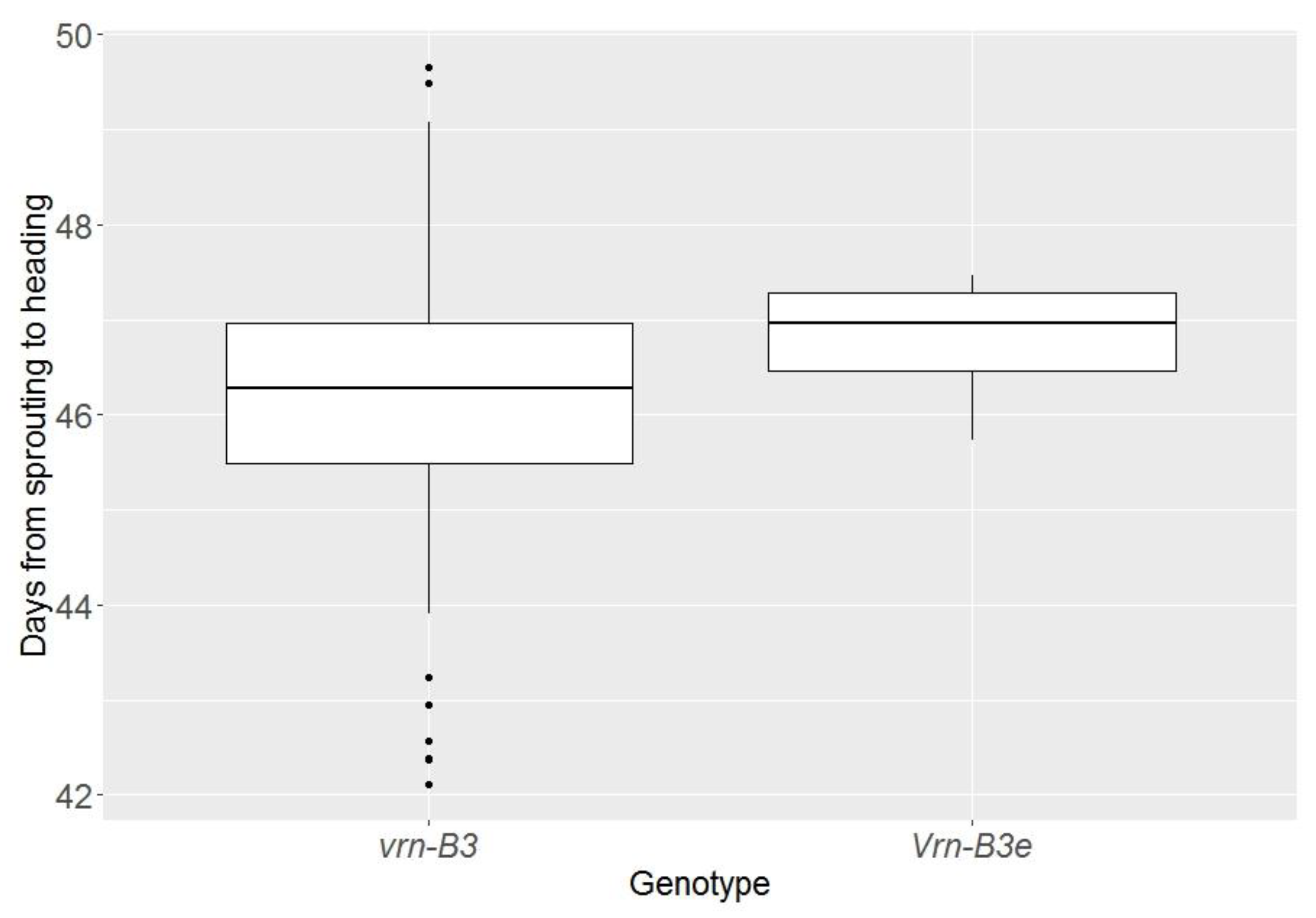
3.4. The Effects of the Vrn-B3d and Vrn-B3e Alleles on Wheat Heading Time
3.5. The Vrn-B3d Allele: Expression-Level Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Snape, J.W.; Butterworth, K.; Whitechurch, E.; Worland, A.J. Waiting for fine times: Genetics of flowering time in wheat. Euphytica 2001, 119, 185–190. [Google Scholar] [CrossRef]
- Yan, L.; Fu, D.; Li, C.; Blechl, A.; Tranquilli, G.; Bonafede, M.; Sanchez, A.; Valarik, M.; Yasuda, S.; Dubcovsky, J. The wheat and barley vernalization gene VRN3 is an orthologue of FT. Proc. Natl. Acad. Sci. USA 2006, 103, 19581–19586. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Dubcovsky, J. Wheat FT protein regulates VRN1 transcription through interactions with FDL2. Plant J. 2008, 55, 543–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, A.; Dubcovsky, J. Wheat TILLING mutants show that the vernalization gene VRN1 down-regulates the flowering repressor VRN2 in leaves but is not essential for flowering. PLoS Genet. 2012, 8, e1003134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.; Loukoianov, A.; Tranquilli, G.; Helguera, M.; Fahima, T.; Dubcovsky, J. Positional cloning of the wheat vernalization gene VRN1. Proc. Natl. Acad. Sci. USA 2003, 100, 6263–6268. [Google Scholar] [CrossRef] [Green Version]
- Pugsley, A.T. A genetic analysis of the spring-winter habit of growth in wheat. Aust. J. Agric. Res. 1971, 22, 21–31. [Google Scholar] [CrossRef]
- Goncharov, N.P. Response to vernalization in wheat: Its quantitative or qualitative nature. Cereal Res. Commun. 2004, 32, 323–330. [Google Scholar] [CrossRef]
- Yan, L.; Helguera, M.; Kato, K.; Fukuyama, S.; Sherman, J.; Dubcovsky, J. Allelic variation at the VRN-1 promoter region in polyploid wheat. Theor. Appl. Genet. 2004, 109, 1677–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, D.; Szűcs, P.; Yan, L.; Helguera, M.; Skinner, J.S.; Von Zitzewitz, J.; Hayes, P.M.; Dubcovsky, J. Large deletions within the first intron in VRN-1 are associated with spring growth habit in barley and wheat. Mol. Genet. Genom. 2005, 273, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Shcherban, A.; Emtseva, M.; Efremova, T. Molecular genetical characterization of vernalization genes Vrn-A1, Vrn-B1 and Vrn-D1 in spring wheat germplasm from Russia and adjacent regions. Cereal Res. Commun. 2012, 40, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Scarth, R.; Law, C.N. The control of the day-length response in wheat by the group 2 chromosomes. Z. Pflanzenzücht. 1984, 92, 140–150. [Google Scholar]
- Worland, A.J.; Börner, A.; Korzun, V.; Li, W.M.; Petrovíc, S.; Sayers, E.J. The influence of photoperiod genes on the adaptability of European winter wheats. Euphytica 1998, 100, 385–394. [Google Scholar] [CrossRef]
- Hunt, L.A. Photoperiodic responses of winter wheats from different climatic regions. J. Plant Breed 1979, 82, 70–80. [Google Scholar]
- Kumar, S.; Sharma, V.; Chaudhary, S.; Tyagi, A.; Mishra, P.; Priyadarshini, A.; Singh, A. Genetics of flowering time in bread wheat Triticum aestivum: Complementary interaction between vernalization-insensitive and photoperiod-insensitive mutations imparts very early flowering habit to spring wheat. J. Genet. 2012, 91, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Turck, F.; Fornara, F.; Coupland, G. Regulation and Identity of Florigen: FLOWERING LOCUS T Moves Center Stage. Annu. Rev. Plant Biol. 2008, 59, 573–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Gao, M.; Zhang, J.; Zuo, A.; Shang, X.; Cui, D. Molecular characterization of vernalization and response genes in bread wheat from the Yellow and Huai Valley of China. BMC Plant Biol. 2013, 13, 199. [Google Scholar] [CrossRef] [Green Version]
- Lapochkina, I.F.; Volkova, G.A. Development of collection of spring common wheat lines substituted and supplemented by chromosomes Aegilops speltoides Tausch. Genetika 1994, 30, 86–87. [Google Scholar]
- Adonina, I.G.; Petrash, N.V.; Timonova, E.M.; Khristov, Y.A.; Salina, E.A. Construction and study of leaf rust-resistant common wheat lines with translocations of Aegilops speltoides Tausch. genetic material. Russ. J. Genet. 2012, 48, 404–409. [Google Scholar] [CrossRef]
- Salina, E.A.; Leonova, I.N.; Shcherban, A.B. Method of Creation of Lines of Spring Soft Wheat with Elongated Period of Earing and Complex Resistance to Fungal Diseases. Patent RU 2535985, 10 October 2014. [Google Scholar]
- Petrash, N.V.; Leonova, I.N.; Adonina, I.G.; Salina, E.A. Effect of translocations from Aegilops speltoides Tausch on resistance to fungal diseases and productivity in common wheat. Russ. J. Genet. 2016, 52, 1253–1262. [Google Scholar] [CrossRef]
- Plaschke, J.; Ganal, M.W.; Roder, M.S. Detection of genetic diversity in closely related bread wheat. Theor. Appl. Genet. 1995, 91, 1001–1007. [Google Scholar] [CrossRef]
- Beales, J.; Turner, A.; Griffiths, S.; Snape, J.W.; Laurie, D.A. A Pseudo-Response Regulator is misexpressed in the photoperiod insensitive Ppd-D1a mutant of wheat (Triticum aestivum L.). Theor. Appl. Genet. 2007, 115, 721–733. [Google Scholar] [CrossRef]
- Shaw, L.M.; Turner, A.S.; Laurie, D.A. The impact of photoperiod insensitive Ppd-1a mutations on the photoperiod pathway across the three genomes of hexaploid wheat (Triticum aestivum). Plant J. 2012, 71, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Wicker, T.; Matthews, D.E.; Keller, B. TREP: A database for Triticeae repetitive elements. Trends Plant Sci. 2002, 7, 561–562. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Crismani, W.; Baumann, U.; Sutton, T.; Shirley, N.; Webster, T.; Spangenberg, G.; Langridge, P.; Able, J.A. Microarray expression analysis of meiosis and microsporogenesis in hexaploid bread wheat. BMC Genom. 2006, 7, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paolacci, A.R.; Tanzarella, O.A.; Porceddu, E.; Ciaffi, M. Identification and validation of reference genes for quantitative RT-PCR normalization in wheat. BMC Mol. Biol. 2009, 10, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Likhenko, I.E.; Stasyuk, A.I.; Shcherban’, A.B.; Zyryanova, A.F.; Likhenko, N.I.; Salina, E.A. Study of allelic composition of Vrn-1 and Ppd-1 genes in early–ripening and middle–early varieties of spring soft wheat in siberia. Russ. J. Genet. Appl. Res. 2015, 5, 198–207. [Google Scholar] [CrossRef]
- Shcherban, A.B.; Börner, A.; Salina, E.A. Effect of VRN-1 and PPD-D1 genes on heading time in European bread wheat cultivars. Plant Breed. 2015, 134, 49–55. [Google Scholar] [CrossRef]
- Iqbal, M.; Navabi, A.; Yang, R.C.; Salmon, D.F.; Spaner, D. Molecular characterization of vernalization response genes in Canadian spring wheat. Genome 2007, 50, 511–516. [Google Scholar] [CrossRef]
- Santra, D.K.; Santra, M.; Allan, R.E.; Campbell, K.G.; Kidwell, K.K. Genetic and molecular characterization of vernalization genes Vrn-A1, Vrn-B1, and Vrn-D1 in spring wheat germplasm from the pacific northwest region of the U.S.A. Plant Breed. 2009, 128, 576–584. [Google Scholar] [CrossRef]
- Muterko, A.; Kalendar, R.; Salina, E. Allelic variation at the VERNALIZATION-A1, VRN-B1, VRN-B3, and PHOTOPERIOD-A1 genes in cultivars of Triticum durum Desf. Planta 2016, 244, 1253–1263. [Google Scholar] [CrossRef]
- McFadden, E.S. A successful transfer of emmer characters to vulgare wheat. J. Am. Soc. Agron. 1930, 22, 1020–1034. [Google Scholar] [CrossRef] [Green Version]
- Muterko, A.; Salina, E. Divergence of VRN-B3 alleles during the evolution of domesticated wheat. Mol. Genet. Genom. 2019, 294, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.K.; Xiao, Y.G.; Zhang, Y.; Xia, X.C.; Dubcovsky, J.; He, Z.H. Allelic variation at the vernalization genes Vrn-A1, Vrn-B1, Vrn-D1, and Vrn-B3 in Chinese wheat cultivars and their association with growth habit. Crop Sci. 2008, 48, 458–470. [Google Scholar] [CrossRef] [Green Version]
- Derakhshan, B.; Mohammadi, S.A.; Moghaddam, M.; Kamali, M.R.J. Molecular characterization of vernalization genes in Iranian wheat landraces. Crop Breed. J. 2013, 3, 1–11. [Google Scholar]
- Sanmiguel, P.; Gaut, B.S.; Tikhonov, A.; Nakajima, Y.; Bennetzen, J.L. The paleontology of intergene retrotransposons of maize. Nat. Genet. 1998, 20, 43–45. [Google Scholar] [CrossRef]
- Charles, M.; Belcram, H.; Carter, M.; Huteau, V. Dynamics and Differential Proliferation of Transposable Elements During the Evolution of the B and A Genomes of Wheat. Genetics 2008, 180, 1071–1086. [Google Scholar] [CrossRef] [Green Version]
- Salina, E.A.; Sergeeva, E.M.; Adonina, I.G.; Shcherban, A.B.; Belcram, H.; Huneau, C.; Chalhoub, B. The impact of Ty3-gypsy group LTR retrotransposons Fatima on B-genome specificity of polyploid wheats. BMC Plant Biol. 2011, 11, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, M.; Torkamaneh, D.; Mehrazar, E. Molecular examination and genotype diversity of vernalization sensitivity and photoperiod response in old and modern bread wheat cultivars grown in Iran. J. Crop Sci. Biotechnol. 2012, 15, 259–265. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence | Gene Region | Annealing Temp. °C | Allelic Variant | Product Size (bp) | Source |
|---|---|---|---|---|---|---|
| Vrn1AF Vrn1-Int1R | gaaaggaaaaattctgctcg gcaggaaatcgaaatcgaag | VRN-A1 promoter | 60 | vrn-A1 Vrn-A1a Vrn-A1b | 734 876 and 965 714 | [8] |
| Ex1/C/F Intr1/B/R3 | gttctccaccgagtcatggt ctcatgccaaaaattgaagatga | VRN-B1 intron-1 | 58 | Vrn-B1a Vrn-B1b Vrn-B1c | 1091 1055 705 | [9] |
| Ex1/C/F Intr1/B/R3 | gttctccaccgagtcatggt caaatgaaaaggaatgagagca | VRN-B1 intron-1 | 60 | vrn-B1 | 1531 | [9] |
| Intr1/D/F Intr1/D/R3 Intr1/D/R4 | gttgtctgcctcatcaaatcc ggtcactggtggtctgtgc aaatgaaaaggaacgagagcg | VRN-D1 intron-1 | 61 | vrn-D1 Vrn-D1a | 997 1671 | [9] |
| Ppd-D1_F Ppd-D1_R1 Ppd-D1_R2 | acgcctcccactacactg gttggttcaaacagagagc cactggtggtagctgagatt | PPD-D1 promoter | 54 | Ppd-D1a Ppd-D1b | 288 414 | [22] |
| FT-B-INS-F FT-B-INS-R | cataatgccaagccggtgagtac atgtctgccaattagctagc | VRN-B3 promoter | 57 | Vrn-B3a | 1200 | [2] |
| FT-B-NOINS-F FT-B-NOINS-R | atgctttcgcttgccatcc ctatccctaccggccattag | VRN-B3 promoter | 57 | vrn-B3 | 1140 | [2] |
| TaFT-F3 TaFT-R3 | cagcagcccagggttgag atctgggtctaccatcacgagtg | VRN-B3 exons 1 and 2 | 60 | - | 72 | [2] |
| TaFT-B1-F TaFT-B1-R | gtcgttcgggcaggag tggaagagtacgagcacga | VRN-B3 exon 3 | 60 | - | 81 | [23] |
| FTpr-F FTpr-R | cgaaagcggagggtatattaaa cccgaacatagaagaagcatag | VRN-B3 promoter | 60 | vrn-B3 Vrn-B3e Vrn-B3d | 1384 1544 3001 | This study |
| Vrn-B3e-F Vrn-B3e-R | cgccagacagcattcctatta tactggtgtctcttgccattc | VRN-B3 promoter | 59 | Vrn-B3e | 302 | This study |
| Vrn-B3d-F Vrn-B3d-R | gttacaccacatccacagaac ccaatactaccggccactaa | VRN-B3 promoter | 62 | Vrn-B3d | 243 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berezhnaya, A.; Kiseleva, A.; Leonova, I.; Salina, E. Allelic Variation Analysis at the Vernalization Response and Photoperiod Genes in Russian Wheat Varieties Identified Two Novel Alleles of Vrn-B3. Biomolecules 2021, 11, 1897. https://doi.org/10.3390/biom11121897
Berezhnaya A, Kiseleva A, Leonova I, Salina E. Allelic Variation Analysis at the Vernalization Response and Photoperiod Genes in Russian Wheat Varieties Identified Two Novel Alleles of Vrn-B3. Biomolecules. 2021; 11(12):1897. https://doi.org/10.3390/biom11121897
Chicago/Turabian StyleBerezhnaya, Alina, Antonina Kiseleva, Irina Leonova, and Elena Salina. 2021. "Allelic Variation Analysis at the Vernalization Response and Photoperiod Genes in Russian Wheat Varieties Identified Two Novel Alleles of Vrn-B3" Biomolecules 11, no. 12: 1897. https://doi.org/10.3390/biom11121897