1. Introduction
Gastric cancer is one of the most common causes of cancer death in the world, particularly in developing countries. In 2012, the global incidence and death due to gastric cancer were 950,000 and 723,000, respectively [
1]. Based on Lauren’s classification, gastric cancer can be histopathologically classified into two major subtypes, a diffuse type and an intestinal type [
2]. We had previously performed cDNA microarray analysis to identify genes differentially expressed between diffuse and intestinal types [
3]. Among 23,040 genes examined, we observed overexpression of IQ-domain GTPase activating protein 3 (
IQGAP3) in both types of gastric cancer.
IQGAP scaffold proteins are conserved during evolution and the gene family comprises three isoforms,
IQGAP1,
IQGAP2 and
IQGAP3. The three isoforms were found in most vertebrates including humans, and the homologous proteins were identified in many eukaryotic species [
4]. IQGAPs are found to be responsible for regulating diverse cellular processes including cytokinesis, cell migration, cell proliferation, intracellular signaling, vesicle trafficking, and cytoskeletal dynamics [
5,
6,
7,
8,
9,
10,
11,
12]. All three genes were suggested to be associated with different types of cancers [
4]. However, in contrast to
IQGAP1 and
IQGAP3, which were suggested to be oncogenes,
IQGAP2 was implicated to be a putative tumor-suppressor gene [
13,
14].
IQGAP1, the most well-investigated member of mammalian
IQGAP genes, is ubiquitously expressed. It is up-regulated in various types of cancer and likely involved in metastasis and neoplastic transformation [
11,
14,
15]. Previous studies suggested a direct interaction between IQGAP1 and Cdc42 and Rac1, which are members of the small Rho GTPase family [
4,
16]. The up-regulation of IQGAP1 promotes cell migration through inhibition of the intrinsic GTPase activities of Cdc42 and Rac1 [
8,
17]. Among small Rho GTPases, Cdc42 and Rac1 induce the formation of filopodia, lamellipodia and stress fibers [
18]. Activation of Rho GTPases induces remodeling of the cytoskeleton required for these morphological changes.
A previous study identified two non-synonymous somatic mutations at the Cdc42 and Rac1 activation binding sites of
IQGAP1 in diffuse-type gastric cancers [
19]. In addition, Wu et al. [
20] showed that IQGAP1 was highly expressed in gastric cancer tissues and cell lines, and stimulated cell migration by interacting with RhoC GTPase. The same group further showed that the IQGAP1-RhoC complex significantly stimulated the proliferation of gastric cancer cells [
21]. Interestingly, knockdown of IQGAP1 alone, but not RhoC, attenuated migration and proliferation of gastric cancer cells, indicating the crucial role of IQGAP1 in gastric cancer tumorigenesis. In line with other studies,
IQGAP2 was suggested to be a tumor-suppressor gene in gastric cancer cell lines [
22]. It was down-regulated in half of the gastric cancer cell lines, which was likely due to aberrant methylation in the promoter region.
Recently, multiple studies have identified strong associations between the expression of IQGAP3 and poor prognosis in various types of cancer. Two studies showed that IQGAP3 promoted the growth and metastasis of lung cancer cells by modulating EGFR-ERK signaling [
23,
24]. Particularly, a high expression level of
IQGAP3 was observed in metastatic samples of lung cancer, which was identified as a marker of poor prognosis. IQGAP3 protein levels were significantly elevated in the plasma of hepatocellular carcinoma (HCC) patients, so the authors suggested that it can be used as a potential biomarker for detecting HCC [
25]. Furthermore, overexpression of IQGAP3 was associated with tumorigenesis of skin and microsatellite-stable stage III colorectal adenocarcinoma carrying
p53 mutations [
26,
27]. Similar to IQGAP1, IQGAP3 was found to be an effector of Rac1 and Cdc42 in mammalian neural cells, and to interact with Ras in epithelial cells [
28,
29]. A study showed that cell apoptosis, metastasis and Cdc42 pathways were strongly associated with IQGAP3 expression in pancreatic cancer patients [
30]. In addition, increased IQGAP3 promotes cell proliferation and invasion in breast cancer [
31], and correlates with poor prognosis in various cancers based on a recent pan-cancer study [
32]. Taken together, the above evidence suggested the role of IQGAP3 in promoting invasion and migration of cancer cells.
In this study, we hypothesized that the up-regulation of IQGAP3 is associated with the invasion/migration of gastric cancer cells. We, therefore, conducted overexpression and knockdown of IQGAP3 in different cell lines, and examined interactions between IQGAP3 and small GTPases to characterize its functional role in regulating invasion and/or migration ability. The results of this study should provide a better understanding of the progression of gastric cancer, and thereby facilitate the development of novel strategies for diagnosis and/or treatment of human tumors involving invasion and metastasis.
2. Materials and Methods
2.1. Cell Lines
Human gastric cell lines, MKN1, mouse fibroblast cell line (NIH3T3), and transformed human embryonic kidney cell line (293T) were purchased from the American Type Culture Collection (ATCC, Rockville, MD, USA). Human diffuse-type gastric cancer cell lines, ST-4 and TMK-1, were kindly provided by Dr. Tsuruo (Cancer Institute, Tokyo, Japan) and Dr. Yasui (Hiroshima University School of Medicine, Japan), respectively. All cells were cultured as monolayers in appropriate media; RPMI1640 (Sigma-Aldrich, St. Louis, MO, USA) for MKN1; DMEM (Sigma-Aldrich, St. Louis, MO, USA) for TMK-1, 293T and NIH3T3; each was supplemented with 10% fetal bovine serum (Cansera International, Etobicoke, ON, Canada) and 1% antibiotic/antimycotic solution (Sigma-Aldrich, St. Louis, MO, USA). Cells were maintained at 37 °C in an atmosphere of humidified air with 5% CO2.
2.2. Quantitative RT-PCR
Total RNA was extracted from the cultured cells using TRIZOL reagent (Invitrogen, Waltham, MA, USA) according to the manufacturer’s protocol. Extracted RNA was treated with DNaseI (Roche Diagnostics, Mannheim, Germany) and reverse transcribed to single-stranded cDNAs using oligo(dT)12-18 primer with Superscript II reverse transcriptase (Invitrogen, Waltham, MA, USA). The expression of the glutaminyl-tRNA synthetase gene (QARS) served as quantitative controls. Real-time PCR reaction was performed by TaqMan assay system according to the manufacturer’s protocol (Applied Biosystems, Foster City, CA, USA). The sequences of primers and probe for IQGAP3 and QARS are as follows: Forward primer, 5′-GGTGTCTTGGTGGAAATTGAAGAT-3′ and reverse primer, 5′-CGGCGTGATGTCAAAGATGA-3′; probe, 5′-FAM-CGCCTCTCACTTCAGA-MGB-3′ for IQGAP3; forward primer, 5′-GGTGGATGCAGCATTAGTGGA-3′ and reverse primer, 5′-AAGACGCTCAAACTGGAACTTGTC-3′; probe, 5′-FAM-CTCTGTGGCCCTGGCAAAACCCTT-MGB-3′ for QARS.
2.3. Preparation of Polyclonal Antibody against IQGAP3 and Immunohistochemistry
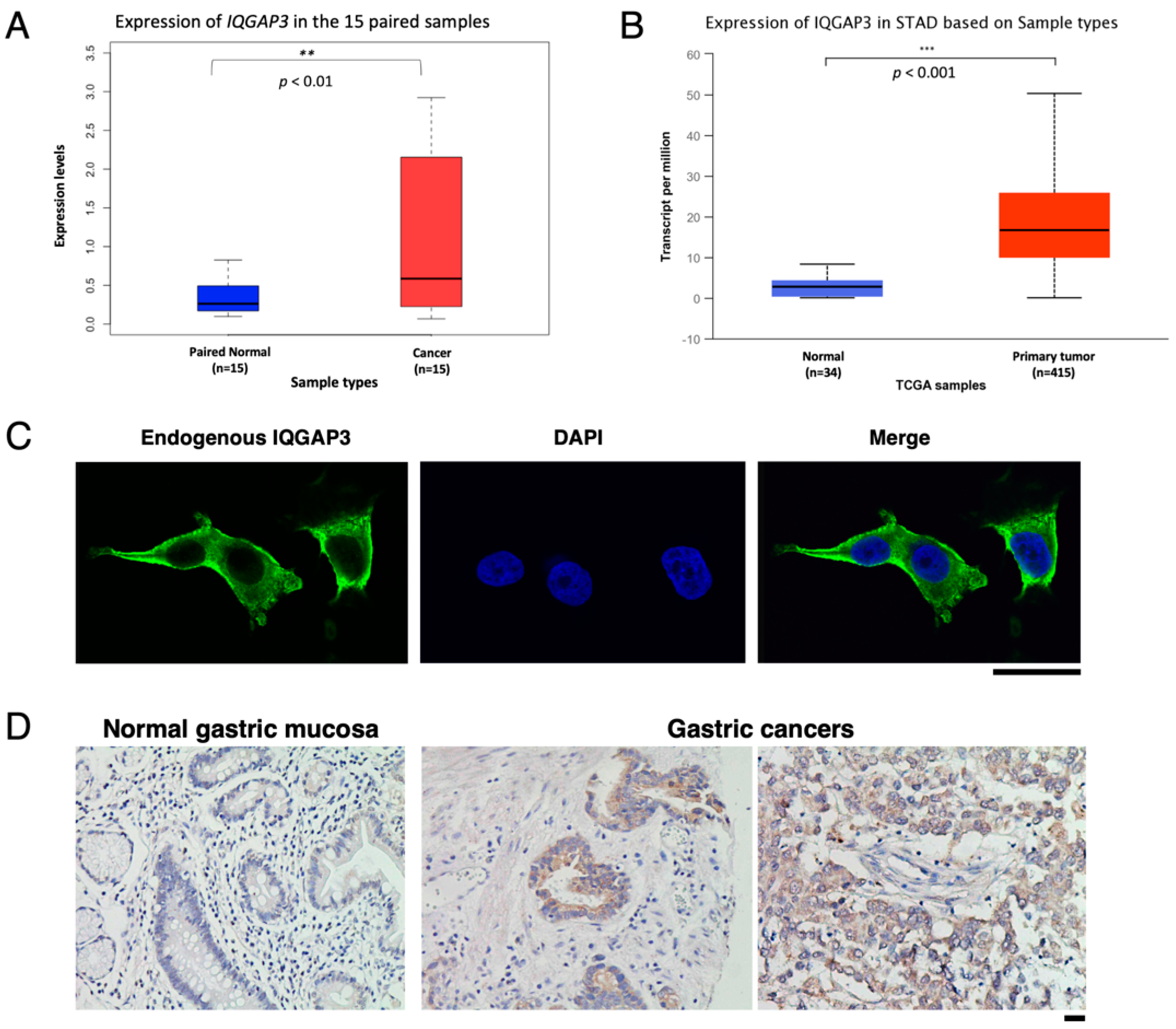
We selected two peptides from the hydrophobic regions of IQGAP3 sequence; HDDRESQDIYDHYLTQAC and CRRQYLRRLHYFQKNVNS for anti-IQGAP3. The high performance liquid chromatography (HPLC)-purified peptides were inoculated into rabbits 7 times, and the immune sera were purified using Affi-gel (Bio-Rad, Hercules, CA, USA) coupling with IQGAP3 peptides on affinity chromatography columns according to standard methodology. Tumor-tissue microarrays using formalin-fixed gastric cancers were obtained from AccuMax Array (ISU ABXIS, Seongnam-si, Korea), and subjected to the SAB-PO peroxidase immunostaining system (Nichirei, Tokyo, Japan) according to the manufacturer’s recommendations. Antigens were retrieved from deparaffinized and re-hydrated tissues by pre-treating the slides in citrate buffer (pH 6.0) for 10 min at 108 °C in an autoclave. Immunohistochemical staining was then performed using anti-IQGAP3 (1:500) antibody. IQGAP3 positivity was assessed semi-quantitatively as negative (no detectable staining or positive staining in <10% of tumor cells), and positive (positive staining in >10% of tumor cells), by three independent investigators with no prior knowledge of clinicopathological data. Cases were considered as positive if at least one of two tissue cores showed positive staining, and as negative if both tissue cores failed to show positive staining.
2.4. TCGA Database Analyses
The Stomach Adenocarcinoma dataset from TCGA (TCGA-STAD) was used to perform the following analyses. IQGAP3 expression between normal and cancer tissues were analyzed using UALCAN (
http://ualcan.path.uab.edu) [
33]. Different clinical characteristics were investigated including tumor grades, cancer stages, and nodal metastasis status, etc. Previously reported genetic alterations in
IQGAP3 were summarized using TCGA-STAD in cBioPortal (
http://cbioportal.org) [
34,
35]. The Kaplan–Meier Plotter database (
https://kmplot.com/analysis/) was used for survival analysis between patients with low and high expression of
IQGAP3 [
36].
IQGAP3 mRNA expression level in gastric cancer subtypes (diffuse and intestinal) were analyzed using Oncomine database (
www.oncomine.org). Asian-specific survival analysis was performed using the UCSC Xena database (
https://xena.ucsc.edu/). Student’s
t-test and log-rank test were used for box plots and survival analyses, respectively.
2.5. Construction of cDNA Expression Clone and Cell Transfection
The entire coding region of human IQGAP3 was amplified by RT and the PCR product was cloned into an appropriate enzyme site of expression vectors using primers 5′-TTCAAGCTTGGAAGAAGGAGGAACATGGAGAG-3′ (forward) and 5′-CTCGAATTCTTCCGCAAAAACTTCTTGTTGAGG-3′ (reverse) for pcDNA3.1-myc/His (Invitrogen, Waltham, MA, USA). Cells plated onto 10-cm dishes (1 × 106 cells/dish) were transfected using FuGENE6 reagent according to the manufacturer’s recommendations (Roche, Basel, Switzerland). Transfected cells were maintained for 7 days in cultured media supplemented with 0.5–0.7 μg/μL geneticin.
2.6. Construction of psiH1BX-IQGAP3
The plasmid expressing siRNA specific to
IQGAP3 was prepared by cloning the double-stranded oligonucleotides of the target transcript into psiH1BX3.0 vector that was constructed in our group as described previously [
37]: 5′-TCCCGCAAACCTGGATGCCATAATTCAAGAGATTATGGCATCCAGGTTTGC-3′ and 5′-AAAAGCAAACCTGGATGCCATAATCTCTTGAATTATGGCATCCAGGTTTGC-3′ for si-IQGAP3 (si7 in
Figure S1). Two additional siRNA expression plasmids were also constructed: A siRNA against
EGFP, and another siRNA against
IQGAP3 (si6 in
Figure S1), which was generated with the following primers: 5′-TCCCGCACTAAGACCACCTTCTATTCAAGAGATAGAAGGTGGTCTTAGTGC-3′ and 5′-AAAAGCACTAAGACCACCTTCTATCTCTTGAATAGAAGGTGGTCTTAGTGC-3′. The forward oligonucleotides and their complements were each phosphorylated by incubation with T4-polynucleotide kinase at 37 °C for 30 min, followed by boiling and slow cooling to anneal the two oligonucleotides. Each product was ligated into psiH1BX3.0 to create a siRNA expression vector. TMK-1 and MKN1 cells plated onto 10-cm dishes (1 × 10
6 cells/dish) were transfected with each siRNA plasmid using FuGENE6 reagent according to the manufacturer’s recommendations (Roche, Basel, Switzerland). Transfected cells were maintained for 7 days in cultured media supplemented with 0.5 μg/μL geneticin for TMK-1, and 0.7 μg/μL geneticin for MKN1. Total protein was extracted from the cells and the knockdown effect of each siRNA at the IQGAP3 protein level was examined by western blotting using an in-house antibody against IQGAP3. Since an
EGFP-siRNA and the other
IQGAP3-siRNA (si6) did not show any knockdown effect on IQGAP3, they were subsequently used as control siRNAs, namely si-control-1 and si-control-2, respectively (
Figure S1).
2.7. Matrigel Invasion Assay and Wound-Healing Migration Assay
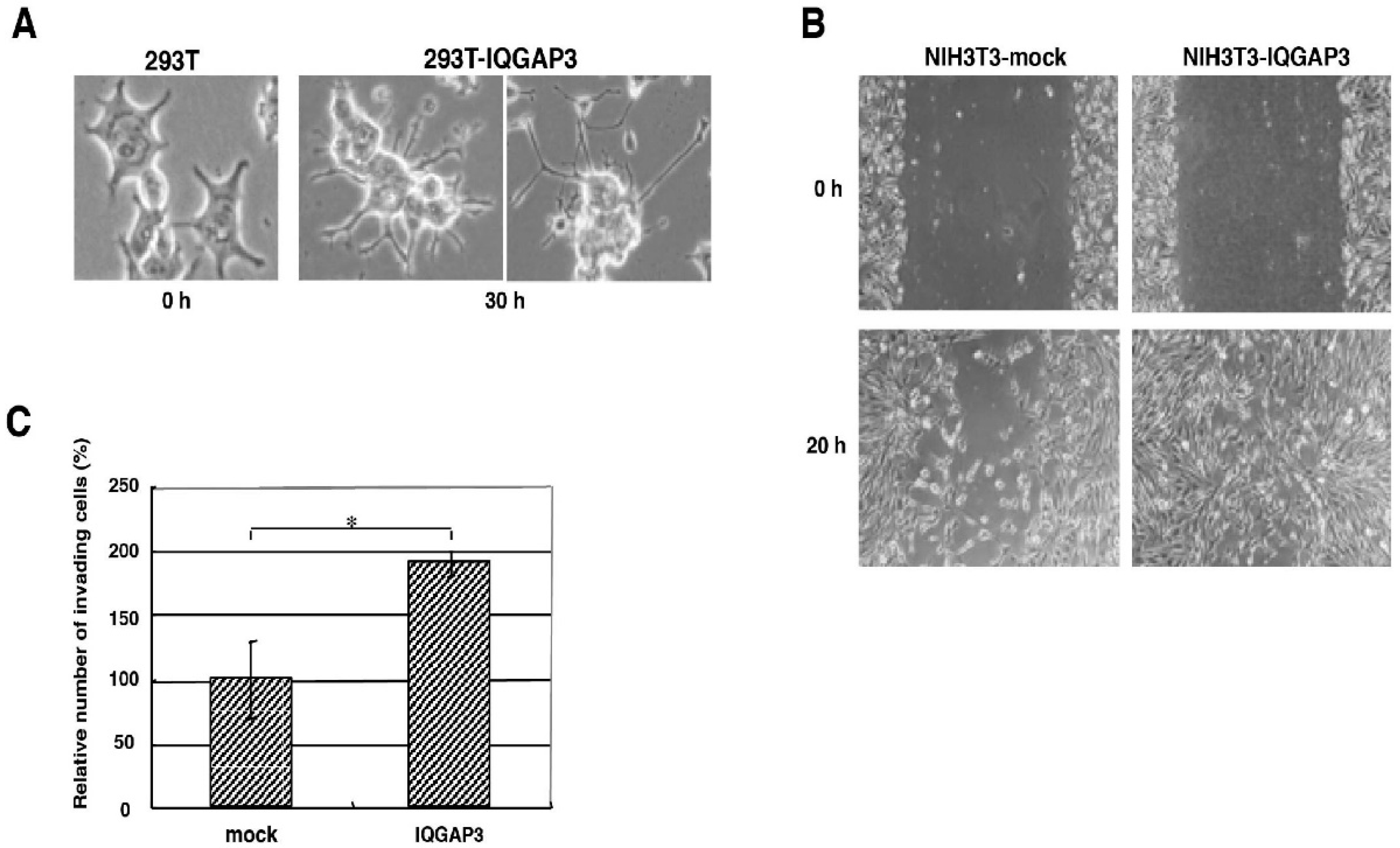
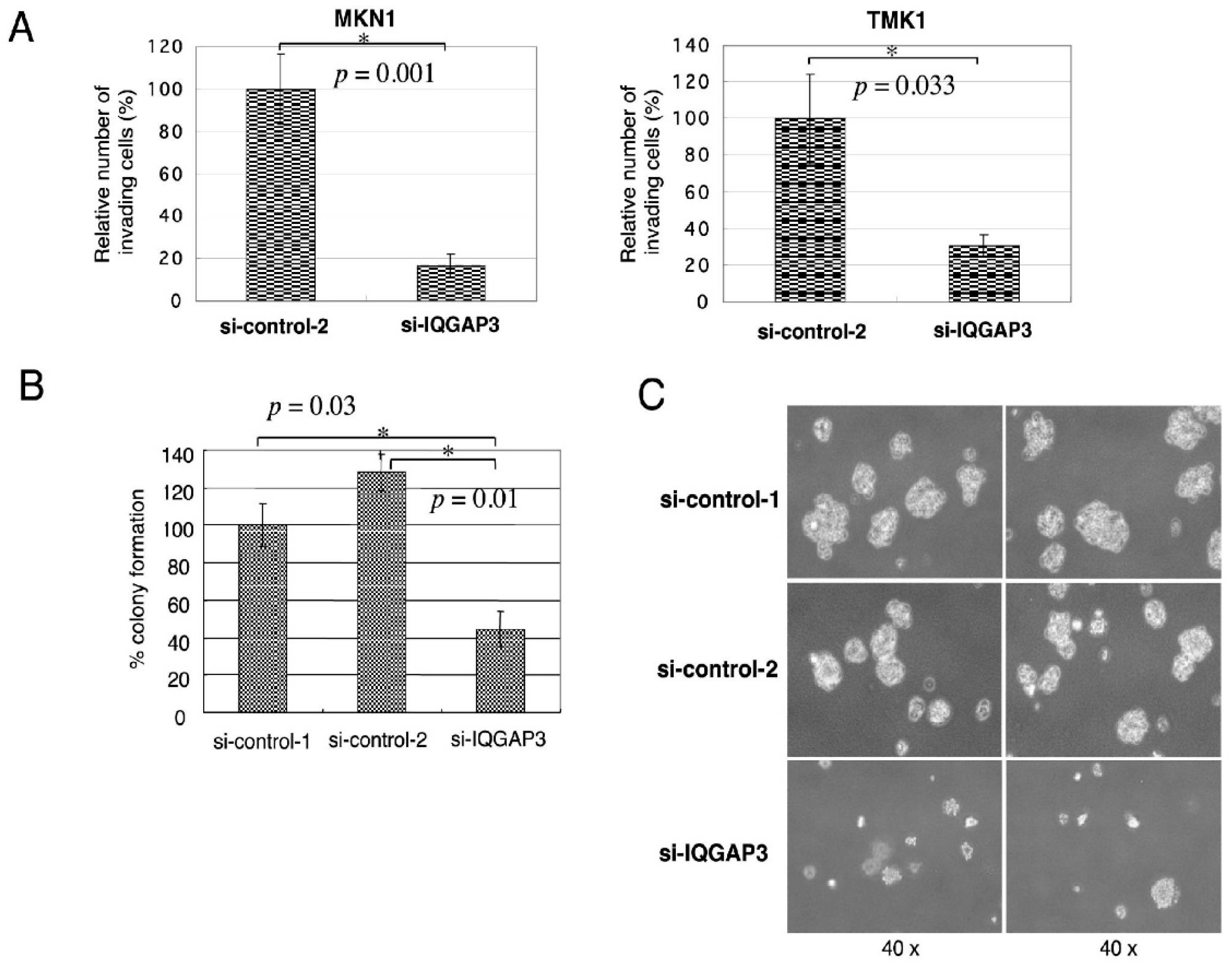
Cell invasion was assessed using BD BioCoatTM MatrigelTM Invasion Chambers with 8 µm inserts in 24-well plates (BD Biosciences, San Jose, CA, USA). After 36 h of incubation, the number of migrated NIH3T3 cells was estimated by counting three independent visual fields in a microscope with a 100× objective. Another Matrigel invasion assay was carried out using TMK-1 and MKN1 cells transfected with si-IQGAP3 or control siRNAs. All the invasion assays were replicated three times independently. For the wound-healing migration assay, cells were grown to confluency in BD BioCoatTM Fibronectin 2-well CultureSlides (BD Biosciences, San Jose, CA, USA) for 2 days, and scraped in the form of a cross made through the confluent monolayers with a 200 μL plastic pipette tip. Several wounded areas were marked for orientation and then photographed by phase-contrast microscopy. Student’s t-test (two-tailed) was used to study the differences between NIH3T3 cells transfected with pcDNA3.1-myc/His-IQGAP3 or mock vector, and si-IQGAP3 or control siRNAs, respectively.
2.8. Cell-Cell Adhesion Assay
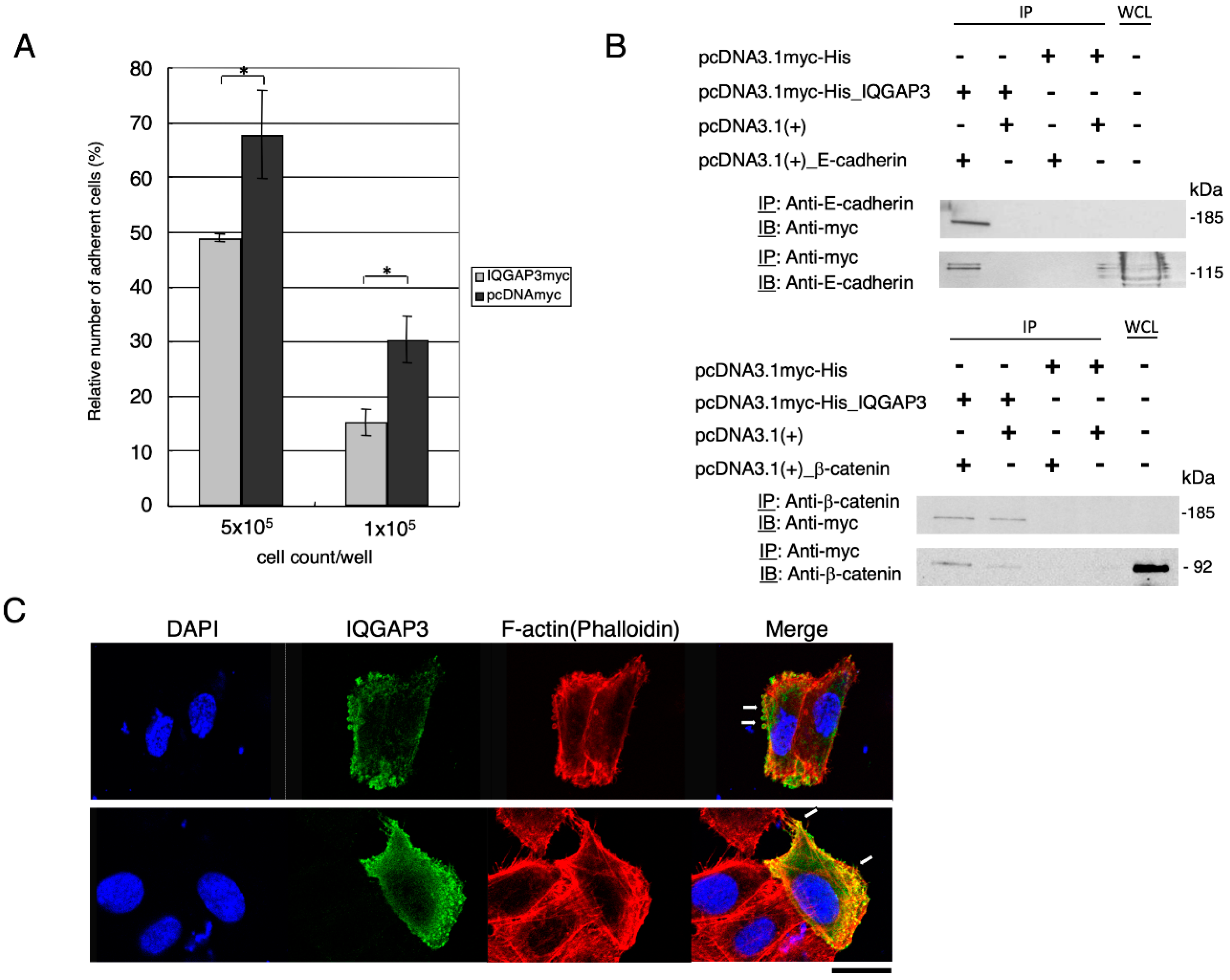
Unlabeled 293T cells were plated onto a 96-well Collagen type I-coated microplate (Iwaki, Shizuoka, Japan), and maintained until they became confluent monolayer. 293T cells expressing exogenous IQGAP3 or mock were then labeled with calcein-AM using Vybrant Cell Adhesion Assay Kit according to the supplier’s protocol (Molecular Probes, Eugene, OR, USA). The calcein-labeled 293T cells were seeded over the unlabeled cells and allowed to adhere to the underlying cells. After non-adherent calcein-labeled cells were removed by washing four-times with culture media, the fluorescent signal was detected by a fluorescence microplate reader (Fluoroskan Ascent FL, Thermo Fisher Scientific, Waltham, MA, USA) using a filter set of 494/517 nm.
2.9. Immunoprecipitation and Western Blotting
The cells were harvested at 48 h after transfection and lysed in CelLytic-M Mammalian Cell Lysis/Extraction reagent (Sigma-Aldrich, St. Louis, MO, USA) plus 1x Protease Inhibitor Cocktail EDTA (-) (Roche, Basel, Switzerland). Cell lysates were immunoprecipitated with anti-c-myc (9E10), anti-e-cadherin (HECD-1, Takara, Shiga, Japan), anti-β-catenin (14, BD Biosciences, San Jose, CA, USA), coupling with rec-Protein G (or A) Sepharose 4B (Zymed, San Francisco, CA, USA) overnight at 4 °C. After washing the beads five times with 1 × TBS buffer, 5 × Laemmli sample buffer was added and then boiled for 5 min to separate the precipitated protein complex. SDS-PAGE was subsequently performed and immunoblotting was carried out with the first antibodies, and HRP conjugated sheep anti-mouse IgG or donkey anti-rabbit IgG (Amersham Pharmacia, Little Chalfont, UK) served as the secondary antibody for ECL detection system (Amersham Pharmacia, Little Chalfont, UK).
2.10. Small GTPase Activation Assay
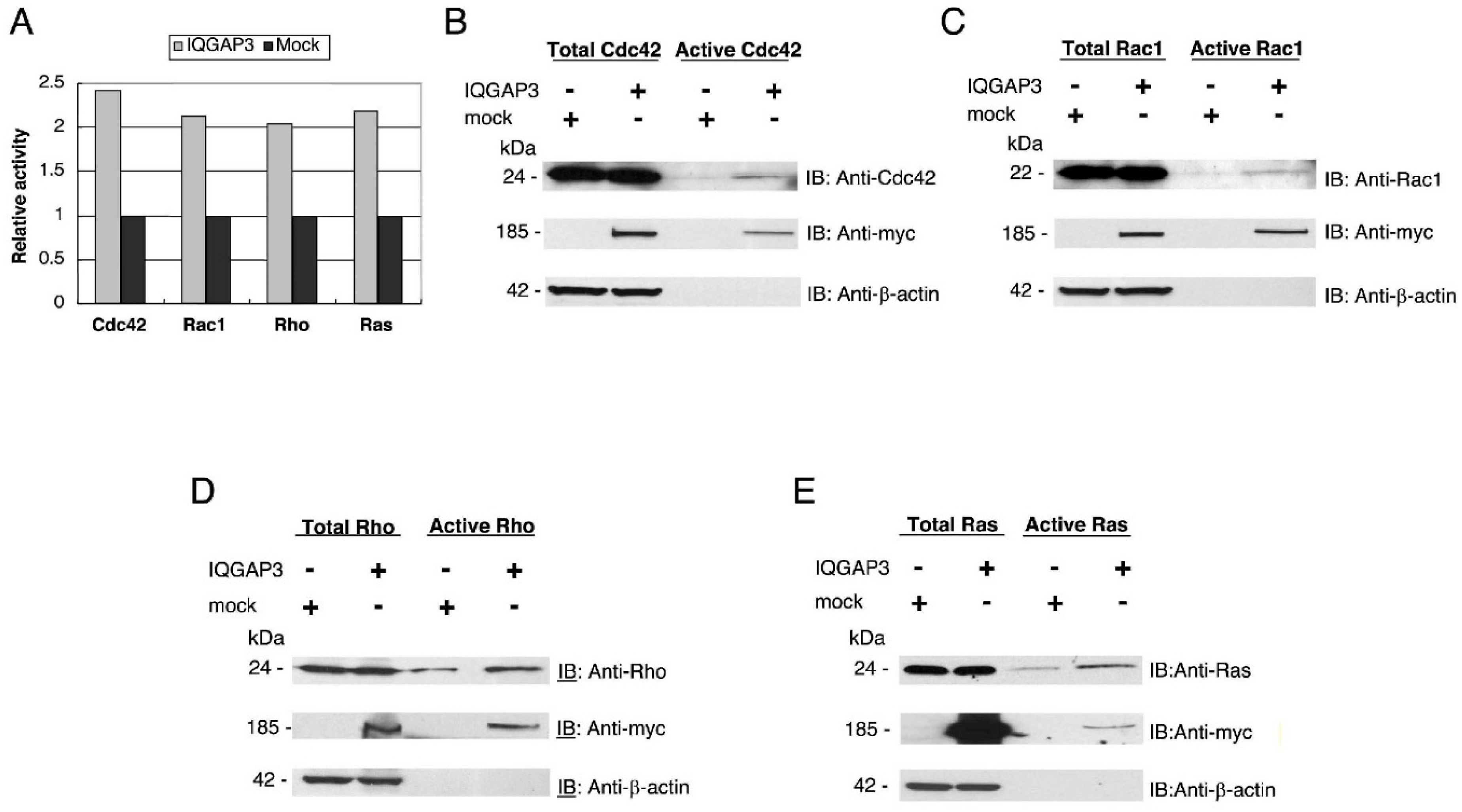
Small GTPase activation assays were performed using Thermo Scientific Pierce Active GTPase Pull-down and Detection Kits (Thermo Fisher Scientific, Waltham, MA, USA) according to the supplier’s protocols. The binding region of the downstream effector for each small GTPase was expressed as a GST-fusion protein, and used to pull down the active or GTP-bound GTPase. Extracts (2 mg) from 293T cells transfected with pcDNA3.1-myc/His-IQGAP3, or pcDNA3.1-myc/His in 700 μL lysis/binding/washing buffer were incubated with GST-Raf1-Ras Binding Domain (for Ras), GST-Pak1-p21 Binding Domain (for Cdc42 and Rac1), and GST-Rhotekin-Rho Binding Domain (for Rho) in the presence of SwellGel Immobilized Glutathione at 4 °C for 1 h in a spin column. The resins were washed three times and eluted by boiling at 95 °C for 5 min. The eluates were separated by SDS-PAGE, and active Ras, Cdc42, Rac1, or Rho were detected by western blotting using specific antibodies. These assays were performed at least 3 times to ensure the validity of the results. Phosphorimager quantification analysis was carried out (Molecular Imager FX, Bio-Rad Laboratories, Hercules, CA, USA), and relative Western blotting band intensities (the levels of active small GTPases in IQGAP3-transfected cells/those in mock-transfected cells) were quantified.
2.11. Co-Immunocytochemical Staining
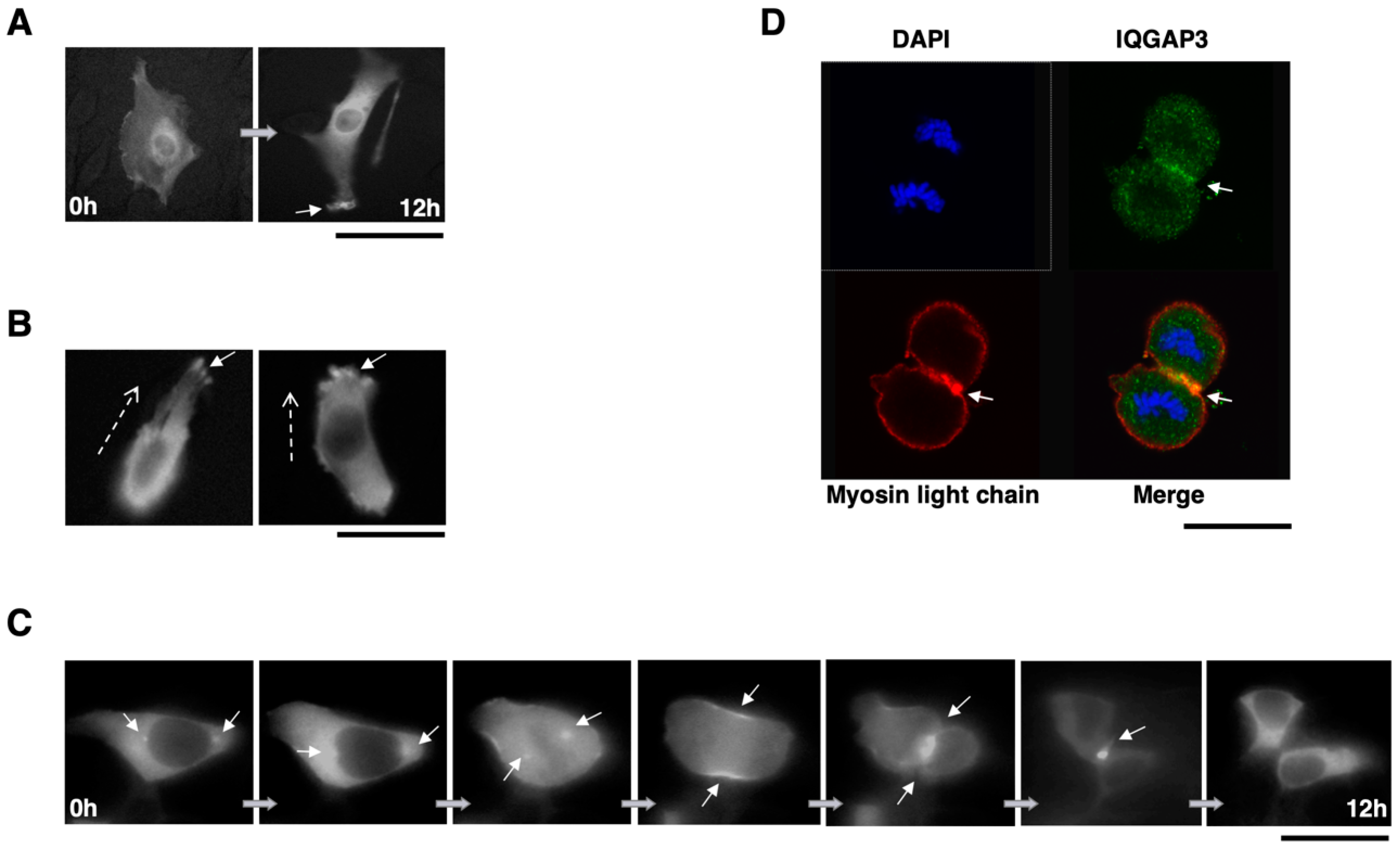
Cells expressing endogenous IQGAP3 or myc-tagged IQGAP3 were fixed and stained with in-house anti-IQGAP3 or anti-c-myc (9E10) (Santa Cruz Biotechnologies, Santa Cruz, CA, USA), respectively. Subsequently, Alexa Fluor® 488 goat anti-rabbit IgG secondary antibody or Alexa Fluor® 488 goat anti-mouse IgG secondary antibody (Molecular Probes, Invitrogen, Carlsbad, CA, USA) were used for visualization. Nuclei were counterstained with 4′,6′-diamidine-2′-phenylindole dihydrochloride (DAPI). Actin cytoskeleton was visualized with Alexa Fluor® 594 phalloidin (Molecular Probes, Invitrogen, Carlsbad, CA, USA). Monoclonal Anti-Myosin (Light Chains 20 kDa, clone MY21, Sigma-Aldrich) and Alexa Fluor® 594 goat anti-mouse IgM (μ chain) secondary antibody (Molecular Probes, Invitrogen, Carlsbad, CA, USA) were used to localize myosin light chain in cells. Fluorescent images were obtained with a TCS SP2 AOBS Spectral Confocal Scanning System (Leica, Bensheim, Germany).
2.12. Time-Lapse Microscopy
We prepared plasmids expressing EGFP-fused IQGAP3 (EGFP-IQGAP3) protein using pEGFP-N1 vector (BD Biosciences Clontech, Mountain View, CA, USA), and transfected them into NIH3T3 cells on a 35 mm glass-bottom culture dish (MatTek Inc., Ashland, MA, USA). Time-lapse images were captured every 15 min for 8–16 h by the Live Cell Imaging System (Power IX81, Olympus, Tokyo, Japan).
4. Discussion
In this study, we have demonstrated that the up-regulation of IQGAP3 promotes invasion and migration in gastric cancer cells, most likely through interacting with e-cadherin and/or β-catenin in association with the reduction of cell-cell adhesion. The co-localization of IQGAP3 and f-actin was also observed at the leading edge of cells, indicating the involvement of IQGAP3 in migration. In contrast, knockdown of IQGAP3 in two gastric cancer cell lines successfully reduced the number of invading cells and colonies in matrigel invasion and soft agar assays, respectively. Furthermore, IQGAP3 shared its interacting proteins, Cdc42, Rac1, Ras, and Rho, with IQGAP1. These results agree with our hypothesis that IQGAP3 may act as an oncogene in gastric cancer. In addition, we revealed that IQGAP3 possibly plays an important role in cytokinesis in gastric cancer cells.
IQGAP1 is frequently up-regulated in a subset of gastric, colon, and ovarian cancers and is localized on chromosome band 15q26, a region that shows amplification in gastric cancer. In addition, two gastric cancer cell lines with
IQGAP1 overexpression had amplification of
IQGAP1 [
38]. The chromosome region 1q22 that contains
IQGAP3 was also found to have copy number gains in gastric cancer [
39]. Recently, Wu et al. [
24] showed that poor prognosis of lung cancer was associated with increased expression of
IQGAP3 resulting from copy number gains. Therefore, genomic amplification might also play a role in
IQGAP3’s elevated expression in gastric cancer. Just recently, elevated expression of IQGAP3 was identified by immunohistochemistry (IHC) staining in the 165 gastric cancer tissues, and was significantly correlated with poor survival [
40]. Of note, there was no significant association between IQGAP3 levels and tumor staging, or lymph node involvement in our IHC study, which may be due to the small sample size.
Kumar et al. has previously examined the expression of
IQGAP3 by analyzing large scale cancer databases including TCGA [
32]. They found
IQGAP3 is overexpressed in gastric cancer and the overall survival is significantly lower in patients who have high
IQGAP3 expression. In this study, we have used several web-based software to further conduct in-depth clinical correlation analyses using the TCGA database. In line with their results, we identified the overexpression of
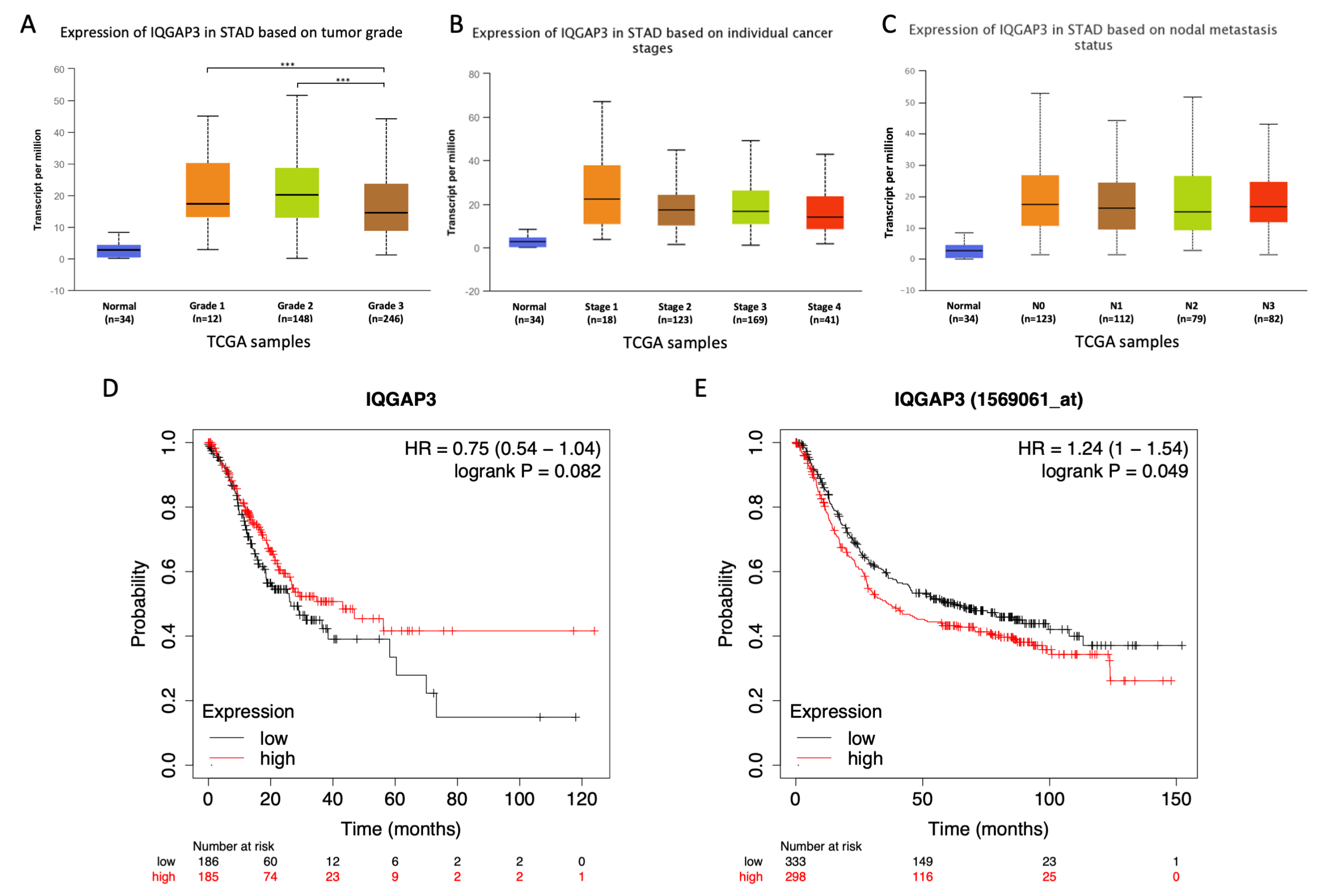
IQGAP3 in the gastric cancer samples of both subtypes, diffuse and intestinal. However, we did not observe differences in the expression of
IQGAP3 between cancer stages, nodal metastasis stages, and tumor grades. Interestingly, we observed the elevated expression of
IQGAP3 from a very early stage of cancer progression (stage 1) and nodal metastasis (N0) (
Figure 2B,C). Although we did not observe the increased expression of
IQGAP3 in relation to the more advanced stages and nodal metastasis status, it is likely that the initial induction of
IQGAP3 expression may play important roles in early gastric carcinogenesis. We, thus, proposed that
IQGAP3 expression may be used as a potential biomarker for early detection of gastric cancer. However, further studies are required to elucidate this possibility.
We also observed different trends regarding the effect of
IQGAP3 expression on overall survival when using different datasets, namely chip-based microarray dataset and sequencing-base RNA-Seq dataset. This observation was first mentioned by Kumar et al. [
32]. It may result from a smaller sample size of the RNA-Seq dataset or some unknown specific characteristics of patients included in these two cohorts. Nevertheless, as our patients were all Asians, we further looked into the overall survival rate of Asian patients with different expression levels of
IQGAP3 in the TCGA database. We did not observe significantly different overall survival rates with regard to differential
IQGAP3 expression, which may again be due to the small number of the Asian population in the database. In light of these findings, future large-scale studies are necessary to address whether higher expression of
IQGAP3 is associated with poorer prognosis in gastric cancer patients, particularly in Asian populations.
We showed a significant reduction in the adhesion of cells overexpressing IQGAP3. Fram et al. [
41] showed that IQGAP1 plays a role in the disruption of cell-cell adhesion by forming a complex with
e-cadherin. Yang et al. [
23] showed an increase of
e-cadherin expression resulted from the knockdown of IQGAP3 in the lung cancer cell line, suggesting the involvement of interaction between
e-cadherin and IQGAP3 protein in cancers. However, the interaction with β-catenin has not been reported in IQGAP3. Our results agree with the above studies and showed that the reduction of cell-cell adhesion was likely to be due to the interaction between IQGAP3 and
e-cadherin proteins, and further suggested a possible mechanism for interaction between β-catenin and IQGAP3 proteins in reducing cell-cell adhesion.
In addition to its role in morphogenesis and migration, IQGAP3 protein showed a unique subcellular localization during cytokinesis. Adachi et al. proposed that IQGAP3 regulated mammalian cell cytokinesis through interacting with anillin [
42]. The knockdown of IQGAP3 led to incomplete cleavage furrow ingression and formation of multinucleated cells. In addition, their study showed that IQGAP proteins co-localize with myosin light chain and RhoA. In agreement with this, we showed that IQGAP3 protein localized at the spindle pole body, contractile ring and concentrated at the cleavage furrow during mitosis and cytokinesis, and also co-localized with myosin light chain in gastric cancer cells. Thus, our studies on both non-cancerous and gastric cancer cell lines provided another strong supporting evidence for the involvement of IQGAP3 in cytokinesis.
Recently, Yang et al. [
23] found that
IQGAP3 had significantly higher expression levels in lung cancers, compared to adjacent non-cancerous tissues of 25 cases. They further showed that the enforced expression of IQGAP3 in Hela cells, which had a relatively low endogenous IQGAP3 expression, promoted cell proliferation, as well as the capability for migration and invasion. In addition, knockdown of IQGAP3 by siRNA in A549 lung cancer cells suppressed cell proliferation and decreased the migration and invasion ability. In line with the literature, our results showed that IQGAP3 plays important role in cell migration and invasion but not proliferation in gastric cancer, suggesting that its function in promoting cell proliferation might be tissue-specific.
The IQGAP3 protein shares high similarity with IQGAP1; it shares 84%, 46%, 73%, 85%, and 68% amino acid similarity with IQGAP1 in calponin homology domain, IQ repeats, WW domain, RasGAP-related domain, and RasGAP
c-terminus domain, respectively [
28]. IQGAP1 stimulates filopodia formation, and promotes cell motility and invasion in mammalian and cancer cells, particularly through the interaction with β-catenin,
e-cadherin, and small GTPase, i.e., Cdc42, Rac1, and RhoC. Since our study used kits including universal Rho and Ras GTPases, we are unable to distinguish the interactions between IQGAP3 and RhoA/B/C or H-Ras/R-Ras. From the literature, RhoC was proposed to regulate proliferation through interaction with IQGAP1 in gastric cancer [
20], suggesting that RhoC and IQGAP1 could stimulate the proliferation of gastric cancer. The up-regulation of RhoC has been shown to increase invasion in cancer cells [
43]. A recent study also proposed that IQGAP3 is a Cdc42 pathway-related protein [
30], since knockdown of IQGAP3 in a pancreatic cancer cell line decreased the expression of Cdc42. This is in line with our results which demonstrate that IQGAP3 interacts with Cdc42. Furthermore, one study showed the interaction between IQGAP3 and Ras [
29], while others suggested no direct interaction between the two proteins [
28]. Although a recent report has proposed that endogenous IQGAP3 does not functionally interact with Ras [
44], in our study we observed the protein-protein interactions between overexpressed exogenous IQGAP3 and endogenous Ras. This finding might be non-specific due to the use of supra-physiological concentration of IQGAP3. However, the possibility that enhanced endogenous IQGAP3 in gastric cancer may permit interaction with Ras cannot be completely ruled out. Taken together, although our results showed that IQGAP3 activated Rho and Ras, further studies are necessary to elucidate the mechanisms underlying the interactions of IQGAP3 with Rho or Ras in cancer.
IQGAP3 has also been associated with markers of apoptosis and metastasis. Shi and colleagues have shown that epithelial-mesenchymal transition (EMT) markers including
e-cadherin, fibronectin, vimentin,
n-cadherin are overexpressed in HCC cells with up-regulation of IQGAP3, while knockdown of IQGAP3 showed the reduction of the above markers [
45]. Furthermore, IQGAP3 functions as an important regulator of metastasis and EMT by constitutively activating the TGF-beta signaling pathway; hence, it may also play some roles in promoting cancer stemness through regulation of tumor microenvironment [
46]. Similar results were also observed in pancreatic cancer [
30], where reduced expression of IQGAP3 resulted in early apoptosis and subcutaneous tumor growth inhibition in the mouse model [
30].
Monteleon et al. [
26] suggested the possibility of using a “decoy” peptide of IQGAP1 and IQGAP3 as targeted therapy. They showed that squamous cell carcinoma had a significant fitness disadvantage after expressing a decoy IQ motif. In contrast, the fitness of normal tissue was not affected by expressing the decoy IQ motif. As most current therapies are very harmful to normal cells as well, targeting IQGAP proteins might be a promising approach for the development of less-toxic molecular targeted therapy in the future.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






