Characterization of Chenopodin Isoforms from Quinoa Seeds and Assessment of Their Potential Anti-Inflammatory Activity in Caco-2 Cells

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chenopodin Extraction and Purification
2.2. SDS-PAGE and IEF/SDS-PAGE
2.3. Protein Identification
2.4. Structural Predictions
2.5. Caco-2 Cell Cultivation
2.6. Transient Transfection and Immunomodulation Assay in Transfected Caco-2 Cells
2.7. Gene Expression Studies by qPCR
2.8. Determination of Saponins
2.9. Cell Vitality Assays
2.10. Statistical Analysis
3. Results
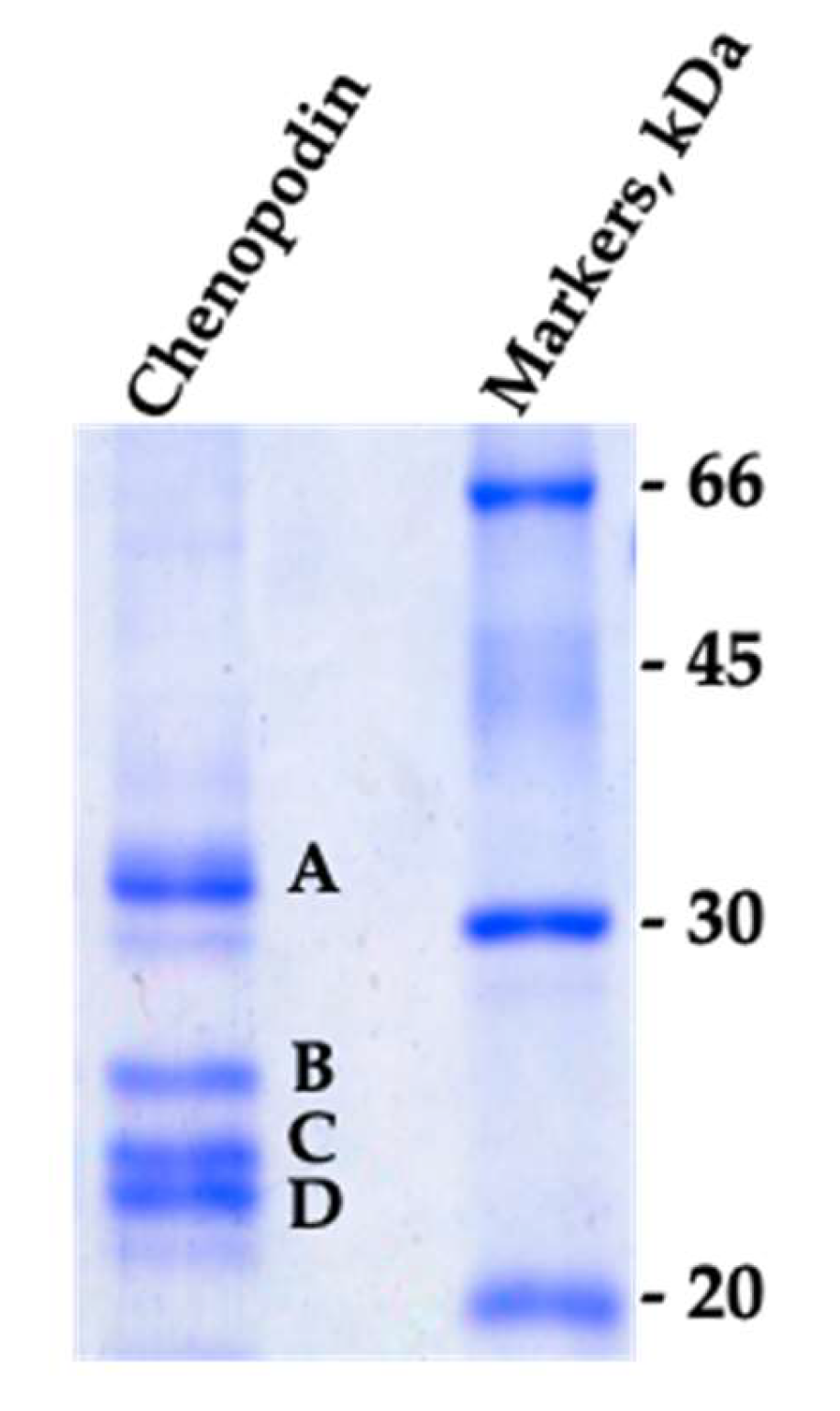
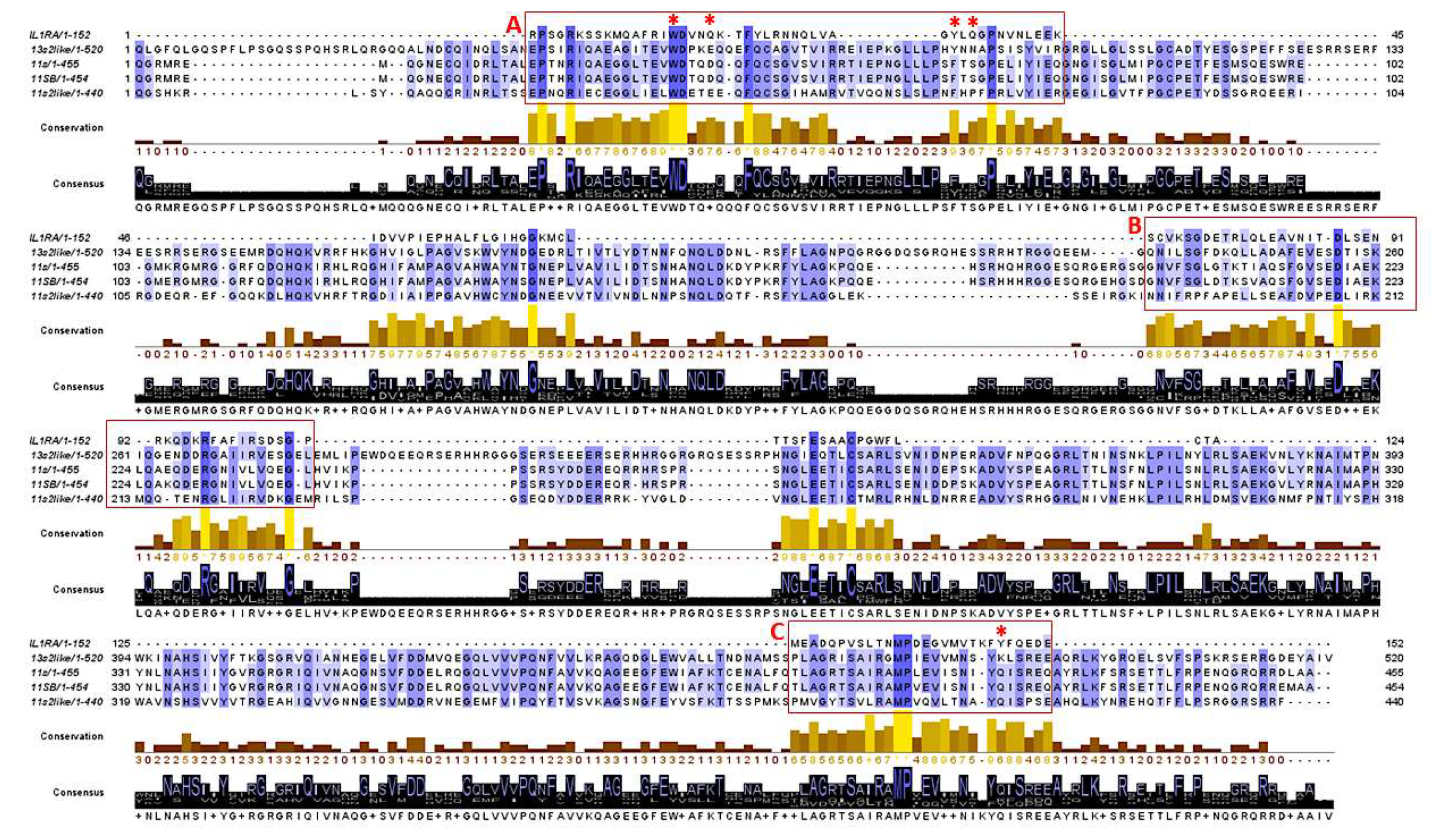
3.1. Characterization of the Isolate Chenopodin
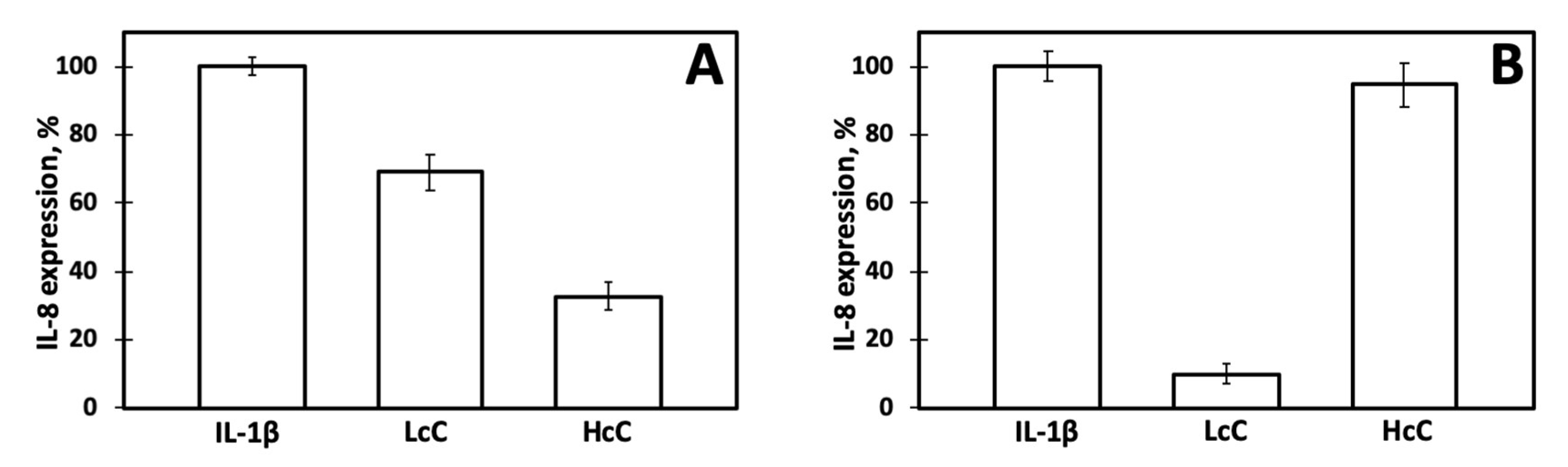
3.2. Immunomodulation Effects of Native Chenopodins
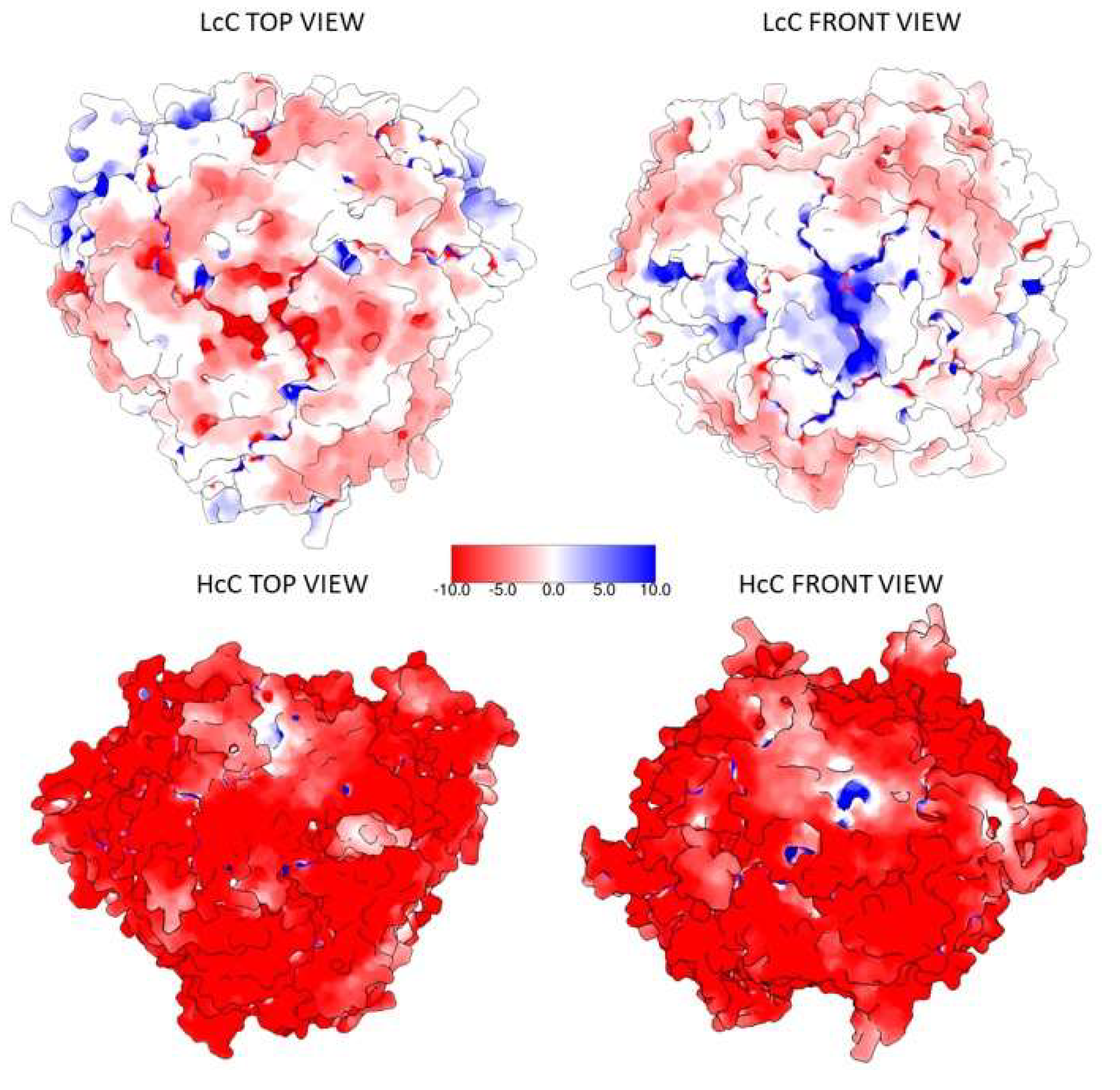
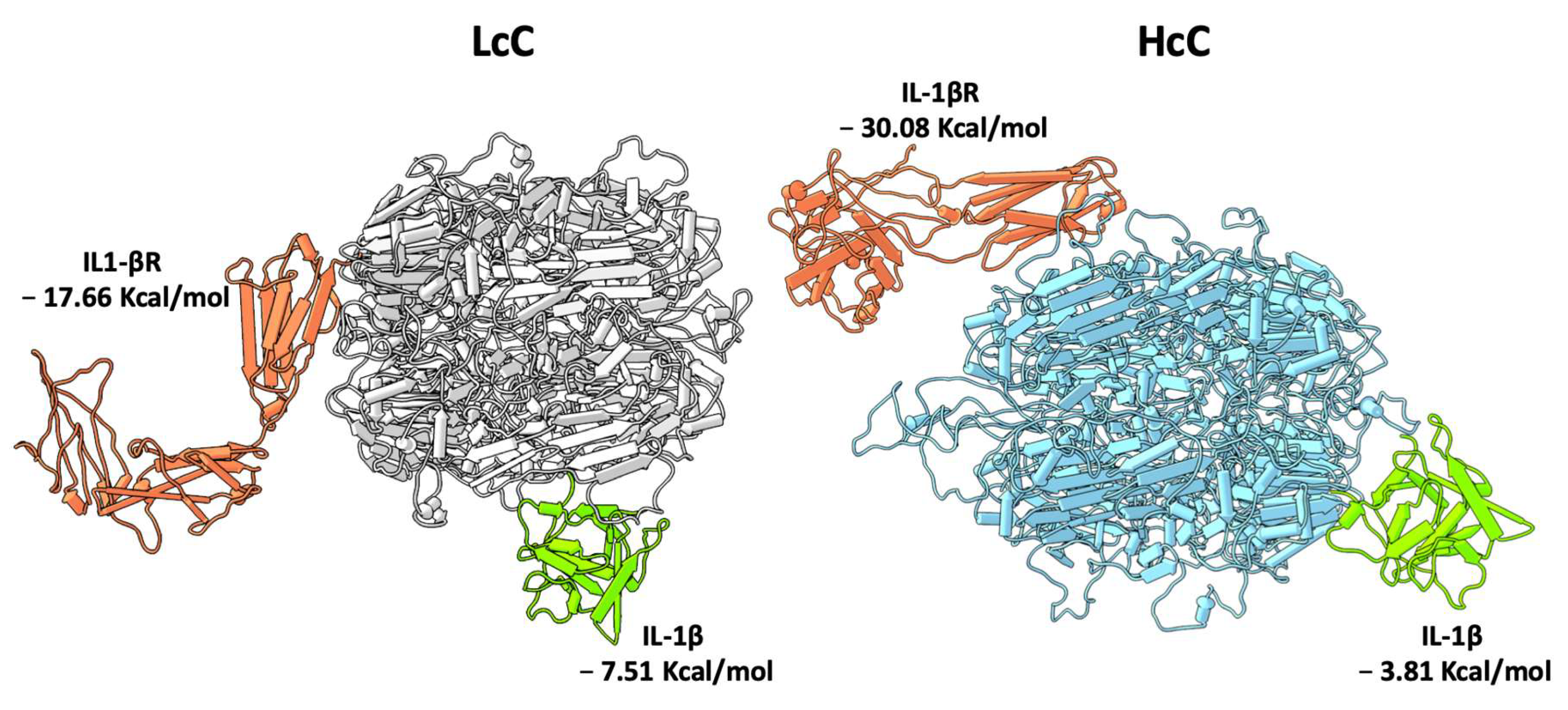
3.3. In Silico Protein–Protein Interaction Analyses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Toapanta, A.; Carpio, C.; Vilcacundo, R.; Carrillo, W. Analysis of protein isolate from quinoa (Chenopodium Quinoa Willd). Asian J. Pharm. Clin. Res. 2016, 9, 332–334. [Google Scholar]
- Repo-Carrasco, R.; Espinoza, C.; Jacobsen, S.E. Nutritional value and use of the Andean crops quinoa (Chenopodium quinoa) and kaniwa (Chenopodium pallidicaule). Food Rev. Int. 2003, 19, 179–189. [Google Scholar] [CrossRef]
- Fairbanks, D.J.; Burgener, K.W.; Robinson, L.R.; Andersen, W.R.; Ballon, E. Electrophoretic characterization of quinoa seed proteins. Plant Breed. 1990, 104, 190–195. [Google Scholar] [CrossRef]
- Utsumi, S. Plant Food Protein Engineering. Adv. Food Nutr. Res. 1992, 36, 89–208. [Google Scholar]
- Brinegar, C.; Goundan, S. Isolation and characterization of chenopodin, the 11S seed storage protein of quinoa (Chenopodium quinoa). J. Agric. Food Chem. 1993, 41, 182–185. [Google Scholar] [CrossRef]
- Srivastava, L.M. Seed food reserves and their accumulation. In Plant Growth and Development; Academic Press: Oxford, UK, 2002; pp. 503–520. [Google Scholar]
- Jacobsen, S.-E. The worldwide potential for quinoa (Chenopodium quinoa, Willd.). Food Rev. Int. 2003, 19, 167–177. [Google Scholar] [CrossRef]
- Bazile, D.; Jacobsen, S.-E.; Verniau, A. The global expansion of quinoa: Trends and limits. Front. Plant Sci. 2016, 7, 622. [Google Scholar] [CrossRef] [Green Version]
- Gordillo-Bastidas, E.; Díaz-Rizzolo, D.; Roura, R.; Rizzolo, D.A.D.; Massanés, T.; Gomis, R. Quinoa (Chenopodium quinoa Willd), from nutritional value to potential health benefits: An integrative review. J. Nutr. Food Sci. 2016, 6, 497. [Google Scholar]
- Graf, B.L.; Rojas-Silva, P.; Rojo, L.E.; Delatorre-Herrera, J.; Baldeón, M.E.; Raskin, I. Innovations in health value and functional food development of quinoa (Chenopodium quinoa Willd.). Compr. Rev. Food Sci. Food Saf. 2015, 14, 431–445. [Google Scholar] [CrossRef] [Green Version]
- Maradini-Filho, A.M. Quinoa: Nutritional aspects. J. Nutraceuticals Food Sci. 2017, 2, 1–5. [Google Scholar]
- Liu, R.H. Dietary bioactive compounds and their health implications. J. Food Sci. 2013, 78 (Suppl. S1), A18–A25. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, P.V.M.; Andrade, P.A.; Hermsdorff, H.H.M.; Dos Santos, C.A.; Cotta, R.M.M.; Estanislau, J.A.S.G.; Campos, A.A.O.; Rosa, C.O.B. Dietary non-nutrients in the prevention of non-communicable diseases: Potentially related mechanisms. Nutrition 2019, 66, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Díaz, A.; Del-Toro-Sánchez, C.L.; Rodríguez-Figueroa, J.C.; Valdéz-Hurtado, S.; Wong-Corral, F.J.; Borboa-Flores, J.; González-Osuna, M.F.; Perez-Perez, L.M.; González-Vega, R.I. Legume proteins as a promising source of anti-inflammatory peptides. Curr. Protein Pept. Sci 2019, 20, 1204–1217. [Google Scholar]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Ben-Neriah, Y.; Karin, M. Inflammation meets cancer, with NF-κB as the matchmaker. Nat. Immunol. 2011, 12, 715–723. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Ariel, A.; Timor, O. Hanging in the balance: Endogenous anti-inflammatory mechanisms in tissue repair and fibrosis. J. Pathol. 2013, 229, 250–263. [Google Scholar] [CrossRef]
- Zhang, L.; Wei, X.; Zhang, R.; Petitte, J.N.; Si, D.; Li, Z.; Cheng, J.; Du, M. Design and development of a novel peptide for treating intestinal inflammation. Front. Immunol. 2019, 10, 1841. [Google Scholar] [CrossRef] [Green Version]
- Artusa, V. Development of a RT-qPCR Assay to Determine the Anti-Inflammatory Potential of Quinoa (Chenopodium quinoa Willd.) Seed Proteins in Human Intestinal Caco-2 Cells. Master’s Thesis, Università degli Studi di Milano, Milano, Italy, 2018. [Google Scholar]
- Scarafoni, A.; Giani, D.; Cerletti, P. An endopeptidase in dormant lupin seed. Phytochemistry 1992, 37, 3715–3723. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Scarafoni, A.; Ronchi, A.; Prinsi, B.; Espen, L.; Assante, G.; Venturini, G.; Duranti, M. The proteome of exudates from germinating Lupinus albus seeds is secreted through a selective dual-step process and contains proteins involved in plant defense. FEBS J. 2013, 280, 1443–1459. [Google Scholar] [CrossRef] [Green Version]
- Hellman, U.; Wernstedt, C.; Gonez, J.; Heldin, C. Improvement of an ‘In-Gel’ digestion procedure for the micropreparation of internal protein fragments for amino acid sequencing. Anal. Biochem. 1995, 224, 451–455. [Google Scholar] [CrossRef]
- Bona, E.; Massa, N.; Novello, G.; Boatti, L.; Cesaro, P.; Todeschini, V.; Magnelli, V.; Manfredi, M.; Marengo, E.; Mignone, F.; et al. Metaproteomic characterization of the Vitis vinifera rhizosphere. FEMS Microbiol. Ecol. 2019, 95, fiy204. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NY, USA, 2005; pp. 571–607. [Google Scholar]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucl. Acids Res. 2018, 46, 296–303. [Google Scholar] [CrossRef] [Green Version]
- Baker, N.; Sept, D.; Joseph, S.; Holst, M.; McCammon, J. Electrostatics of nanosystems: Application to microtubules and the ribosome. Proc. Natl. Acad. Sci. USA 2001, 98, 10037–10041. [Google Scholar] [CrossRef] [Green Version]
- Dolinsky, T.J.; Czodrowski, P.; Li, H.; Nielsen, J.E.; Jensen, J.H.; Klebe, G.; Baker, N.A. PDB2PQR: Expanding and upgrading automated preparation of biomolecular structures for molecular simulations. Nucl. Acids Res. 2007, 35, 522–525. [Google Scholar] [CrossRef]
- Sanner, M.F.; Olson, A.J.; Spehner, J.C. Reduced surface: An efficient way to compute molecular surfaces. Biopolymers 1996, 38, 305–320. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera-a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Duhovny, D.; Nussinov, R.; Wolfson, H.J. Efficient unbound docking of rigid molecules. In Algorithms in bioinformatics; Guigó, R., Gusfield, D., Eds.; Springer: Berlin, Germany, 2002; Volume 2452, pp. 185–200. [Google Scholar]
- Schneidman-Duhovny, D.; Inbar, Y.; Nussinov, R.; Wolfson, H.J. PatchDock and SymmDock: Servers for rigid and symmetric docking. Nucl. Acids Res. 2005, 33, 363–367. [Google Scholar] [CrossRef] [Green Version]
- Andrusier, N.; Nussinov, R.; Wolfson, H.J. FireDock: Fast interaction refinement in molecular docking. Proteins 2007, 69, 139–159. [Google Scholar] [CrossRef]
- Di Tommaso, P.; Moretti, S.; Xenarios, I.; Orobitg, M.; Montanyola, A.; Chang, J.M.; Taly, J.F.; Notredame, C. T-Coffee: A web server for the multiple sequence alignment of protein and RNA sequences using structural information and homology extension. Nucl. Acids Res. 2011, 39, 13–17. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2-A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Moran, T.; Swanson, E.; Julian, C.; Harris, J.; Bonen, D.K.; Hedl, M.; Nicolae, D.L.; Abraham, C.; Cho, J.H. Regulation of IL-8 and IL-1 expression in Crohn’s disease associated NOD2/CARD15 mutations. Hum. Mol. Genet. 2004, 13, 1715–1725. [Google Scholar] [CrossRef] [Green Version]
- Bottero, V.; Imbert, V.; Frelin, C.; Formento, J.L.; Peyron, J.F. Monitoring NF-κB transactivation potential via Real-Time PCR quantification of IκB-α Gene Expression. Mol. Diagn. 2003, 7, 187–194. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Aoun, M.; Corsetto, P.A.; Nugue, G.; Montorfano, G.; Ciusani, D.C.; Hogarth, P.; Gregory, A.; Hayflick, S.; Zorzi, G.; Rizzo, A.M.; et al. Changes in red blood cell membrane lipid composition: A new perspective into the pathogenesis of PKAN. Mol. Genet. Metab. 2017, 121, 180–189. [Google Scholar] [CrossRef]
- McDonald, J.G.; Thompson, B.M.; Mc Crum, E.C.; Russell, D.W. Extraction and analysis of sterols in biological matrices by high performance liquid chromatography electrospray ionization mass spectrometry. Methods Enzymol. 2007, 432, 145–170. [Google Scholar]
- Barbiroli, A.; Capraro, J.; Marulo, S.; Gamba, M.; Scarafoni, A. Effects on the Caco-2 cells of a hypoglycemic protein from lupin seeds in a solution and adsorbed on polystyrene nanoparticles to mimic a complex food matrix. Biomolecules 2019, 9, 606. [Google Scholar] [CrossRef] [Green Version]
- Daig, R.; Rogler, G.; Aschenbrenner, E.; Vogl, D.; Falk, W.; Gross, V.; Schölmerich, J.; Andus, T. Human intestinal epithelial cells secrete interleukin-1 receptor antagonist and interleukin-8 but not interleukin-1 or interleukin-6. Gut 2000, 46, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Philpott, D.J.; Yamaoka, S.; Israël, A.; Sansonetti, P.J. Invasive Shigella flexneri activates NF-kappa B through a lipopolysaccharide-dependent innate intracellular response and leads to IL-8 expression in epithelial cells. J. Immunol. 2000, 165, 903–914. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, D.E.; Ho, Y.S.; Lightfoot, D.J.; Schmöckel, S.M.; Li, B.; Borm, T.J.A.; Ohyanagi, H.; Mineta, K.; Michell, C.T.; Saber, N.; et al. The genome of Chenopodium quinoa. Nature 2017, 542, 307–312. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Tsao, R. Phytochemicals in quinoa and amaranth grains and their antioxidant, anti-inflammatory, and potential health beneficial effects: A review. Mol. Nutr. Food Res. 2017, 61, 1600767. [Google Scholar] [CrossRef]
- Yao, Y.; Yang, X.; Shi, Z.; Ren, G. Anti-inflammatory activity of saponins from quinoa (Chenopodium quinoa Willd.) seeds in lipopolysaccharide-stimulated RAW 264.7 macrophages cells. J. Food Sci. 2014, 79, 1018–1023. [Google Scholar] [CrossRef]
- Vigers, G.P.A.; Anderson, L.J.; Caffes, P.; Brandhuber, B.J. Crystal structure of the type-I interleukin-1 receptor complexed with interleukin-1β. Nature 1997, 386, 190–194. [Google Scholar] [CrossRef]
- Dinarello, C.A. The interleukin-1 family: 10 years of discovery. FASEB J. 1994, 8, 1314–1325. [Google Scholar] [CrossRef] [Green Version]
- Greenfeder, S.A.; Nunes, P.; Kwee, L.; Labow, M.; Chizzonite, R.A.; Ju, G. Molecular cloning and characterization of a second subunit of the interleukin 1 receptor complex. J. Biol. Chem. 1995, 270, 13757–13765. [Google Scholar] [CrossRef] [Green Version]
- Castañeda, S.; Atienza-Mateo, B.; Martín-Varillas, J.L.; Serra López-Matencio, J.M.; González-Gay, M.A. Anakinra for the treatment of adult-onset Still’s disease. Expert Rev. Clin. Immunol. 2018, 14, 979–992. [Google Scholar] [CrossRef] [Green Version]
- Schreuder, H.; Tardif, C.; Trump-Kallmeyer, S.; Soffientini, A.; Sarubbi, E.; Akeson, A.; Bowlin, T.; Yanofsky, S.; Barrett, R.W. A new cytokine-receptor binding mode revealed by the crystal structure of the IL-1 receptor with an antagonist. Nature 1997, 386, 194–200. [Google Scholar] [CrossRef]
- Ren, G.; Zhu, Y.; Shi, Z.; Li, J. Detection of lunasin in quinoa (Chenopodium quinoa Willd.) and the in vitro evaluation of its antioxidant and anti-inflammatory activities. J. Sci. Food Agric. 2017, 97, 4110–4116. [Google Scholar] [CrossRef]
- Hernández-Ledesma, B.; Hsieh, C.C.; de Lumen, B.O. Antioxidant and anti-inflammatory properties of cancer preventive peptide lunasin in RAW 264.7 macrophages. Biochem. Biophys. Res. Commun. 2009, 390, 803–808. [Google Scholar] [CrossRef]
- Cam, A.; de Mejia, E.G. RGD-peptide lunasin inhibits Akt-mediated NF-κB activation in human macrophages through interaction with the αVβ3 integrin. Mol. Nutr. Food Res. 2012, 56, 1569–1581. [Google Scholar] [CrossRef]
- Aguzzi, M.S.; Fortugno, P.; Giampietri, C.; Ragone, G.; Capogrossi, M.C.; Facchiano, A. Intracellular targets of RGDS peptide in melanoma cells. Mol. Cancer 2010, 9, 84. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Ye, F.; Ginsberg, M.H. Regulation of integrin activation. Annu. Rev. Cell Dev. Biol. 2011, 27, 321–345. [Google Scholar] [CrossRef]
- Humtsoe, J.O.; Bowling, R.A., Jr.; Feng, S.; Wary, K.K. Murine lipid phosphate phosphohydrolase-3 acts as a cell-associated integrin ligand. Biochem. Biophys. Res. Commun. 2005, 335, 906–919. [Google Scholar] [CrossRef]
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed storage proteins: Structures and biosynthesis. Plant Cell 1995, 7, 945–956. [Google Scholar]
- Muto, E.; Dell’Agli, M.; Sangiovanni, E.; Mitro, N.; Fumagalli, M.; Crestani, M.; De Fabiani, E.; Caruso, D. Olive oil phenolic extract regulates interleukin-8 expression by transcriptional and posttranscriptional mechanisms in Caco-2 cells. Mol. Nutr. Food Res. 2015, 59, 1217–1221. [Google Scholar] [CrossRef]
- Varilek, G.W.; Neil, G.A.; Bishop, W.P. Caco-2 cells express type I interleukin-1 receptors: Ligand binding enhances proliferation. Am. J. Physiol. 1994, 267, G1101–G1107. [Google Scholar] [CrossRef]
- Böcker, U. Influence of therapeutic intervention with interleukins on epithelial cell function. In Cytokines and cell Homeostasis in the Gastroinstestinal Tract, Proceedings of Falk Symposium 113 Held in Regensburg, Germany, 16–18 September 1999; Andus, T., Rogler, G., Schlottmann, K., Frick, E., Adler, G., Schmiegel, W., Zeitz, M., Schölmerich, J., Eds.; Kluwer Academic Publishers: Berlin, Germany, 1999; pp. 61–71. [Google Scholar]
- Klampfer, L. Cytokines, inflammation and colon cancer. Curr. Cancer Drug Targets. 2011, 11, 451–464. [Google Scholar] [CrossRef]
- Leonard, F.; Collnot, E.M.; Lehr, C.M. A three-dimensional coculture of enterocytes, monocytes and dendritic cells to model inflamed intestinal mucosa in vitro. Mol. Pharm. 2010, 7, 2103–2119. [Google Scholar] [CrossRef]
- Romier, B.; Van De Walle, J.; During, A.; Larondelle, Y.; Schneider, Y.J. Modulation of signalling nuclear factor-kappaB activation pathway by polyphenols in human intestinal Caco-2 cells. Br. J. Nutr. 2008, 100, 542–551. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.C. Non-canonical NF-κB signaling pathway. Cell Res. 2011, 21, 71–85. [Google Scholar] [CrossRef] [Green Version]
- Chalamaiah, M.; Yu, W.; Wu, J. Immunomodulatory and anticancer protein hydrolysates (peptides) from food proteins: A review. Food Chem. 2018, 245, 205–222. [Google Scholar] [CrossRef]
- Iskandar, M.M.; Dauletbaev, N.; Kubow, S.; Mawji, N.; Lands, L.C. Whey protein hydrolysates decrease IL-8 secretion in lipopolysaccharide (LPS)-stimulated respiratory epithelial cells by affecting LPS binding to Toll-like receptor 4. Br. J. Nutr. 2013, 110, 58–68. [Google Scholar] [CrossRef] [Green Version]
- Chahal, D.S.; Sivamani, R.K.; Isseroff, R.R.; Dasu, M.R. Plant-based modulation of Toll-like receptors: An emerging therapeutic model. Phytother Res. 2013, 27, 1423–1438. [Google Scholar] [CrossRef]
- Capraro, J.; Clemente, A.; Rubio, L.A.; Magni, C.; Scarafoni, A.; Duranti, M. Assessment of the lupin seed glucose-lowering protein intestinal absorption by using in vitro and ex vivo models. Food Chem. 2011, 125, 1279–1283. [Google Scholar] [CrossRef]
- Hoese, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, G.A.; Opazo-Navarrete, M.; Meurs, M.; Minor, M.; Sala, G.; van Boekel, M.; Stieger, M.; Janssen, A.E. Denaturation and in vitro gastric digestion of heat-treated quinoa protein isolates obtained at various extraction pH. Food Biophys. 2016, 11, 184–197. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Band | Accession ID | Description | MW |
|---|---|---|---|
| A | AAS67036.1 | 11S seed storage globulin | 54007 |
| ABI94736.1 | 11S seed storage globulin B | 53942 | |
| XP_021768838.1 | 11S globulin seed storage protein 2-like | 52806 | |
| XP_021752668.1 | 13S globulin seed storage protein 2-like | 61191 | |
| B | XP_021752668.1 | 13S globulin seed storage protein 2-like | 61191 |
| C | AAS67036.1 | 11S seed storage globulin | 54007 |
| ABI94736.1 | 11S seed storage globulin B | 53942 | |
| XP_021768838.1 | 11S globulin seed storage protein 2-like | 52806 | |
| XP_021752668.1 | 13S globulin seed storage protein 2-like | 61191 | |
| D | AAS67036.1 | 11S seed storage globulin | 54007 |
| ABI94736.1 | 11S seed storage globulin B | 53942 |
| Accession ID | Description | pI | Charge |
|---|---|---|---|
| XP_021752668.1 | 13S globulin seed storage protein 2-like | 5.96 | −101 e |
| XP_021768838.1 | 11S globulin seed storage protein 2-like | 6.35 | −79 e |
| ABI94736.1 | 11S seed storage globulin B | 6.47 | −45 e |
| AAS67036.1 | 11S seed storage globulin | 7.34 | −38 e |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capraro, J.; De Benedetti, S.; Di Dio, M.; Bona, E.; Abate, A.; Corsetto, P.A.; Scarafoni, A. Characterization of Chenopodin Isoforms from Quinoa Seeds and Assessment of Their Potential Anti-Inflammatory Activity in Caco-2 Cells. Biomolecules 2020, 10, 795. https://doi.org/10.3390/biom10050795
Capraro J, De Benedetti S, Di Dio M, Bona E, Abate A, Corsetto PA, Scarafoni A. Characterization of Chenopodin Isoforms from Quinoa Seeds and Assessment of Their Potential Anti-Inflammatory Activity in Caco-2 Cells. Biomolecules. 2020; 10(5):795. https://doi.org/10.3390/biom10050795
Chicago/Turabian StyleCapraro, Jessica, Stefano De Benedetti, Marina Di Dio, Elisa Bona, Ambra Abate, Paola Antonia Corsetto, and Alessio Scarafoni. 2020. "Characterization of Chenopodin Isoforms from Quinoa Seeds and Assessment of Their Potential Anti-Inflammatory Activity in Caco-2 Cells" Biomolecules 10, no. 5: 795. https://doi.org/10.3390/biom10050795