
Non-Targeted Metabolomic Study of Fetal Growth Restriction
Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Object Collection
2.2. Clinical Characteristics
2.3. Instruments and Reagents
2.4. Sample Preparation
2.5. Mass Spectrometry Detection
2.6. Statistical Analysis
3. Result
3.1. Clinical Characteristics
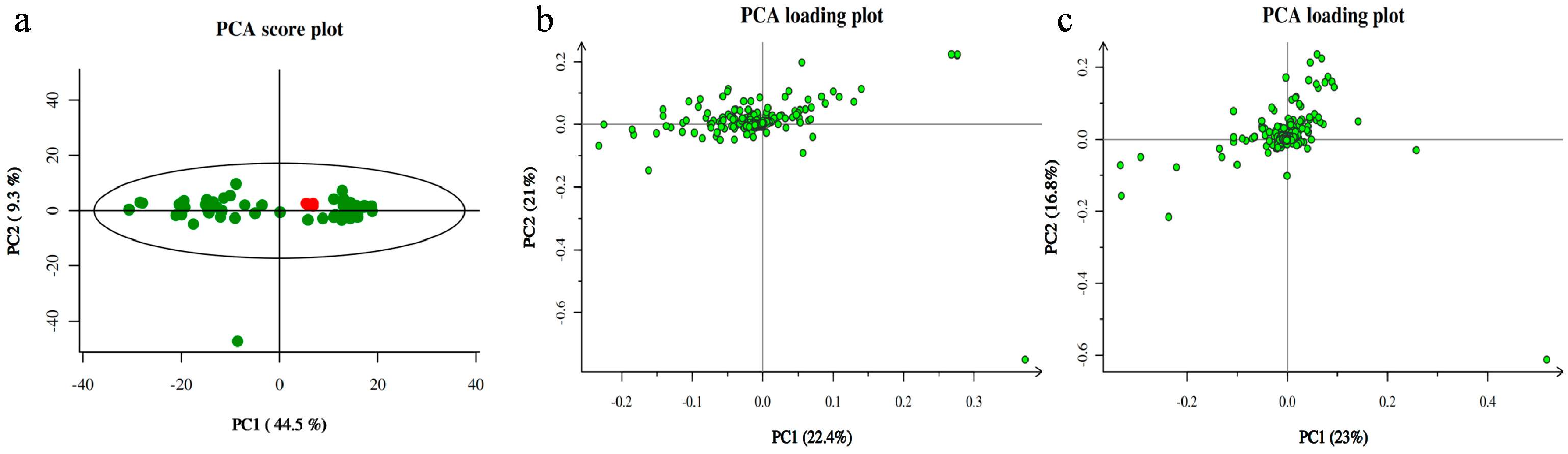
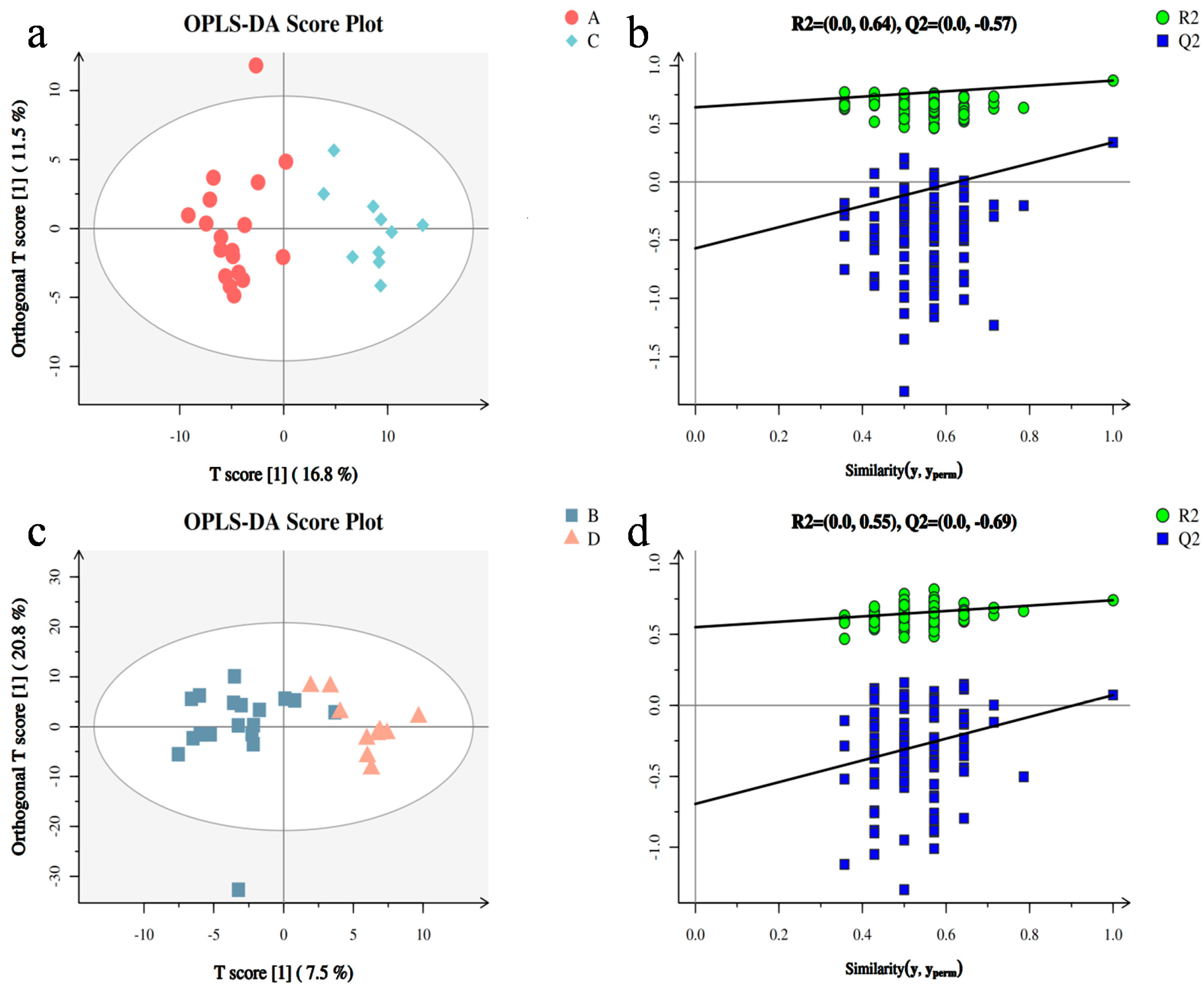
3.2. Metabolomic Analysis between Two Groups of Samples
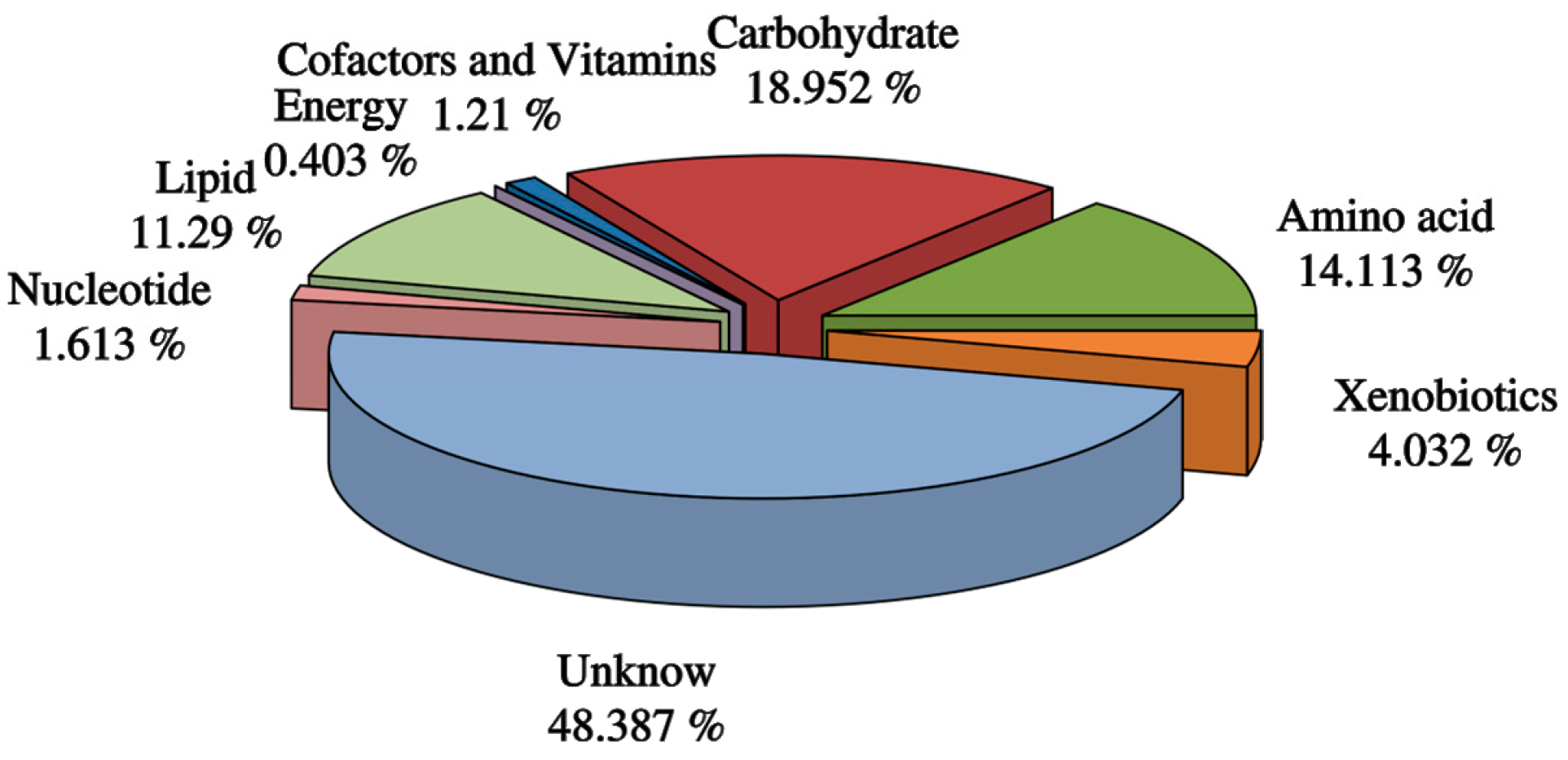
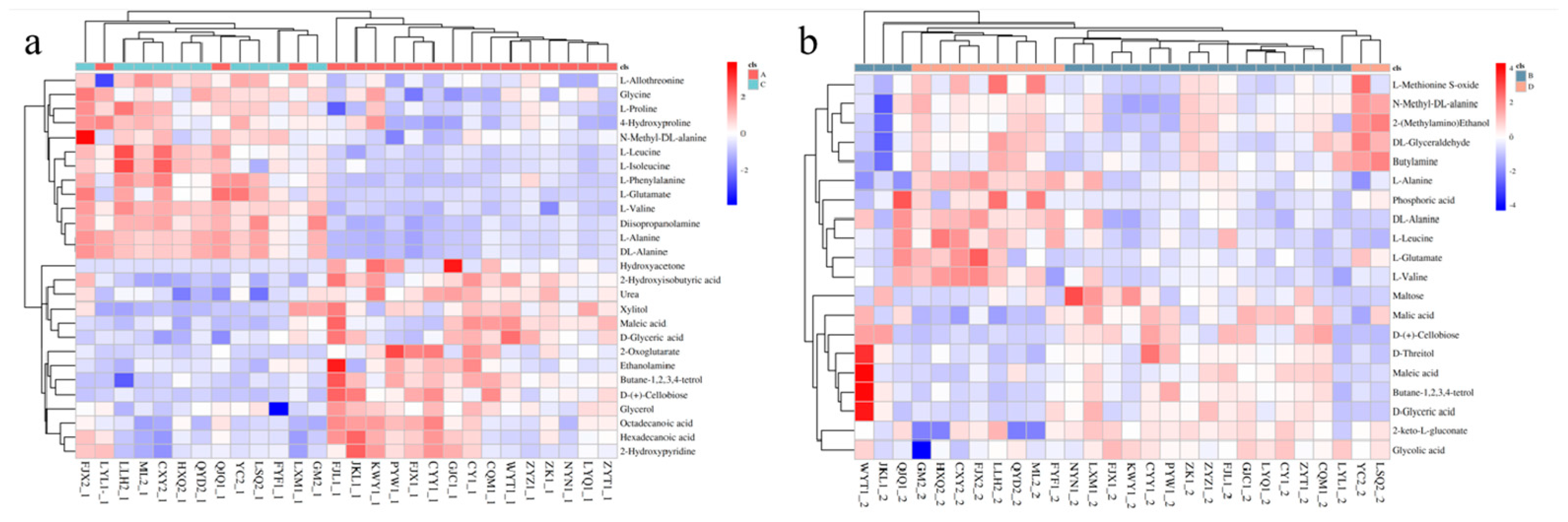
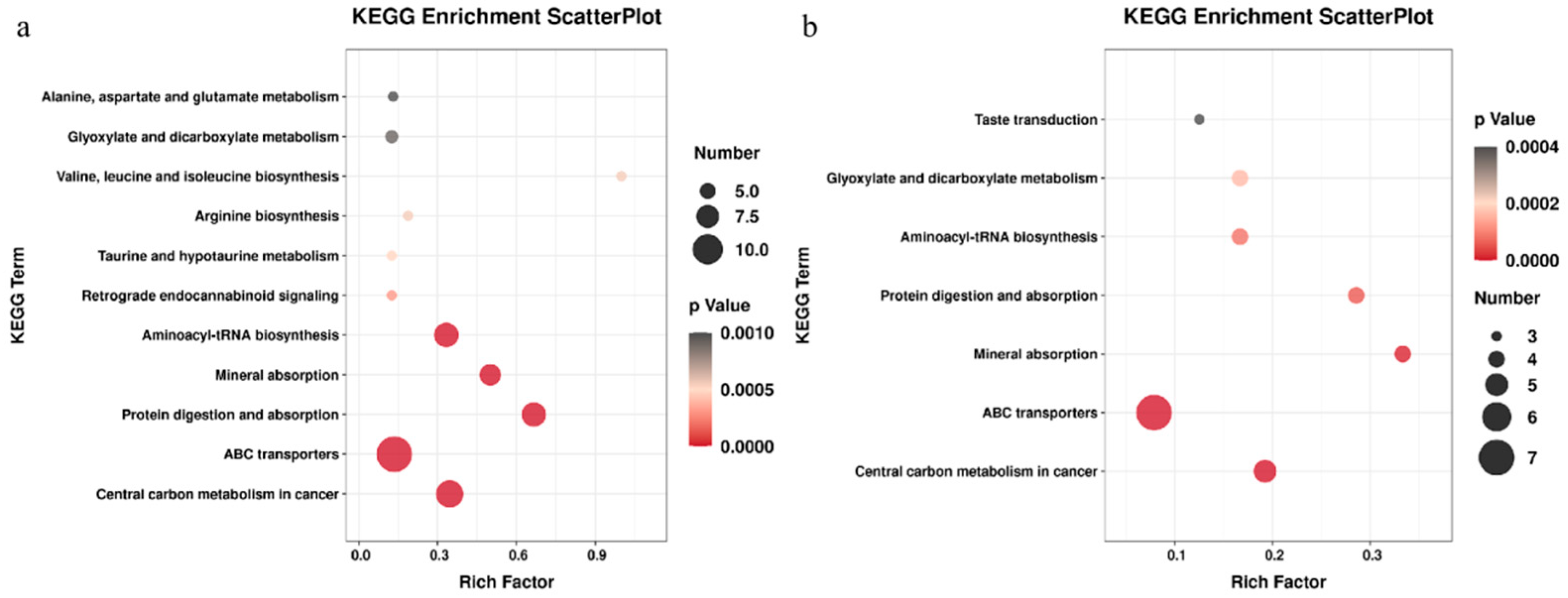
3.3. Screening of Differential Metabolites and Analysis of Metabolic Pathways
3.4. Related Metabolic Pathway Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miao, Z.; Chen, M.; Wu, H.; Ding, H.; Shi, Z. Comparative proteomic profile of the human placenta in normal and fetal growth restriction subjects. Cell Physiol. Biochem. 2014, 34, 1701–1710. [Google Scholar] [CrossRef] [Green Version]
- Nardozza, L.M.; Caetano, A.C.; Zamarian, A.C.; Mazzola, J.B.; Silva, C.P.; Marcal, V.M.; Lobo, T.F.; Peixoto, A.B.; Araujo Junior, E. Fetal growth restriction: Current knowledge. Arch. Gynecol. Obstet. 2017, 295, 1061–1077. [Google Scholar] [CrossRef] [PubMed]
- Zur, R.L.; Kingdom, J.C.; Parks, W.T.; Hobson, S.R. The Placental Basis of Fetal Growth Restriction. Obs. Gynecol. Clin. N. Am. 2020, 47, 81–98. [Google Scholar] [CrossRef] [PubMed]
- Moco, S.; Buescher, J.M. Metabolomics Going Deeper, Going Broader, Going Further. Methods Mol. Biol. 2023, 2554, 155–178. [Google Scholar]
- Sun, L.; Hu, Y.; Qi, H. A Summary of Chinese Expert Consensus on Fetal Growth Restriction (An Update on the 2019 Version). Matern.-Fetal Med. 2022, 4, 162–168. [Google Scholar]
- Cheng, Y.K.Y.; Lu, J.; Leung, T.Y.; Chan, Y.M.; Sahota, D.S. Prospective assessment of INTERGROWTH-21st and World Health Organization estimated fetal weight reference curves. Ultrasound Obs. Gynecol. 2018, 51, 792–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, N.S.; Barr, P.; Aliev, F.; Stephenson, M.; Kuo, S.I.; Chan, G.; Dick, D.M.; Edenberg, H.J.; Hesselbrock, V.; Kamarajan, C.; et al. Principal Component Analysis Reduces Collider Bias in Polygenic Score Effect Size Estimation. Behav. Genet. 2022, 52, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Triba, M.N.; Le Moyec, L.; Amathieu, R.; Goossens, C.; Bouchemal, N.; Nahon, P.; Rutledge, D.N.; Savarin, P. PLS/OPLS models in metabolomics: The impact of permutation of dataset rows on the K-fold cross-validation quality parameters. Mol. Biosyst. 2015, 11, 13–19. [Google Scholar] [CrossRef]
- Worley, B.; Powers, R. PCA as a practical indicator of OPLS-DA model reliability. Curr. Metab. 2016, 4, 97–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Lin, S.S.; Yang, J.T.; Tang, L.J.; Yu, R.Q. Detection of inborn errors of metabolism utilizing GC-MS urinary metabolomics coupled with a modified orthogonal partial least squares discriminant analysis. Talanta 2017, 165, 545–552. [Google Scholar] [CrossRef]
- Blasco, H.; Blaszczynski, J.; Billaut, J.C.; Nadal-Desbarats, L.; Pradat, P.F.; Devos, D.; Moreau, C.; Andres, C.R.; Emond, P.; Corcia, P.; et al. Comparative analysis of targeted metabolomics: Dominance-based rough set approach versus orthogonal partial least square-discriminant analysis. J. Biomed. Inf. 2015, 53, 291–299. [Google Scholar] [CrossRef]
- Dai, C.; Fei, Y.; Li, J.; Shi, Y.; Yang, X. A Novel Review of Homocysteine and Pregnancy Complications. Biomed. Res. Int. 2021, 2021, 6652231. [Google Scholar] [CrossRef]
- Vaughan, O.R.; Maksym, K.; Silva, E.; Barentsen, K.; Anthony, R.V.; Brown, T.L.; Hillman, S.L.; Spencer, R.; David, A.L.; Rosario, F.J.; et al. Placenta-specific Slc38a2/SNAT2 knockdown causes fetal growth restriction in mice. Clin. Sci. 2021, 135, 2049–2066. [Google Scholar] [CrossRef] [PubMed]
- Cleal, J.K.; Lofthouse, E.M.; Sengers, B.G.; Lewis, R.M. A systems perspective on placental amino acid transport. J. Physiol. 2018, 596, 5511–5522. [Google Scholar] [CrossRef] [Green Version]
- Regnault, T.R.; de Vrijer, B.; Galan, H.L.; Wilkening, R.B.; Battaglia, F.C.; Meschia, G. Umbilical uptakes and transplacental concentration ratios of amino acids in severe fetal growth restriction. Pediatr. Res. 2013, 73, 602–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porter, A.C.; Gumina, D.L.; Armstrong, M.; Maclean, K.N.; Reisdorph, N.; Galan, H.L.; Stabler, S.P.; Bailey, B.A.; Hobbins, J.C.; Hurt, K.J. Maternal Amino Acid Profiles to Distinguish Constitutionally Small versus Growth-Restricted Fetuses Defined by Doppler Ultrasound: A Pilot Study. Am. J. Perinatol. 2020, 37, 1084–1093. [Google Scholar] [CrossRef]
- Kim, J.; Lee, S.; Lee, J.; Park, J.C.; Kim, K.H.; Ko, J.M.; Park, S.H.; Kim, S.K.; Mook-Jung, I.; Lee, J.Y. Neurotoxicity of phenylalanine on human iPSC-derived cerebral organoids. Mol. Genet. Metab. 2022, 136, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Dai, Z.; Zhang, Y.; Chen, J.; Yang, Y.; Wu, G.; Tso, P.; Wu, Z. Maternal L-proline supplementation enhances fetal survival, placental development, and nutrient transport in micedagger. Biol. Reprod. 2019, 100, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Yue, M.; Fang, S.L.; Zhuo, Z.; Li, D.D.; Feng, J. Zinc glycine chelate absorption characteristics in Sprague Dawley rat. J. Anim. Physiol. Anim. Nutr. 2015, 99, 457–464. [Google Scholar] [CrossRef]
- Wang, X.; Geng, F.; Wu, J.; Kou, Y.; Xu, S.; Sun, Z.; Feng, S.; Ma, L.; Luo, Y. Effects of beta-conglycinin on growth performance, immunoglobulins and intestinal mucosal morphology in piglets. Arch. Anim. Nutr. 2014, 68, 186–195. [Google Scholar] [CrossRef]
- Garcia, I.S.; Teixeira, S.A.; Costa, K.A.; Marques, D.B.D.; Rodrigues, G.A.; Costa, T.C.; Guimaraes, J.D.; Otto, P.I.; Saraiva, A.; Ibelli, A.M.G.; et al. l-Arginine supplementation of gilts during early gestation modulates energy sensitive pathways in pig conceptuses. Mol. Reprod. Dev. 2020, 87, 819–834. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | FGR Group (n = 18) | Control Group (n = 10) | p Value |
|---|---|---|---|
| Age (Y) | 29.9 ± 3.9 | 31.2 ± 5.4 | 0.51 |
| Height (cm) | 1.6 ± 0.1 | 1.6 ± 0.1 | 0.81 |
| Weight (kg) | 55.4 ± 7.1 | 54.5 ± 7.4 | 0.79 |
| BMI | 22.3 ± 2.7 | 21.7 ± 2.6 | 0.59 |
| Growth meridian < 2SD, (n (%) | 72.22% (13/18) | 0 | — |
| Puncture gestational age (wks) | 30.1 ± 3.4 | 19.1 ± 1.6 | <0.01 |
| Chromosome abnormality | 11.1% (2/18) | — | — |
| Cesarean section rate (%) | 61.1% (11/18) | 40% (4/10) | — |
| Gestational age at delivery (wks) | 36.8 ± 2.1 | 38.6 ± 2.2 | 0.38 |
| Infant female, n (%) | 50% (n = 9) | 40% (n = 4) | — |
| Birth weight (kg) | 2.2 ± 0.5 | 3.3 ± 0.6 | <0.01 |
| Amniotic Fluid Supernatant | Amniotic Fluid Cell Sediment | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Metabolite | Rt | Mz | VIP | FC_A/C | p Value | Metabolite | Rt | Mz | VIP | FC_B/D | p Value |
| l-glutamic acid | 9.35 | 246.18 | 2.08 | 0.202 | <0.001 | l-glutamic acid | 9.35 | 246.18 | 1.5 | 0.432 | 0.004 |
| leucine | 7.08 | 158.16 | 2.91 | 0.388 | <0.001 | phosphoric acid | 7.05 | 299.13 | 2.83 | 0.504 | 0.041 |
| Phenylalanine | 9.45 | 218.15 | 1 | 0.391 | <0.001 | L-methionine S-oxide | 10.3 | 128.1 | 1.39 | 0.527 | 0.002 |
| isoleucine | 7.24 | 158.16 | 1.95 | 0.396 | <0.001 | L-valine | 6.68 | 144.14 | 3.18 | 0.564 | 0.008 |
| valine | 6.68 | 144.14 | 5.82 | 0.434 | <0.001 | L-Alanine | 5.84 | 116.23 | 5.1 | 0.6 | 0.008 |
| Diisopropylamine | 8.44 | 232.16 | 1.52 | 0.497 | <0.001 | l-leucine | 7.08 | 158.16 | 1.2 | 0.701 | 0.028 |
| Isothreonine | 7.85 | 218.17 | 1.8 | 0.636 | <0.001 | DL-alanine | 5.83 | 116.1 | 4.3 | 0.708 | 0.015 |
| proline | 7.31 | 142.13 | 1.98 | 0.66 | 0.004 | DL glyceraldehyde | 6.76 | 147.09 | 1.27 | 0.795 | 0.002 |
| DL-alanine | 5.83 | 116.1 | 5.53 | 0.675 | <0.001 | N-methyl-D-L-alanine | 6.36 | 130.12 | 2.78 | 0.814 | 0.01 |
| L-Alanine | 5.84 | 116.23 | 5.56 | 0.676 | <0.001 | 2-(methylamino) ethanol | 5.66 | 116.07 | 1.26 | 0.828 | 0.012 |
| 4-hydroxyproline | 8.79 | 230.19 | 1.39 | 0.694 | 0.019 | Butylamine | 5.9 | 174.15 | 1.9 | 0.865 | 0.025 |
| N-methyl-D-L-alanine | 6.36 | 130.12 | 1.48 | 0.824 | 0.005 | Glycolic acid | 5.62 | 147.09 | 1.68 | 1.196 | 0.03 |
| glycine | 7.35 | 174.14 | 1.97 | 0.876 | 0.024 | malic acid | 8.52 | 147.09 | 1.82 | 1.426 | <0.001 |
| Hexadecane acid | 11.66 | 117.05 | 1.79 | 1.095 | 0.011 | 2-Keto-L-gluconate | 10.85 | 305.28 | 1.32 | 1.515 | 0.022 |
| 2-Hydroxypyridine | 5.37 | 152.11 | 2.68 | 1.107 | 0.022 | malt dust | 14.66 | 361.23 | 1.09 | 1.731 | 0.05 |
| Octadecanoic acid | 12.57 | 117.05 | 2.25 | 1.131 | 0.002 | D-glycerate | 7.47 | 147.09 | 1.82 | 1.787 | 0.021 |
| urea | 6.88 | 147.1 | 3.16 | 1.155 | 0.023 | Maleic acid | 7.38 | 147.09 | 2.12 | 1.797 | 0.031 |
| 2-hydroxyisobutyric acid | 5.62 | 147.09 | 1.41 | 1.223 | 0.004 | Butane 1,2,3,4-tetraol | 8.62 | 147.09 | 1.28 | 1.813 | 0.01 |
| Ethanolamine | 7.07 | 174.14 | 1.53 | 1.263 | 0.032 | Threitol | 8.62 | 147.09 | 1.46 | 1.969 | 0.008 |
| glycerol | 7.06 | 147.09 | 2.45 | 1.317 | 0.006 | D-(+)-cellulose | 14.33 | 204.14 | 1.2 | 4.079 | <0.001 |
| D-glycerate | 7.47 | 147.09 | 1.67 | 1.471 | 0.014 | ||||||
| xylitol | 9.9 | 147.09 | 1.15 | 1.637 | 0.011 | ||||||
| Butane 1,2,3,4-tetraol | 8.62 | 147.09 | 1.48 | 1.711 | <0.001 | ||||||
| Maleic acid | 7.38 | 147.09 | 3 | 2.056 | 0.004 | ||||||
| 2-oxyglutaric acid | 9.07 | 147.09 | 1.17 | 2.563 | 0.007 | ||||||
| D-(+)-cellulose | 14.33 | 204.14 | 1.38 | 4.538 | 0.001 | ||||||
| Hydroxyacetone | 8.93 | 219.17 | 1.08 | 4.669 | 0.042 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, F.; Li, Z.; Xu, Y.; Huang, S.; Li, Y.; Jiang, W. Non-Targeted Metabolomic Study of Fetal Growth Restriction. Metabolites 2023, 13, 761. https://doi.org/10.3390/metabo13060761
Chen F, Li Z, Xu Y, Huang S, Li Y, Jiang W. Non-Targeted Metabolomic Study of Fetal Growth Restriction. Metabolites. 2023; 13(6):761. https://doi.org/10.3390/metabo13060761
Chicago/Turabian StyleChen, Fang, Zhi Li, Yanwen Xu, Shuang Huang, Yanqiu Li, and Weiying Jiang. 2023. "Non-Targeted Metabolomic Study of Fetal Growth Restriction" Metabolites 13, no. 6: 761. https://doi.org/10.3390/metabo13060761