
Potential Benefits of Omega-3 Polyunsaturated Fatty Acids (N3PUFAs) on Cardiovascular Health Associated with COVID-19: An Update for 2023


Abstract
:1. Introduction
2. Risk of Cardiovascular Health Deterioration during and Post-COVID-19
2.1. Epidemiological Evidence: Endothelial Dysfunction Is Linked to COVID-19-Associated Cardiac Microthrombotic Dysfunction
2.2. Prothrombotic State in COVID Infection
3. Beneficial Potential of N3PUFAs as Conjunctive Supplements for Cardiovascular Health in Acute and Post-COVID-19
3.1. N3PUFA and Endothelial Function
3.2. N3PUFA and Immunothrombosis
3.3. N3PUFA and Inflammation
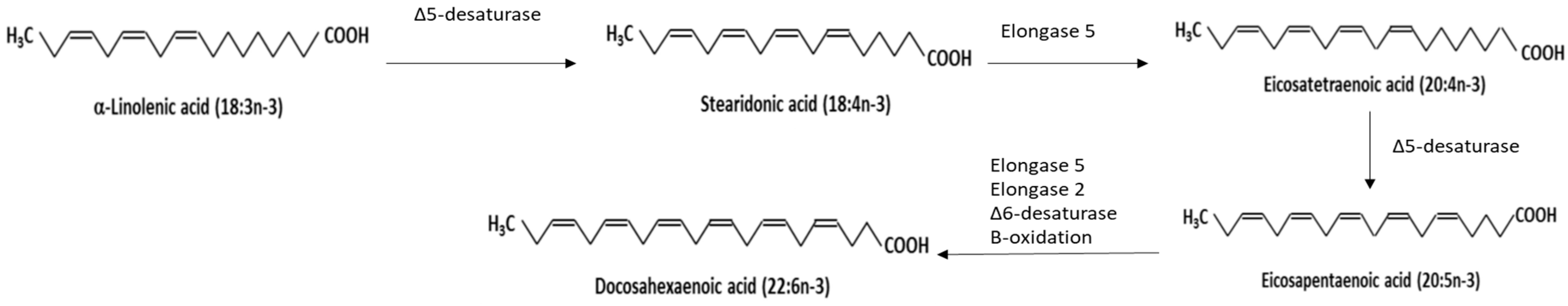
4. N3PUFA Form and Bioavailability
5. A High N3PUFA Dose Can Be Essential in Protecting Cardiovascular Health from COVID-19
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. WHO Coronavirus (COVID-19) Dashboard Overview of Current Cases; WHO: Geneva, Switzerland, 2022. [Google Scholar]
- Huertas, A.; Montani, D.; Savale, L.; Pichon, J.; Tu, L.; Parent, F.; Guignabert, C.; Humbert, M. Endothelial Cell Dysfunction: A Major Player in SARS-CoV-2 Infection (COVID-19)? Eur. Respir. J. 2020, 56, 2001634. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Ilyas, I.; Weng, J. Endothelial Dysfunction in COVID-19: An Overview of Evidence, Biomarkers, Mechanisms and Potential Therapies. Acta Pharmacol. Sin. 2022, 44, 695–709. [Google Scholar] [CrossRef] [PubMed]
- Yau, J.W.; Hwee, T.; Subodh, V. Endothelial Cell Control of Thrombosis. BMC Cardiovasc. Disord. 2015, 15, 130. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A.; et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in COVID-19. N. Engl. J. Med. 2020, 383, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Hamming, I.; Timens, W.; Bulthuis, M.L.C.; Lely, A.T.; Navid, G.J.; van Goor, H. Tissue Distribution of ACE2 Protein, the Functional Receptor for SARS Coronavirus. A First Step in Understanding SARS Pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Muniyappa, R.; Gubbi, S. COVID-19 Pandemic, Coronaviruses, and Diabetes Mellitus. Am. J. Physiol.-Endocrinol. Metab. 2020, 318, E736–E741. [Google Scholar] [CrossRef]
- Singh, A.K.; Gillies, C.L.; Singh, R.; Singh, A.; Chudasama, Y.; Coles, B.; Seidu, S.; Zaccardi, F.; Davies, M.J.; Khunti, K.; et al. Prevalence of Co-morbidities and Their Association with Mortality in Patients with COVID-19: A Systematic Review and Meta-analysis. Diabetes Obes. Metab. 2020, 22, 1915–1924. [Google Scholar] [CrossRef]
- Clerkin, K.J.; Fried, J.A.; Jayant, R.; Gabriel, S.; Jan, M.G.; Amirali, M.; Sneha, S.J.; Daniel, B.; Deepa, K.; LeRoy, R.; et al. COVID-19 and Cardiovascular Disease. Circulation 2020, 141, 1648–1655. [Google Scholar] [CrossRef]
- Bansal, M. Cardiovascular Disease and COVID-19. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 247–250. [Google Scholar] [CrossRef]
- Shah, W.; Hillman, T.; Playford, E.D.; Hishmeh, L. Managing the Long Term Effects of COVID-19: Summary of NICE, SIGN, and RCGP Rapid Guideline. BMJ 2021, 372, n136. [Google Scholar] [CrossRef]
- Rezel-Potts, E.; Douiri, A.; Sun, X.; Chowienczyk, P.J.; Shah, A.M.; Gulliford, M.C. Cardiometabolic Outcomes up to 12 Months after COVID-19 Infection. A Matched Cohort Study in the UK. PLoS Med. 2022, 19, e1004052. [Google Scholar] [CrossRef]
- Jung, E.M.; Stroszczynski, C.; Jung, F. Contrast Enhanced Ultrasonography (CEUS) to Detect Abdominal Microcirculatory Disorders in Severe Cases of COVID-19 Infection: First Experience. Clin. Hemorheol. Microcirc. 2020, 74, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Koutsiaris, A.G.; Riri, K.; Boutlas, S.; Panagiotou, T.N.; Kotoula, M.; Daniil, Z.; Tsironi, E.E. COVID-19 Hemodynamic and Thrombotic Effect on the Eye Microcirculation after Hospitalization: A Quantitative Case-Control Study. Clin. Hemorheol. Microcirc. 2022, 82, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Knight, R.; Walker, V.; Ip, S.; Cooper, J.A.; Bolton, T.; Keene, S.; Denholm, R.; Akbari, A.; Abbasizanjani, H.; Torabi, F.; et al. Association of COVID-19 with Arterial and Venous Vascular Diseases: A Population-Wide Cohort Study of 48 Million Adults in England and Wales. medRxiv 2021. [Google Scholar] [CrossRef]
- Calder, P.C. Nutrition, Immunity and COVID-19. BMJ Nutr. Prev. Health 2020, 3, 74–92. [Google Scholar] [CrossRef]
- Mentella, M.C.; Scaldaferri, F.; Gasbarrini, A.; Miggiano, G.A.D. The Role of Nutrition in the COVID-19 Pandemic. Nutrients 2021, 13, 1093. [Google Scholar] [CrossRef]
- Ramírez-Santana, M.; Zapata Barra, R.; Ñunque González, M.; Müller, J.M.; Vásquez, J.E.; Ravera, F.; Lago, G.; Cañón, E.; Castañeda, D.; Pradenas, M. Inverse Association between Omega-3 Index and Severity of COVID-19: A Case–Control Study. Int. J. Environ. Res. Public Health 2022, 19, 6445. [Google Scholar] [CrossRef]
- Hathaway, D.; Pandav, K.; Patel, M.; Riva-Moscoso, A.; Singh, B.M.; Patel, A.; Min, Z.C.; Singh-Makkar, S.; Sana, M.K.; Sanchez-Dopazo, R.; et al. Omega 3 Fatty Acids and COVID-19: A Comprehensive Review. Infect Chemother. 2020, 52, 478–495. [Google Scholar] [CrossRef]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic Characterisation and Epidemiology of 2019 Novel Coronavirus: Implications for Virus Origins and Receptor Binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef]
- Wu, Z.; McGoogan, J.M. Characteristics of and Important Lessons From the Coronavirus Disease 2019 (COVID-19) Outbreak in China: Summary of a Report of 72 314 Cases From the Chinese Center for Disease Control and Prevention. JAMA 2020, 323, 1239–1242. [Google Scholar] [CrossRef]
- Menter, T.; Haslbauer, J.D.; Nienhold, R.; Savic, S.; Hopfer, H.; Deigendesch, N.; Frank, S.; Turek, D.; Willi, N.; Pargger, H.; et al. Postmortem Examination of COVID-19 Patients Reveals Diffuse Alveolar Damage with Severe Capillary Congestion and Variegated Findings in Lungs and Other Organs Suggesting Vascular Dysfunction. Histopathology 2020, 77, 198–209. [Google Scholar] [CrossRef]
- Wichmann, D.; Sperhake, J.-P.; Lütgehetmann, M.; Steurer, S.; Edler, C.; Heinemann, A.; Heinrich, F.; Mushumba, H.; Kniep, I.; Schröder, A.S.; et al. Autopsy Findings and Venous Thromboembolism in Patients With COVID-19: A Prospective Cohort Study. Ann. Intern. Med. 2020, 173, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, S.; Fischer, I.; Naderi, S.; Faghih Jouibari, M.; Abdolreza, S.; Karimialavijeh, E.; Aslzadeh, S.; Mashayekhi, M.; Zojaji, M.; Kahlert, U.D.; et al. Systemic Inflammatory Index Is a Novel Predictor of Intubation Requirement and Mortality after SARS-CoV-2 Infection. Pathogens 2021, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- Ball, L.; Silva, P.L.; Giacobbe, D.R.; Bassetti, M.; Zubieta-Calleja, G.R.; Rocco, P.R.M.; Pelosi, P. Understanding the Pathophysiology of Typical Acute Respiratory Distress Syndrome and Severe COVID-19. Expert Rev. Respir. Med. 2022, 16, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Argenziano, M.G.; Bruce, S.L.; Slater, C.L.; Tiao, J.R.; Baldwin, M.R.; Barr, R.G.; Chang, B.P.; Chau, K.H.; Choi, J.J.; Gavin, N.; et al. Characterization and Clinical Course of 1000 Patients with Coronavirus Disease 2019 in New York: Retrospective Case Series. BMJ 2020, 369, m1996. [Google Scholar] [CrossRef] [PubMed]
- Lala, A.; Johnson, K.W.; Januzzi, J.L.; Russak, A.J.; Paranjpe, I.; Richter, F.; Zhao, S.; Somani, S.; Van Vleck, T.; Vaid, A.; et al. Prevalence and Impact of Myocardial Injury in Patients Hospitalized With COVID-19 Infection. J. Am.Coll. Cardiol. 2020, 76, 533–546. [Google Scholar] [CrossRef]
- Klok, F.A.; Kruip, M.J.H.A.; van der Meer, N.J.M.; Arbous, M.S.; Gommers, D.A.M.P.J.; Kant, K.M.; Kaptein, F.H.J.; van Paassen, J.; Stals, M.A.M.; Huisman, M.V.; et al. Incidence of Thrombotic Complications in Critically Ill ICU Patients with COVID-19. Thromb. Res. 2020, 191, 145–147. [Google Scholar] [CrossRef]
- Poissy, J.; Goutay, J.; Caplan, M.; Parmentier, E.; Duburcq, T.; Lassalle, F.; Jeanpierre, E.; Rauch, A.; Labreuche, J.; Susen, S.; et al. Pulmonary Embolism in Patients With COVID-19: Awareness of an Increased Prevalence. Circulation 2020, 142, 184–186. [Google Scholar] [CrossRef]
- Jenner, W.J.; Gorog, D.A. Incidence of Thrombotic Complications in COVID-19. J. Thromb. Thrombolysis 2021, 52, 999–1006. [Google Scholar] [CrossRef]
- Lippi, G.; Favaloro, E.J. D-Dimer Is Associated with Severity of Coronavirus Disease 2019: A Pooled Analysis. Thromb. Haemost. 2020, 120, 876–878. [Google Scholar] [CrossRef]
- Xiong, X.; Chi, J.; Gao, Q. Prevalence and Risk Factors of Thrombotic Events on Patients with COVID-19: A Systematic Review and Meta-analysis. Thromb. J. 2021, 19, 32. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Huang, X.; Ding, D.; Zhang, J.; Xu, L.; Hu, Z.; Xu, W.; Tao, Z. Comparative Study of Acute Lung Injury in COVID-19 and Non-COVID-19 Patients. Front. Med. 2021, 8, 666629. [Google Scholar] [CrossRef] [PubMed]
- Stals, M.A.M.; Grootenboers, M.J.J.H.; van Guldener, C.; Kaptein, F.H.J.; Braken, S.J.E.; Chen, Q.; Chu, G.; van Driel, E.M.; Iglesias Del Sol, A.; de Jonge, E.; et al. Risk of Thrombotic Complications in Influenza versus COVID-19 Hospitalized Patients. Res. Pract. Thromb. Haemost. 2021, 5, 412–420. [Google Scholar] [CrossRef]
- Williamson, E.J.; Walker, A.J.; Bhaskaran, K.; Bacon, S.; Bates, C.; Morton, C.E.; Curtis, H.J.; Mehrkar, A.; Evans, D.; Inglesby, P. Factors Associated with COVID-19-Related Death Using OpenSAFELY. Nature 2020, 584, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Holman, N.; Knighton, P.; Kar, P.; O’Keefe, J.; Curley, M.; Weaver, A.; Barron, E.; Bakhai, C.; Khunti, K.; Wareham, N.J.; et al. Risk Factors for COVID-19-Related Mortality in People with Type 1 and Type 2 Diabetes in England: A Population-Based Cohort Study. Lancet Diabetes Endocrinol. 2020, 8, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.X.; Tyagi, T.; Jain, K.; Gu, V.W.; Lee, S.H.; Hwa, J.M.; Kwan, J.M.; Krause, D.S.; Lee, A.I.; Halene, S.; et al. Thrombocytopathy and Endotheliopathy: Crucial Contributors to COVID-19 Thromboinflammation. Nat. Rev. Cardiol. 2021, 18, 194–209. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef] [PubMed]
- Katsoularis, I.; Fonseca-Rodríguez, O.; Farrington, P.; Jerndal, H.; Lundevaller, E.H.; Sund, M.; Lindmark, K.; Connolly, A.-M.F. Risks of Deep Vein Thrombosis, Pulmonary Embolism, and Bleeding after COVID-19: Nationwide Self-Controlled Cases Series and Matched Cohort Study. BMJ 2022, 377, e069590. [Google Scholar] [CrossRef] [PubMed]
- Giacca, M.; Shah, A.M. The Pathological Maelstrom of COVID-19 and Cardiovascular Disease. Nat. Cardiovasc. Res. 2022, 1, 200–210. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, M.; Chen, X.; Montaner, L.J. Cytokine Storm and Leukocyte Changes in Mild versus Severe SARS-CoV-2 Infection: Review of 3939 COVID-19 Patients in China and Emerging Pathogenesis and Therapy Concepts. J. Leukoc. Biol. 2020, 108, 17–41. [Google Scholar] [CrossRef]
- Du, F.; Liu, B.; Zhang, S. COVID-19: The Role of Excessive Cytokine Release and Potential ACE2 down-Regulation in Promoting Hypercoagulable State Associated with Severe Illness. J. Thromb. Thrombolysis 2021, 51, 313–329. [Google Scholar] [CrossRef] [PubMed]
- Tay, M.Z.; Poh, C.M.; Rénia, L.; MacAry, P.A.; Ng, L.F.P. The Trinity of COVID-19: Immunity, Inflammation and Intervention. Nat. Rev. Immunol. 2020, 20, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Loo, J.; Spittle, D.A.; Newnham, M. COVID-19, Immunothrombosis and Venous Thromboembolism: Biological Mechanisms. Thorax 2021, 76, 412–420. [Google Scholar] [CrossRef] [PubMed]
- López, D.; Orta, X.; Casós, K.; Sáiz, M.P.; Puig-Parellada, P.; Farriol, M.; Mitjavila, M.T. Upregulation of Endothelial Nitric Oxide Synthase in Rat Aorta after Ingestion of Fish Oil-Rich Diet. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H567–H572. [Google Scholar] [CrossRef] [PubMed]
- Petrosini, L.; Cutuli, D.; Caporali, P.; Ronci, M. N–3 Polyunsaturated Fatty Acids Supplementation Decreases Asymmetric Dimethyl Arginine and Arachidonate Accumulation in Aging Spontaneously Hypertensive Rats. Eur. J. Nutr. 2005, 44, 327–333. [Google Scholar]
- Chisaki, K.; Okuda, Y.; Suzuki, S.; Miyauchi, T.; Soma, M.; Ohkoshi, N.; Sone, H.; Yamada, N.; Nakajima, T. Eicosapentaenoic Acid Suppresses Basal and Insulin-Stimulated Endothelin-1 Production in Human Endothelial Cells. Hypertens. Res. 2003, 26, 655–661. [Google Scholar] [CrossRef]
- Christensen, P.; Larsen, T.M.; Westerterp-Plantenga, M.; Macdonald, I.; Martinez, J.A.; Handjiev, S.; Poppitt, S.D.; Ritz, C.; Taylor, M.A.; Raben, A.; et al. Men and Women Respond Differently to Rapid Weight Loss: Metabolic Outcomes after a Low-Energy Diet in 2,500 Overweight, Pre-Diabetic Individuals in the PREVIEW Intervention Study. Diab. Obesity Metab. 2018, 20, 2840–2851. [Google Scholar] [CrossRef]
- Wu, S.-Y.; Mayneris-Perxachs, J.; Lovegrove, J.A.; Todd, S.; Yaqoob, P. Fish-Oil Supplementation Alters Numbers of Circulating Endothelial Progenitor Cells and Microparticles Independently of ENOS Genotype1,2,3,4. Am. J. Clin. Nutr. 2014, 100, 1232–1243. [Google Scholar] [CrossRef]
- Wu, J.H.Y.; Cahill, L.E.; Mozaffarian, D. Effect of Fish Oil on Circulating Adiponectin: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. J. Clin. Endocrinol. Metab. 2013, 98, 2451–2459. [Google Scholar] [CrossRef]
- Din, J.N.; Archer, R.M.; Harding, S.A.; Sarma, J.; Lyall, K.; Flapan, A.D.; Newby, D.E. Effect of ω-3 Fatty Acid Supplementation on Endothelial Function, Endogenous Fibrinolysis and Platelet Activation in Male Cigarette Smokers. Heart 2013, 99, 168–174. [Google Scholar] [CrossRef]
- Monahan, K.D.; Feehan, R.P.; Blaha, C.; McLaughlin, D.J. Effect of Omega-3 Polyunsaturated Fatty Acid Supplementation on Central Arterial Stiffness and Arterial Wave Reflections in Young and Older Healthy Adults. Physiol. Rep. 2015, 3, e12438. [Google Scholar] [CrossRef] [PubMed]
- Singhal, A.; Lanigan, J.; Storry, C.; Low, S.; Birbara, T.; Lucas, A.; Deanfield, J. Docosahexaenoic Acid Supplementation, Vascular Function and Risk Factors for Cardiovascular Disease: A Randomized Controlled Trial in Young Adults. J. Am. Heart Assoc. 2013, 2, e000283. [Google Scholar] [CrossRef] [PubMed]
- Grenon, S.M.; Owens, C.D.; Nosova, E.V.; Hughes-Fulford, M.; Alley, H.F.; Chong, K.; Perez, S.; Yen, P.K.; Boscardin, J.; Hellmann, J.; et al. Short-Term, High-Dose Fish Oil Supplementation Increases the Production of Omega-3 Fatty Acid–Derived Mediators in Patients With Peripheral Artery Disease (the OMEGA-PAD I Trial). J. Am. Heart Assoc. 2015, 4, e002034. [Google Scholar] [CrossRef] [PubMed]
- Isaksen, T.; Evensen, L.H.; Johnsen, S.H.; Jacobsen, B.K.; Hindberg, K.; Brækkan, S.K.; Hansen, J. Dietary Intake of Marine N-3 Polyunsaturated Fatty Acids and Future Risk of Venous Thromboembolism. Res. Pract. Thromb. Haemost. 2018, 3, 59–69. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, J.; Guo, H.; Liang, J.; Li, Y. Associations of Fish and Omega-3 Fatty Acids Consumption With the Risk of Venous Thromboembolism. A Meta-Analysis of Prospective Cohort Studies. Front. Nutr. 2020, 7, 614784. [Google Scholar] [CrossRef]
- Bonutti, P.M.; Sodhi, N.; Patel, Y.H.; Sultan, A.A.; Khlopas, A.; Chughtai, M.; Kolisek, F.R.; Williams, N.; Mont, M.A. Novel Venous Thromboembolic Disease (VTED) Prophylaxis for Total Knee Arthroplasty—Aspirin and Fish Oil. Ann. Transl. Med. 2017, 5, S30. [Google Scholar] [CrossRef]
- Zheng, X.; Jia, R.; Li, Y.; Liu, T.; Wang, Z. Omega-3 Fatty Acids Reduce Post-Operative Risk of Deep Vein Thrombosis and Pulmonary Embolism after Surgery for Elderly Patients with Proximal Femoral Fractures: A Randomized Placebo-Controlled, Double-Blind Clinical Trial. Int. Orthop. 2020, 44, 2089–2093. [Google Scholar] [CrossRef]
- Lordan, S.; Smyth, T.J.; Soler-Vila, A.; Stanton, C.; Ross, R.P. The α-Amylase and α-Glucosidase Inhibitory Effects of Irish Seaweed Extracts. Food Chem. 2013, 141, 2170–2176. [Google Scholar] [CrossRef]
- Chap, H. Forty Five Years with Membrane Phospholipids, Phospholipases and Lipid Mediators: A Historical Perspective. Biochimie 2016, 125, 234–249. [Google Scholar] [CrossRef]
- Fadeel, B.; Xue, D. The Ins and Outs of Phospholipid Asymmetry in the Plasma Membrane: Roles in Health and Disease. Crit. Rev. Biochem. Mol. Biol. 2009, 44, 264–277. [Google Scholar] [CrossRef]
- Yeung, J.; Apopa, P.L.; Vesci, J.; Stolla, M.; Rai, G.; Simeonov, A.; Jadhav, A.; Fernandez-Perez, P.; Maloney, D.J.; Boutaud, O.; et al. 12-Lipoxygenase Activity Plays an Important Role in PAR4 and GPVI-Mediated Platelet Reactivity. Thromb. Haemost. 2013, 110, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Ikei, K.N.; Yeung, J.; Apopa, P.L.; Ceja, J.; Vesci, J.; Holman, T.R.; Holinstat, M. Investigations of Human Platelet-Type 12-Lipoxygenase: Role of Lipoxygenase Products in Platelet Activation1. J. Lipid Res. 2012, 53, 2546–2559. [Google Scholar] [CrossRef] [PubMed]
- Adili, R.; Hawley, M.; Holinstat, M. Regulation of Platelet Function and Thrombosis by Omega-3 and Omega-6 Polyunsaturated Fatty Acids. Prostaglandins Lipid Mediat. 2018, 139, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Schoene, N.; Harris, W. Mean Platelet Volume as an Indicator of Platelet Activation: Methodological Issues. Platelets 2002, 13, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial Reactive Oxygen Species (ROS) and ROS-Induced ROS Release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef] [PubMed]
- Laforge, M.; Elbim, C.; Frère, C.; Hémadi, M.; Massaad, C.; Nuss, P.; Benoliel, J.-J.; Becker, C. Tissue Damage from Neutrophil-Induced Oxidative Stress in COVID-19. Nat. Rev. Immunol. 2020, 20, 515–516. [Google Scholar] [CrossRef]
- Kashani, K.B. Hypoxia in COVID-19: Sign of Severity or Cause for Poor Outcomes. Mayo Clin. Proc. 2020, 95, 1094–1096. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Covassin, N.; Fan, Z.; Singh, P.; Gao, W.; Li, G.; Kara, T.; Somers, V.K. Association Between Hypoxemia and Mortality in Patients With COVID-19. Mayo Clin. Proc. 2020, 95, 1138–1147. [Google Scholar] [CrossRef]
- Ribeiro, D.; Sousa, A.; Nicola, P.; Ferreira de Oliveira, J.M.P.; Rufino, A.T.; Silva, M.; Freitas, M.; Carvalho, F.; Fernandes, E. β-Carotene and Its Physiological Metabolites: Effects on Oxidative Status Regulation and Genotoxicity in in Vitro Models. Food Chem. Toxicol. 2020, 141, 111392. [Google Scholar] [CrossRef]
- Gutiérrez, S.; Svahn, S.L.; Johansson, M.E. Effects of Omega-3 Fatty Acids on Immune Cells. Int. J. Mol. Sci. 2019, 20, 5028. [Google Scholar] [CrossRef]
- Williams-Bey, Y.; Boularan, C.; Vural, A.; Huang, N.-N.; Hwang, I.-Y.; Shan-Shi, C.; Kehrl, J.H. Omega-3 Free Fatty Acids Suppress Macrophage Inflammasome Activation by Inhibiting NF-ΚB Activation and Enhancing Autophagy. PLoS ONE 2014, 9, e97957. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.; Jeon, H.; Kim, I.-H.; Jeong, H.S.; Lee, J. Anti-Inflammatory Effects of Stearidonic Acid Mediated by Suppression of NF-ΚB and MAP-Kinase Pathways in Macrophages. Lipids 2017, 52, 781–787. [Google Scholar] [CrossRef] [PubMed]
- Su, K.-P.; Lai, H.-C.; Yang, H.-T.; Su, W.-P.; Peng, C.-Y.; Chang, J.P.-C.; Chang, H.-C.; Pariante, C.M. Omega-3 Fatty Acids in the Prevention of Interferon-Alpha-Induced Depression: Results from a Randomized, Controlled Trial. Biol. Psychiatry 2014, 76, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Cucchi, D.; Camacho-Muñoz, D.; Certo, M.; Niven, J.; Smith, J.; Nicolaou, A.; Mauro, C. Omega-3 Polyunsaturated Fatty Acids Impinge on CD4+ T Cell Motility and Adipose Tissue Distribution via Direct and Lipid Mediator-Dependent Effects. Cardiovasc. Res. 2020, 116, 1006–1020. [Google Scholar] [CrossRef]
- Kang, K.W.; Kim, S.; Cho, Y.-B.; Ryu, S.R.; Seo, Y.-J.; Lee, S.-M. Endogenous N-3 Polyunsaturated Fatty Acids Are Beneficial to Dampen CD8+ T Cell-Mediated Inflammatory Response upon the Viral Infection in Mice. Int. J. Mol. Sci. 2019, 20, 4510. [Google Scholar] [CrossRef]
- Jump, D.B. The Biochemistry of N-3 Polyunsaturated Fatty Acids. J. Biol. Chem. 2002, 277, 8755–8758. [Google Scholar] [CrossRef]
- Serhan, C.N. Pro-Resolving Lipid Mediators Are Leads for Resolution Physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef]
- Meydani, S.N.; Endres, S.; Woods, M.M.; Goldin, B.R.; Soo, C.; Morrill-Labrode, A.; Dinarello, C.A.; Gorbach, S.L. Oral (n-3) Fatty Acid Supplementation Suppresses Cytokine Production and Lymphocyte Proliferation: Comparison between Young and Older Women. J. Nutr. 1991, 121, 547–555. [Google Scholar] [CrossRef]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 Is an Omega-3 Fatty Acid Receptor Mediating Potent Anti-Inflammatory and Insulin-Sensitizing Effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Wu, J.H.Y. Omega-3 Fatty Acids and Cardiovascular Disease: Effects on Risk Factors, Molecular Pathways, and Clinical Events. J. Am. Coll. Cardiol. 2011, 58, 2047–2067. [Google Scholar] [CrossRef]
- Kaur, G.; Cameron-Smith, D.; Garg, M.; Sinclair, A.J. Docosapentaenoic Acid (22:5n-3): A Review of Its Biological Effects. Prog. Lipid Res. 2011, 50, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.J.; Miles, E.A.; Burdge, G.C.; Yaqoob, P.; Calder, P.C. Metabolism and Functional Effects of Plant-Derived Omega-3 Fatty Acids in Humans. Prog. Lipid Res. 2016, 64, 30–56. [Google Scholar] [CrossRef] [PubMed]
- Innes, J.K.; Calder, P.C. Marine Omega-3 (N-3) Fatty Acids for Cardiovascular Health: An Update for 2020. Int. J. Mol. Sci. 2020, 21, 1362. [Google Scholar] [CrossRef]
- Calder, P.C. Very Long-Chain n-3 Fatty Acids and Human Health: Fact, Fiction and the Future. Proc. Nutr. Soc. 2018, 77, 52–72. [Google Scholar] [CrossRef]
- Burri, L.; Hoem, N.; Banni, S.; Berge, K. Marine Omega-3 Phospholipids: Metabolism and Biological Activities. Int. J. Mol. Sci. 2012, 13, 15401–15419. [Google Scholar] [CrossRef]
- Richter, C.K.; Bowen, K.J.; Mozaffarian, D.; Kris-Etherton, P.M.; Skulas-Ray, A.C. Total Long-Chain n-3 Fatty Acid Intake and Food Sources in the United States Compared to Recommended Intakes: NHANES 2003–2008. Lipids 2017, 52, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Ackman, R.G. The Absorption of Fish Oils and Concentrates. Lipids 1992, 27, 858–862. [Google Scholar] [CrossRef]
- Dyerberg, J.; Madsen, P.; Møller, J.M.; Aardestrup, I.; Schmidt, E.B. Bioavailability of Marine N-3 Fatty Acid Formulations. Prostaglandins Leukot. Essent. Fat. Acids 2010, 83, 137–141. [Google Scholar] [CrossRef]
- West, A.L.; Burdge, G.C.; Calder, P.C. Lipid Structure Does Not Modify Incorporation of EPA and DHA into Blood Lipids in Healthy Adults: A Randomised-Controlled Trial. Br. J. Nutr. 2016, 116, 788–797. [Google Scholar] [CrossRef]
- Sadovsky, R.; Kris-Etherton, P. Prescription Omega-3-Acid Ethyl Esters for the Treatment of Very High Triglycerides. Postgrad. Med. 2009, 121, 145–153. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the Substantiation of Health Claims Related to Eicosapentaenoic Acid (EPA), Docosahexaenoic Acid (DHA), Docosapentaenoic Acid (DPA) and Maintenance of Normal Cardiac Function (ID 504, 506, 516, 527, 538, 703, 1128, 1317, 1324, 1325), Maintenance of Normal Blood Glucose Concentrations (ID 566), Maintenance of Normal Blood Pressure (ID 506, 516, 703, 1317, 1324), Maintenance of Normal Blood HDL-Cholesterol Concentrations (ID 506), Maintenance of Normal (Fasting) Blood Concentrations of Triglycerides (ID 506, 527, 538, 1317, 1324, 1325), Maintenance of Normal Blood LDL-Cholesterol Concentrations (ID 527, 538, 1317, 1325, 4689), Protection of the Skin from Photo-Oxidative (UV-Induced) Damage (ID 530), Improved Absorption of EPA and DHA (ID 522, 523), Contribution to the Normal Function of the Immune System by Decreasing the Levels of Eicosanoids, Arachidonic Acid-Derived Mediators and pro-Inflammatory Cytokines (ID 520, 2914), and “Immunomodulating Agent” (4690) Pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2010, 8, 1796. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Fats and Fatty Acids in Human Nutrition. Report of an Expert Consultation, 10–14 November 2008, Geneva. Available online: https://agris.fao.org/agris-search/search.do?recordID=XF2016049106 (accessed on 12 March 2023).
- Public Health England. SACN Advice on Fish Consumption. Available online: https://www.gov.uk/government/publications/sacn-advice-on-fish-consumption (accessed on 12 March 2023).
- Meyer, B.J.; Mann, N.J.; Lewis, J.L.; Milligan, G.C.; Sinclair, A.J.; Howe, P.R.C. Dietary Intakes and Food Sources of Omega-6 and Omega-3 Polyunsaturated Fatty Acids. Lipids 2003, 38, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Howe, P.; Meyer, B.; Record, S.; Baghurst, K. Dietary Intake of Long-Chain ω-3 Polyunsaturated Fatty Acids: Contribution of Meat Sources. Nutrition 2006, 22, 47–53. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, J.H.; Jacob, D.; Lavie, C.J. Omega-3 Fatty Acid Therapy: The Tide Turns for a Fish Story. Mayo Clin. Proc. 2017, 92, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Marchioli, R. Dietary Supplementation with N-3 Polyunsaturated Fatty Acids and Vitamin E after Myocardial Infarction: Results of the GISSI-Prevenzione Trial. Lancet 1999, 354, 447–455. [Google Scholar] [CrossRef]
- Yokoyama, M.; Origasa, H.; Matsuzaki, M.; Matsuzawa, Y.; Saito, Y.; Ishikawa, Y.; Oikawa, S.; Sasaki, J.; Hishida, H.; Itakura, H.; et al. Effects of Eicosapentaenoic Acid on Major Coronary Events in Hypercholesterolaemic Patients (JELIS): A Randomised Open-Label, Blinded Endpoint Analysis. Lancet 2007, 369, 1090–1098. [Google Scholar] [CrossRef]
- Gissi-HF Investigators. Effect of N-3 Polyunsaturated Fatty Acids in Patients with Chronic Heart Failure (the GISSI-HF Trial): A Randomised, Double-Blind, Placebo-Controlled Trial. Lancet 2008, 372, 1223–1230. [CrossRef]
- Einvik, G.; Klemsdal, T.O.; Sandvik, L.; Hjerkinn, E.M. A Randomized Clinical Trial on N-3 Polyunsaturated Fatty Acids Supplementation and All-Cause Mortality in Elderly Men at High Cardiovascular Risk. Eur. J. Cardiovasc. Prev. Rehabil. 2010, 17, 588–592. [Google Scholar] [CrossRef]
- Galan, P.; Kesse-Guyot, E.; Czernichow, S.; Briancon, S.; Blacher, J.; Hercberg, S. Effects of B Vitamins and Omega 3 Fatty Acids on Cardiovascular Diseases: A Randomised Placebo Controlled Trial. BMJ 2010, 341, c6273. [Google Scholar] [CrossRef]
- Kromhout, D.; Giltay, E.J.; Geleijnse, J.M. Alpha Omega Trial Group N-3 Fatty Acids and Cardiovascular Events after Myocardial Infarction. N. Engl. J. Med. 2010, 363, 2015–2026. [Google Scholar] [CrossRef]
- Rauch, B.; Schiele, R.; Schneider, S.; Diller, F.; Victor, N.; Gohlke, H.; Gottwik, M.; Steinbeck, G.; Del Castillo, U.; Sack, R.; et al. OMEGA, a Randomized, Placebo-Controlled Trial to Test the Effect of Highly Purified Omega-3 Fatty Acids on Top of Modern Guideline-Adjusted Therapy after Myocardial Infarction. Circulation 2010, 122, 2152–2159. [Google Scholar] [CrossRef] [PubMed]
- The ORIGIN Trial Investigators. N–3 Fatty Acids and Cardiovascular Outcomes in Patients with Dysglycemia. N. Engl. J. Med. 2012, 367, 309–318. [Google Scholar] [CrossRef] [PubMed]
- The Risk and Prevention Study Collaborative Group. N–3 Fatty Acids in Patients with Multiple Cardiovascular Risk Factors. N. Engl. J. Med. 2013, 368, 1800–1808. [Google Scholar] [CrossRef] [PubMed]
- Writing Group for the AREDS2 Research Group. Effect of Long-Chain ω-3 Fatty Acids and Lutein + Zeaxanthin Supplements on Cardiovascular Outcomes: Results of the Age-Related Eye Disease Study 2 (AREDS2) Randomized Clinical Trial. JAMA Intern. Med. 2014, 174, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Heydari, B.; Abdullah, S.; Pottala, J.V.; Shah, R.; Abbasi, S.; Mandry, D.; Francis, S.A.; Lumish, H.; Ghoshhajra, B.B.; Hoffman, U.; et al. Effect of Omega-3 Acid Ethyl Esters on Left Ventricular Remodeling After Acute Myocardial Infarction: The OMEGA-REMODEL Randomized Clinical Trial. Circulation 2016, 134, 378. [Google Scholar] [CrossRef]
- The ASCEND Study Collaborative Group. Effects of N−3 Fatty Acid Supplements in Diabetes Mellitus. N. Engl. J. Med. 2018, 379, 1540–1550. [Google Scholar] [CrossRef] [PubMed]
- Manson, J.E.; Cook, N.R.; Lee, I.-M.; Christen, W.; Bassuk, S.S.; Mora, S.; Gibson, H.; Albert, C.M.; Gordon, D.; Copeland, T.; et al. Marine N−3 Fatty Acids and Prevention of Cardiovascular Disease and Cancer. N. Engl. J. Med. 2019, 380, 23–32. [Google Scholar] [CrossRef]
- Bhatt, D.L.; Steg, P.G.; Miller, M.; Brinton, E.A.; Jacobson, T.A.; Ketchum, S.B.; Doyle, R.T.; Juliano, R.A.; Jiao, L.; Granowitz, C.; et al. Cardiovascular Risk Reduction with Icosapent Ethyl for Hypertriglyceridemia. N. Engl. J. Med. 2019, 380, 11–22. [Google Scholar] [CrossRef]
- Hu, Y.; Hu, F.B.; Manson, J.E. Marine Omega-3 Supplementation and Cardiovascular Disease: An Updated Meta-Analysis of 13 Randomized Controlled Trials Involving 127 477 Participants. J. Am. Heart Assoc. 2019, 8, e013543. [Google Scholar] [CrossRef]
- Bernasconi, A.A.; Wiest, M.M.; Lavie, C.J.; Milani, R.V.; Laukkanen, J.A. Effect of Omega-3 Dosage on Cardiovascular Outcomes: An Updated Meta-Analysis and Meta-Regression of Interventional Trials. Mayo Clin. Proc. 2021, 96, 304–313. [Google Scholar] [CrossRef]
- Maki, K.C.; Palacios, O.M.; Bell, M.; Toth, P.P. Use of Supplemental Long-Chain Omega-3 Fatty Acids and Risk for Cardiac Death: An Updated Meta-Analysis and Review of Research Gaps. J. Clin. Lipidol. 2017, 11, 1152–1160.e2. [Google Scholar] [CrossRef] [PubMed]
- Elagizi, A.; Lavie, C.J.; O’Keefe, E.; Marshall, K.; O’Keefe, J.H.; Milani, R.V. An Update on Omega-3 Polyunsaturated Fatty Acids and Cardiovascular Health. Nutrients 2021, 13, 204. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Pathological Significance | Effects of N3PUFA 1 |
|---|---|
| Endothelial function | ↑ NO 2 and eNOS 3 activity |
| ↓ endothelin-1 | |
| ↑ endothelium-mediated vasodilation | |
| ↑ VSMC 4 relaxation | |
| ↑ adiponectin | |
| Immunothrombosis | ↓ platelet activation |
| ↓ platelet aggregation and TX 5 release via COX-1 6 and 12-LOX 7 | |
| ↓ affinity of TxA2 8 | |
| ↑ PGI2 9 production | |
| ↓ fibrinogen level | |
| ↑ tPA 10 level | |
| ↓ proinflammatory cytokines (ILs 11, TNF 12) | |
| Inflammation | ↓ regulation of AP-1 13 and NFκB 14 |
| ↓ T-lymphocyte proliferation |
| Year | Trial | Population | No. of Subjects | Age (years) | Male (%) | Subject Characteristics | N3PUFA 1, Dose | Control | Study Period | Result |
|---|---|---|---|---|---|---|---|---|---|---|
| 1999 | GISSI-P [98] | Italian | 11,324 | 59 | 84.7 | Surviving recent (≤3 months) myocardial infarction | N3PUFA, 1 g/day | Vitamin E, 300 mg/day | 3.5 years | ↓ RR 2 of death = 10% (95% CI: 1–18%); ↓ RR of CVD = 17% (95% CI: 3–29%) |
| 2007 | JELIS [99] | Japanese | 18,645 | Average 61 | 31.4 | Total cholesterol ≥6.5 mmol/L | EPA 3, 1.8 g/day; statin | Statin only | 5 years | ↓ 19% RR in major cardiovascular events |
| 2008 | GISSI-HF [98,100] | Italian | 955 | ≥18 | 77.8 | With chronic heart failure of New York Heart Association classes II–IV, irrespective of cause, and left ventricular ejection fraction | N3PUFA, 1 g/day | Placebo | 3.9 years | ↓ HR 4 of death = 0.91 (95% CI: 0.833–0.998); ↓ HR of hospital admission for cardiovascular reasons = 0.92 (95% CI: 0.849–0.999) |
| 2010 | DOIT [101] | Norwegian | 563 | 64–76 | 100 | Without overt cardiovascular disease | N3PUFA, 2.4 g/day | Placebo (corn oil) | 3 years | ↓ HR of death = 0.57 (95% CI: 0.29–1.10); ↓ HR of cardiovascular events = 0.86 (95% CI: 0.57–1.38) |
| 2010 | SU.FOL.OM3 [102] | French | 2501 | 45–80 | 79.5 | With a history of myocardial infarction, unstable angina, or ischemic stroke | 5-methyltetrahydrofolate, 560 μg/day; vitamin B6, 3 mg/day; vitamin B12, 20 μg/day; N3PUFA, 0.6 g/day | Placebo | 4.7 years | No significant effect on major cardiovascular events |
| 2010 | Alpha-OMEGA [103] | Dutch | 4837 | 60–80 | 78.0 | Had a myocardial infarction, received state-of-the-art antihypertensive, antithrombotic, and lipid-modifying therapy | N3PUFA, 0.376 g/day (EPA, 0.226 g/day; DHA 5, 0.150 g/day) | ALA, 1.9 g/day | NA | → HR of major cardiovascular events = 1.01 (95% CI: 0.87–1.17) |
| 2010 | OMEGA [104] | German | 3851 | 64 | 74.4 | 3 to 14 days after acute myocardial infarction | N3PUFA (EE form), 1 g/day | Placebo | 1 year | No significant difference in sudden cardiac death, total mortality, major adverse cerebrovascular and cardiovascular events |
| 2012 | ORIGIN [105] | Canadian | 12,536 | ≥50 | 40.0 | At high risk for cardiovascular events and had impaired fasting glucose, impaired glucose tolerance, or diabetes | N3PUFA (EE form), 0.9 g/day | Placebo | 6.2 years | → HR of time to death or admission to the hospital for cardiovascular causes, 0.97 (95% CI: 0.88–1.08) |
| 2013 | R and P [106] | Italian | 12,513 | ≥65 | 61.5 | with multiple cardiovascular risk factors or atherosclerotic vascular disease but not myocardial infarction | N3PUFA (EE form), 1 g/day | Placebo | 1 year | → HR of the rates of major cardiovascular events, 1.01 (95% CI: 0.93–1.10) |
| 2014 | AREDS-2 [107] | American | 4203 | 50–85 | 56.8 | With stable, existing CVD (>12 months since initial event) | N3PUFA, 1 g/day (EPA, 650 mg/day; DHA, 350 mg/day); lutein, 10 mg/day; zeaxanthin, 2 mg/day | Placebo | 4.8 years | → HR of risk of CVD or secondary CVD outcomes, 0.95; 95% CI, 0.78–1.17 |
| 2016 | OMEGA- REMODEL [108] | American | 358 | >21 | 65.0 | With an acute MI | N3PUFA, 4 g/day (EPA, 465mg/day; DHA, 375 mg/day) | Corn oil (linoleic acid, no N3PUFA, 600 mg/day) | 6 months | ↓ LVESVI 6 (–5.8%, p = 0.017); ↓ Non-infarct myocardial fibrosis (−5.6%, p = 0.026) |
| 2018 | ASCEND [109] | British | 15,480 | ≥40 | 62.6 | With diabetes but without evidence of atherosclerotic cardiovascular disease | N3PUFA, 1 g/day | Olive oil, 1 g/day | 7.4 years | No significant difference in serious vascular event or revascularization |
| 2019 | VITAL [110] | American | 25,871 | >50 (males) >55 (females) | 49.9 | Healthy | N3PUFA, 1 g/day; vitamin D3 2000 IU/day | Placebo | 5.3 years | No significant difference in serious vascular event; ↓ HR of MI = 0.71 (95% CI:0.59–0.9) |
| 2019 | REDUCE-IT [111] | 71% (US, Canada, Netherlands, Australia, New Zealand, and South Africa), 25.8% (Eastern European), 3.2% (Asia-Pacific) | 8179 | ≥45 (established CVD) ≥50 (established T2DM) | 71.2 | With established cardiovascular disease or with diabetes and other risk factors, receiving statin therapy, fasting triglyceride level of 135 to 499 mg per deciliter (1.52 to 5.63 mmol per liter), a low-density lipoprotein cholesterol level of 41 to 100 mg per deciliter (1.06 to 2.59 mmol per liter) | EPA (icosapent ethyl highly purified EPA formulation), 4 g/day | Placebo | 4.9 years | ↓ HR of major cardiovascular events = 0.75 (95% CI: 0.68–0.83) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, L.W.; Quek, S.-Y.; Lu, S.-P.; Chen, J.-H. Potential Benefits of Omega-3 Polyunsaturated Fatty Acids (N3PUFAs) on Cardiovascular Health Associated with COVID-19: An Update for 2023. Metabolites 2023, 13, 630. https://doi.org/10.3390/metabo13050630
Lu LW, Quek S-Y, Lu S-P, Chen J-H. Potential Benefits of Omega-3 Polyunsaturated Fatty Acids (N3PUFAs) on Cardiovascular Health Associated with COVID-19: An Update for 2023. Metabolites. 2023; 13(5):630. https://doi.org/10.3390/metabo13050630
Chicago/Turabian StyleLu, Louise Weiwei, Siew-Young Quek, Shi-Ping Lu, and Jie-Hua Chen. 2023. "Potential Benefits of Omega-3 Polyunsaturated Fatty Acids (N3PUFAs) on Cardiovascular Health Associated with COVID-19: An Update for 2023" Metabolites 13, no. 5: 630. https://doi.org/10.3390/metabo13050630