
Study of the Metabolite Changes in Ganoderma lucidum under Pineapple Leaf Residue Stress via LC-MS/MS Coupled with a Non-Targeted Metabolomics Approach
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Fermentation Substrates
2.2. Extraction of Metabolites
2.3. Detection Conditions for LC-MS/MS
2.4. Database Search and Data Analysis
3. Results and Discussion
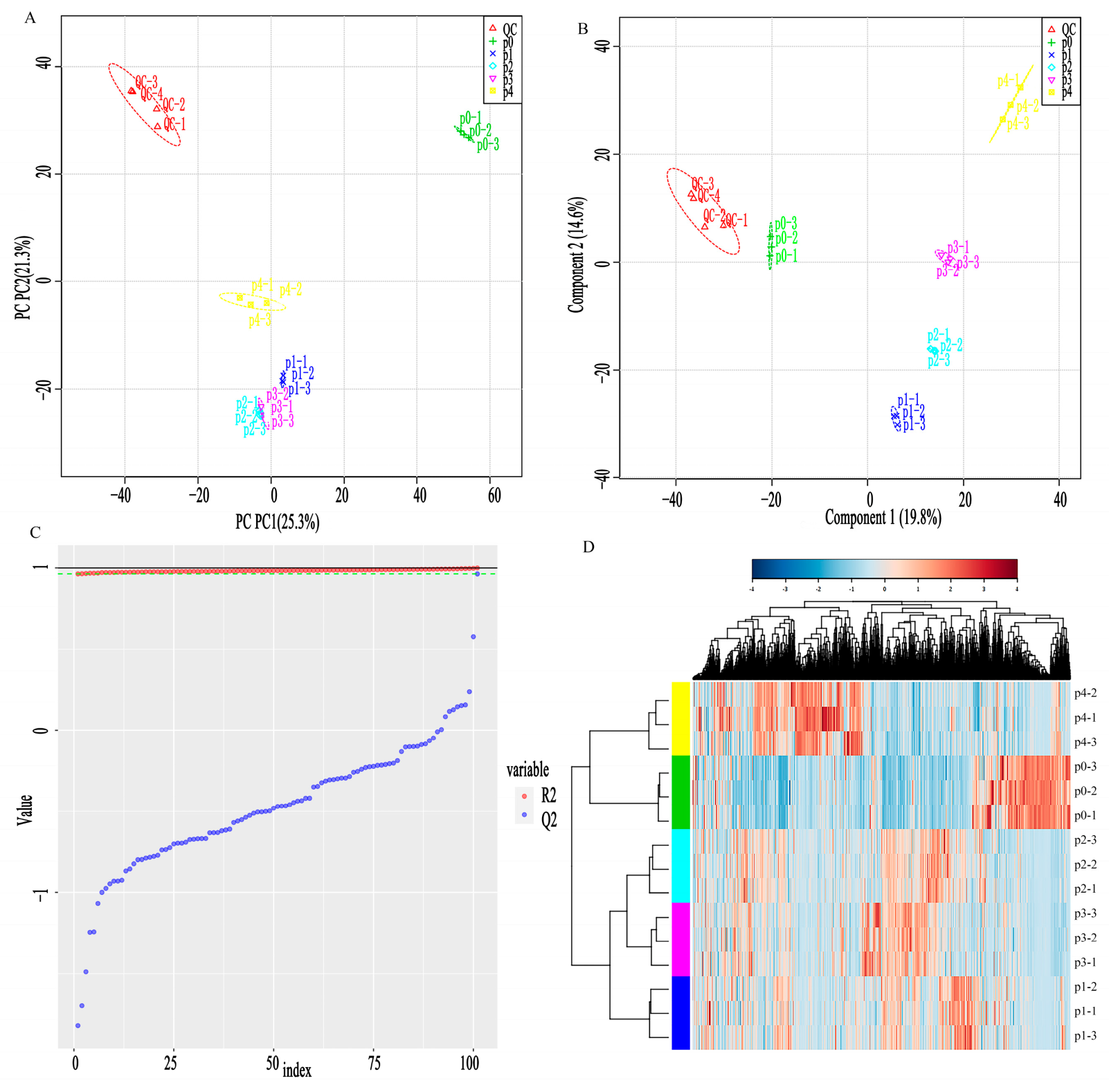
3.1. Total Ionogram Analysis of Samples with LC-MS/MS Detection, PCA, and PLS-DA Analysis
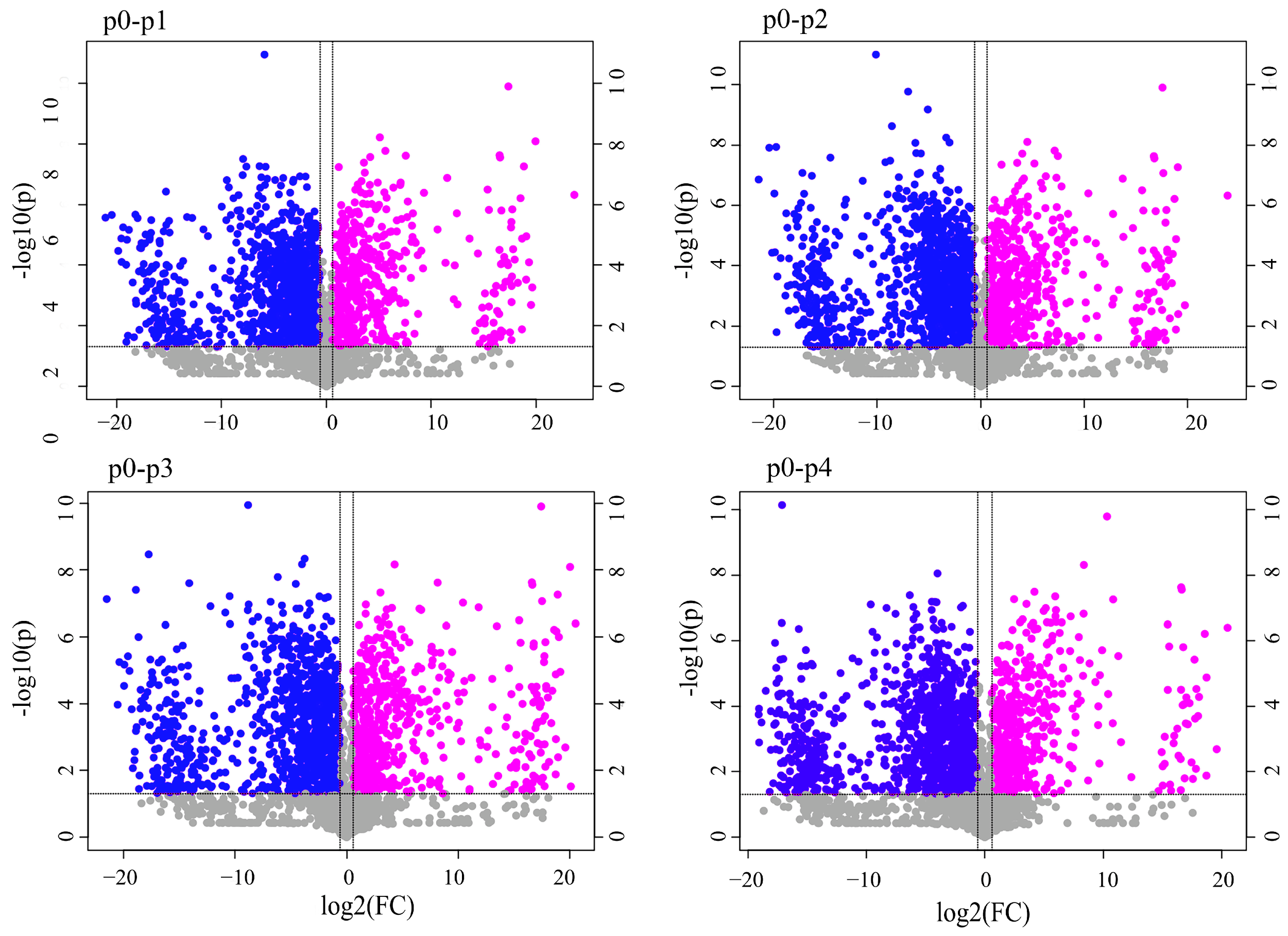
3.2. Screening for Differential Metabolites between Samples
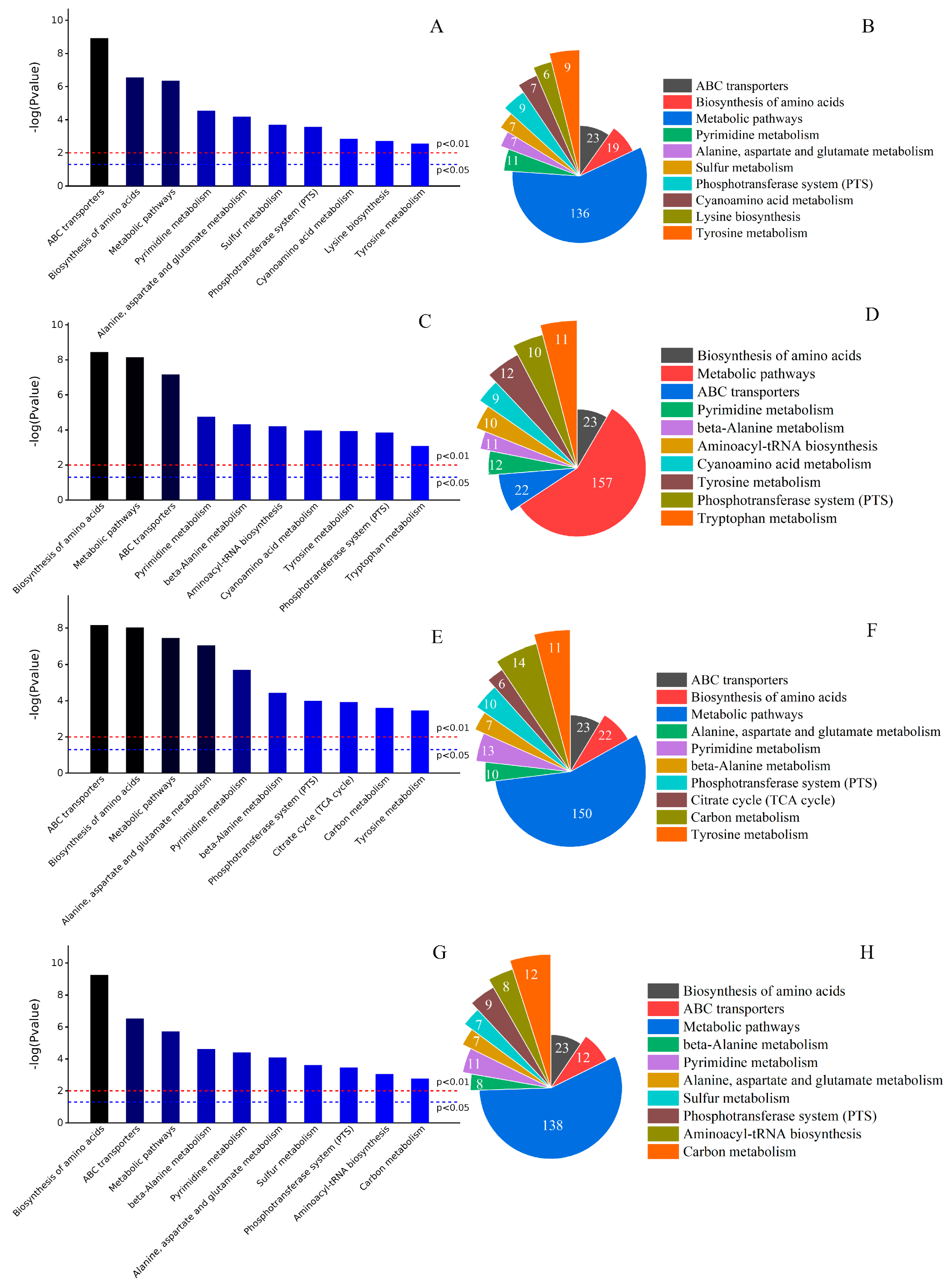
3.3. Differential Metabolite KEGG Metabolic Pathway Analysis
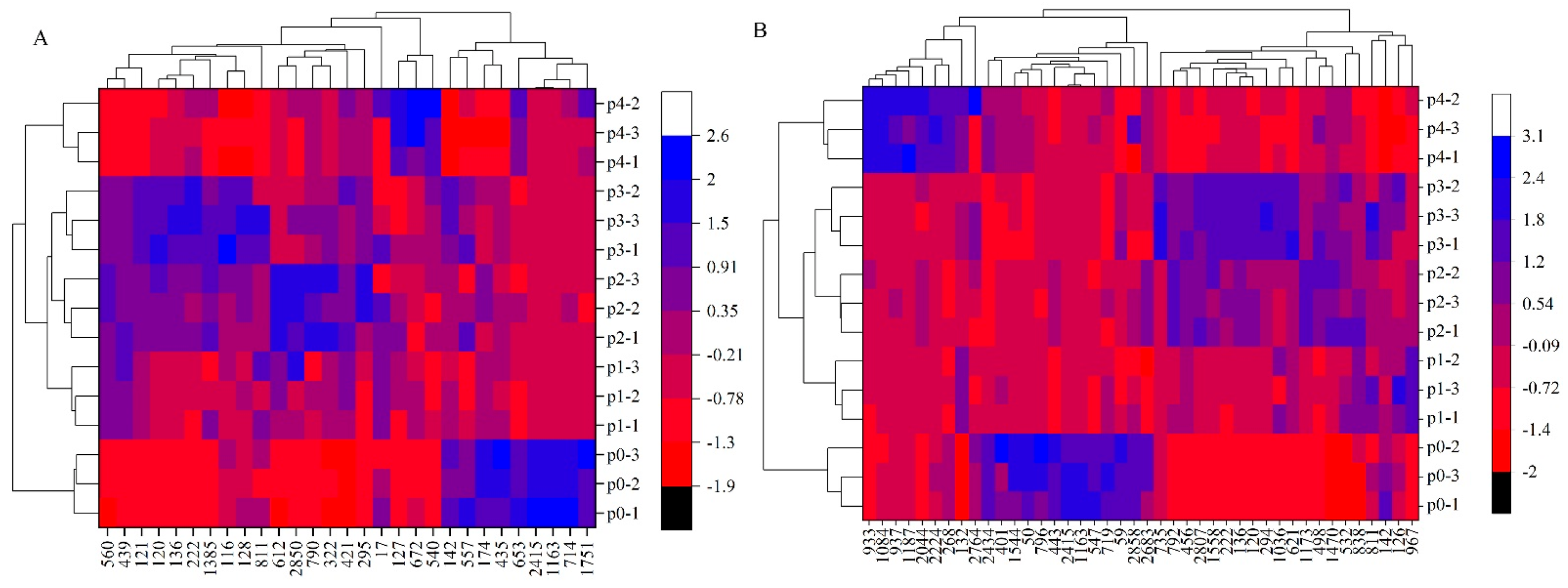
3.4. Differential Metabolite Heatmap Analysis of the Biosynthesis of Amino Acids and ABC Transporters
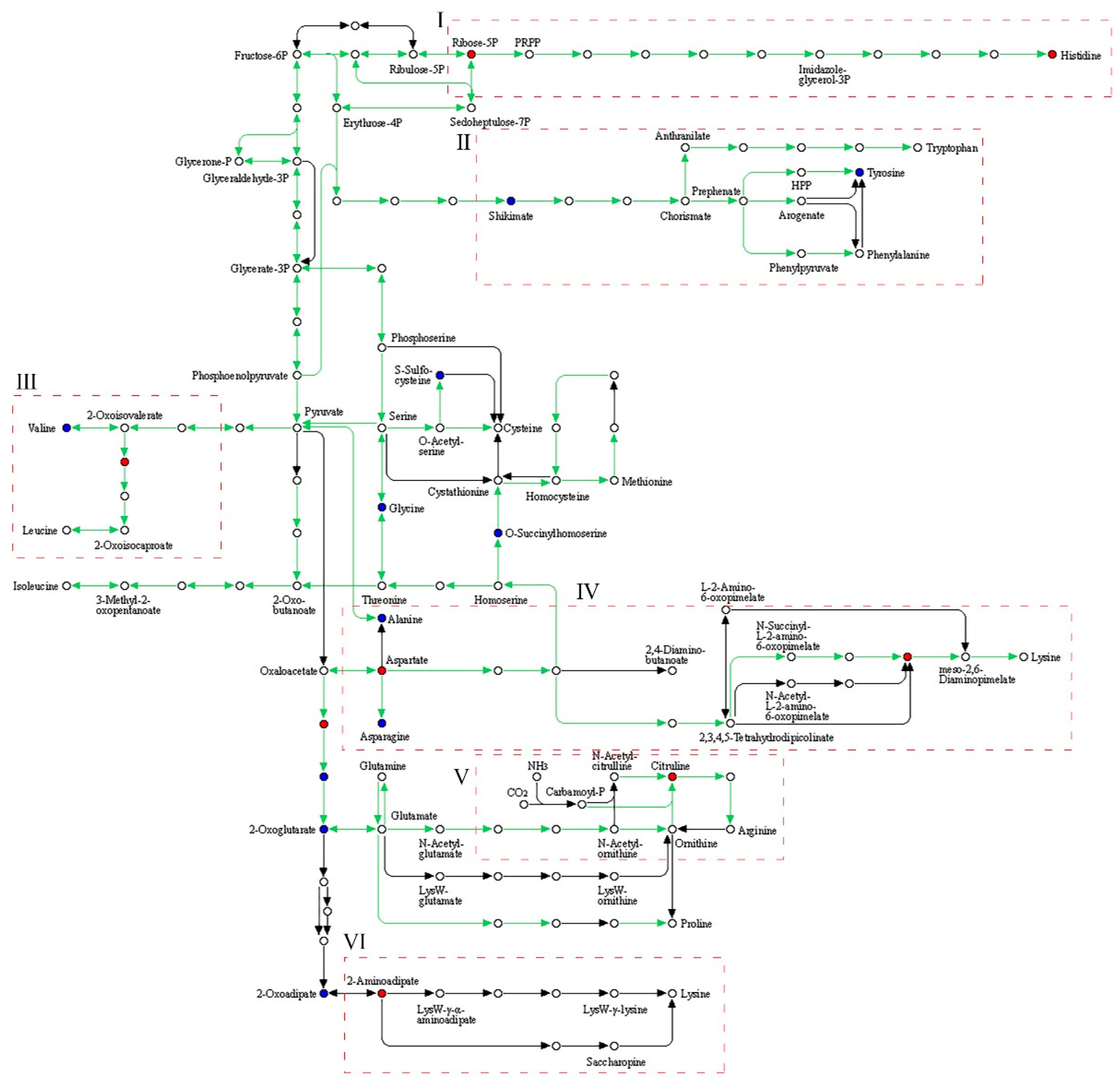
3.5. Metabolic Pathway Analysis of the Biosynthesis of Amino Acids
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, C.H.; He, H.; He, X.G.; Shao, X.H.; Kuang, S.Z.; Xiao, W.Q. Pineapple cultivar structure and extrension of new idependent breeding pineapple cultivars in China. China Trop. Agric. 2021, 4, 13–15. [Google Scholar] [CrossRef]
- Van Tran, T.; Nguyen, D.T.C.; Nguyen, T.T.T.; Nguyen, D.H.; Alhassan, M.; Jalil, A.; Nabgan, W.; Lee, T. A critical review on pineapple (Ananas comosus) wastes for water treatment, challenges and future prospects towards circular economy. Sci. Total. Environ. 2023, 856, 158817. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ou, Z.Q.; Deng, Y.G.; Cui, Z.D.; Li, L.; Zhang, J.; Wang, Y.Q.; Wei, L.J.; Liu, S.L. Development and experiments of type 1JHB-150 pineapple leaf shattering and returning machine. J. Agric. Mech. Res. 2019, 41, 103–109. [Google Scholar] [CrossRef]
- Zhou, W.; Ye, C.; Geng, L.; Chen, G.; Wang, X.; Chen, W.; Sa, R.; Zhang, J.; Zhang, X. Purification and characterization of bromelain from pineapple (Ananas comosus L.) peel waste. J. Food Sci. 2021, 86, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Ramli, A.N.M.; Manas, N.H.A.; Hamid, A.A.A.; Hamid, H.A.; Illias, R.M. Comparative structural analysis of fruit and stem bromelain from Ananas comosus. Food Chem. 2018, 266, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Asano, S.; Ogata, H.; Ito, S.; Nakase, T.; Takeda, S.; Miyoshi, K.; Numata, Y.; Takahashi, K.; Kajikawa, H. Digestive, fermentative, and physical properties of pineapple residue as a feed for cattle. Anim. Sci. J. 2021, 92, e13535. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Fu, B.M.; Wang, Y.; Ye, C. Pharmacological activity of pineapple protease and its application in livestock and poultry diseases. Anim. Breed. Feed. 2020, 2, 64–66. [Google Scholar] [CrossRef]
- Bahri, S.; Hadati, K.S.; Satrimafitrah, P. Production of protein hydrolysate from tofu dregs using the crude extract of bromelain from pineapple core (Ananas comosus l). J. Physics: Conf. Ser. 2021, 1763, 012008. [Google Scholar] [CrossRef]
- Wu, Z.M.; Peng, S.; Wang, Z.; Yang, K.; Qiu, X.Y.; Wu, P.X.; Xu, Y.W.; Wang, J.L.; Yin, F.Q. Evaluation of Lactic acid bacteria and cellulase on silage fermentation quality and fermentation in vitro of pineapple residue. Chin. J. Anim. Nutr. 2022, 12, 7981–7992. [Google Scholar] [CrossRef]
- Deng, Y.G.; Wang, J.L.; Sun, W.S.; Jiao, J.; Zheng, Y.; Wang, G. Effect of dry matter concentration on dry anaerobic fermentation for biogas production in pineapple leaf residues. Anhui Agric. Sci. 2010, 38, 18607–18610. [Google Scholar] [CrossRef]
- Sajjanshetty, R.; Kulkarni, N.S.; Shankar, K.; Jayalakshmi, S.; Sreeramulu, K. Enhanced production and in-situ removal of butanol during the fermentation of lignocellulosic hydrolysate of pineapple leaves. Ind. Crop. Prod. 2021, 173, 114147. [Google Scholar] [CrossRef]
- Ling, G.W.; Jia, C.Y.; Ze, Y.S.; Qi, W.; Fang, Z.; Zhi, H.J.; Cong, L.X.; Fan, R.P.; Zhong, A.L.; Li, N. Ganoderic acids-rich ethanol extract from Ganoderma lucidum protects against alcoholic liver injury and modulates intestinal microbiota in mice with excessive alcohol intake. Curr. Res. Food Sci. 2022, 5, 515–530. [Google Scholar] [CrossRef]
- Wu, T.; Liu, X.; Wang, T.; Tian, L.; Qiu, H.; Ge, F.; Zhu, J.; Shi, L.; Jiang, A.; Yu, H.; et al. Heme Oxygenase/Carbon Monoxide Participates in the Regulation of Ganoderma lucidum Heat-Stress Response, Ganoderic Acid Biosynthesis, and Cell-Wall Integrity. Int. J. Mol. Sci. 2022, 23, 13147. [Google Scholar] [CrossRef]
- Liu, L.; Feng, J.; Gao, K.; Zhou, S.; Yan, M.; Tang, C.; Zhou, J.; Liu, Y.; Zhang, J. Influence of carbon and nitrogen sources on structural features and immunomodulatory activity of exopolysaccharides from Ganoderma lucidum. Process. Biochem. 2022, 119, 96–105. [Google Scholar] [CrossRef]
- Tang, C.W.; Zhang, J.S.; Liu, Y.F.; Tang, Y.; Wang, P.; Feng, J. Effects of pH on metabolites and antioxidant activities of Ganoderma lingzhi liquid fermentation. Mycosystema 2023, 42, 570–583. [Google Scholar] [CrossRef]
- Gu, L.; Zhong, X.; Lian, D.; Zheng, Y.; Wang, H.; Liu, X. Triterpenoid biosynthesis and the transcriptional response elicited by nitric oxide in submerged fermenting Ganoderma lucidum. Process. Biochem. 2017, 60, 19–26. [Google Scholar] [CrossRef]
- Zhang, B.; Zhou, J.; Li, Q.; Gan, B.; Peng, W.; Zhang, X.; Tan, W.; Jiang, L.; Li, X. Manganese affects the growth and metabolism of Ganoderma lucidum based on LC-MS analysis. PeerJ 2019, 7, e6846. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Liu, L.-P.; Wang, Y.-Z.; Yang, L.; Zhang, C.; Yue, M.-X.; Dabbour, M.; Mintah, B.K.; Wang, L. Effect of ultrasonication on the metabolome and transcriptome profile changes in the fermentation of Ganoderma lucidum. Microbiol. Res. 2021, 254, 126916. [Google Scholar] [CrossRef]
- Ma, X.; Wang, P.; Zhou, S.; Sun, Y.; Liu, N.; Li, X.; Hou, Y. De novo transcriptome sequencing and comprehensive analysis of the drought-responsive genes in the desert plant Cynanchum komarovii. BMC Genom. 2015, 16, 753. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Song, S.Q.; Yue, S.N.; Lian, L.D.; Zhao, M.W. Biological function of the transcription factor AreA in Ganoderma lingzhi. Mycosystema 2020, 1, 57–65. [Google Scholar] [CrossRef]
- Ahmed, O.H.; Husni, M.H.A.; Anuar, A.R.; Hanafi, M.M. Production of Humic Acid from Pineapple Leaf Residue. J. Sustain. Agric. 2003, 22, 113–124. [Google Scholar] [CrossRef]
- Ahmed, O.H.; Husni, A.M.; Anuar, R.A.; Hanafi, M.M. Alternative means of recycling pineapple leaf residues. Fruits 2003, 58, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.L.; Tang, M.; Zhang, X.M.; Tang, J. Analysis of metabolites change from flowering to withering of Rhododendron delavayi based on LC-MS/MS. Guihaia 2022, 42, 1170–1180. [Google Scholar] [CrossRef]
- Li, Z.N.; Liu, C.; Wu, Q.F.; Wang, F.; Chen, H.P.; Liu, Y.P. Metabolomics analysis of eucommiae cortex obtained by different processing and drying methods based on ultra-performance liquid chromatography-tandem mass spectrometry. J. Instrum. Anal. 2022, 41, 998–1006. [Google Scholar] [CrossRef]
- Ji, B.C.; Li, Z.G.; Huang, Y.C.; Zhou, W.X.; Yang, Y.; Du, T. Comparative analysis of metabolomes in fruits of different mulberry varieties based on LC-MS technology. Acta Sericologica Sin. 2022, 48, 189–197. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef]
- Yi-Jun, L.; Yang-Yang, Q.; Bing, S.; Yang-Yang, L.; Xing-Hao, T.; Hong-Jun, O.; Yahui, L.; Ge, T.; Zi-Wei, Y.; Fei, C.; et al. Effects of four drying methods on Ganoderma lucidum volatile organic compounds analyzed via headspace solid-phase microextraction and comprehensive two-dimensional chromatography-time-of-flight mass spectrometry. Microchem. J. 2021, 166, 106258. [Google Scholar] [CrossRef]
- Yang, J.B.; Wang, Q.; Wang, X.T.; Chen, X.L.; Wang, Y.; Gao, H.Y.; Song, Y.F.; Wei, F.; Ma, S.C. Difference analysis of chemical composition of polygoni multiflori radix and polygoni multiflori radix preparata based on plant metabonomics. Chin. J. Pharmacovigil. 2022, 19, 615–619. [Google Scholar] [CrossRef]
- Huang, R.S.; Zhan, R.J.; Chen, B.H.; Wang, Y.; Liao, B.Y.; Zhang, Q.W.; Li, C.N. Comparative study on the quality of mulberry leaf vegetable from different mulberry varieties based on metabolomics. Acta Sericologica Sin. 2022, 48, 340–350. [Google Scholar] [CrossRef]
- Gu, Y.; Zang, P.; Li, J.; Yan, Y.; Wang, J. Plasma metabolomics in a deep vein thrombosis rat model based on ultra-high performance liquid chromatography-electrostatic field orbitrap high resolution mass spectrometry. Chin. J. Chromatogr. 2022, 40, 736–745. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhang, F.-L.; Li, J.-R.; Peng, P.-Z.; Liu, B.; Zhao, L.-N. Ganoderma lucidum protease hydrolyzate on lipid metabolism and gut microbiota in high-fat diet fed rats. Food Biosci. 2021, 47, 101460. [Google Scholar] [CrossRef]
- Lv, X.-C.; Wu, Q.; Cao, Y.-J.; Lin, Y.-C.; Guo, W.-L.; Rao, P.-F.; Zhang, Y.-Y.; Chen, Y.-T.; Ai, L.-Z.; Ni, L. Ganoderic acid A from Ganoderma lucidum protects against alcoholic liver injury through ameliorating the lipid metabolism and modulating the intestinal microbial composition. Food Funct. 2022, 13, 5820–5837. [Google Scholar] [CrossRef]
- Viroel, F.J.M.; Laurino, L.F.; Caetano, L.A.; Jozala, A.F.; Spim, S.R.V.; Pickler, T.B.; Sercundes, M.K.; Gomes, M.C.; Hataka, A.; Grotto, D.; et al. Ganoderma lucidum Modulates Glucose, Lipid Peroxidation and Hepatic Metabolism in Streptozotocin-Induced Diabetic Pregnant Rats. Antioxidants 2022, 11, 1035. [Google Scholar] [CrossRef]
- Li, H.; Liu, J.; Hou, Z.; Luo, X.; Lin, J.; Jiang, N.; Hou, L.; Ma, L.; Li, C.; Qu, S. Activation of mycelial defense mechanisms in the oyster mushroom Pleurotus ostreatus induced by Tyrophagus putrescentiae. Food Res. Int. 2022, 160, 111708. [Google Scholar] [CrossRef]
- Kato, M.; Urabe, S.; Matsuzawa, S.; Kato, A.; Fukazawa, M.; Hiyama, E.; Kurii, A.; Mikami, N.; Kitajima, Y.; Hida, M.; et al. High-volume pre-dilution on-line hemodiafiltration is the adequate blood purification method from the viewpoint of amino acid nutrition. Ren. Replace. Ther. 2022, 8, 51. [Google Scholar] [CrossRef]
- Li, J.L.; Guo, C.Z.; Wang, S.J.; Wang, J.B.; Sui, S.S. Research advance on biological activities and application of lysine. China Feed. 2020, 21, 63–66. [Google Scholar] [CrossRef]
- Zhang, Y.M. Effects of Dietary Lysine Supplementation on Wool Production and Gut Microbes of Angora Rabbits. Master’s Thesis, Shandong Agricultural University, Taian, China, 2022. [Google Scholar] [CrossRef]
- An, S.H.; Kang, H.-K.; Kong, C. Standardized ileal digestible lysine requirements of 21–28 days old male broilers. Anim. Feed. Sci. Technol. 2022, 292, 115409. [Google Scholar] [CrossRef]
- Baldi, G.; Soglia, F.; Laghi, L.; Meluzzi, A.; Petracci, M. The role of histidine dipeptides on postmortem acidification of broiler muscles with different energy metabolism. Poult. Sci. 2020, 100, 1299–1307. [Google Scholar] [CrossRef]
- Kasiga, T.; White, B.M.; Bruce, T.J.; Brown, M.L. Effect of fish meal replacement with CarinataBrassica carinatain low animal protein diets of rainbow troutOncorhynchus mykiss (Walbaum) on trypsin activity, protein and amino acid digestibility and bioavailability. Aquac. Res. 2020, 51, 2134–2149. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Qian, Y.; Wang, C.; He, Y.; Zhu, C.; Chen, G.; Lin, L.; Chen, Y. Study of the Metabolite Changes in Ganoderma lucidum under Pineapple Leaf Residue Stress via LC-MS/MS Coupled with a Non-Targeted Metabolomics Approach. Metabolites 2023, 13, 487. https://doi.org/10.3390/metabo13040487
Liu Y, Qian Y, Wang C, He Y, Zhu C, Chen G, Lin L, Chen Y. Study of the Metabolite Changes in Ganoderma lucidum under Pineapple Leaf Residue Stress via LC-MS/MS Coupled with a Non-Targeted Metabolomics Approach. Metabolites. 2023; 13(4):487. https://doi.org/10.3390/metabo13040487
Chicago/Turabian StyleLiu, Yijun, Yangyang Qian, Chunyu Wang, Yingying He, Chuxing Zhu, Gang Chen, Lijing Lin, and Yuliang Chen. 2023. "Study of the Metabolite Changes in Ganoderma lucidum under Pineapple Leaf Residue Stress via LC-MS/MS Coupled with a Non-Targeted Metabolomics Approach" Metabolites 13, no. 4: 487. https://doi.org/10.3390/metabo13040487