
NMR-Based Metabolomics Reveals Effects of Water Stress in the Primary and Specialized Metabolisms of Bauhinia ungulata L. (Fabaceae)

 , ,
, ,
Abstract
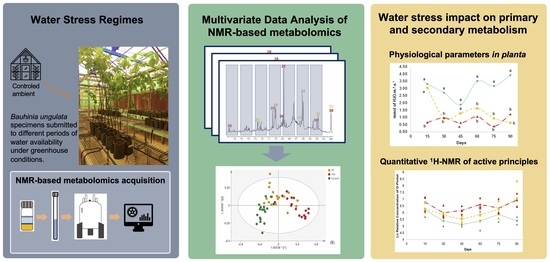
:
1. Introduction
2. Materials and Methods
2.1. Plant Material and Cultivation
2.2. Drought Conditions
2.3. Sampling Procedure
2.4. Preparation of the Extracts
2.5. UHPLC-ESI-HRMS Analysis
2.6. UHPLC-HRMS2 Data Processing
2.7. Molecular Network Analysis and Computational Annotation
2.8. 1H-NMR and 2D-NMR Experiments
2.9. Quantitative Analysis by 1H-NMR (qHNMR)
2.10. Physiological Parameters Analyses
2.11. Statistical Analysis
3. Results and Discussion
3.1. Metabolite Profiling via LC-HRMS
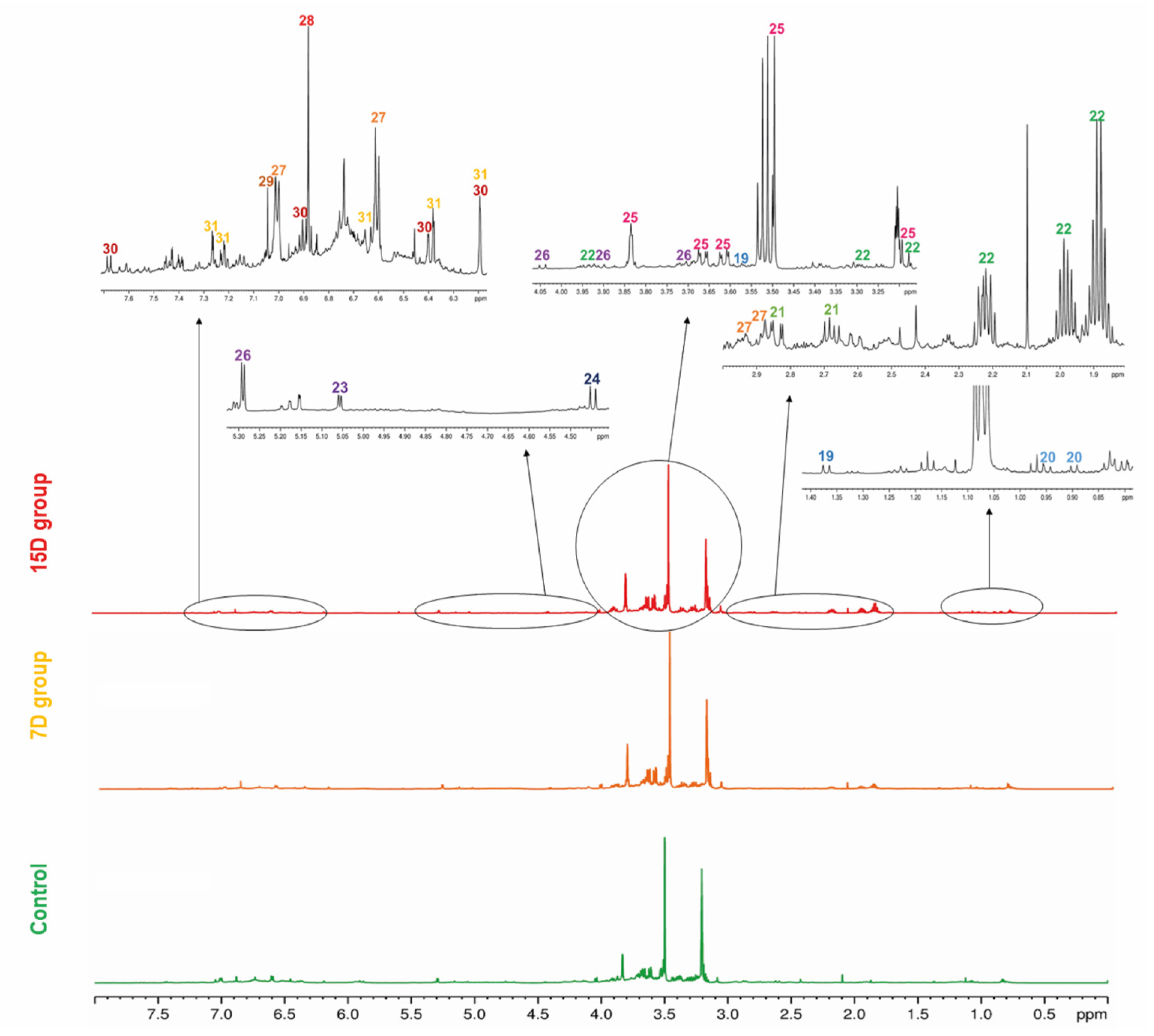
3.2. Metabolite Profiling via NMR
3.3. NMR-Based Metabolomics
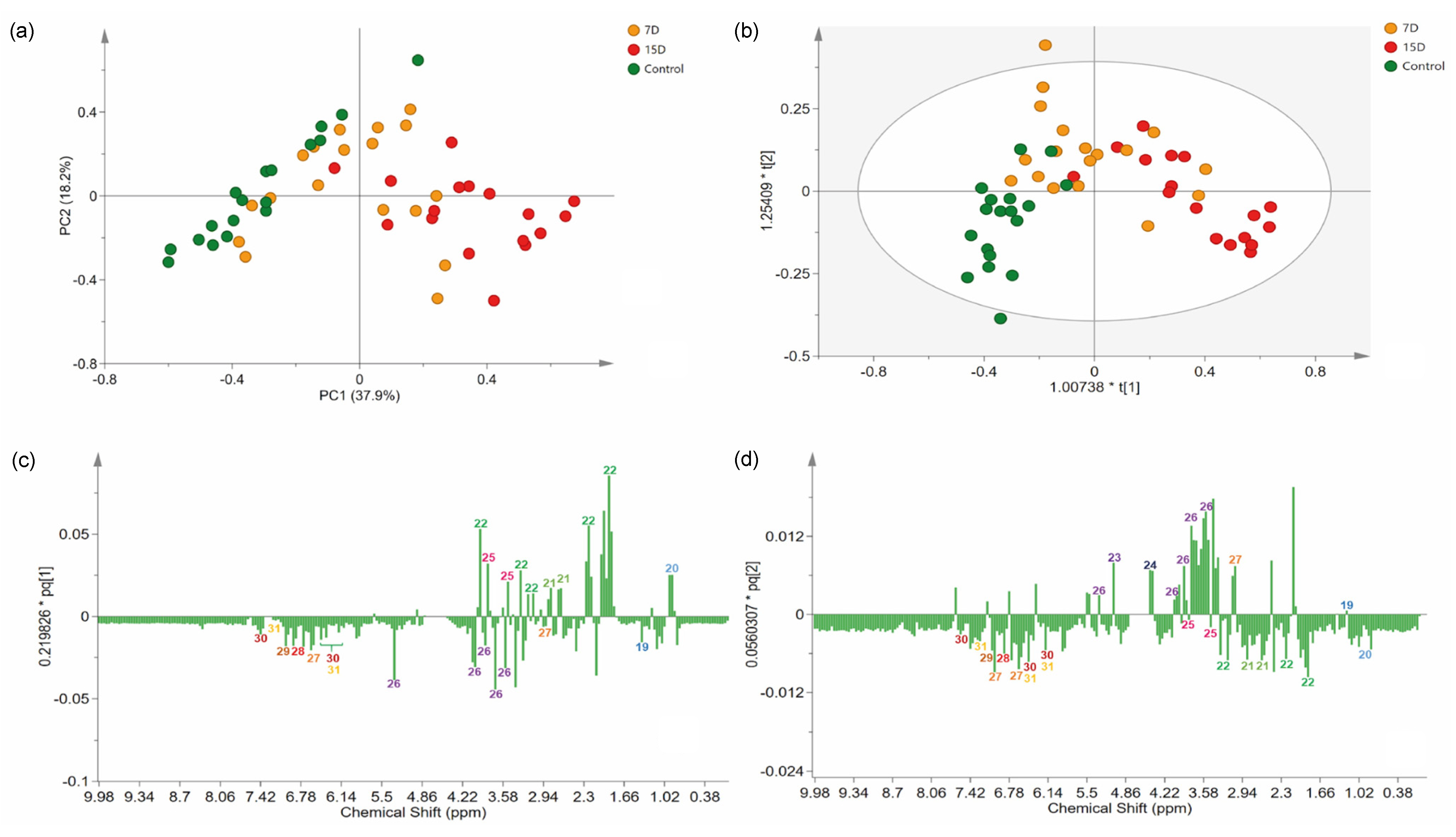
3.4. Multivariate Data Analyses
3.5. Identification of Stress Biomarkers for Drought
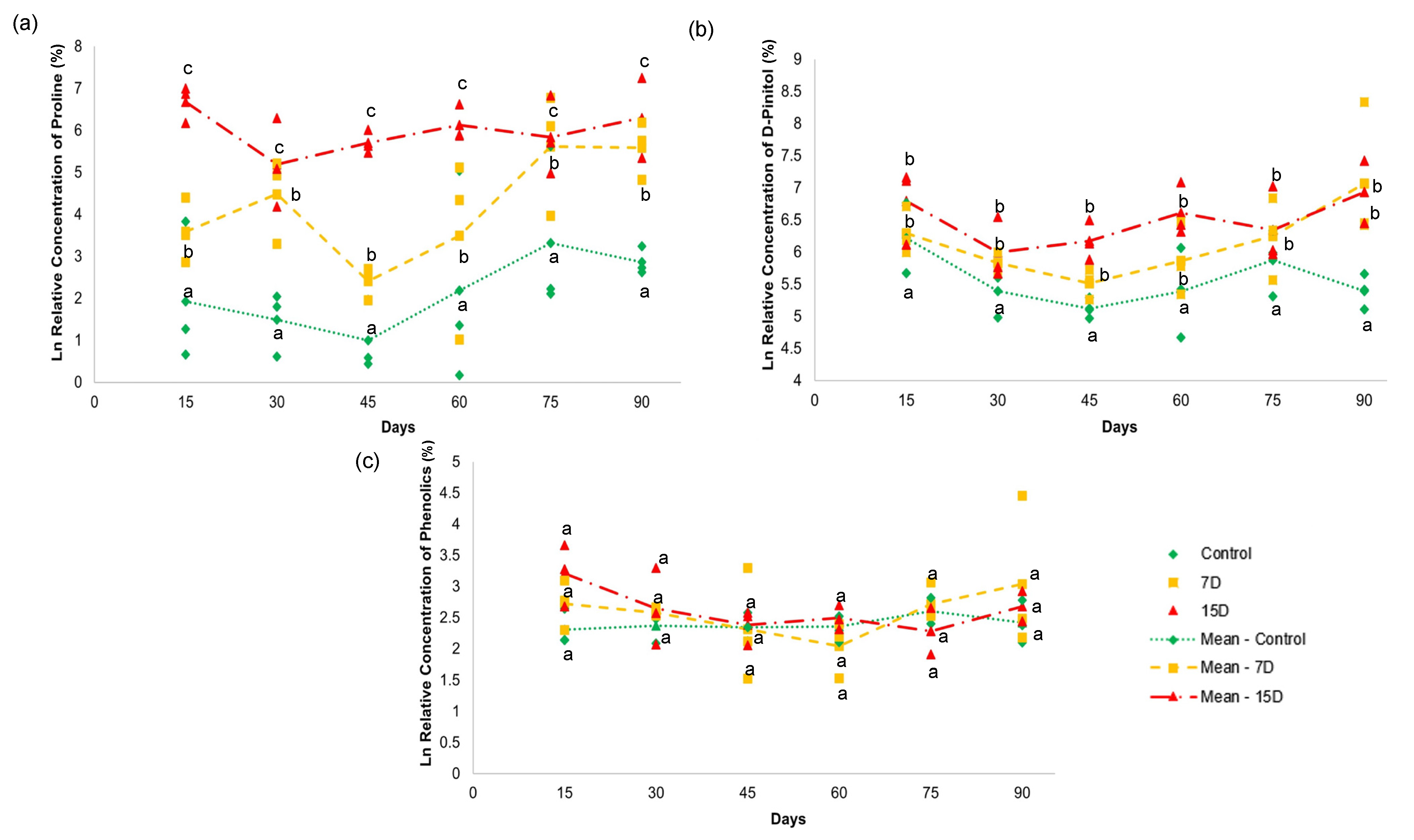
3.6. Quantitative Analysis by NMR
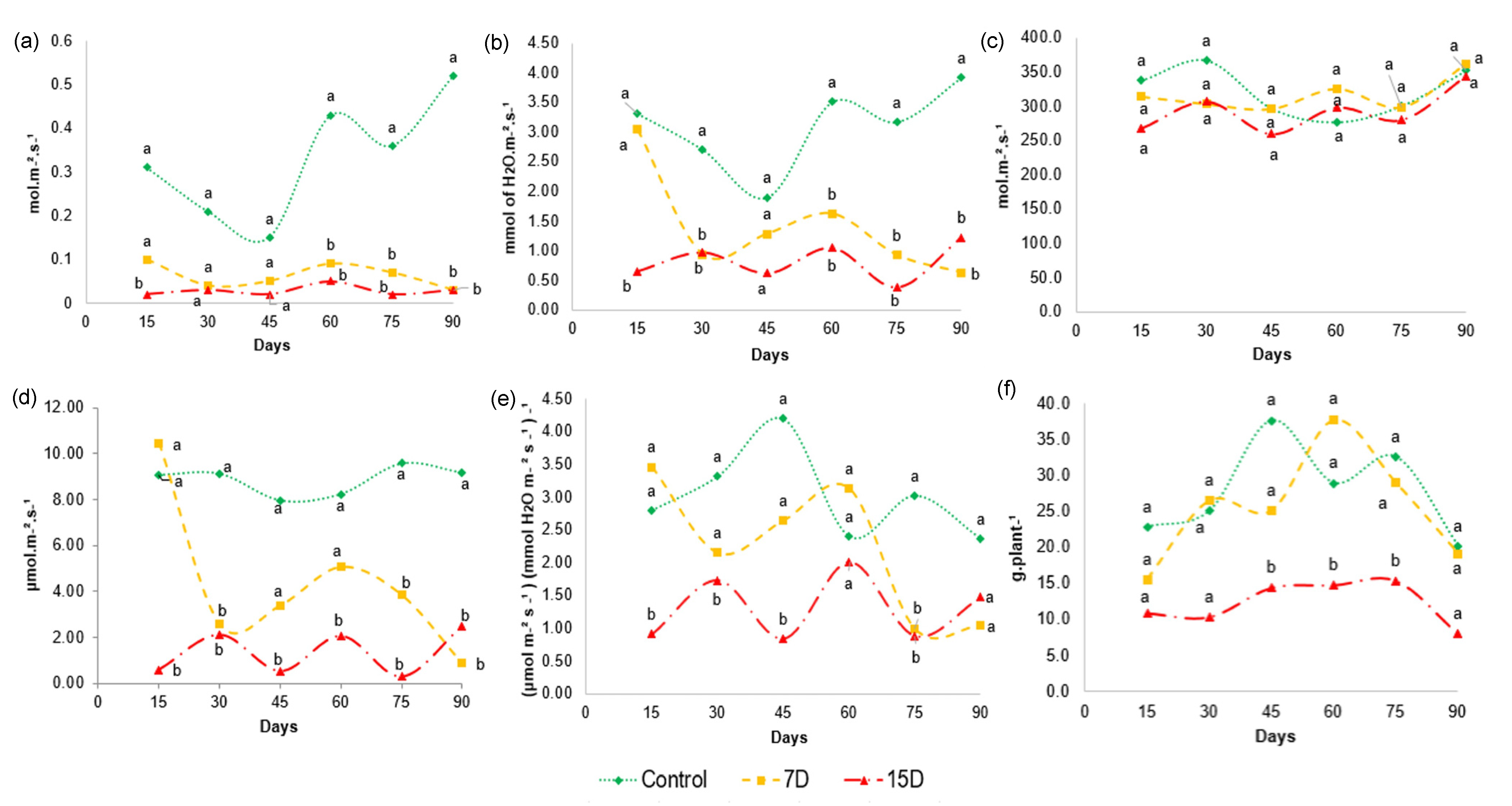
3.7. Physiological Parameters
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Morais, S.M.; Dantas, J.D.P.; Da Silva, A.R.A.; Magalhães, E.F. Plantas medicinais usadas pelos índios Tapebas do Ceará. Rev. Bras. Farm. 2005, 15, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Cavassan, O.; Weiser, V.D.L. Vascular flora of the cerrado of Bauru-SP. Biota Neotrop. 2015, 15. [Google Scholar] [CrossRef] [Green Version]
- Bieski, I.G.C.; Rios-Santos, F.; de Oliveira, R.M.; Espinosa, M.M.; Macedo, M.L.; Albuquerque, U.P.; de Oliveira Martins, D.T. Ethnopharmacology of Medicinal Plants of the Pantanal Region (Mato Grosso, Brazil). Evid. Based Complement. Altern. Med. 2012, 2012, 272749. [Google Scholar] [CrossRef] [Green Version]
- Neto, M.M.; Filho, R.B.; Lima, M.; Silveira, E. Flavonoids and alkaloids from leaves of Bauhinia ungulata L. Biochem. Syst. Ecol. 2008, 36, 227–229. [Google Scholar] [CrossRef]
- De Sousa, L.M.; de Carvalho, J.L.; da Silva, H.C.; Lemos, T.L.G.; Arriaga, A.M.C.; Braz-Filho, R.; Militão, G.C.G.; Silva, T.D.S.; Ribeiro, P.R.V.; Santiago, G.M.P. New Cytotoxic Bibenzyl and Other Constituents from Bauhinia ungulate L. (Fabaceae). Chem. Biodivers. 2016, 13, 1630–1635. [Google Scholar] [CrossRef]
- Paula, C.S. Phytochemical Study and Biological Properties of Bauhinia ungulata L. Leaves, Fabaceae. Ph.D. Thesis, Federal University of Paraná, Curitiba, Brazil, 2014. [Google Scholar]
- Borges, C.V.; Minatel, I.O.; Gomez-Gomez, H.A.; Lima, G.P.P. Medicinal Plants: Influence of Environmental Factors on the Content of Secondary Metabolites. In Medicinal Plants and Environmental Challenges; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 259–277. [Google Scholar]
- Kliebenstein, D.J. Making new molecules—Evolution of structures for novel metabolites in plants. Curr. Opin. Plant Biol. 2013, 16, 112–117. [Google Scholar] [CrossRef]
- Thakur, M.; Bhattacharya, S.; Khosla, P.K.; Puri, S. Improving production of plant secondary metabolites through biotic and abiotic elicitation. J. Appl. Res. Med. Aromat. Plants 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Li, Y.; Kong, D.; Fu, Y.; Sussman, M.R.; Wu, H. The effect of developmental and environmental factors on secondary metabolites in medicinal plants. Plant Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef]
- Ramakrishna, A.; Ravishankar, G.A. Influence of Abiotic Stress Signals on Secondary Metabolites in Plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar]
- Jeelani, S.M.; Rather, G.A.; Sharma, A.; Lattoo, S.K. In perspective: Potential medicinal plant resources of Kashmir Himalayas, their domestication and cultivation for commercial exploitation. J. Appl. Res. Med. Aromat. Plants 2018, 8, 10–25. [Google Scholar] [CrossRef]
- Doerfler, H.; Lyon, D.; Nägele, T.; Sun, X.; Fragner, L.; Hadacek, F.; Egelhofer, V.; Weckwerth, W. Granger causality in integrated GC–MS and LC–MS metabolomics data reveals the interface of primary and secondary metabolism. Metabolomics 2013, 9, 564–574. [Google Scholar] [CrossRef] [Green Version]
- Fazani, R.; Sanches, E.; Alves Da Silva, E. Changes in Leaf Water Potential and Photosynthesis of Bauhinia Forficata Link under Water Deficit and after Rehydration. Hoehnea 2013, 40, 181–190. [Google Scholar]
- Glaubitz, U.; Li, X.; Schaedel, S.; Erban, A.; Sulpice, R.; Kopka, J.; Hincha, D.K.; Zuther, E. Integrated analysis of rice transcriptomic and metabolomic responses to elevated night temperatures identifies sensitivity- and tolerance-related profiles. Plant Cell Environ. 2017, 40, 121–137. [Google Scholar] [CrossRef] [PubMed]
- Vaz, A.M.S.D.F.; Tozzi, A.M.G.A. Bauhinia ser. Cansenia (Leguminosae: Caesalpinioideae) no Brasil. Rodriguesia 2003, 54, 55–143. [Google Scholar] [CrossRef] [Green Version]
- Pennington, R.T.; Lewis, G.; Ratter, J. An Overview of the Plant Diversity, Biogeography and Conservation of Neotropical Savannas and Seasonally Dry Forests; CRC Press: Boca Raton, FL, USA, 2006; pp. 1–29. [Google Scholar]
- Kim, H.K.; Choi, Y.H.; Verpoorte, R. NMR-based metabolomic analysis of plants. Nat. Protoc. 2010, 5, 536–549. [Google Scholar] [CrossRef] [PubMed]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [Green Version]
- Nothias, L.F.; Petras, D.; Schmid, R.; Dührkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-Based Molecular Networking in the GNPS Analysis Environment. bioRxiv 2019, 10, 812404. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Allard, P.-M.; Péresse, T.; Bisson, J.; Gindro, K.; Marcourt, L.; Pham, V.C.; Roussi, F.; Litaudon, M.; Wolfender, J.-L. Integration of Molecular Networking and In-Silico MS/MS Fragmentation for Natural Products Dereplication. Anal. Chem. 2016, 88, 3317–3323. [Google Scholar] [CrossRef]
- Pauli, G.F.; Jaki, B.U.; Lankin, D.C. A Routine Experimental Protocol for qHNMR Illustrated with Taxol. J. Nat. Prod. 2007, 70, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Holzgrabe, U. Quantitative NMR spectroscopy in pharmaceutical applications. Prog. Nucl. Magn. Reson. Spectrosc. 2010, 57, 229–240. [Google Scholar] [CrossRef]
- Bharti, S.K.; Roy, R. Quantitative 1H NMR spectroscopy. TrAC Trends Anal. Chem. 2012, 35, 5–26. [Google Scholar] [CrossRef]
- Cechin, I.; Corniani, N.; Fumis, T.D.F.; Cataneo, A.C. Ultraviolet-B and water stress effects on growth, gas exchange and oxidative stress in sunflower plants. Radiat. Environ. Biophys. 2008, 47, 405–413. [Google Scholar] [CrossRef]
- Košir, I.J.; Kidrič, J. Identification of Amino Acids in Wines by One- and Two-Dimensional Nuclear Magnetic Resonance Spectroscopy. J. Agric. Food Chem. 2001, 49, 50–56. [Google Scholar] [CrossRef]
- Jafari, T.; Rahikainen, M.; Puljula, E.; Sinkkonen, J.; Kangasjärvi, S. The impact of light intensity on metabolomic profile of Arabidopsis thaliana wild type and reticulata mutant by NMR spectroscopy. Phytochem. Lett. 2018, 26, 170–178. [Google Scholar] [CrossRef]
- Pogliani, L.; Ellenberger, M.; Valat, J. A NMR investigation of proline and its derivatives. II—Conformational implications of the1H NMR spectrum of L-proline at different pH. Org. Magn. Reson. 1975, 7, 61–71. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhao, Y.; Yang, J.; Jiang, Y.; Lu, F.; Jia, Y.; Yang, B. Metabolomic analyses of banana during postharvest senescence by 1H-high resolution-NMR. Food Chem. 2017, 218, 406–412. [Google Scholar] [CrossRef]
- Pfeffer, P.E.; Valentine, K.M.; Parrish, F.W. Deuterium-induced differential isotope shift carbon-13 NMR. 1. Resonance reassignments of mono- and disaccharides. J. Am. Chem. Soc. 1979, 101, 1265–1274. [Google Scholar] [CrossRef]
- Raya-Gonzalez, D.; Pamatz-Bolanõs, T.; del Rio-Torres, R.E.; Munõz, R.E.M.; Ron-Echeverria, O.; Martinez-Pacheco, M.M. D-(+)-Pinitol, a Component of the Heartwood of Enterolobium cyclocarpum (Jacq.) Griseb. Z. Fur Nat.—Sect. C J. Biosci. 2008, 63, 922–924. [Google Scholar] [CrossRef]
- Waibel, M.; De Angelis, M.; Stossi, F.; Kieser, K.J.; Carlson, K.E.; Katzenellenbogen, B.S.; Katzenellenbogen, J.A. Bibenzyl- and stilbene-core compounds with non-polar linker atom substituents as selective ligands for estrogen receptor beta. Eur. J. Med. Chem. 2009, 44, 3412–3424. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.R.; Kim, E.N.; Jeong, G.S. Simultaneous Quantitative Analysis of Flavonoids Isolated from the Leaves of Diospyros Kaki. Korean J. Pharmacogn. 2020, 51, 139–145. [Google Scholar] [CrossRef]
- Sánchez-Hidalgo, M.; León-González, A.J.; Gálvez-Peralta, M.; González-Mauraza, N.H.; Martin-Cordero, C. d-Pinitol: A cyclitol with versatile biological and pharmacological activities. Phytochem. Rev. 2021, 20, 211–224. [Google Scholar] [CrossRef]
- Saldanha, L.L.; Delgado, A.Q.; Marcourt, L.; Camaforte, N.A.D.P.; Vareda, P.M.P.; Ebrahimi, S.N.; Vilegas, W.; Dokkedal, A.L.; Queiroz, E.F.; Wolfender, J.-L.; et al. Hypoglycemic active principles from the leaves of Bauhinia holophylla: Comprehensive phytochemical characterization and in vivo activity profile. PLoS ONE 2021, 16, e0258016. [Google Scholar] [CrossRef]
- Santos, K.M.; Gomes, I.N.F.; Silva-Oliveira, R.J.; Pinto, F.E.; Oliveira, B.G.; Chagas, R.C.R.; Romão, W.; Reis, R.M.V.; Ribeiro, R.I.M.A. Bauhinia variegata candida Fraction Induces Tumor Cell Death by Activation of Caspase-3, RIP, and TNF-R1 and Inhibits Cell Migration and Invasion In Vitro. BioMed. Res. Int. 2018, 2018, 4702481. [Google Scholar] [CrossRef] [Green Version]
- Dewangan, P.; Verma, A.; Kesharwani, D. Isolation of D-Pinitol: A Bioactive Carbohydrate from the Leaves of Bauhinia variegata L. Int. J. Pharm. Sci. Rev. Res. 2014, 24, 43–45. [Google Scholar]
- Lambert, C.; Richard, T.; Renouf, E.; Bisson, J.; Waffo-Téguo, P.; Bordenave, L.; Ollat, N.; Mérillon, J.-M.; Cluzet, S. Comparative Analyses of Stilbenoids in Canes of Major Vitis vinifera L. Cultivars. J. Agric. Food Chem. 2013, 61, 11392–11399. [Google Scholar] [CrossRef]
- Guo, P.; Anderson, J.D.; Bozell, J.J.; Zivanovic, S. The effect of solvent composition on grafting gallic acid onto chitosan via carbodiimide. Carbohydr. Polym. 2016, 140, 171–180. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Li, C.; Zhang, Z.; Ma, F.; Li, M. Response of sugar metabolism in apple leaves subjected to short-term drought stress. Plant Physiol. Biochem. 2019, 141, 164–171. [Google Scholar] [CrossRef]
- Kantar, M.; Lucas, S.J.; Budak, H. Drought Stress: Molecular Genetics and Genomics Approaches; Academic Press: Cambridge, MA, USA, 2011; Volume 57. [Google Scholar]
- Kobylińska, A.; Borek, S.; Posmyk, M.M. Melatonin redirects carbohydrates metabolism during sugar starvation in plant cells. J. Pineal Res. 2018, 64, e12466. [Google Scholar] [CrossRef]
- Du, Y.; Zhao, Q.; Chen, L.; Yao, X.; Zhang, W.; Zhang, B.; Xie, F. Effect of drought stress on sugar metabolism in leaves and roots of soybean seedlings. Plant Physiol. Biochem. 2020, 146, 1–12. [Google Scholar] [CrossRef]
- Shao, H.-B.; Chu, L.-Y.; Jaleel, C.A.; Zhao, C.-X. Water-deficit stress-induced anatomical changes in higher plants. Comptes Rendus Biol. 2008, 331, 215–225. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef] [Green Version]
- McManus, M.T.; Bieleski, R.L.; Caradus, J.R.; Barker, D.J. Pinitol accumulation in mature leaves of white clover in response to a water deficit. Environ. Exp. Bot. 2000, 43, 11–18. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Iqbal, M.; Rasheed, R.; Hussain, I.; Perveen, S.; Mahmood, S. Dynamic Proline Metabolism: Importance and Regulation in Water-Limited Environments. In Plant Metabolites and Regulation under Environmental Stress; Elsevier: Amsterdam, The Netherlands, 2018; pp. 323–336. ISBN 9780128126905. [Google Scholar]
- Choi, Y.-S.; Lee, M.R.; Kim, C.S.; Lee, K.-H. Detection of proline using a novel paper-based analytical device for on-site diagnosis of drought stress in plants. Rev. Sci. Instruments 2019, 90, 45002. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline Mechanisms of Stress Survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.; Lamant, A. Pinitol and myo-inositol accumulation in water-stressed seedlings of maritime pine. Phytochemistry 1988, 27, 3423–3427. [Google Scholar] [CrossRef]
- Noiraud, N.; Maurousset, L.; Lemoine, R. Transport of Polyols in Higher Plants. Plant Physiol. Biochem. 2001, 39, 717–728. [Google Scholar] [CrossRef]
- Srivastava, K.; Tiwari, M.; Dubey, A.; Dubey, A. D-Pinitol—A Natural Phytomolecule and Its Pharmacological Effect. Int. J. Pharm. Life Sci. 2020, 11, 6609–6623. [Google Scholar]
- Hernández-Mijares, A.; Bañuls, C.; Peris, J.E.; Monzó, N.; Jover, A.; Bellod, L.; Victor, V.M.; Rocha, M. A single acute dose of pinitol from a naturally-occurring food ingredient decreases hyperglycaemia and circulating insulin levels in healthy subjects. Food Chem. 2013, 141, 1267–1272. [Google Scholar] [CrossRef]
- Navarro, J.; Decara, J.; Medina-Vera, D.; Tovar, R.; Suarez, J.; Pavón, J.; Serrano, A.; Vida, M.; Gutierrez-Adan, A.; Sanjuan, C.; et al. D-Pinitol from Ceratonia siliqua Is an Orally Active Natural Inositol That Reduces Pancreas Insulin Secretion and Increases Circulating Ghrelin Levels in Wistar Rats. Nutrients 2020, 12, 2030. [Google Scholar] [CrossRef]
- Zhang, C.; Wu, W.; Xin, X.; Li, X.; Liu, D. Extract of ice plant (Mesembryanthemum crystallinum) ameliorates hyperglycemia and modulates the gut microbiota composition in type 2 diabetic Goto-Kakizaki rats. Food Funct. 2019, 10, 3252–3261. [Google Scholar] [CrossRef]
- Radwan, A.; Kleinwächter, M.; Selmar, D. Impact of drought stress on specialised metabolism: Biosynthesis and the expression of monoterpene synthases in sage (Salvia officinalis). Phytochemistry 2017, 141, 20–26. [Google Scholar] [CrossRef]
- Mechri, B.; Tekaya, M.; Hammami, M.; Chehab, H. Effects of drought stress on phenolic accumulation in greenhouse-grown olive trees (Olea europaea). Biochem. Syst. Ecol. 2020, 92, 104112. [Google Scholar] [CrossRef]
- Hodaei, M.; Rahimmalek, M.; Arzani, A.; Talebi, M. The effect of water stress on phytochemical accumulation, bioactive compounds and expression of key genes involved in flavonoid biosynthesis in Chrysanthemum morifolium L. Ind. Crop. Prod. 2018, 120, 295–304. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Oh, M.-M. Mild water deficit increases the contents of bioactive compounds in dropwort. Hortic. Environ. Biotechnol. 2017, 58, 458–466. [Google Scholar] [CrossRef]
- Ahmad, P.; Jaleel, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of enzymatic and nonenzymatic antioxidants in plants during abiotic stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef]
- Hichem, H.; Mounir, D.; Naceur, E.A. Differential responses of two maize (Zea mays L.) varieties to salt stress: Changes on polyphenols composition of foliage and oxidative damages. Ind. Crop. Prod. 2009, 30, 144–151. [Google Scholar] [CrossRef]
- Gharibi, S.; Tabatabaei, B.E.S.; Saeidi, G.; Talebi, M.; Matkowski, A. The effect of drought stress on polyphenolic compounds and expression of flavonoid biosynthesis related genes in Achillea pachycephala Rech.f. Phytochemistry 2019, 162, 90–98. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Liakopoulos, G.; Nikolopoulos, D.; Bresta, P.; Stavroulaki, V.; Sumbele, S. Carbon Gain vs. Water Saving, Growth vs. Defence: Two Dilemmas with Soluble Phenolics as a Joker. Plant Sci. 2014, 227, 21–27. [Google Scholar] [CrossRef]
- Huang, J.; Hammerbacher, A.; Forkelová, L.; Hartmann, H. Release of resource constraints allows greater carbon allocation to secondary metabolites and storage in winter wheat. Plant Cell Environ. 2017, 40, 672–685. [Google Scholar] [CrossRef] [Green Version]
- Pérez-López, U.; Sgherri, C.; Miranda-Apodaca, J.; Micaelli, F.; Lacuesta, M.; Mena-Petite, A.; Quartacci, M.F.; Muñoz-Rueda, A. Concentration of phenolic compounds is increased in lettuce grown under high light intensity and elevated CO2. Plant Physiol. Biochem. 2018, 123, 233–241. [Google Scholar] [CrossRef]
- Per, T.S.; Khan, N.A.; Reddy, P.S.; Masood, A.; Hasanuzzaman, M.; Khan, M.I.R.; Anjum, N.A. Approaches in modulating proline metabolism in plants for salt and drought stress tolerance: Phytohormones, mineral nutrients and transgenics. Plant Physiol. Biochem. 2017, 115, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Zali, A.G.; Ehsanzadeh, P. Exogenous proline improves osmoregulation, physiological functions, essential oil, and seed yield of fennel. Ind. Crop. Prod. 2018, 111, 133–140. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, L.; Shen, Q.; Yang, J.; Han, X.; Tian, F.; Wu, J. Effects of Water Stress on Photosynthesis, Yield, and Water Use Efficiency in Winter Wheat. Water 2020, 12, 2127. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Moller, I.M.; Murphy, A. Fisiologia e Desenvolvimento Vegetal Diversidade Vegetal; Artmed Editora: Porto Alegre, Brazil, 2017; Volume 6, ISBN 978-85-8271-366-2. [Google Scholar]
- Heckenberger, U.; Roggatz, U.; Schurr, U. Effect of Drought Stress on the Cytological Status in Ricinus Communis. J. Exp. Bot. 1998, 49, 181–189. [Google Scholar] [CrossRef]







{kind=link}
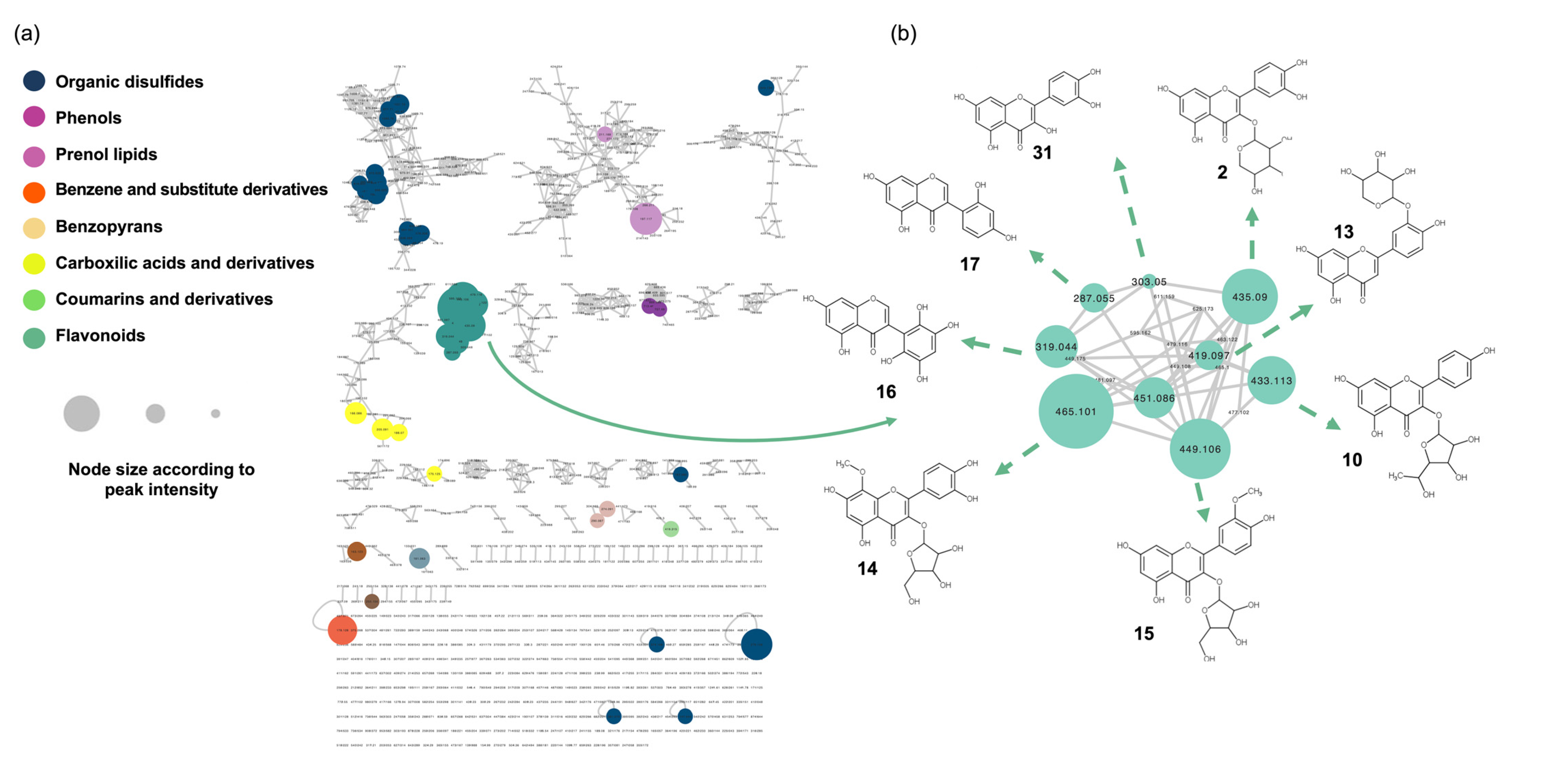{kind=link}
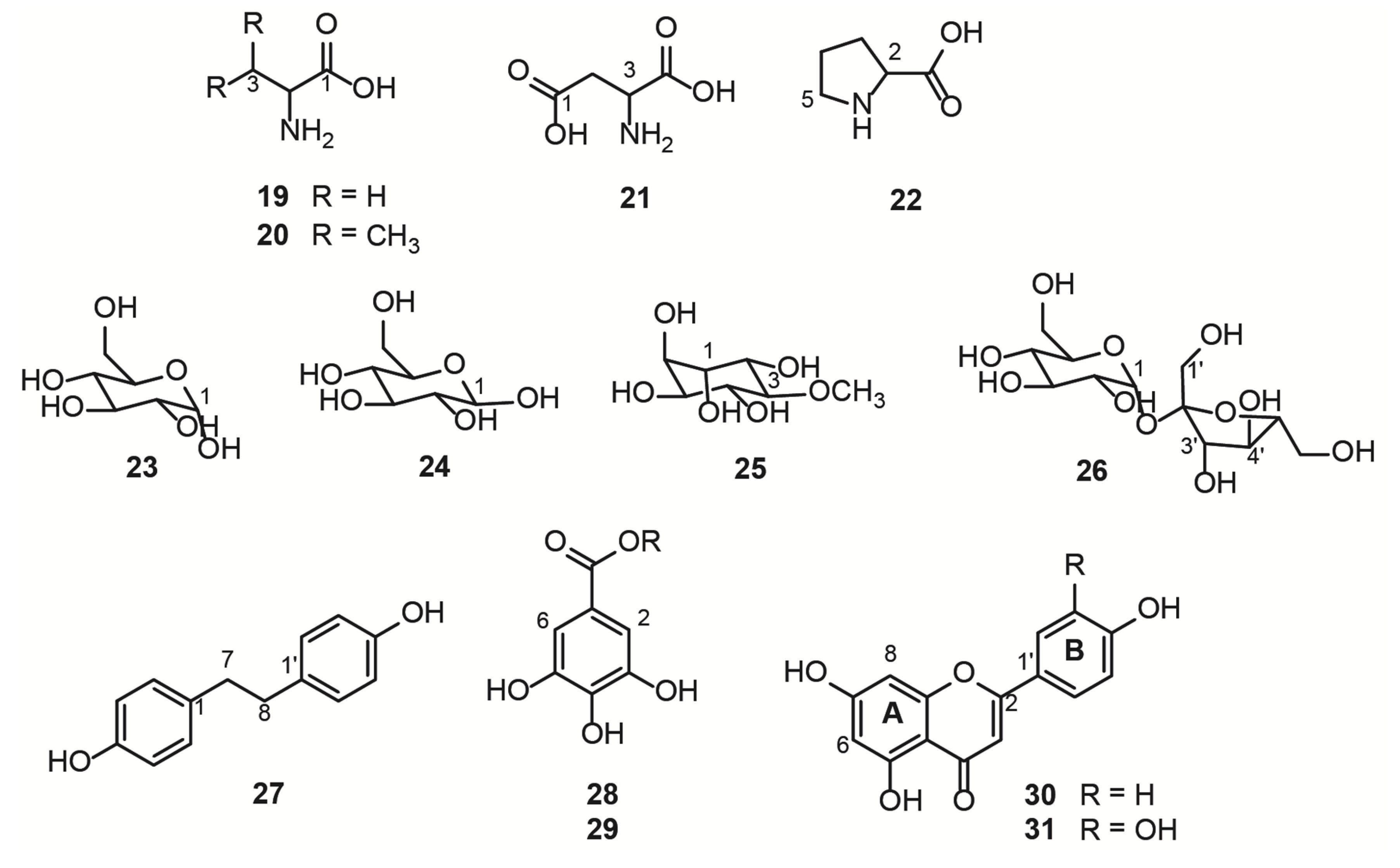{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Annotation (Correspondent Compound in Database *) | Molecular Formula (Neutral) | m/z (M + H)+ | Retention Time (min) | Chemical Class ** | Partial InChIKey |
|---|---|---|---|---|---|---|
| 1 | Quercetin-3-O-β-L-Rhamnofuranoside | C21H20O11 | 449.1055 | 1.61 | Flavonoids | OEKUVLQNKPXSOY |
| 2 | Quercetin-3-O-Arabinopyranoside | C20H18O11 | 435.0896 | 1.55 | Flavonoids | PZZRDJXEMZMZFD |
| 3 | [(3R,4R,5R,6S)-6-[2-(5,7-Dihydroxy-4-oxochromen-2-yl)-4,5-dihydroxyphenoxy]-4,5-dihydroxyoxan-3-yl] acetate | C22H20O12 | 477.1023 | 1.84 | Flavonoids | TUOJCDPSHAVURV |
| 4 | Quercetin 7-Rutinoside | C27H30O16 | 611.1589 | 1.38 | Flavonoids | IVTMALDHFAHOGL |
| 5 | Tagetiin | C21H20O13 | 481.0965 | 1.23 | Flavonoids | YUANNBKEZDNSIV |
| 6 | 3′,4′,5,7-Tetrahydroxyflavone-4′-O-[α-L-Rhamnopyranosyl-(1→6)-β-D-glucopyranoside] | C27H30O15 | 595.1625 | 1.47 | Flavonoids | WDQNUWOVEJHDOP |
| 7 | Mearnsitrin | C22H22O12 | 479.1158 | 1.65 | Flavonoids | NAQNISJXKDSYJD |
| 8 | Tricetin 3-Glucoside | C21H20O12 | 465.1012 | 1.38 | Flavonoids | XYILCYMQHZSECK |
| 9 | Tricetin | C15H10O7 | 303.0497 | 1.55 | Flavonoids | ARSRJFRKVXALTF |
| 10 | Kaempferol 3-O-α-L-rhamnofuranoside | C21H20O10 | 433.1127 | 1.81 | Flavonoids | FFFIPDPCGREKEW |
| 11 | Myricetin 3-O-α-Arabinofuranoside | C20H18O12 | 451.0856 | 1.35 | Flavonoids | OXJKSVCEIOYZQL |
| 12 | Nitensoside B | C28H32O16 | 625.1730 | 1.55 | Flavonoids | MTUPEWBIUKFRBD |
| 13 | Luteolin 3′-xyloside | C20H18O10 | 419.0966 | 1.74 | Flavonoids | ZUMPYZVELBOZDM |
| 14 | Corniculatusin 3-α-L-arabinofuranoside | C21H20O12 | 465.1004 | 1.62 | Flavonoids | DYNQYIRMFWJOJH |
| 15 | Isorhamnetin 3-O-α-L-arabinofuranoside | C21H20O11 | 449.1079 | 1.82 | Flavonoids | OOZLPFOTSYKMTJ |
| 16 | 2′,3′,5,5′,6′,7-Hexahydroxyisoflavone | C15H10O8 | 319.0437 | 1.38 | Isoflavones | XFCGHCBRDUZOSI |
| 17 | 2′-Hydroxygenistein | C15H10O6 | 287.0546 | 1.81 | Isoflavones | GSSOWCUOWLMMRJ |
| 18 | 4′,5,6,7,8-Pentahydroxyisoflavone-8-Me ether, 6-O-α-L-rhamnopyranoside | C22H22O11 | 463.1221 | 1.85 | Isoflavones | VEDUBLIYMIMISG |
| 31 | Quercetin | C15H10O7 | 303.0498 | 2.09 | Flavonoids | REFJWTPEDVJJIY |
| ID | Compound | Reference | Solvent |
|---|---|---|---|
| 19 | Alanine | [27] | D2O |
| 20 | Valine | [27] | D2O |
| 21 | Aspartate | [28] | D2O |
| 22 | Proline | [29] | D2O |
| 23 | α-D-glucose | [30,31] | CD3OD/D2O |
| 24 | β-D-glucose | [30,31] | CD3OD/D2O |
| 25 | D-pinitol | [32] | D2O |
| 26 | Sucrose | [30,31] | CD3OD/D2O |
| 27 | 4,4′-Dihydroxybibenzyl | [33] | acetone-d6 |
| 28 | Gallic acid derivative I | n.a. * | -- |
| 29 | Gallic acid derivative II | n.a. * | -- |
| 30 | Kaempferol | [34] | CD3OD |
| 31 | Quercetin | [34] | CD3OD |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borim de Souza, A.J.; Ocampos, F.M.M.; Catoia Pulgrossi, R.; Dokkedal, A.L.; Colnago, L.A.; Cechin, I.; Saldanha, L.L. NMR-Based Metabolomics Reveals Effects of Water Stress in the Primary and Specialized Metabolisms of Bauhinia ungulata L. (Fabaceae). Metabolites 2023, 13, 381. https://doi.org/10.3390/metabo13030381
Borim de Souza AJ, Ocampos FMM, Catoia Pulgrossi R, Dokkedal AL, Colnago LA, Cechin I, Saldanha LL. NMR-Based Metabolomics Reveals Effects of Water Stress in the Primary and Specialized Metabolisms of Bauhinia ungulata L. (Fabaceae). Metabolites. 2023; 13(3):381. https://doi.org/10.3390/metabo13030381
Chicago/Turabian StyleBorim de Souza, Ana Júlia, Fernanda Maria Marins Ocampos, Rafael Catoia Pulgrossi, Anne Lígia Dokkedal, Luiz Alberto Colnago, Inês Cechin, and Luiz Leonardo Saldanha. 2023. "NMR-Based Metabolomics Reveals Effects of Water Stress in the Primary and Specialized Metabolisms of Bauhinia ungulata L. (Fabaceae)" Metabolites 13, no. 3: 381. https://doi.org/10.3390/metabo13030381