
Ultraviolet-B and Heavy Metal-Induced Regulation of Secondary Metabolites in Medicinal Plants: A Review
Abstract
:1. Introduction
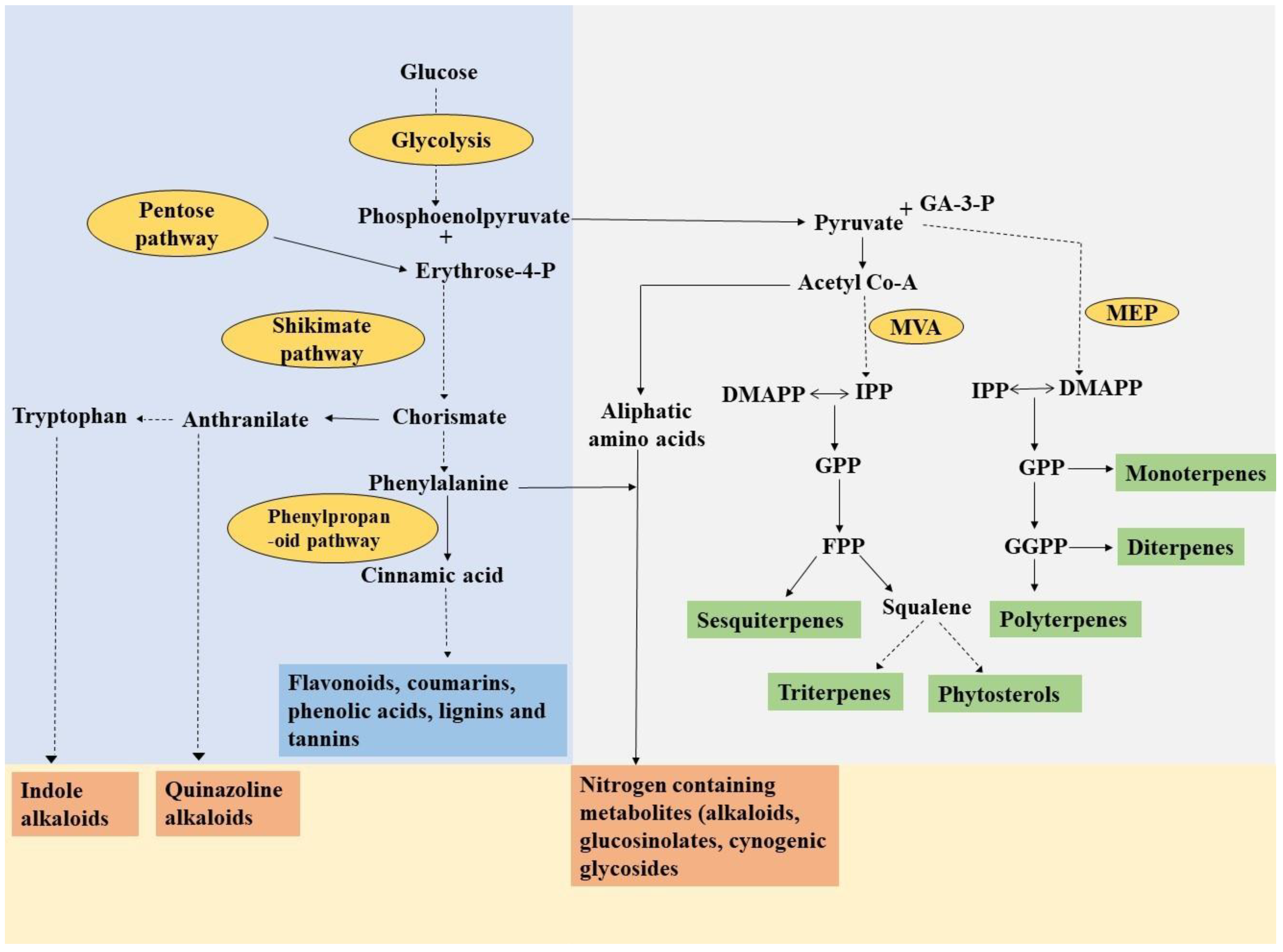
2. Secondary Metabolites: A Boon from Plants
3. UV-B Perception, Signal Transduction and Regulation of Genes of Secondary Metabolites’ Biosynthesis Pathway
4. UV-B Mediated Biosynthesis and Accumulation of Secondary Metabolites in Medicinal Plants
5. Role of Developmental Stages on Accumulation of Secondary Metabolites in Medicinal Plants under UV-B Stress
6. Impact of Heavy Metals on Signaling, Biosynthesis and Accumulation of Secondary Metabolites in Medicinal Plants
7. Interactive Effect of UV-B and Heavy Metals on Secondary Metabolites of Medicinal Plants
8. Conclusions and Future Prospects
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ncube, B.; Finnie, J.F.; Van Staden, J. Quality from the Field: The Impact of Environmental Factors as Quality Determinants in Medicinal Plants. S. Afr. J. Bot. 2012, 82, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Csepregi, K.; Coffey, A.; Cunningham, N.; Prinsen, E.; Hideg, É.; Jansen, M.A.K. Developmental Age and UV-B Exposure Co-Determine Antioxidant Capacity and Flavonol Accumulation in Arabidopsis Leaves. Environ. Exp. Bot. 2017, 140, 19–25. [Google Scholar] [CrossRef]
- Hideg, É.; Jansen, M.A.K.; Strid, Å. UV-B Exposure, ROS, and Stress: Inseparable Companions or Loosely Linked Associates? Trends Plant Sci. 2013, 18, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, A.; Jaiswal, D.; Agrawal, S.B. Ultraviolet-B Mediated Biochemical and Metabolic Responses of a Medicinal Plant Adhatoda vasica Nees. at Different Growth Stages. J. Photochem. Photobiol. B Biol. 2021, 216, 112142. [Google Scholar] [CrossRef]
- Kumar, D.; Seth, C.S. Photosynthesis, lipid peroxidation, and antioxidative responses of Helianthus annuus L. against chromium (VI) accumulation. Int. J. Phytoremed. 2022, 24, 590–599. [Google Scholar] [CrossRef]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Heavy Metal Stress and Some Mechanisms of Plant Defense Response Abolghassem. Sci. World J. 2015, 25, 27–54. [Google Scholar] [CrossRef] [Green Version]
- Kováčik, J.; Klejdus, B. Dynamics of Phenolic Acids and Lignin Accumulation in Metal-Treated Matricaria chamomilla Roots. Plant Cell Rep. 2008, 27, 605–615. [Google Scholar] [CrossRef]
- Anjitha, K.S.; Sameena, P.P.; Puthur, J.T. Functional Aspects of Plant Secondary Metabolites in Metal Stress Tolerance and Their Importance in Pharmacology. Plant Stress 2021, 2, 100038. [Google Scholar] [CrossRef]
- Yadav, M.; Gupta, P.; Seth, C.S. Foliar Application of α-Lipoic Acid Attenuates Cadmium Toxicity on Photosynthetic Pigments and Nitrogen Metabolism in Solanum lycopersicum L. Acta Physiol. Plant. 2022, 44, 112. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Kumar, R.; Altaf, M.M.; Kumar, A.; Khan, L.U.; Saqib, M.; Nawaz, M.A.; Saddiq, B.; Bahadur, S.; et al. Phytohormones Mediated Modulation of Abiotic Stress Tolerance and Potential Crosstalk in Horticultural Crops. J. Plant Growth Regul. 2022, 1–27. [Google Scholar] [CrossRef]
- Choudhary, K.K.; Agrawal, S.B. Cultivar Specificity of Tropical Mung Bean (Vigna radiata L.) to Elevated ultraviolet-B: Changes in Antioxidative Defense System, Nitrogen Metabolism and Accumulation of Jasmonic and Salicylic acids. Environ. Exp. Bot. 2014, 99, 122–132. [Google Scholar] [CrossRef]
- Paeizi, M.; Karimi, F.; Razavi, K. Changes in Medicinal Alkaloids Production and Expression of Related Regulatory and Biosynthetic Genes in Response to Silver Nitrate Combined with Methyl Jasmonate in Catharanthus roseus in Vitro Propagated Shoots. Plant Physiol. Biochem. 2018, 132, 623–632. [Google Scholar] [CrossRef]
- Fan, F.Y.; Li, C.L.; Luo, Z.; Tang, G.Z. UV-B Radiation-Induced Changes in Tea Metabolites and Related Gene Expression. In Stress Physiology of Tea in the Face of Climate Change; Han, W.Y., Li, X., Ahammed, G., Eds.; Springer: Singapore, 2018; pp. 111–133. [Google Scholar] [CrossRef]
- Mahajan, M.; Kuiry, R.; Pal, P.K. Understanding the Consequence of Environmental Stress for Accumulation of Secondary Metabolites in Medicinal and Aromatic Plants. J. Appl. Res. Med. Aromat. Plants 2020, 18, 100255. [Google Scholar] [CrossRef]
- Jan, R.; Asaf, S.; Numan, M.; Kim, K.M.; Lubna. Plant Secondary Metabolite Biosynthesis and Transcriptional Regulation in Response to Biotic and Abiotic Stress Conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as Antioxidants in Plants: Location and Functional Significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef]
- Facchini, P.J. Alkaloid Biosynthesis in Plants: Biochemistry, Cell Biology, Molecular Regulation, and Metabolic Engineering Applications. Annu. Rev. Plant Biol. 2001, 52, 29–66. [Google Scholar] [CrossRef] [Green Version]
- Zagoskina, N.V.; Dubravina, G.A.; Alyavina, A.K.; Goncharuk, E.A. Effect of Ultraviolet (UV-B) Radiation on the Formation and Localization of Phenolic Compounds in Tea Plant Callus Cultures. Russ. J. Plant Physiol. 2003, 50, 270–275. [Google Scholar] [CrossRef]
- Jaiswal, D.; Pandey-Rai, S.; Agrawal, S.B.; Apoorva. Untangling the UV-B Radiation-Induced Transcriptional Network Regulating Plant Morphogenesis and Secondary Metabolite Production. Environ. Exp. Bot. 2021, 192, 104655. [Google Scholar] [CrossRef]
- Pandey, A.; Agrawal, S.B. Ultraviolet-B Radiation: A Potent Regulator of Flavonoids Biosynthesis, Accumulation and Functions in Plants. Curr. Sci. 2020, 119, 176–185. [Google Scholar] [CrossRef]
- Jenkins, G.I. The UV-B Photoreceptor UVR8: From Structure to Physiology. Plant Cell 2014, 26, 21–37. [Google Scholar] [CrossRef] [Green Version]
- Tossi, V.E.; Regalado, J.J.; Iannicelli, J.; Laino, L.E.; Burrieza, H.P.; Escandón, A.S.; Pitta-Álvarez, S.I. Beyond Arabidopsis: Differential UV-B Response Mediated by UVR8 in Diverse Species. Front. Plant Sci. 2019, 10, 780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stracke, R.; Favory, J.J.; Gruber, H.; Bartelniewoehner, L.; Bartels, S.; Binkert, M.; Funk, M.; Weisshaar, B.; Ulm, R. The Arabidopsis BZIP Transcription Factor HY5 Regulates Expression of the PFG1/MYB12 Gene in Response to Light and Ultraviolet-B Radiation. Plant Cell Environ. 2010, 33, 88–103. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, X.; Jang, Z.; Chen, Z.; Ruo, X.; Jin, W.; Wu, Y.; Shi, X.; Xu, M. UV RESISTANCE LOCUS 8 from Chrysanthemum morifolium Ramat (CmUVR8) Plays Important Roles in UV-B Signal Transduction and UV-B-Induced Accumulation of Flavonoids. Front. Plant Sci. 2018, 9, 955. [Google Scholar] [CrossRef] [PubMed]
- Shamala, L.F.; Zhou, H.C.; Han, Z.X.; Wei, S. UV-B Induces Distinct Transcriptional Re-Programing in UVR8-Signal Transduction, Flavonoid, and Terpenoids Pathways in Camellia sinensis. Front. Plant Sci. 2020, 11, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afreen, F.; Zobayed, S.M.A.; Kozai, T. Spectral Quality and UV-B Stress Stimulate Glycyrrhizin Concentration of Glycyrrhiza uralensis in Hydroponic and Pot System. Plant Physiol. Biochem. 2005, 43, 1074–1081. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, Y.; Guo, Q.; Wang, W.; Liu, L.; Cao, L.; Li, C. Short-Term UV-B Radiation Effects on Morphology, Physiological Traits and Accumulation of Bioactive Compounds in Prunella vulgaris L. J. Plant Interact. 2017, 12, 348–354. [Google Scholar] [CrossRef]
- Rai, K.; Agrawal, S.B. Effect on Essential Oil Components and Wedelolactone Content of a Medicinal Plant Eclipta alba Due to Modifications in the Growth and Morphology under Different Exposures of Ultraviolet-B. Physiol. Mol. Biol. Plants 2020, 26, 773–792. [Google Scholar] [CrossRef]
- Kumari, R.; Agrawal, S.B.; Singh, S.; Dubey, N.K. Supplemental Ultraviolet-B Induced Changes in Essential Oil Composition and Total Phenolics of Acorus calamus L. (Sweet Flag). Ecotoxicol. Environ. Saf. 2009, 72, 2013–2019. [Google Scholar] [CrossRef]
- Takshak, S.; Agrawal, S.B. Secondary Metabolites and Phenylpropanoid Pathway Enzymes as Influenced under Supplemental Ultraviolet-B Radiation in Withania somnifera Dunal, an Indigenous Medicinal Plant. J. Photochem. Photobiol. B Biol. 2014, 140, 332–343. [Google Scholar] [CrossRef]
- Zhou, X.; Lyu, J.; Sun, L.; Dong, J.; Xu, H. Metabolic Programming of Rhododendron chrysanthum Leaves Following Exposure to UVB Irradiation. Funct. Plant Biol. 2021, 48, 1175–1185. [Google Scholar] [CrossRef]
- Zhang, X.; Ding, X.; Ji, Y.; Wang, S.; Chen, Y.; Luo, J.; Shen, Y.; Peng, L. Measurement of Metabolite Variations and Analysis of Related Gene Expression in Chinese Liquorice (Glycyrrhiza uralensis) Plants under UV-B Irradiation. Sci. Rep. 2018, 8, 6144. [Google Scholar] [CrossRef] [Green Version]
- Zhan, J.; Yang, Q.; Lin, Z.; Zheng, T.; Wang, M.; Sun, W.; Bu, T.; Tang, Z.; Li, C.; Han, X.; et al. Enhanced Antioxidant Capacity and Upregulated Transporter Genes Contribute to the UV-B-Induced Increase in Blinin in Conyza blinii. Environ. Sci. Pollut. Res. 2021, 28, 13275–13287. [Google Scholar] [CrossRef]
- Yoshida, H.; Shimada, K.; Hikosaka, S.; Goto, E. Effect of UV-B Irradiation on Bioactive Compounds of Red Perilla (Perilla frutescens (L.) Britton) Cultivated in a Plant Factory with Artificial Light. Horticulturae 2022, 8, 725. [Google Scholar] [CrossRef]
- Wu, K.; Liu, J.; Liu, Y.; Shao, W.; Abozeid, A.; Tang, Z.; Mu, L. Tissue-Specific Metabolic Response of Acanthopanax senticosus (Rupr.et. Maxim.) Harms Leaves to Enhanced UV-B Radiation. Acta Physiol. Plant. 2022, 44, 131. [Google Scholar] [CrossRef]
- Inostroza-Blancheteau, C.; Reyes-Díaz, M.; Arellano, A.; Latsague, M.; Acevedo, P.; Loyola, R.; Arce-Johnson, P.; Alberdi, M. Effects of UV-B Radiation on Anatomical Characteristics, Phenolic Compounds and Gene Expression of the Phenylpropanoid Pathway in Highbush Blueberry Leaves. Plant Physiol. Biochem. 2014, 85, 85–95. [Google Scholar] [CrossRef]
- Pandey, N.; Pandey-Rai, S. Short Term UV-B Radiation-Mediated Transcriptional Responses and Altered Secondary Metabolism of in Vitro Propagated Plantlets of Artemisia annua L. Plant Cell Tissue Organ Cult. 2014, 116, 371–385. [Google Scholar] [CrossRef]
- Lv, M.; Su, H.Y.; Li, M.L.; Yang, D.L.; Yao, R.Y.; Li, M.F.; Wei, J.H. Effect of UV-B Radiation on Growth, Flavonoid and Podophyllotoxin Accumulation, and Related Gene Expression in Sinopodophyllum hexandrum. Plant Biol. 2021, 23, 202–209. [Google Scholar] [CrossRef]
- Jiao, J.; Gai, Q.Y.; Wang, W.; Luo, M.; Gu, C.B.; Fu, Y.J.; Ma, W. Ultraviolet Radiation-Elicited Enhancement of Isoflavonoid Accumulation, Biosynthetic Gene Expression, and Antioxidant Activity in Astragalus membranaceus Hairy Root Cultures. J. Agric. Food Chem. 2015, 63, 8216–8224. [Google Scholar] [CrossRef]
- Li, J.; Han, X.; Wang, C.; Tang, L.; Zhang, W.; Qi, W. The Response of Achyranthes bidentata Blume to Short-Term UV-B Exposure. Russ. J. Plant Physiol. 2019, 66, 160–170. [Google Scholar] [CrossRef]
- Choi, J.; Kim, J.; Yoon, H.I.; Son, J.E. Effect of Far-Red and UV-B Light on the Growth and Ginsenoside Content of Ginseng (Panax ginseng C. A. Meyer) Sprouts Aeroponically Grown in Plant Factories. Hortic. Environ. Biotechnol. 2022, 63, 77–87. [Google Scholar] [CrossRef]
- Jaiswal, D.; Pandey, A.; Agrawal, M.; Agrawal, S.B. Photosynthetic, Biochemical and Secondary Metabolite Changes in a Medicinal Plant Chlorophytum borivillianum (Safed Musli) against Low and High Doses of UV-B Radiation. Photochem. Photobiol. 2022, 99, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Interdonato, R.; Rosa, M.; Nieva, C.B.; González, J.A.; Hilal, M.; Prado, F.E. Effects of Low UV-B Doses on the Accumulation of UV-B Absorbing Compounds and Total Phenolics and Carbohydrate Metabolism in the Peel of Harvested Lemons. Environ. Exp. Bot. 2011, 70, 204–211. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, J.; Abozeid, A.; Wu, K.X.; Guo, X.R.; Mu, L.Q.; Tang, Z.H. UV-B Radiation Largely Promoted the Transformation of Primary Metabolites to Phenols in Astragalus mongholicus Seedlings. Biomolecules 2020, 10, 504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidović, M.; Morina, F.; Milić, S.; Albert, A.; Zechmann, B.; Tosti, T.; Winkler, J.B.; Jovanović, S.V. Carbon Allocation from Source to Sink Leaf Tissue in Relation to Flavonoid Biosynthesis in Variegated Pelargonium zonale under UV-B Radiation and High PAR Intensity. Plant Physiol. Biochem. 2015, 93, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.S.; Zheng, L.P.; Tian, H.; Li, W.Y.; Wang, J.W. Transcriptome Responses Involved in Artemisinin Production in Artemisia annua L. under UV-B Radiation. J. Photochem. Photobiol. B Biol. 2014, 140, 292–300. [Google Scholar] [CrossRef]
- Dolzhenko, Y.; Bertea, C.M.; Occhipinti, A.; Bossi, S.; Maffei, M.E. UV-B Modulates the Interplay between Terpenoids and Flavonoids in Peppermint (Mentha × piperita L.). J. Photochem. Photobiol. B Biol. 2010, 100, 67–75. [Google Scholar] [CrossRef]
- Sankari, M.; Hridya, H.; Sneha, P.; George Priya Doss, C.; Ramamoorthy, S. Effect of UV Radiation and Its Implications on Carotenoid Pathway in Bixa orellana L. J. Photochem. Photobiol. B Biol. 2017, 176, 136–144. [Google Scholar] [CrossRef]
- Rodriguez-Morrison, V.; Llewellyn, D.; Zheng, Y. Cannabis Inflorescence Yield and Cannabinoid Concentration Are Not Increased With Exposure to Short-Wavelength Ultraviolet-B Radiation. Front. Plant Sci. 2021, 12, 725078. [Google Scholar] [CrossRef]
- Sun, M.; Gu, X.; Fu, H.; Zhang, L.; Chen, R.; Cui, L.; Zheng, L.; Zhang, D.; Tian, J. Change of Secondary Metabolites in Leaves of Ginkgo biloba L. in Response to UV-B Induction. Innov. Food Sci. Emerg. Technol. 2010, 11, 672–676. [Google Scholar] [CrossRef]
- Rai, K.; Agrawal, S.B. An Assessment of Dose-Dependent UV-B Sensitivity in Eclipta alba: Biochemical Traits, Antioxidative Properties, and Wedelolactone Yield. Environ. Sci. Pollut. Res. 2021, 28, 45434–45449. [Google Scholar] [CrossRef]
- Rizi, M.R.; Azizi, A.; Sayyari, M.; Mirzaie-Asl, A.; Conti, L. Increased Phenylpropanoids Production in UV-B Irradiated Salvia verticillata as a Consequence of Altered Genes Expression in Young Leaves. Plant Physiol. Biochem. 2021, 167, 174–184. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, X.; Guo, Q.; Liu, L.; Li, C.; Cao, L.; Qin, Q.; Zhao, M.; Wang, W. Effects of UV-B Radiation on the Content of Bioactive Components and the Antioxidant Activity of Prunella vulgaris L. Spica during Development. Molecules 2018, 23, 989. [Google Scholar] [CrossRef] [Green Version]
- Müller, V.; Albert, A.; Barbro Winkler, J.; Lankes, C.; Noga, G.; Hunsche, M. Ecologically Relevant UV-B Dose Combined with High PAR Intensity Distinctly Affect Plant Growth and Accumulation of Secondary Metabolites in Leaves of Centella asiatica L. Urban. J. Photochem. Photobiol. B Biol. 2013, 127, 161–169. [Google Scholar] [CrossRef]
- Lajayer, B.A.; Ghorbanpour, M.; Nikabadi, S. Heavy Metals in Contaminated Environment: Destiny of Secondary Metabolite Biosynthesis, Oxidative Status and Phytoextraction in Medicinal Plants. Ecotoxicol. Environ. Saf. 2017, 145, 377–390. [Google Scholar] [CrossRef]
- Nasim, S.A.; Dhir, B. Heavy Metals Alter the Potency of Medicinal Plants. Rev. Environ. Contam. Toxicol. 2010, 203, 139–149. [Google Scholar] [CrossRef]
- Kumar, S.; Trivedi, P.K. Heavy Metal Stress Signaling in Plants. In Plant Metal Interaction; Ahmad, P., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; Volume 2, pp. 585–603. [Google Scholar]
- Xing, B.; Yang, D.; Guo, W.; Liang, Z.; Yan, X.; Zhu, Y.; Liu, Y. Ag+ as a More Effective Elicitor for Production of Tanshinones than Phenolic Acids in Salvia miltiorrhiza Hairy Roots. Molecules 2015, 20, 309–324. [Google Scholar] [CrossRef] [Green Version]
- Pesaraklu, A.; Radjabian, T.; Salami, S.A. Methyl Jasmonate and Ag+ as Effective Elicitors for Enhancement of Phenolic Acids Contents in Salvia officinalis and Salvia verticillata, as Two Traditional Medicinal Plants. S. Afr. J. Bot. 2021, 141, 105–115. [Google Scholar] [CrossRef]
- Mishra, B.; Singh Sangwan, N. Amelioration of Cadmium Stress in Withania somnifera by ROS Management: Active Participation of Primary and Secondary Metabolism. Plant Growth Regul. 2019, 87, 403–412. [Google Scholar] [CrossRef]
- Rai, R.; Pandey, S.; Rai, S.P. Arsenic-Induced Changes in Morphological, Physiological, and Biochemical Attributes and Artemisinin Biosynthesis in Artemisia annua, an Antimalarial Plant. Ecotoxicology 2011, 20, 1900–1913. [Google Scholar] [CrossRef]
- Biswas, T.; Parveen, O.; Pandey, V.P.; Mathur, A.; Dwivedi, U.N. Heavy Metal Accumulation Efficiency, Growth and Centelloside Production in the Medicinal Herb Centella asiatica (L.) Urban under Different Soil Concentrations of Cadmium and Lead. Ind. Crops Prod. 2020, 157, 112948. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Kong, Y.C.; Zain, N.A.M. Effect of Cadmium and Copper Exposure on Growth, Secondary Metabolites and Antioxidant Activity in the Medicinal Plant Sambung Nyawa (Gynura procumbens (Lour.) Merr). Molecules 2017, 22, 1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, P.; Shi, Y.; Li, Z.; Chen, Q.; Xu, T.R.; Cui, X.; Guan, H.; Guo, L.; Yang, Y. Impaired Terpenoid Backbone Biosynthesis Reduces Saponin Accumulation in Panax notoginseng under Cd Stress. Funct. Plant Biol. 2019, 46, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Haq, K.; Rupasinghe, H.P.V.; Saxena, P.K. Nickel Contamination Affects Growth and Secondary Metabolite Composition of St. John’s Wort (Hypericum perforatum L.). Environ. Exp. Bot. 2003, 49, 251–257. [Google Scholar] [CrossRef]
- Babula, P.; Klejdus, B.; Kovacik, J.; Hedbavny, J.; Hlavna, M. Lanthanum Rather than Cadmium Induces Oxidative Stress and Metabolite Changes in Hypericum perforatum. J. Hazard. Mater. 2015, 286, 334–342. [Google Scholar] [CrossRef]
- Zeng, J.; Li, X.; Wang, X.; Zhang, K.; Wang, Y.; Kang, H.; Chen, G.; Lan, T.; Zhang, Z.; Yuan, S.; et al. Cadmium and Lead Mixtures Are Less Toxic to the Chinese Medicinal Plant Ligusticum chuanxiong Hort. Than Either Metal Alone. Ecotoxicol. Environ. Saf. 2020, 193, 110342. [Google Scholar] [CrossRef]
- Ahmad, B.; Jaleel, H.; Sadiq, Y.; Khan, M.M.A.; Shabbir, A. Response of Exogenous Salicylic Acid on Cadmium Induced Photosynthetic Damage, Antioxidant Metabolism and Essential Oil Production in Peppermint. Plant Growth Regul. 2018, 86, 273–286. [Google Scholar] [CrossRef]
- Prasad, A.; Singh, A.K.; Chand, S.; Chanotiya, C.S.; Patra, D.D. Effect of Chromium and Lead on Yield, Chemical Composition of Essential Oil, and Accumulation of Heavy Metals of Mint Species. Commun. Soil Sci. Plant Anal. 2010, 41, 2170–2186. [Google Scholar] [CrossRef]
- Hojati, M.; Modarres-Sanavy, S.A.M.; Enferadi, S.T.; Majdi, M.; Ghanati, F.; Farzadfar, S. Differential Deployment of Parthenolide and Phenylpropanoids in Feverfew Plants Subjected to Divalent Heavy Metals and Trans-Cinnamic Acid. Plant Soil 2016, 399, 41–59. [Google Scholar] [CrossRef]
- Park, Y.J.; Kim, N.S.; Sathasivam, R.; Chung, Y.S.; Park, S.U. Impact of Copper Treatment on Phenylpropanoid Biosynthesis in Adventitious Root Culture of Althaea officinalis L. Prep. Biochem. Biotechnol. 2022, 52, 283–291. [Google Scholar] [CrossRef]
- Rai, V.; Vajpayee, P.; Singh, S.N.; Mehrotra, S. Effect of Chromium Accumulation on Photosynthetic Pigments, Oxidative Stress Defense System, Nitrate Reduction, Proline Level and Eugenol Content of Ocimum tenuiflorum L. Plant Sci. 2004, 167, 1159–1169. [Google Scholar] [CrossRef]
- Gondor, O.K.; Szalai, G.; Kovács, V.; Janda, T.; Pál, M. Impact of UV-B on Drought- or Cadmium-Induced Changes in the Fatty Acid Composition of Membrane Lipid Fractions in Wheat. Ecotoxicol. Environ. Saf. 2014, 108, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Regier, N.; Beauvais-Flück, R.; Slaveykova, V.I.; Cosio, C. Elodea nuttallii Exposure to Mercury Exposure under Enhanced Ultraviolet Radiation: Effects on Bioaccumulation, Transcriptome, Pigment Content and Oxidative Stress. Aquat. Toxicol. 2016, 180, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.; Agrawal, M.; Agrawal, S.B. Individual and Combined Effects of Chromium and Ultraviolet-B Radiation on Defense System, Ultrastructural Changes, and Production of Secondary Metabolite Psoralen in a Medicinal Plant Psoralea corylifolia L. Environ. Sci. Pollut. Res. 2022, 30, 4372–4385. [Google Scholar] [CrossRef]
- Rojas-Lillo, Y.; Alberdi, M.; Acevedo, P.; Inostroza-Blancheteau, C.; Rengel, Z.; Mora, M.D.L.L.; Reyes-Díaz, M. Manganese Toxicity and UV-B Radiation Differentially Influence the Physiology and Biochemistry of Highbush Blueberry (Vaccinium corymbosum) Cultivars. Funct. Plant Biol. 2014, 41, 156–167. [Google Scholar] [CrossRef]
- Pandey, A.; Jaiswal, D.; Agrawal, M.; Agrawal, S.B. Changes in Ultrastructure, Photosynthetic Abilities, and Secondary Metabolite Due to Individual and Interactive Effects of Chromium and Ultraviolet-B Radiation in Adhatoda vasica. Photosynthetica 2022, 61, 20–30. [Google Scholar] [CrossRef]
- Azadi, M.; Siavash Moghaddam, S.; Rahimi, A.; Pourakbar, L.; Popović-Djordjević, J. Biosynthesized Silver Nanoparticles Ameliorate Yield, Leaf Photosynthetic Pigments, and Essential Oil Composition of Garden Thyme (Thymus vulgaris L.) Exposed to UV-B Stress. J. Environ. Chem. Eng. 2021, 9, 105919. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Medicinal Plants | Active Metabolites | Medicinal Properties/Uses | UV-B Dose and Duration | Responses | References |
|---|---|---|---|---|---|
| Glycyrrhiza uralensis L. | Glycyrrhizin | Natural sweetener, antitumor and anti-HIV activity | High intensity UV-B radiation (3 days at 1.13 W m−2). Low intensity UV-B radiation (15 days at 0.43 W m−2) | Both high and low intensity UV-B treatment increased glycyrrhizin content by 1.5 fold | [26] |
| Prunella vulgaris L. | Rosmarinic acid, caffeic acid and hyperoside | Thyroid gland malfunction, sedative and antifebrile agent | 35 μW cm−2 nm−1 30 min for 15 days | Caffeic acid, rosmarinic acid and hyperoside increased by 22.5, 53.4 and 121%, respectively | [27] |
| Eclipta alba L. (Hassk) | Wedelolactone | Liver cirrhosis, hepatitis and baldness | Intermittent and continuous exposure to UV-B (ambient + 7.2 kJ m−2 d−1) for 130 and 240 h, respectively | Wedelolactone content increased by 74.5 and 8.3%, respectively at intermittent and continuous exposure | [28] |
| Acorus calamus L. | β-asarone, aristolene and caryophyllene oxide | Antispasmodic, carminative, anthelminthic, hypotensive and anti-depressant properties | Ambient + 1.8 kJ m−2 d−1 for 3 h | β-asarone decreased by 8.4% whereas aristolene and caryophyllene oxide increased by 47.6 and 66.6%, respectively | [29] |
| Withania somnifera Dunal | Withanolide A and withaferin | Nervine tonic, antiarthritic, immune modulator and anticancerous activities | Ambient + 3.6 kJ m−2 d−1 for 3 h | Withanolide A decreased by 41.2% in leaves whereas withaferin A increased by 12.4% in above-ground parts | [30] |
| Rhododendron chrysanthum Pall. | Flavonoids | Inflammation, pain, skin ailments, common cold and gastrointestinal disorders | 2.3 W m−2 for 8 h | Flavonoids content increased by 62% | [31] |
| Glycyrrhiza uralensis L. | Apigenin, flavonoids | Anti-inflammatory, antiviral, antimicrobial, antioxidative and anticancer | 0.024 W m−2 for 12, 24, 48 and 96 h | Apigenin content increased by tenfold as compared to control with 12 h irradiation of UV-B | [32] |
| Conyza blinii H. Lev. | Blinin | Anti-inflammatory, analgesic, antitumor | 20, 40 and 80 μW cm−2 | Blinin content increased under all doses of UV-B | [33] |
| Perilla frutescens L | Rosmarinic acid and anthocyanins | Antibacterial, antioxidant | 4 W m−2 with continuous irradiation (4 W 24 h) and 6 W m−2 with 16 h irradiation (6 W 16 h) for 3 d | Rosmarinic acid contents in 4 W 24 h and 6 W 16 h treatments were 103 and 168% higher however, anthocyanin content decreased | [34] |
| Acanthopanax senticosus Rupr. and Maxim | Eleutheroside B and E (saponin) | Anti-diabetic, neuroprotective, anti-inflammatory, cardioprotective | 1.9 kJ m−2 d−1 (medium dose) and 3.4 kJ m−2 d−1 (high dose) for 2 h | Eleutheroside B and E content increased by 1.5 and 1.9 times under a medium dose and by 1.9 and 3.5 times at a higher dose | [35] |
| Vaccinium corymbosum L. cv. Brigitta and Bluegold | Delphinidin (anthocyanins) and chlorogenic acid (phenolic acid) | Antioxidant | 0.07, 0.12 or 0.19 W m−2 UV-B radiation for 0, 6, 24, 48 and 74 h. | Delphinidin was abundant in Bluegold and increased by 175% as compared to control at the highest dose whereas chlorogenic acid increased by 83% in Brigitta at a lower dose | [36] |
| Artemisia annua L. | Artemisinin (sesquiterpene lactone) | Antimalarial | 2.8 W m−2 of UV-B radiation for different short-term (1, 2, 3 and 4 h) | Artemisinin content increased by 103% after 3 h of UV-B irradiation | [37] |
| Sinopodophyllum hexandrum Royle | Podophyllotoxin | Anticancer | 107 μW cm−2 irradiance at 306 nm; | Podophyllotoxin showed 0.79- and 0.86-fold decreases under UV-B on a dry weight and per plant basis, respectively | [38] |
| Astragalus membranaceus L. | Isoflavonoids | Antioxidant, cardioprotective, anti-inflammatory, antiviral and immunomodulatory | 5.4–172.8 kJ m–2 d–1 for 0.5–16 h | Under optimal elicitation dose (86.4 kJ m–2 d–1), total isoflavonoids content increased by 2.29-fold against the control | [39] |
| Achyranthes bidentata Blume | Oleanolic acid and ecdysterone | Anti-inflammatory, anti-arthritic, diuretic and antibacterial agent | Duration (dose): 1 h (0.738 kJ m−2), 2 h (1.476 kJ m–2), 3 h (2.214 kJ m−2) and 4 h (2.952 kJ m−2) | The greatest increase in oleanolic acid was observed in the 3 h UV-B treatment with an increase of 201.53%, a peak increase of 255.54 and 347.96% total ecdysterone content in leaves and roots, respectively was observed after 2 h of UV-B treatment | [40] |
| Panax ginseng CA Meyer | Ginsenoside | Antidiabetic, anticancer, neuroprotective and immunomodulatory effect | 0.36, 0.72 and 1.08 kJ m−2 d−1 | Non- significant variation was observed in ginsenoside content under UV-B | [41] |
| Chlorophytum borivillianum Sant. et. Fer. | Saponin | Aphrodisiac, revitalizer, sex tonic and remedy for diabetes | Low (ambient + 3.2 kJ m−2 d−1) and high (ambient + 7.2 kJ m−2 d−1) UV-B dose | Under low dose of UV-B, content of saponin enhanced by 26% | [42] |
| Plants | Active Metabolites | Importance | Heavy Metal | Effect | References |
|---|---|---|---|---|---|
| Mentha piperita L. | Menthol, menthone and menthyl acetate | Essential oil is used as an antispasmodic, carminative, analgesic, antimicrobial, antiviral and vasodilating agent | 30, 60 and 120 mg kg−1 Cd in the soil | All the three levels of Cd enhanced the essential oil concentration; however, Cd-60 increased essential oil concentration maximally by up to 16.1%. Menthol concentration declined to 3.2% by Cd-120 treatment; however, Cd-30 and Cd-60 showed an increase of 8 and 1.03%, respectively. The content of menthone and menthyl acetate increased maximally at Cd-60 and Cd-120, respectively | [68] |
| Centella asiatica L. | Asiaticoside, madecassoside, asiatic acid and madecassic acid collectively known as centellosides | Nootropic disorders, memory loss, insomnia and dermatological issues | 50, 100, 150 and 200 mg kg−1 Cd in the soil | In terms of total centelloside content, maximum content was recovered from leaves from Cd-200 treatment. However, Cd-50 and Cd-100 treatments failed to elicit the production of centellosides | [62] |
| Tanacetum parthenium L. | Camphor | Essential oil exhibits anti-inflammatory, antioxidant, antifungal and insecticidal effects | 5, 35 and 70 μM Cd/Cu | Exposure to Cd at the level of 5, 35 and 70 μM decreased essential oil content by 21, 38 and 41%, respectively. However, essential oil content of plants exposed to 5 μM Cu increased by 6% compared to control plants. The relative content of camphor as the major component decreased by 15 and 12% under 70-μM-Cd and 5-μM-Cu treatments, respectively | [70] |
| Panax notoginseng (Burk.) F.H. Chen. | Panax notoginseng saponins (PNS) | Promotes blood circulation, counteracts blood stasis, relieves swelling and pain, | 2.5, 5.0 or 10 mM Cd | Cd significantly decreases the accumulation of PNS in the rhizome and main root while promoting the accumulation of PNS in the rootlet | [64] |
| Catharanthus roseus L. | Vincristine, vinblastine, ajmalicine, vindoline and catharanthine | Anticancer | 50 and 100 μM Ag | Vindoline, catharanthine, vincristine, vinblastine and ajmalicine contents increased significantly under all employed treatments after seven days of treatment | [12] |
| Althaea officinalis L. | Phenolics | Anti-inflammatory, antimicrobial, antitussive, demulcent and immunomodulatory | 0.5, 1 and 2 mM Cu | Treatment with 0.5 mM CuCl2 led to 1.67 times higher total phenolics | [71] |
| Salvia miltiorrhiza Bunge | Phenolic acid (rosmarinic acid, caffeic acid ferulic acid, salvianolic acid B, danshensu and cinnamic acid) and tanshinones | Cholestatic liver injury, anti-platelet, cardioprotective and anti- inflammatory effects | 15 μM Ag | Rosmarinic acid, caffeic acid and ferulic acid were significantly increased, while accumulations of salvianolic acid B, danshensu and cinnamic acid were decreased. Further, the content of tanshinones also increased | [58] |
| Artemisia annua L. | Artemisinin (sesquiterpene lactone) | Antimalarial | 1500, 3000 and 4500 μg L−1 As | The concentration of artemisinin increased in a dose- and time-dependent manner, the maximum being 22% at 4500 μg L−1 as of 7 days over the control | [61] |
| Ocimum tenuiflorum L. | Eugenol | Antiseptic, antispasmodic and antibacterial and insect repellent properties | 10.0, 20.0, 50.0 and 100.0 μM Cr | All the chromium concentrations increased eugenol content by14.46, 24.61, 16.80 and 3.83% under 10, 20, 50 and 100 μM Cr, respectively | [72] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pandey, A.; Agrawal, M.; Agrawal, S.B. Ultraviolet-B and Heavy Metal-Induced Regulation of Secondary Metabolites in Medicinal Plants: A Review. Metabolites 2023, 13, 341. https://doi.org/10.3390/metabo13030341
Pandey A, Agrawal M, Agrawal SB. Ultraviolet-B and Heavy Metal-Induced Regulation of Secondary Metabolites in Medicinal Plants: A Review. Metabolites. 2023; 13(3):341. https://doi.org/10.3390/metabo13030341
Chicago/Turabian StylePandey, Avantika, Madhoolika Agrawal, and Shashi Bhushan Agrawal. 2023. "Ultraviolet-B and Heavy Metal-Induced Regulation of Secondary Metabolites in Medicinal Plants: A Review" Metabolites 13, no. 3: 341. https://doi.org/10.3390/metabo13030341