
Metabolomics Assessment of Volume Overload-Induced Heart Failure and Oxidative Stress in the Kidney

 and
and 
Abstract
:
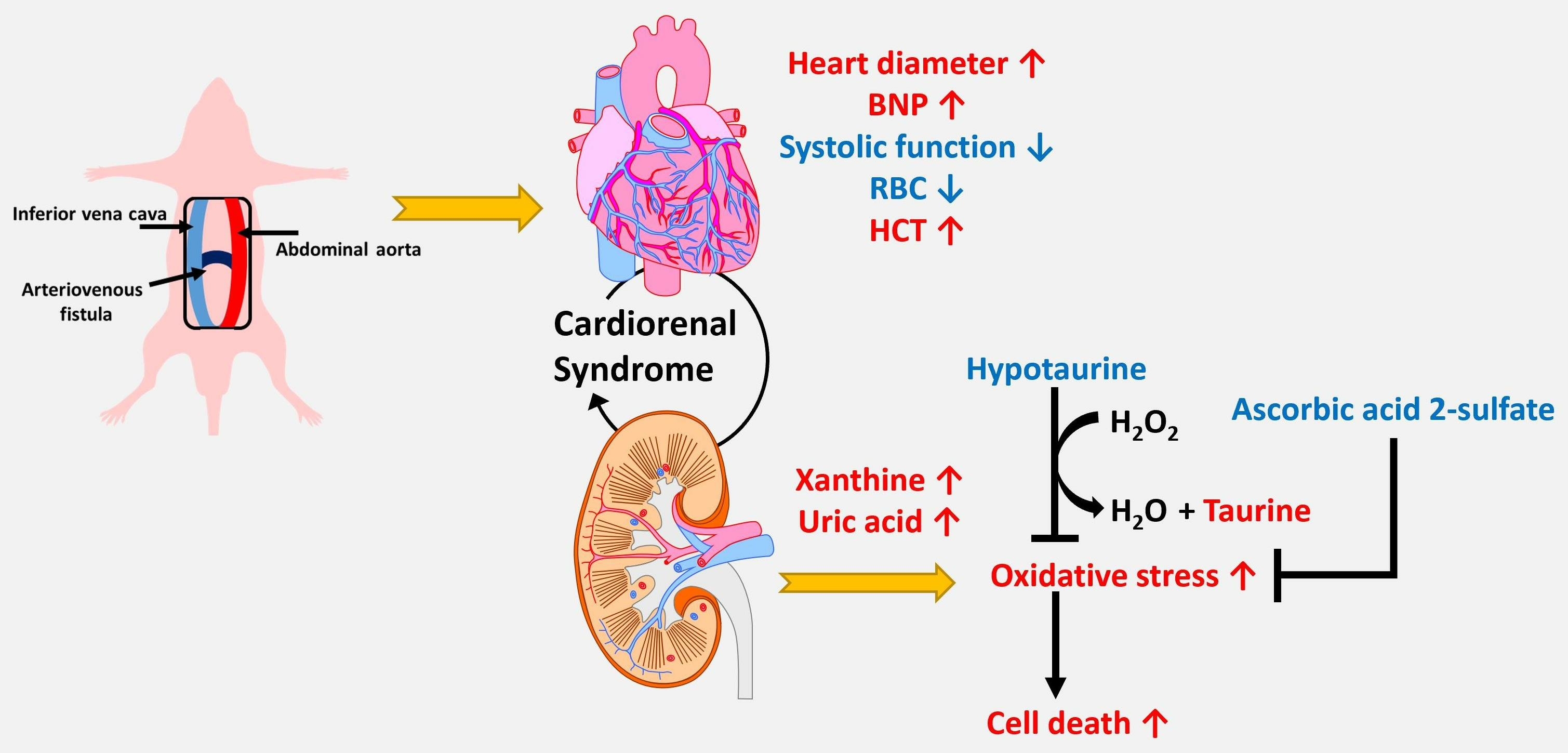
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Establishment of the Rat Aortocaval Fistula (ACF) Model
2.3. Biochemical Analysis
2.4. Echocardiographic Evaluation of Cardiac Function
2.5. Kidney Sample Preparation for Global Analysis of Hydrophilic Metabolites
2.6. Determination of the Hydrophilic Profile of Metabolites in Kidney Tissue Using UPLC TOF-MS
2.7. Cell Culture and Cell Viability Determination
2.8. Statistical Analysis
3. Results
3.1. Hemodynamic Characteristics and Cardiac Structure
3.2. Metabolomics Analysis of Kidney Metabolites in Control and ACF Rats
3.3. Accumulation of Xanthine and Uric Acid in Kidney Tissue Indicates a Higher Oxidative Stress Status in Kidney Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Disease, G.B.D.; Injury, I.; Prevalence, C. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar]
- Roger, V.L. Epidemiology of Heart Failure: A Contemporary Perspective. Circ. Res. 2021, 128, 1421–1434. [Google Scholar] [CrossRef] [PubMed]
- Groenewegen, A.; Rutten, F.H.; Mosterd, A.; Hoes, A.W. Epidemiology of heart failure. Eur. J. Heart Fail. 2020, 22, 1342–1356. [Google Scholar] [CrossRef] [PubMed]
- Savarese, G.; Becher, P.M.; Lund, L.H.; Seferovic, P.; Rosano, G.M.C.; Coats, A.J.S. Global burden of heart failure: A comprehensive and updated review of epidemiology. Cardiovasc. Res. 2023, 118, 3272–3287. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Pan, W.; Xu, Y.; Zhang, J.; Wan, J.; Jiang, H. Microglia-Mediated Neuroinflammation: A Potential Target for the Treatment of Cardiovascular Diseases. J. Inflamm. Res. 2022, 15, 3083–3094. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, C.J.; Duhamel, T.A.; Dhalla, N.S. Mechanisms for the transition from physiological to pathological cardiac hypertrophy. Can. J. Physiol. Pharmacol. 2020, 98, 74–84. [Google Scholar] [CrossRef]
- Liu, P.P. Cardiorenal syndrome in heart failure: A cardiologist’s perspective. Can. J. Cardiol. 2008, 24 (Suppl. B), 9B–25B. [Google Scholar] [CrossRef]
- Szymanski, M.K.; de Boer, R.A.; Navis, G.J.; van Gilst, W.H.; Hillege, H.L. Animal models of cardiorenal syndrome: A review. Heart Fail. Rev. 2012, 17, 411–420. [Google Scholar] [CrossRef]
- Damman, K.; Testani, J.M. The kidney in heart failure: An update. Eur. Heart J. 2015, 36, 1437–1444. [Google Scholar] [CrossRef]
- Hillege, H.L.; Nitsch, D.; Pfeffer, M.A.; Swedberg, K.; McMurray, J.J.; Yusuf, S.; Granger, C.B.; Michelson, E.L.; Östergren, J.; Cornel, J.H.; et al. Renal Function as a Predictor of Outcome in a Broad Spectrum of Patients with Heart Failure. Circulation 2006, 113, 671–678. [Google Scholar] [CrossRef]
- Goh, C.Y.; Vizzi, G.; De Cal, M.; Ronco, C. Cardiorenal syndrome: A complex series of combined heart/kidney disorders. Contrib. Nephrol. 2011, 174, 33–45. [Google Scholar] [PubMed]
- Kottgen, A.; Russell, S.D.; Loehr, L.R.; Crainiceanu, C.M.; Rosamond, W.D.; Chang, P.P.; Chambless, L.E.; Coresh, J. Reduced kidney function as a risk factor for incident heart failure: The atherosclerosis risk in communities (ARIC) study. J. Am. Soc. Nephrol. 2007, 18, 1307–1315. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.L.; Lichtman, J.H.; Bracken, M.B.; Shlipak, M.G.; Phillips, C.O.; DiCapua, P.; Krumholz, H.M. Renal impairment and outcomes in heart failure: Systematic review and meta-analysis. J. Am. Coll. Cardiol. 2006, 47, 1987–1996. [Google Scholar] [CrossRef]
- Chang, C.J.; Chen, C.C.; Hsu, L.A.; Chang, G.J.; Ko, Y.H.; Chen, C.F.; Chen, M.Y.; Yang, S.H.; Pang, J.H.S. Degradation of the internal elastic laminae in vein grafts of rats with aortocaval fistulae: Potential impact on graft vasculopathy. Am. J. Pathol. 2009, 174, 1837–1846. [Google Scholar] [CrossRef] [PubMed]
- Tung, Y.-C.; Cheng, M.-L.; Wu, L.-S.; Tang, H.-Y.; Huang, C.-Y.; Chang, G.-J.; Chang, C.-J. Derangements and Reversibility of Energy Metabolism in Failing Hearts Resulting from Volume Overload: Transcriptomics and Metabolomics Analyses. Int. J. Mol. Sci. 2022, 23, 6809. [Google Scholar] [CrossRef] [PubMed]
- Tung, Y.-C.; Wu, L.-S.; Hsiao, F.-C.; Hsu, L.-A.; Yeh, Y.-H.; Chang, C.-H.; Chen, Y.-C.; Chang, C.-J. Upregulation of Myocardial Neutrophil Gelatinase-Associated Lipocalin during Development of Heart Failure Caused by Volume-Overload and the Effect in Regulating Activity of Matrix Metalloproteinase-9. Acta Cardiol. Sin. 2022, 38, 765–777. [Google Scholar] [CrossRef]
- Wang, X.; Ren, B.; Liu, S.; Sentex, E.; Tappia, P.S.; Dhalla, N.S. Characterization of cardiac hypertrophy and heart failure due to volume overload in the rat. J. Appl. Physiol. 2003, 94, 752–763. [Google Scholar] [CrossRef]
- Griffin, J.L.; Atherton, H.; Shockcor, J.; Atzori, L. Metabolomics as a tool for cardiac research. Nat. Rev. Cardiol. 2011, 8, 630–643. [Google Scholar] [CrossRef]
- Lopaschuk, G.D.; Karwi, Q.G.; Tian, R.; Wende, A.R.; Abel, E.D. Cardiac Energy Metabolism in Heart Failure. Circ. Res. 2021, 128, 1487–1513. [Google Scholar] [CrossRef]
- Rosano, G.M.; Vitale, C. Metabolic Modulation of Cardiac Metabolism in Heart Failure. Card. Fail. Rev. 2018, 4, 99–103. [Google Scholar] [CrossRef]
- Cheng, M.L.; Wang, C.H.; Shiao, M.S.; Liu, M.H.; Huang, Y.Y.; Huang, C.Y.; Mao, C.T.; Lin, J.F.; Ho, H.Y.; Yang, N.I. Metabolic disturbances identified in plasma are associated with outcomes in patients with heart failure: Diagnostic and prognostic value of metabolomics. J. Am. Coll. Cardiol. 2015, 65, 1509–1520. [Google Scholar] [CrossRef]
- Tang, H.-Y.; Wang, C.-H.; Ho, H.-Y.; Wu, P.-T.; Hung, C.-L.; Huang, C.-Y.; Yeh, Y.-H.; Cheng, M.-L. Lipidomics reveals accumulation of the oxidized cholesterol in erythrocytes of heart failure patients. Redox Biol. 2017, 14, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.-Y.; Wang, C.-H.; Ho, H.-Y.; Lin, J.-F.; Lo, C.-J.; Huang, C.-Y.; Cheng, M.-L. Characteristic of Metabolic Status in Heart Failure and Its Impact in Outcome Perspective. Metabolites 2020, 10, 437. [Google Scholar] [CrossRef] [PubMed]
- Watson, L.E.; Sheth, M.; Denyer, R.F.; Dostal, D.E. Baseline echocardiographic values for adult male rats. J. Am. Soc. Echocardiogr. 2004, 17, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Beckonert, O.; Keun, H.C.; Ebbels, T.M.D.; Bundy, J.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Metabolic profiling, metabolomic and metabonomic procedures for NMR spectroscopy of urine, plasma, serum and tissue extracts. Nat. Protoc. 2007, 2, 2692–2703. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, D.; Yi, B.; Liao, Z.; Tang, L.; Yin, D.; He, M. Taurine supplementation reduces oxidative stress and protects the liver in an iron-overload murine model. Mol. Med. Rep. 2014, 10, 2255–2262. [Google Scholar] [CrossRef] [PubMed]
- Androne, A.S.; Katz, S.D.; Lund, L.; LaManca, J.; Hudaihed, A.; Hryniewicz, K.; Mancini, D.M. Hemodilution is common in patients with advanced heart failure. Circulation 2003, 107, 226–229. [Google Scholar] [CrossRef]
- Anand, I.; Gupta, P. Anemia and Iron Deficiency in Heart Failure: Current Concepts and Emerging Therapies. Circulation 2018, 138, 80–98. [Google Scholar] [CrossRef]
- Anand, I.S. Anemia and chronic heart failure implications and treatment options. J. Am. Coll. Cardiol. 2008, 52, 501–511. [Google Scholar] [CrossRef]
- Kousa, O.; Mullane, R.; Aboeata, A. Cardiorenal Syndrome. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Panjeta, M.; Tahirović, I.; Sofić, E.; Ćorić, J.; Dervišević, A. Interpretation of Erythropoietin and Haemoglobin Levels in Patients with Various Stages of Chronic Kidney Disease. J. Med. Biochem. 2017, 36, 145–152. [Google Scholar] [CrossRef]
- Buliga-Finis, O.N.; Ouatu, A.; Tanase, D.M.; Gosav, E.M.; Seritean Isac, P.N.; Richter, P.; Rezus, C. Managing Anemia: Point of Convergence for Heart Failure and Chronic Kidney Disease? Life 2023, 13, 1311. [Google Scholar] [CrossRef] [PubMed]
- Yancy, C.W.; Jessup, M.; Bozkurt, B.; Butler, J.; Casey, D.E., Jr.; Colvin, M.M.; Drazner, M.H.; Filippatos, G.S.; Fonarow, G.C.; Givertz, M.M.; et al. 2017 ACC/AHA/HFSA Focused Update of the 2013 ACCF/AHA Guideline for the Management of Heart Failure: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines and the Heart Failure Society of America. J. Am. Coll. Cardiol. 2017, 70, 776–803. [Google Scholar] [PubMed]
- Malik, A.; Brito, D.; Vaqar, S.; Chhabra, L. Congestive Heart Failure. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Cheng, T.-H.; Ma, M.-C.; Liao, M.-T.; Zheng, C.-M.; Lu, K.-C.; Liao, C.-H.; Hou, Y.-C.; Liu, W.-C.; Lu, C.-L. Indoxyl Sulfate, a Tubular Toxin, Contributes to the Development of Chronic Kidney Disease. Toxins 2020, 12, 684. [Google Scholar] [CrossRef] [PubMed]
- Harlacher, E.; Wollenhaupt, J.; Baaten, C.C.F.M.J.; Noels, H. Impact of Uremic Toxins on Endothelial Dysfunction in Chronic Kidney Disease: A Systematic Review. Int. J. Mol. Sci. 2022, 23, 531. [Google Scholar] [CrossRef] [PubMed]
- Eynde, J.V.D.; Jacquemyn, X.; Cloet, N.; Noé, D.; Gillijns, H.; Lox, M.; Gsell, W.; Himmelreich, U.; Luttun, A.; McCutcheon, K.; et al. Arteriovenous Fistulae in Chronic Kidney Disease and the Heart: Physiological, Histological, and Transcriptomic Characterization of a Novel Rat Model. J. Am. Heart Assoc. 2022, 11, e027593. [Google Scholar] [CrossRef]
- Yamamoto, T.; Moriwaki, Y.; Takahashi, S. Effect of ethanol on metabolism of purine bases (hypoxanthine, xanthine, and uric acid). Clin. Chim. Acta 2005, 356, 35–57. [Google Scholar] [CrossRef]
- Gotsman, I.; Keren, A.; Lotan, C.; Zwas, D.R. Changes in uric acid levels and allopurinol use in chronic heart failure: Association with improved survival. J. Card. Fail. 2012, 18, 694–701. [Google Scholar] [CrossRef]
- Omizo, H.; Tamura, Y.; Morimoto, C.; Ueno, M.; Hayama, Y.; Kuribayashi-Okuma, E.; Uchida, S.; Shibata, S. Cardio-renal protective effect of the xanthine oxidase inhibitor febuxostat in the 5/6 nephrectomy model with hyperuricemia. Sci. Rep. 2020, 10, 9326. [Google Scholar] [CrossRef]
- Wang, T.T.; Du, Y.W.; Wang, W.; Li, X.N.; Liu, H.B. Inhibition of Xanthine Oxidase Protects against Sepsis-Induced Acute Kidney Injury by Ameliorating Renal Hypoxia. Oxid. Med. Cell. Longev. 2022, 2022, 4326695. [Google Scholar] [CrossRef]
- Jalal, D.I.; Chonchol, M.; Chen, W.; Targher, G. Uric acid as a target of therapy in CKD. Am. J. Kidney Dis. 2013, 61, 134–146. [Google Scholar] [CrossRef]
- Kanbay, M.; Segal, M.; Afsar, B.; Kang, D.-H.; Rodriguez-Iturbe, B.; Johnson, R.J. The role of uric acid in the pathogenesis of human cardiovascular disease. Heart 2013, 99, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-H.; Wei, F.; Vaziri, N.D.; Cheng, X.-L.; Bai, X.; Lin, R.-C.; Zhao, Y.-Y. Metabolomics insights into chronic kidney disease and modulatory effect of rhubarb against tubulointerstitial fibrosis. Sci. Rep. 2015, 5, 14472. [Google Scholar] [CrossRef] [PubMed]
- Sturman, J.A. Taurine in development. Physiol. Rev. 1993, 73, 119–147. [Google Scholar] [CrossRef]
- Cheropkina, H.; Catucci, G.; Marucco, A.; Fenoglio, I.; Gilardi, G.; Sadeghi, S.J. Human flavin-containing monooxygenase 1 and its long-sought hydroperoxyflavin intermediate. Biochem. Pharmacol. 2021, 193, 114763. [Google Scholar] [CrossRef]
- McGaunn, J.; Baur, J.A. Taurine linked with healthy aging. Science 2023, 380, 1010–1011. [Google Scholar] [CrossRef] [PubMed]
- Chesney, R.W.; Han, X.; Patters, A.B. Taurine and the renal system. J. Biomed. Sci. 2010, 17 (Suppl. S1), S4. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Yum, H.W.; Kim, S.H.; Kim, W.; Kim, S.J.; Kim, C.; Kim, K.; Suh, Y.G.; Surh, Y.J. Protective Effects of Taurine Chloramine on Experimentally Induced Colitis: NFkappaB, STAT3, and Nrf2 as Potential Targets. Antioxidants 2021, 10, 479. [Google Scholar] [CrossRef]
- Trachtman, H.; Futterweit, S.; Prenner, J.; Hanon, S. Antioxidants reverse the antiproliferative effect of high glucose and advanced glycosylation end products in cultured rat mesangial cells. Biochem. Biophys. Res. Commun. 1994, 199, 346–352. [Google Scholar] [CrossRef]
- Han, X.; Yue, J.; Chesney, R.W. Functional TauT protects against acute kidney injury. J. Am. Soc. Nephrol. 2009, 20, 1323–1332. [Google Scholar] [CrossRef]
- Takebayashi, J.; Kaji, H.; Ichiyama, K.; Makino, K.; Gohda, E.; Yamamoto, I.; Tai, A. Inhibition of free radical-induced erythrocyte hemolysis by 2-O-substituted ascorbic acid derivatives. Free Radic. Biol. Med. 2007, 43, 1156–1164. [Google Scholar] [CrossRef]
- Yin, X.; Li, J.; Bose, D.; Okamoto, J.; Kwon, A.; Jackson, A.U.; Silva, L.F.; Oravilahti, A.; Stringham, H.M.; Ripatti, S.; et al. Metabolome-wide Mendelian randomization characterizes heterogeneous and shared causal effects of metabolites on human health. medRxiv 2023. [Google Scholar] [CrossRef]
- Hansen, S.H.; Andersen, M.L.; Cornett, C.; Gradinaru, R.; Grunnet, N. A role for taurine in mitochondrial function. J. Biomed. Sci. 2010, 17 (Suppl. S1), S23. [Google Scholar] [CrossRef] [PubMed]
- Jong, C.J.; Sandal, P.; Schaffer, S.W. The Role of Taurine in Mitochondria Health: More Than Just an Antioxidant. Molecules 2021, 26, 4913. [Google Scholar] [CrossRef]
- De Carvalho, F.G.; Brandao, C.F.C.; Batitucci, G.; Souza, A.d.O.; Ferrari, G.D.; Alberici, L.C.; Muñoz, V.R.; Pauli, J.R.; De Moura, L.P.; Ropelle, E.R.; et al. Taurine supplementation associated with exercise increases mitochondrial activity and fatty acid oxidation gene expression in the subcutaneous white adipose tissue of obese women. Clin. Nutr. 2021, 40, 2180–2187. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (n = 7) | ACF (n = 6) | p-Value | |
|---|---|---|---|
| Weight (g) | 514.29 ± 52.16 | 579.17 ± 73.65 | 0.133 |
| Hematologic data | |||
| WBC (103/μL) | 14.85 ± 5.04 | 16.15 ± 5.06 | 0.366 |
| RBC (106/μL) | 10.59 ± 0.55 | 9.18 ± 1.41 | 0.073 |
| HGB (g/dL) | 16.91 ± 0.62 | 14.65 ± 1.97 * | 0.017 |
| HCT (%) | 52.51 ± 1.33 | 46.10 ± 4.93 ** | 0.008 |
| MCV (fL) | 49.73 ± 2.97 | 50.62 ± 4.02 | 0.628 |
| MCH (pg/cell) | 16.00 ± 0.78 | 16.00 ± 0.73 | 0.943 |
| MCHC (g/dL) | 32.23 ± 1.38 | 31.70 ± 1.33 | 0.471 |
| RDW (fL) | 29.36 ± 0.60 | 30.57 ± 3.33 | 0.836 |
| RDW-CV (%) | 20.79 ± 1.47 | 20.17 ± 1.43 | 0.668 |
| PLT (103/μL) | 959.86 ± 215.68 | 741.33 ± 221.89 | 0.100 |
| PCT (%) | 0.71 ± 0.18 | 0.54 ± 0.15 | 0.101 |
| PDW (fL) | 8.16 ± 0.24 | 8.03 ± 0.68 | 0.564 |
| MPV (fL) | 7.31 ± 0.34 | 7.30 ± 0.38 | 0.942 |
| Biochemistry data | |||
| Glucose (mg/dL) | 87.86 ± 9.35 | 89.67 ± 15.37 | 1.000 |
| Creatinine (mg/dL) | 1.09 ± 0.31 | 1.13 ± 0.14 | 0.775 |
| BNP (ng/mL) | 0.49 ± 0.28 | 1.10 ± 0.62 * | 0.036 |
| ALT (U/L) | 16.68 ± 6.16 | 17.88 ± 4.30 | 0.945 |
| AST (U/L) | 54.11 ± 12.75 | 76.55 ± 19.29 * | 0.022 |
| Phospholipid (mg/dL) | 93.52 ± 8.86 | 97.73 ± 8.90 | 0.445 |
| Cholesterol (mg/dL) | 167.20 ± 10.84 | 167.06 ± 15.79 | 0.945 |
| Triglyceride (mg/dL) | 70.68 ± 16.78 | 77.33 ± 16.00 | 0.534 |
| No. | Metabolites | Control | ACF | VIP | p Value |
|---|---|---|---|---|---|
| 1 | Carnitine | 517.03 ± 45.87 | 606.07 ± 101.21 | 4.59 | 1.15 × 10−4 |
| 2 | Acetylcarnitine | 72.99 ± 13.65 | 28.75 ± 20.74 | 4.04 | 2.39 × 10−12 |
| 3 | Phosphocholine | 283.01 ± 28.89 | 240.81 ± 38.19 | 3.25 | 3.68 × 10−5 |
| 4 | Adenosine | 34.62 ± 16.26 | 11.34 ± 4.55 | 2.74 | 1.31 × 10−8 |
| 5 | Betaine | 143.77 ± 13.45 | 178.44 ± 61.52 | 2.48 | 5.43 × 10−3 |
| 6 | Taurine | 209.68 ± 17.40 | 236.60 ± 31.51 | 2.46 | 3.00 × 10−4 |
| 7 | NAD | 113.83 ± 30.33 | 87.52 ± 16.21 | 2.41 | 3.87 × 10−4 |
| 8 | Creatine | 71.38 ± 18.67 | 48.59 ± 25.99 | 2.21 | 5.94 × 10−4 |
| 9 | Guanine | 57.76 ± 13.18 | 40.64 ± 12.72 | 2.11 | 1.79 × 10−5 |
| 10 | Propionylcarnitine | 15.17 ± 6.40 | 3.17 ± 3.77 | 2.05 | 1.36 × 10−1 |
| 11 | Choline | 78.12 ± 35.95 | 50.35 ± 46.63 | 2.05 | 1.91 × 10−2 |
| 12 | Hydroxyisovaleroyl carnitine | 31.17 ± 5.16 | 20.01 ± 9.03 | 1.79 | 1.02 × 10−6 |
| 13 | Anserine | 23.66 ± 7.91 | 12.19 ± 8.25 | 1.77 | 5.12 × 10−6 |
| 14 | Deoxycytidine | 6.84 ± 3.09 | 0.62 ± 1.16 | 1.52 | 1.69 × 10−12 |
| 15 | N-Methyl histidine | 26.77 ± 3.12 | 19.86 ± 6.61 | 1.36 | 9.22 × 10−6 |
| 16 | N-Methyl aspartic acid | 11.27 ± 5.95 | 4.98 ± 3.70 | 1.25 | 4.24 × 10−5 |
| 17 | Tryptophan | 21.16 ± 6.38 | 28.66 ± 8.91 | 1.25 | 9.05 × 10−4 |
| 18 | Cytosine | 44.11 ± 7.58 | 52.09 ± 11.49 | 1.20 | 4.29 × 10−3 |
| 19 | 5′-Methylthioadenosine | 11.03 ± 1.37 | 7.70 ± 1.57 | 1.08 | 9.21 × 10−11 |
| 20 | Inosine | 949.86 ± 93.69 | 1021.77 ± 95.09 | 3.60 | 8.49 × 10−3 |
| 21 | AMP | 191.35 ± 97.41 | 127.16 ± 78.60 | 3.32 | 1.27 × 10−2 |
| 22 | Pyroglutamic acid | 127.33 ± 23.56 | 100.05 ± 21.89 | 2.66 | 7.91 × 10−5 |
| 23 | Xanthine | 93.10 ± 24.54 | 128.83 ± 59.43 | 2.61 | 5.41 × 10−3 |
| 24 | Glucuronolactone | 95.91 ± 46.76 | 61.38 ± 40.11 | 2.53 | 6.65 × 10−3 |
| 25 | Tyrosine | 20.93 ± 4.00 | 34.82 ± 9.55 | 2.21 | 5.60 × 10−9 |
| 26 | Succinyladenosine | 49.98 ± 17.85 | 33.01 ± 10.59 | 2.06 | 1.62 × 10−4 |
| 27 | Malic acid | 56.50 ± 9.38 | 72.07 ± 15.28 | 2.04 | 4.11 × 10−5 |
| 28 | Gluconic acid | 32.49 ± 11.13 | 20.62 ± 9.06 | 1.74 | 1.20 × 10−4 |
| 29 | Glutamate | 106.18 ± 12.36 | 119.18 ± 17.13 | 1.63 | 2.60 × 10−3 |
| 30 | Uric acid | 24.67 ± 3.15 | 34.37 ± 11.54 | 1.58 | 8.60 × 10−5 |
| 31 | Glucose 6-phosphate | 17.48 ± 4.50 | 10.55 ± 5.18 | 1.43 | 4.16 × 10−6 |
| 32 | Ascorbic acid-2-sulfate | 27.11 ± 6.49 | 18.16 ± 11.54 | 1.41 | 9.53 × 10−4 |
| 33 | Cholic acid | 6.32 ± 7.45 | 14.00 ± 8.03 | 1.32 | 7.85 × 10−4 |
| 34 | Glucose | 9.75 ± 1.83 | 4.97 ± 2.89 | 1.31 | 2.63 × 10−9 |
| 35 | S-Adenosylhomocysteine | 6.89 ± 2.24 | 11.31 ± 3.42 | 1.17 | 9.50 × 10−7 |
| 36 | 3,4-Dihydroxybutyric acid | 2.14 ± 2.94 | 6.39 ± 2.33 | 1.15 | 6.30 × 10−7 |
| 37 | 5-Glutamyl-taurine | 38.93 ± 4.00 | 32.95 ± 8.58 | 1.12 | 1.80 × 10−3 |
| 38 | Phosphoethanolamine | 25.59 ± 7.16 | 31.09 ± 3.73 | 1.09 | 1.39 × 10−3 |
| 39 | N-Acetylglutamine | 4.89 ± 2.20 | 1.71 ± 1.40 | 1.02 | 1.39 × 10−7 |
| 40 | Tyrosol 4-sulfate | 7.35 ± 3.96 | 3.37 ± 2.61 | 1.01 | 1.13 × 10−4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, H.-Y.; Huang, J.-E.; Tsau, M.-T.; Chang, C.-J.; Tung, Y.-C.; Lin, G.; Cheng, M.-L. Metabolomics Assessment of Volume Overload-Induced Heart Failure and Oxidative Stress in the Kidney. Metabolites 2023, 13, 1165. https://doi.org/10.3390/metabo13111165
Tang H-Y, Huang J-E, Tsau M-T, Chang C-J, Tung Y-C, Lin G, Cheng M-L. Metabolomics Assessment of Volume Overload-Induced Heart Failure and Oxidative Stress in the Kidney. Metabolites. 2023; 13(11):1165. https://doi.org/10.3390/metabo13111165
Chicago/Turabian StyleTang, Hsiang-Yu, Jyh-En Huang, Ming-Tong Tsau, Chi-Jen Chang, Ying-Chang Tung, Gigin Lin, and Mei-Ling Cheng. 2023. "Metabolomics Assessment of Volume Overload-Induced Heart Failure and Oxidative Stress in the Kidney" Metabolites 13, no. 11: 1165. https://doi.org/10.3390/metabo13111165