
Metabolomic Insights into the Mechanisms of Ganoderic Acid: Protection against α-Amanitin-Induced Liver Injury


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animal Treatment
2.3. Sample Collection and Preparation
2.4. Oxidative Damage and Inflammatory Markers
2.5. UPLC/Q-TOF-MS Analysis
2.6. Data Processing and Statistical Analysis
2.7. Identification of Potential Biomarkers
3. Results
3.1. Blood Biochemical Indicators
3.2. Histopathological Findings for Liver Tissue
3.3. ROS, IL-6, TNF-α, and COX-2 Levels
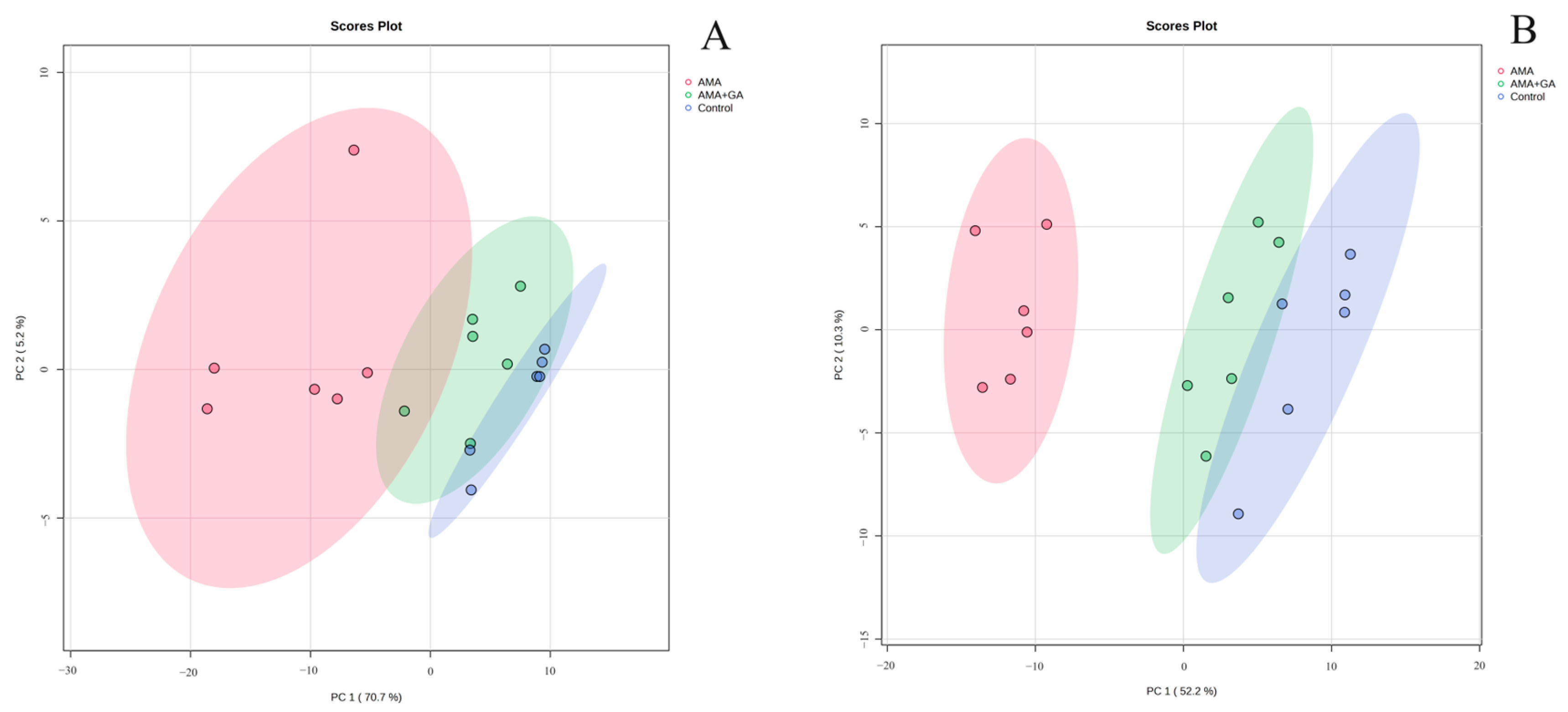
3.4. Multivariate Analysis of Analytes Present in the Blood
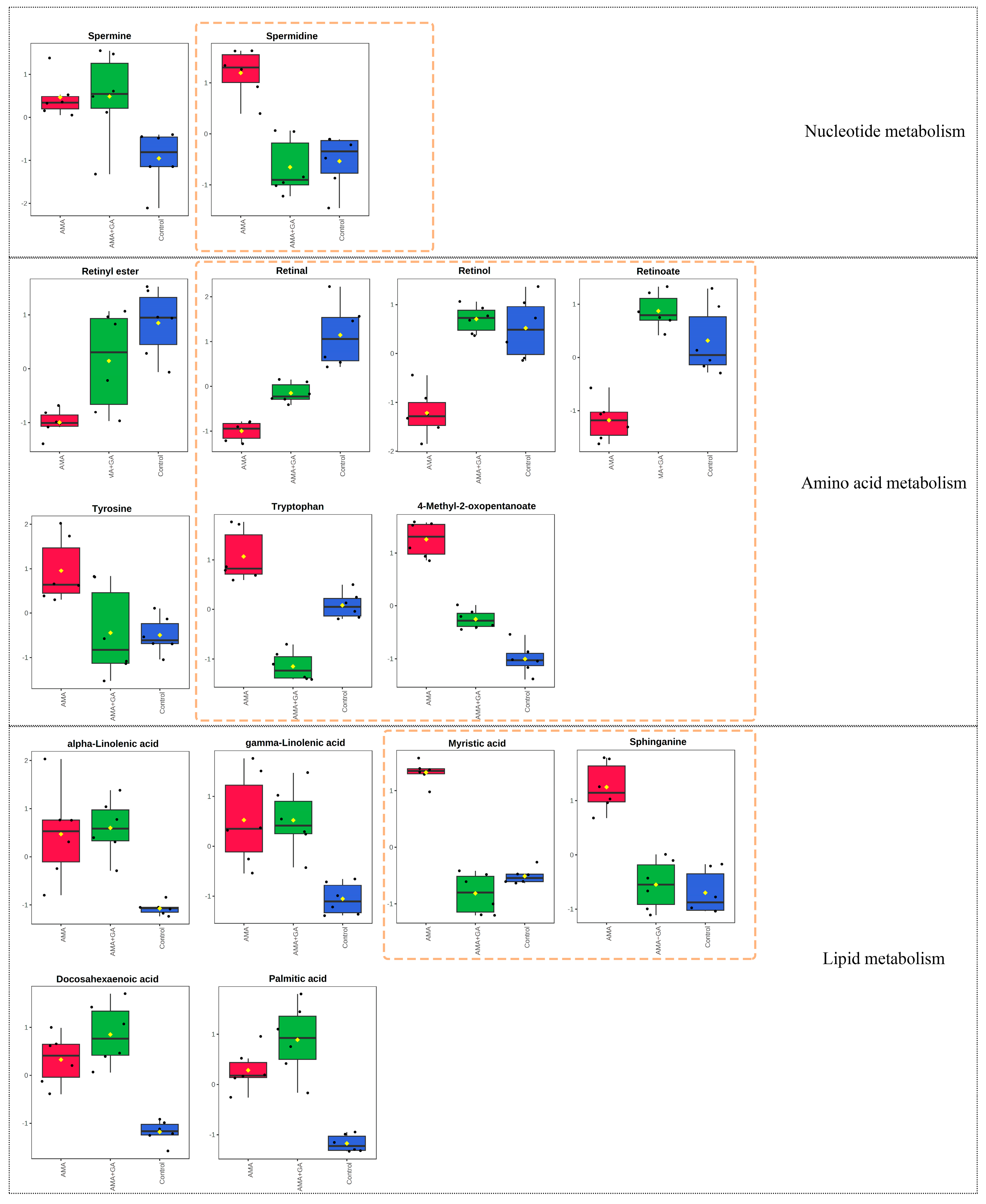
3.5. Metabolites’ Identification and Pathway Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ALP | Alkaline Phosphatase |
| ALT | Alanine Aminotransferase |
| AMA | α-Amanitin |
| AST | Aspartate Aminotransferase |
| BUN | Blood Urea Nitrogen |
| COX-2 | Cyclooxygenase-2 |
| Cr | Creatinine |
| GA | Ganoderic Acid A |
| GGT | Gamma-Glutamyl Transpeptidase |
| IL-6 | Interleukin-6 |
| OPLS-DA | Orthogonal Partial Least Squares Discriminant Analysis |
| PBS | Phosphate Buffered Saline |
| PCA | Principal Component Analysis |
| PLS-DA | Partial Least Squares Discriminant Analysis |
| QC | Quality Control Samples |
| ROS | Reactive Oxygen Species |
| RSD | Relative Standard Deviation |
| TNF-α | Tumor Necrosis Factor-α |
| VIP | Variable Importance in Projection |
References
- Li, C.; Oberlies, N.H. The most widely recognized mushroom: Chemistry of the genus Amanita. Life Sci. 2005, 78, 532–538. [Google Scholar] [CrossRef]
- Liu, X.; Farnung, L.; Wigge, C.; Cramer, P. Cryo-EM structure of a mammalian RNA polymerase II elongation complex inhibited by α-amanitin. J. Biol. Chem. 2018, 293, 7189–7194. [Google Scholar] [CrossRef]
- Rodrigues, D.F.; das Neves, R.P.; Carvalho, A.T.P.; Bastos, M.L.; Costa, V.M.; Carvalho, F. In vitro mechanistic studies on α-amanitin and its putative antidotes. Arch. Toxicol. 2020, 94, 2061–2078. [Google Scholar] [CrossRef]
- Le Daré, B.; Ferron, P.-J.; Gicquel, T. Toxic Effects of Amanitins: Repurposing Toxicities toward New Therapeutics. Toxins 2021, 13, 417. [Google Scholar] [CrossRef]
- Xue, J.; Lou, X.; Ning, D.; Shao, R.; Chen, G. Mechanism and treatment of α-amanitin poisoning. Arch. Toxicol. 2022, 97, 121–131. [Google Scholar] [CrossRef]
- Lin, L.-Y.; Tong, Y.-L.; Lu, Y.-Q. The characteristics of liver injury induced by Amanita and clinical value of α-amanitin detection. Hepatobiliary Pancreat. Dis. Int. 2022, 21, 257–266. [Google Scholar] [CrossRef]
- Chen, X.; Shao, B.; Yu, C.; Yao, Q.; Ma, P.; Li, H.; Cai, W.; Fu, H.; Li, B.; Sun, C. The cyclopeptide -amatoxin induced hepatic injury via the mitochondrial apoptotic pathway associated with oxidative stress. Peptides 2020, 129, 170314. [Google Scholar] [CrossRef]
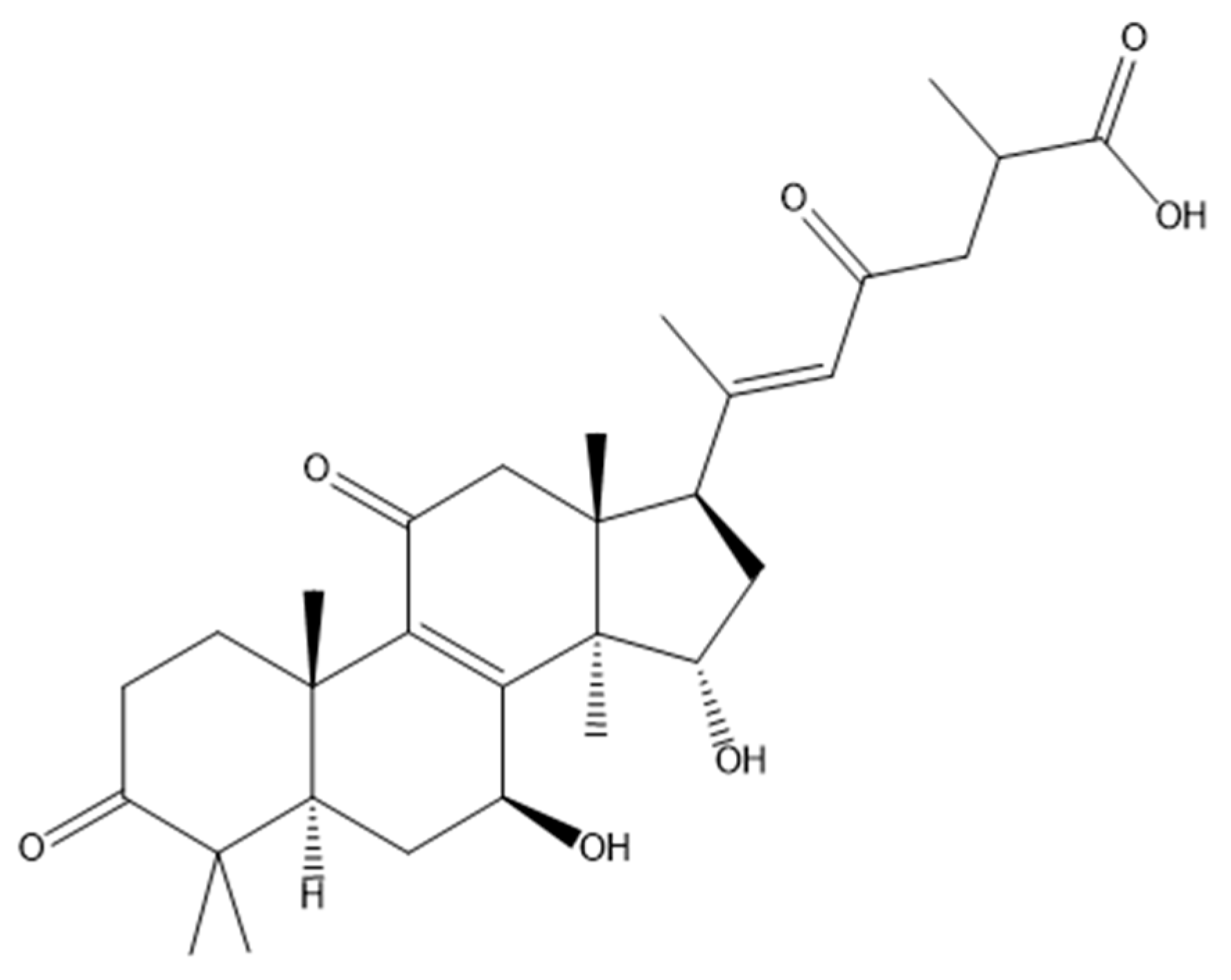
- Gill, B.S.; Navgeet; Kumar, S. Antioxidant potential of ganoderic acid in Notch-1 protein in neuroblastoma. Mol. Cell. Biochem. 2018, 456, 1–14. [Google Scholar] [CrossRef]
- Lixin, X.; Lijun, Y.; Songping, H. Ganoderic acid A against cyclophosphamide-induced hepatic toxicity in mice. J. Biochem. Mol. Toxicol. 2018, 33, e22271. [Google Scholar] [CrossRef]
- Lei, L.; Zheng, C.; Ye, J.F.; Zhu, K.; Zhou, Y.B.; Liu, J.; Gao, M.; Wu, Y.T.; Liu, Y.T.; Liu, L.Y.; et al. Metabolomic Profiling of Mice Exposed to α-amanitin Using Ultra-performance Liquid Chromatography Quadrupole Time-of-flight Tandem Mass Spectrometry. Biomed. Environ. Sci. 2023, 36, 289–294. [Google Scholar]
- Kwiatkowski, M.; Wurlitzer, M.; Krutilin, A.; Kiani, P.; Nimer, R.; Omidi, M.; Mannaa, A.; Bussmann, T.; Bartkowiak, K.; Kruber, S.; et al. Homogenization of tissues via picosecond-infrared laser (PIRL) ablation: Giving a closer view on the in-vivo composition of protein species as compared to mechanical homogenization. J. Proteom. 2016, 134, 193–202. [Google Scholar] [CrossRef]
- Grobler, T.; Opperman, M.; Bester, J.; Swanepoel, A.C.; du Preez, I. Metabolomic Profiling of Hormonal Contraceptive Use in Young Females Using a Commercially Available LC-MS/MS Kit. Metabolites 2023, 13, 1092. [Google Scholar] [CrossRef]
- Siegert, M.J.; Knittel, C.H.; Süssmuth, R.D. A Convergent Total Synthesis of the Death Cap Toxin α-Amanitin. Angew. Chem. Int. Ed. 2019, 59, 5500–5504. [Google Scholar] [CrossRef]
- Chen, X.; Shao, B.; Yu, C.; Yao, Q.; Ma, P.; Li, H.; Li, B.; Sun, C. Energy disorders caused by mitochondrial dysfunction contribute to α-amatoxin-induced liver function damage and liver failure. Toxicol. Lett. 2020, 336, 68–79. [Google Scholar] [CrossRef]
- Garcia, J.; Costa, V.M.; Bovolini, A.; Duarte, J.A.; Rodrigues, D.F.; Bastos, M.d.L.; Carvalho, F. An effective antidotal combination of polymyxin B and methylprednisolone for α-amanitin intoxication. Arch. Toxicol. 2019, 93, 1449–1463. [Google Scholar] [CrossRef]
- Garcia, J.; Costa, V.M.; Carvalho, A.T.P.; Silvestre, R.; Duarte, J.A.; Dourado, D.F.A.R.; Arbo, M.D.; Baltazar, T.; Dinis-Oliveira, R.J.; Baptista, P.; et al. A breakthrough on Amanita phalloides poisoning: An effective antidotal effect by polymyxin B. Arch. Toxicol. 2015, 89, 2305–2323. [Google Scholar] [CrossRef]
- Rahman, M. Ganoderic Acid for Anticancer Applications: Scope of Nanomedicine in Its Effective Delivery. In Nanomedicine for Bioactives: Healthcare Applications; Springer: Singapore, 2020; pp. 1–10. [Google Scholar]
- Liang, C.; Tian, D.; Liu, Y.; Li, H.; Zhu, J.; Li, M.; Xin, M.; Xia, J. Review of the molecular mechanisms of Ganoderma lucidum triterpenoids: Ganoderic acids A, C2, D, F, DM, X and Y. Eur. J. Med. Chem. 2019, 174, 130–141. [Google Scholar] [CrossRef]
- Mansour, H.H.; Hafez, H.F.; Fahmy, N.M. Silymarin Modulates Cisplatin-Induced Oxidative Stress and Hepatotoxicity in Rats. BMB Rep. 2006, 39, 656–661. [Google Scholar] [CrossRef]
- Kwo, P.Y.; Cohen, S.M.; Lim, J.K. ACG Clinical Guideline: Evaluation of Abnormal Liver Chemistries. Off. J. Am. Coll. Gastroenterol. ACG 2017, 112, 18–35. [Google Scholar] [CrossRef]
- Nyblom, H.; Berggren, U.; Balldin, J.; Olsson, R. High ast/alt ratio may indicate advanced alcoholic liver disease rather than heavy drinking. Alcohol Alcohol. 2004, 39, 336–339. [Google Scholar] [CrossRef]
- Chen, L.W.; Huang, M.-S.; Shyu, Y.-C.; Chien, R.-N. Gamma-glutamyl transpeptidase elevation is associated with metabolic syndrome, hepatic steatosis, and fibrosis in patients with nonalcoholic fatty liver disease: A community-based cross-sectional study. Kaohsiung J. Med. Sci. 2021, 37, 819–827. [Google Scholar] [CrossRef]
- Moneim, A.E.A.; Dkhil, M.; Al-Khalifa, M.S.; Al-Quraishy, S.; Zrieq, R. Indigofera oblongifolia mitigates lead-acetate-induced kidney damage and apoptosis in a rat model. Drug Des. Dev. Ther. 2016, 10, 1847–1856. [Google Scholar] [CrossRef]
- Wang, Z.; Gao, Y.; Huang, X.; Huang, S.; Yang, X.; Wang, J.; Zheng, N. Metabolomics analysis underlay mechanisms in the renal impairment of mice caused by combination of aflatoxin M1 and ochratoxin A. Toxicology 2021, 458, 152835. [Google Scholar] [CrossRef]
- Huang, Y.Q.; Tang, Y.-X.; Qiu, B.-H.; Talukder, M.; Li, X.-N.; Li, J.-L. Di-2-ethylhexyl phthalate (DEHP) induced lipid metabolism disorder in liver via activating the LXR/SREBP-1c/PPARa/γ and NF-κB signaling pathway. Food Chem. Toxicol. 2022, 165, 113119. [Google Scholar] [CrossRef]
- Yang, W.C.; Wang, Y.; Zhang, C.; Huang, Y.; Yu, J.; Shi, L.; Zhang, P.; Yin, Y.; Li, R.; Tao, K. Maresin1 Protect Against Ferroptosis-Induced Liver Injury Through ROS Inhibition and Nrf2/HO-1/GPX4 Activation. Front. Pharmacol. 2022, 13, 865689. [Google Scholar] [CrossRef]
- Ferrer, M.D.; Busquests-Cortes, C.; Capó, X.; Tejada, S.; Tur, J.A.; Pons, A.; Sureda, A. Cyclooxygenase-2 Inhibitors as a Therapeutic Target in Inflammatory Diseases. Curr. Med. Chem. 2019, 26, 3225–3241. [Google Scholar] [CrossRef]
- Farzanegi, P.; Dana, A.; Ebrahimpoor, Z.; Asadi, M.; Azarbayjani, M.A. Mechanisms of beneficial effects of exercise training on non-alcoholic fatty liver disease (NAFLD): Roles of oxidative stress and inflammation. Eur. J. Sport Sci. 2019, 19, 994–1003. [Google Scholar] [CrossRef]
- Shastak, Y.; Gordillo, A.; Pelletier, W. The relationship between vitamin A status and oxidative stress in animal production. J. Appl. Anim. Res. 2023, 51, 546–553. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Interactions between reactive oxygen species and autophagy. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2021, 1868, 119041. [Google Scholar] [CrossRef]
- Sinha, K.; Das, J.; Pal, P.B.; Sil, P.C. Oxidative stress: The mitochondria-dependent and mitochondria-independent pathways of apoptosis. Arch. Toxicol. 2013, 87, 1157–1180. [Google Scholar] [CrossRef]
- Frey, S.K.; Vogel, S. Vitamin A Metabolism and Adipose Tissue Biology. Nutrients 2011, 3, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Li, H. Innate immune regulatory networks in hepatic lipid metabolism. J. Mol. Med. 2019, 97, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Pegg, A.E. Functions of Polyamines in Mammals. J. Biol. Chem. 2016, 291, 14904–14912. [Google Scholar] [CrossRef] [PubMed]
- Aouida, M.; Ramotar, D. Identification of essential yeast genes involved in polyamine resistance. Gene 2018, 677, 361–369. [Google Scholar] [CrossRef]
- Pegg, A.E. Toxicity of Polyamines and Their Metabolic Products. Chem. Res. Toxicol. 2013, 26, 1782–1800. [Google Scholar] [CrossRef] [PubMed]
- Merrill, A.H.; Sullards, M.C.; Wang, E.; Voss, K.A.; Riley, R.T. Sphingolipid Metabolism: Roles in Signal Transduction and Disruption by Fumonisins. Environ. Health Perspect. 2001, 109 (Suppl. S2), 283–289. [Google Scholar] [PubMed]
- Linn, S.C.; Kim, H.S.; Keane, E.M.; Andras, L.M.; Wang, E.; Merrill, A.H. Regulation of de novo sphingolipid biosynthesis and the toxic consequences of its disruption. Biochem. Soc. Trans. 2001, 29, 831–835. [Google Scholar] [CrossRef] [PubMed]
- Schmelz, E.M.; Dombrink-Kurtzman, M.A.; Roberts, P.C.; Kozutsumi, Y.; Kawasaki, T.; Merrill, A.H. Induction of Apoptosis by Fumonisin B1in HT29 Cells Is Mediated by the Accumulation of Endogenous Free Sphingoid Bases. Toxicol. Appl. Pharmacol. 1998, 148, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Martínez, L.; Torres, S.; Baulies, A.; Alarcón-Vila, C.; Elena, M.; Fabriàs, G.; Casas, J.; Caballeria, J.; Fernandez-Checa, J.C.; García-Ruiz, C. Myristic acid potentiates palmitic acid-induced lipotoxicity and steatohepatitis associated with lipodystrophy by sustaning de novo ceramide synthesis. Oncotarget 2015, 6, 41479–41496. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Berk, M.; McIntyre, T.M.; Gores, G.J.; Feldstein, A.E. The lysosomal-mitochondrial axis in free fatty acid-induced hepatic lipotoxicity. Hepatology 2007, 47, 1495–1503. [Google Scholar] [CrossRef]
- Holme, E. Disorders of Tyrosine Degradation. In Physician’s Guide to the Laboratory Diagnosis of Metabolic Diseases; Blau, N., Duran, M., Blaskovics, M.E., Gibson, K.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 141–153. [Google Scholar]
- Holme, E.; Mitchell, G.A. Tyrosine Metabolism. In Physician’s Guide to the Diagnosis, Treatment, and Follow-Up of Inherited Metabolic Diseases; Blau, N., Vici, C.D., Ferreira, C.R., Vianey-Saban, C., van Karnebeek, C.D.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 23–31. [Google Scholar]
- Zhang, S.; Zeng, X.; Ren, M.; Mao, X.; Qiao, S. Novel metabolic and physiological functions of branched chain amino acids: A review. J. Anim. Sci. Biotechnol. 2017, 8, 10. [Google Scholar] [CrossRef] [PubMed]
- Sunny, N.E.; Kalavalapalli, S.; Bril, F.; Garrett, T.J.; Nautiyal, M.; Mathew, J.T.; Williams, C.M.; Cusi, K.; Koelmel, J.P.; Abdo, K.; et al. Cross-talk between branched-chain amino acids and hepatic mitochondria is compromised in nonalcoholic fatty liver disease. Am. J. Physiol. Metab. 2015, 309, E311–E319. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | ALT (U/L) | AST (U/L) | ALP (U/L) | GGT (U/L) | BUN (mmol/L) | Cr (μmol/L) |
|---|---|---|---|---|---|---|
| Control | 38.50 ± 4.83 | 61.26 ± 4.15 | 61.23 ± 6.25 | 4.36 ± 1.27 | 8.23 ± 0.82 | 32.45 ± 2.83 |
| AMA | 262.98 ± 34.28 a | 315.12 ± 42.13 a | 333.25 ± 56.47 a | 59.52 ± 10.92 a | 100.02 ± 11.28 a | 130.82 ± 18.43 a |
| AMA+GA (5 mg/kg bw) | 141.39 ± 15.45 b | 258.62 ± 20.98 b | 265.50 ± 34.58 b | 40.46 ± 9.83 b | 85.92 ± 10.52 b | 109.35 ± 14.33 b |
| AMA+GA (10 mg/kg bw) | 99.73 ± 8.41 bc | 200.62 ± 43.91 bc | 200.37 ± 23.21 bc | 33.39 ± 2.90 bc | 60.59 ± 9.66 bc | 87.75 ± 11.26 bc |
| AMA+GA (20 mg/kg bw) | 79.55 ± 9.74 bcd | 138.88 ± 14.30 bcd | 178.62 ± 36.17 bcd | 24.20 ± 2.03 bcd | 40.56 ± 4.65 bcd | 67.90 ± 11.28 bcd |
| AMA+GA (40 mg/kg bw) | 46.61 ± 9.95 bcde | 106.62 ± 10.42 bcde | 142.38 ± 22.13 bcde | 13.71 ± 2.18 bcde | 15.76 ± 5.53 bcde | 51.01 ± 1.62 bcde |
| Group | ALT (U/L) | AST (U/L) | ALP (U/L) | GGT (U/L) | BUN (mmol/L) | Cr (μmol/L) |
|---|---|---|---|---|---|---|
| Control | 40.92 ± 6.94 | 72.75 ± 6.34 | 55.62 ± 11.72 | 4.84 ± 0.46 | 5.64 ± 0.70 | 35.41 ± 1.98 |
| AMA (24 h) | 157.24 ± 22.72 a | 140.86 ± 23.81 a | 114.00 ± 11.03 a | 19.07 ± 9.76 a | 12.93 ± 2.71 a | 80.67 ± 10.92 a |
| AMA+GA (24 h) | 51.21 ± 16.14 b | 118.62 ± 11.21 b | 74.00 ± 7.82 b | 10.39 ± 2.91 b | 9.85 ± 0.96 b | 49.54 ± 13.07 b |
| AMA (48 h) | 216.65 ± 28.01 ab | 192.75 ± 44.87 ab | 253.50 ± 15.70 ab | 27.24 ± 4.70 ab | 23.04 ± 4.47 ab | 160.35 ± 26.43 ab |
| AMA+GA (48 h) | 68.46 ± 8.85 c | 117.86 ± 15.26 c | 101.43 ± 37.66 c | 14.74 ± 1.25 c | 8.96 ± 0.85 c | 54.14 ± 4.91 c |
| Group | ROS (pmoL/mL) | IL-6 (μg/mL) | TNF-α (μg/mL) | COX-2 (mg/mL) |
|---|---|---|---|---|
| Control | 40.67 ± 6.14 | 32.21 ± 7.04 | 444.67 ± 96.90 | 113.33 ± 14.71 |
| AMA (24 h) | 100.38 ± 9.39 a | 103.67 ± 5.20 a | 1813.50 ± 84.60 a | 218.17 ± 15.97 a |
| AMA+GA (24 h) | 69.52 ± 5.33 b | 69.78 ± 7.23 b | 793.03 ± 134.86 b | 152.67 ± 16.79 b |
| AMA (48 h) | 202.65 ± 13.85 ab | 171.67 ± 11.91 ab | 3749.48 ± 109.50 ab | 311.17 ± 15.50 ab |
| AMA+GA (48 h) | 103.50 ± 9.63 c | 96.91 ± 14.04 c | 1272.83 ± 89.96 c | 212.50 ± 11.31 c |
| Metabolite | Retention Time (min) | Mass Found | Adducts | Exact Mass | MS/MS Fragments | MS/MS |
|---|---|---|---|---|---|---|
| CE (eV) | ||||||
| Retinal | 11.92 | 285.2217 | [M+H]+ | 284.214 | 161.0955 | 20 |
| 175.1479 | ||||||
| 119.0849 | ||||||
| Retinyl ester | 13.87 | 303.2311 | [M+H]+ | 302.2246 | 285.221 | 20 |
| 135.1157 | ||||||
| 103.0533 | ||||||
| Retinol | 14.56 | 287.2373 | [M+H]+ | 286.2297 | 121.0351 | 20 |
| 269.2248 | ||||||
| 93.0686 | ||||||
| Retinoate | 11.06 | 301.2167 | [M+H]+ | 300.2089 | 123.1166 | 20 |
| 161.0963 | ||||||
| 81.0696 | ||||||
| Spermidine | 0.68 | 146.1645 | [M+H]+ | 145.1579 | 72.0809 | 20 |
| 84.0809 | ||||||
| 112.1122 | ||||||
| Spermine | 0.7 | 203.2234 | [M+H]+ | 202.2157 | 112.1122 | 20 |
| 129.1389 | ||||||
| 84.0804 | ||||||
| Sphinganine | 8.76 | 302.3058 | [M+H]+ | 301.2981 | 60.0451 | 20 |
| 284.2949 | ||||||
| 252.2844 | ||||||
| Tryptophan | 2.59 | 205.0967 | [M+H]+ | 204.0899 | 146.0602 | 20 |
| 118.0649 | ||||||
| 188.0714 | ||||||
| Palmitic acid | 14.78 | 255.2324 | [M−H]− | 256.2402 | 255.2324 | 20 |
| Docosahexaenoic acid | 13.42 | 327.2325 | [M−H]− | 328.2402 | 283.2424 | 20 |
| 327.2323 | ||||||
| 229.1955 | ||||||
| Myristic acid | 12.51 | 227.2019 | [M−H]− | 228.2089 | 277.2019 | 20 |
| 68.9957 | ||||||
| α-Linolenic acid | 12.66 | 277.2174 | [M−H]− | 278.2246 | 277.2174 | 20 |
| 59.0138 | ||||||
| γ-Linolenic acid | 12.84 | 277.2177 | [M−H]− | 278.2246 | 277.2177 | 20 |
| 59.0141 | ||||||
| Tyrosine | 0.92 | 180.0667 | [M−H]− | 181.0739 | 119.0547 | 20 |
| 163.0535 | ||||||
| 72.0099 | ||||||
| 4-Methyl-2-oxopentanoate | 0.93 | 129.0562 | [M−H]− | 130.063 | 129.0562 | 10 |
| 101.0604 | ||||||
| Hydroxyphenylpyruvic acid | 0.92 | 179.0353 | [M−H]− | 180.0423 | 179.0353 | 20 |
| 135.0452 | ||||||
| 106.9578 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, C.; Lv, S.; Ye, J.; Zou, L.; Zhu, K.; Li, H.; Dong, Y.; Li, L. Metabolomic Insights into the Mechanisms of Ganoderic Acid: Protection against α-Amanitin-Induced Liver Injury. Metabolites 2023, 13, 1164. https://doi.org/10.3390/metabo13111164
Zheng C, Lv S, Ye J, Zou L, Zhu K, Li H, Dong Y, Li L. Metabolomic Insights into the Mechanisms of Ganoderic Acid: Protection against α-Amanitin-Induced Liver Injury. Metabolites. 2023; 13(11):1164. https://doi.org/10.3390/metabo13111164
Chicago/Turabian StyleZheng, Chong, Shaofang Lv, Jianfang Ye, Lu Zou, Kai Zhu, Haichang Li, Yongxi Dong, and Lei Li. 2023. "Metabolomic Insights into the Mechanisms of Ganoderic Acid: Protection against α-Amanitin-Induced Liver Injury" Metabolites 13, no. 11: 1164. https://doi.org/10.3390/metabo13111164