
Manifestations of Liver Impairment and the Effects of MH-76, a Non-Quinazoline α1-Adrenoceptor Antagonist, and Prazosin on Liver Tissue in Fructose-Induced Metabolic Syndrome
 , , ,
, , , 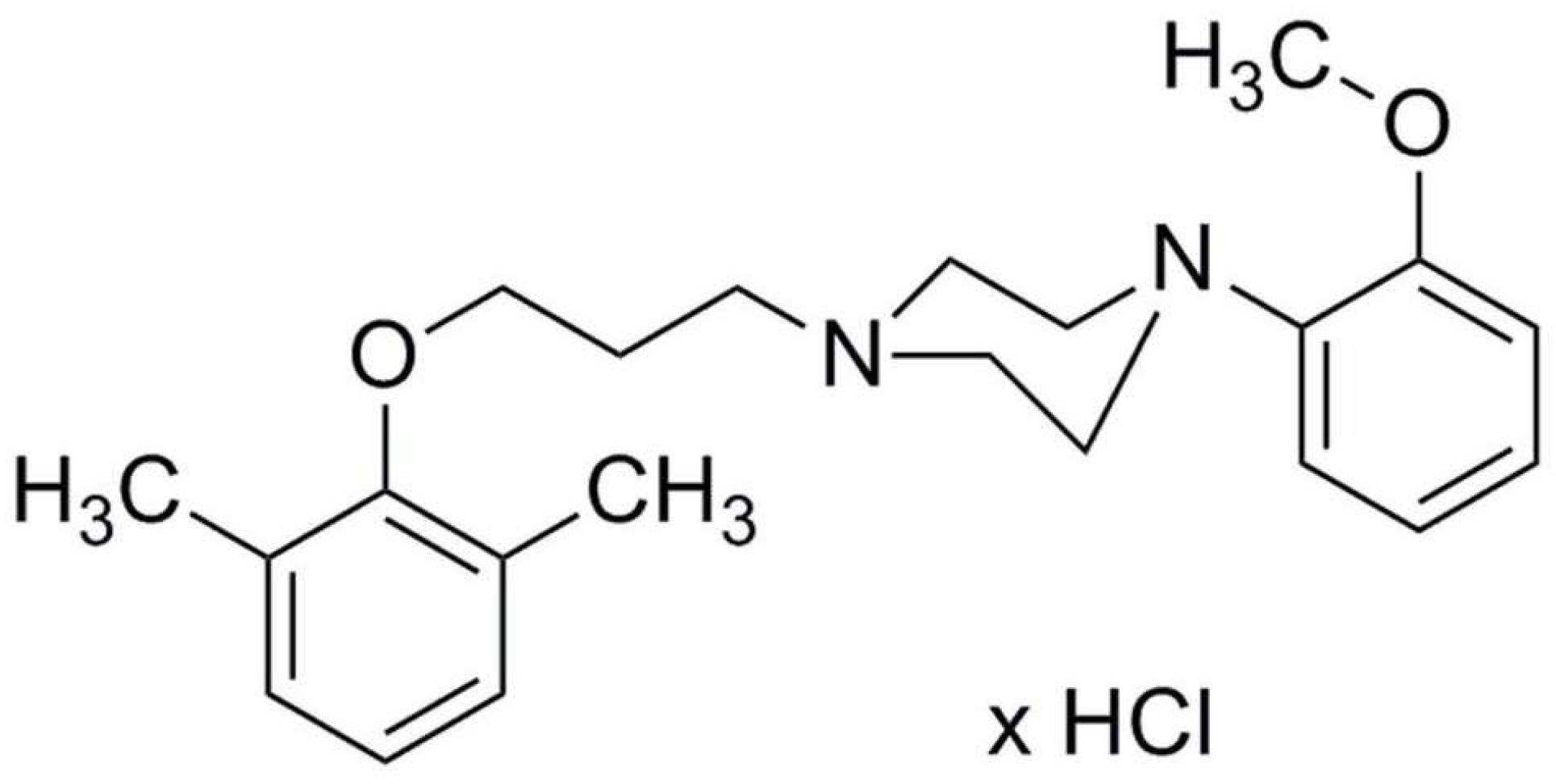{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Protocol
2.2. Drugs and Chemicals
2.3. Biochemical Assays
2.3.1. Preparation of Tissue Homogenates
2.3.2. Determination of the Non-Protein Thiols Levels (NPSH)
2.3.3. The Ferric Reducing Antioxidant Power (FRAP) Assay
2.4. Histopathological Evaluation and Image Analysis and Identifying and Quantifying Liver Fibrosis
2.5. Hepatotoxicity Assay
2.6. Statistical Analysis
3. Results
3.1. Effect of MH-76 and Prazosin on the Concentration of Proinflammatory Factors in Liver Tissue
3.2. Effect of MH-76 and Prazosin on Non-Proteine Thiols and Total Reducing Activity in Liver Tissue
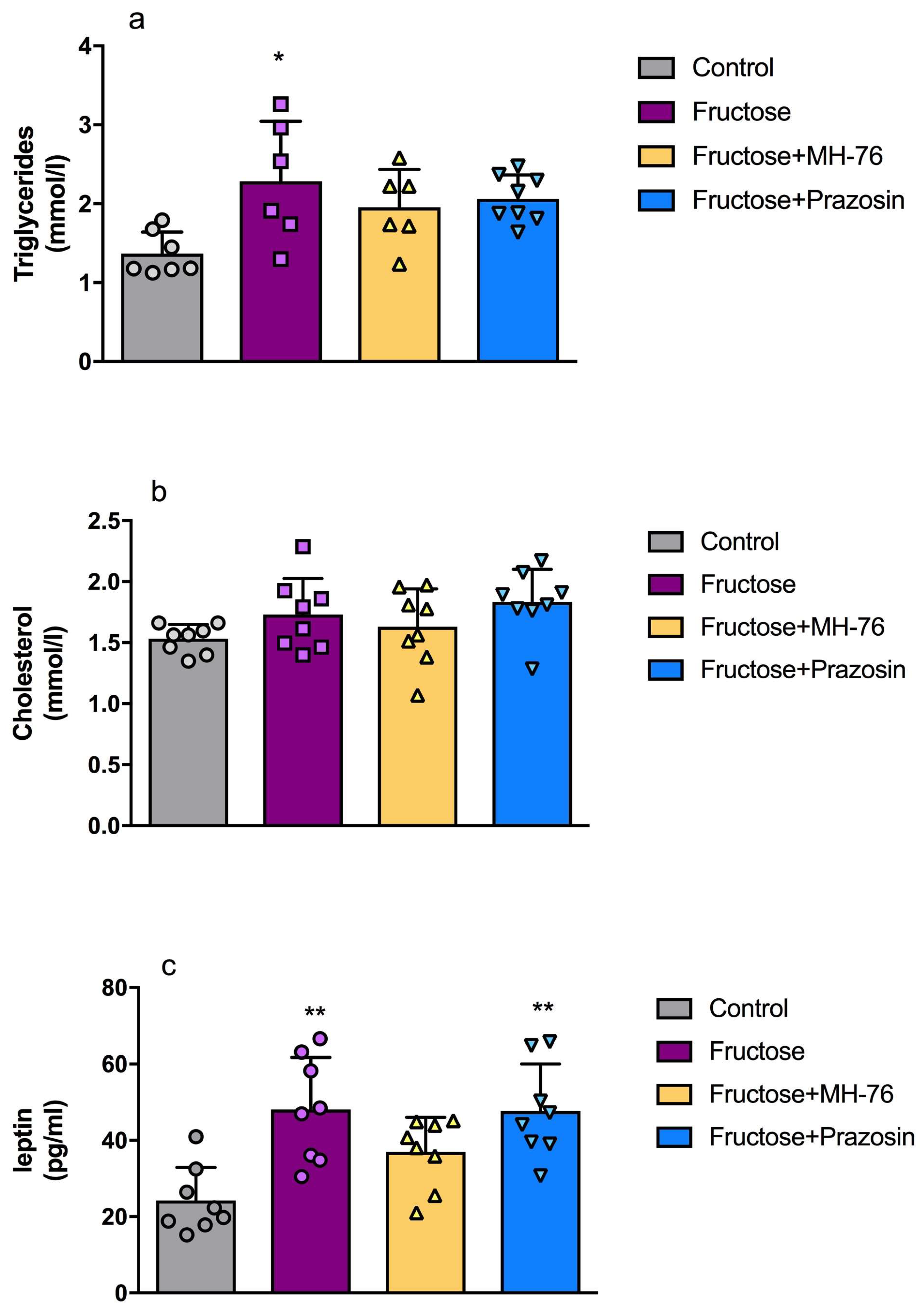
3.3. Effect of MH-76 and Prazosin on the Lipid Content and Leptin Concentration in Liver Tissue
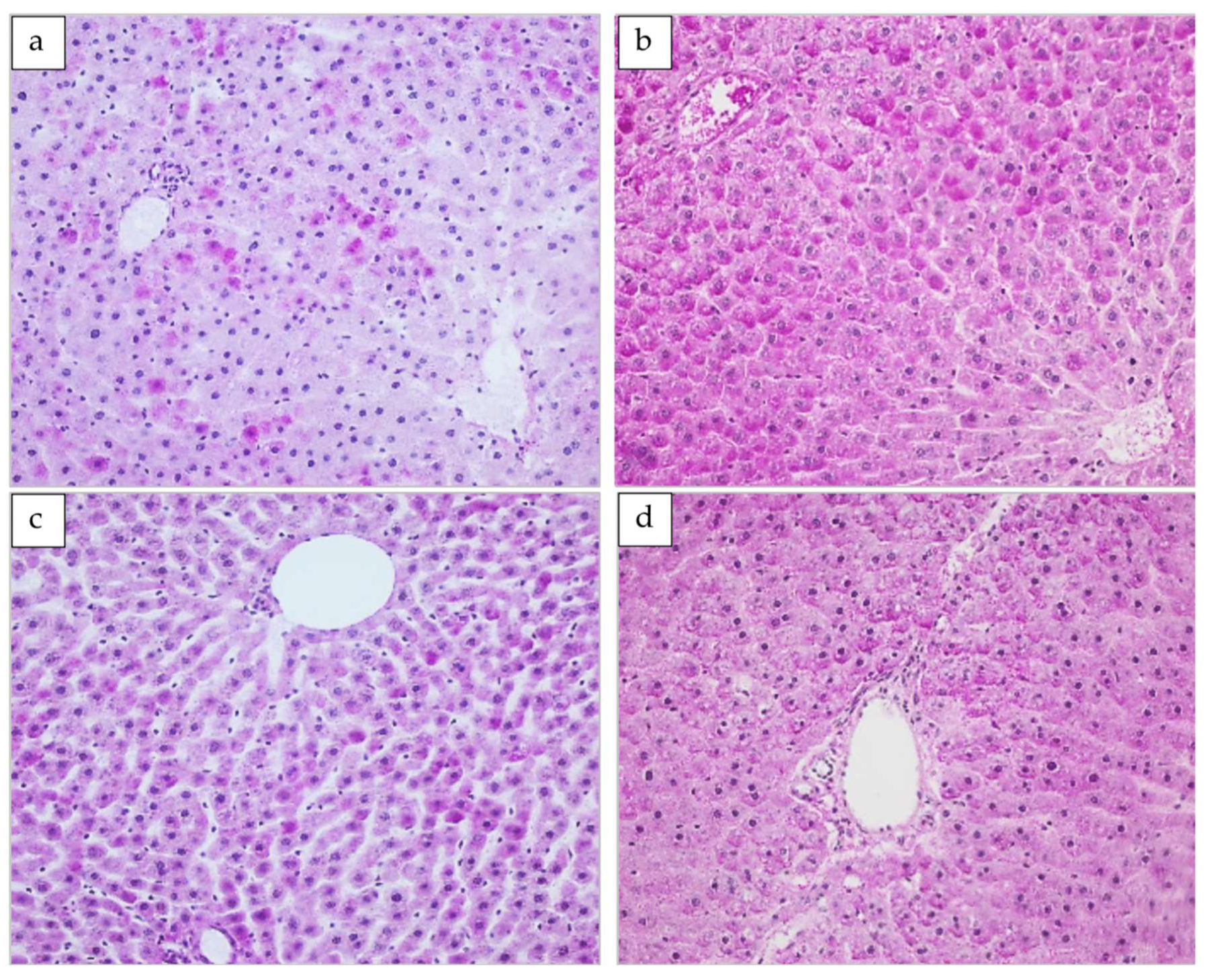
3.4. Histopathological Examination of LiverTissue
3.4.1. The Basic Lobular Structure
3.4.2. Glycogen Deposits
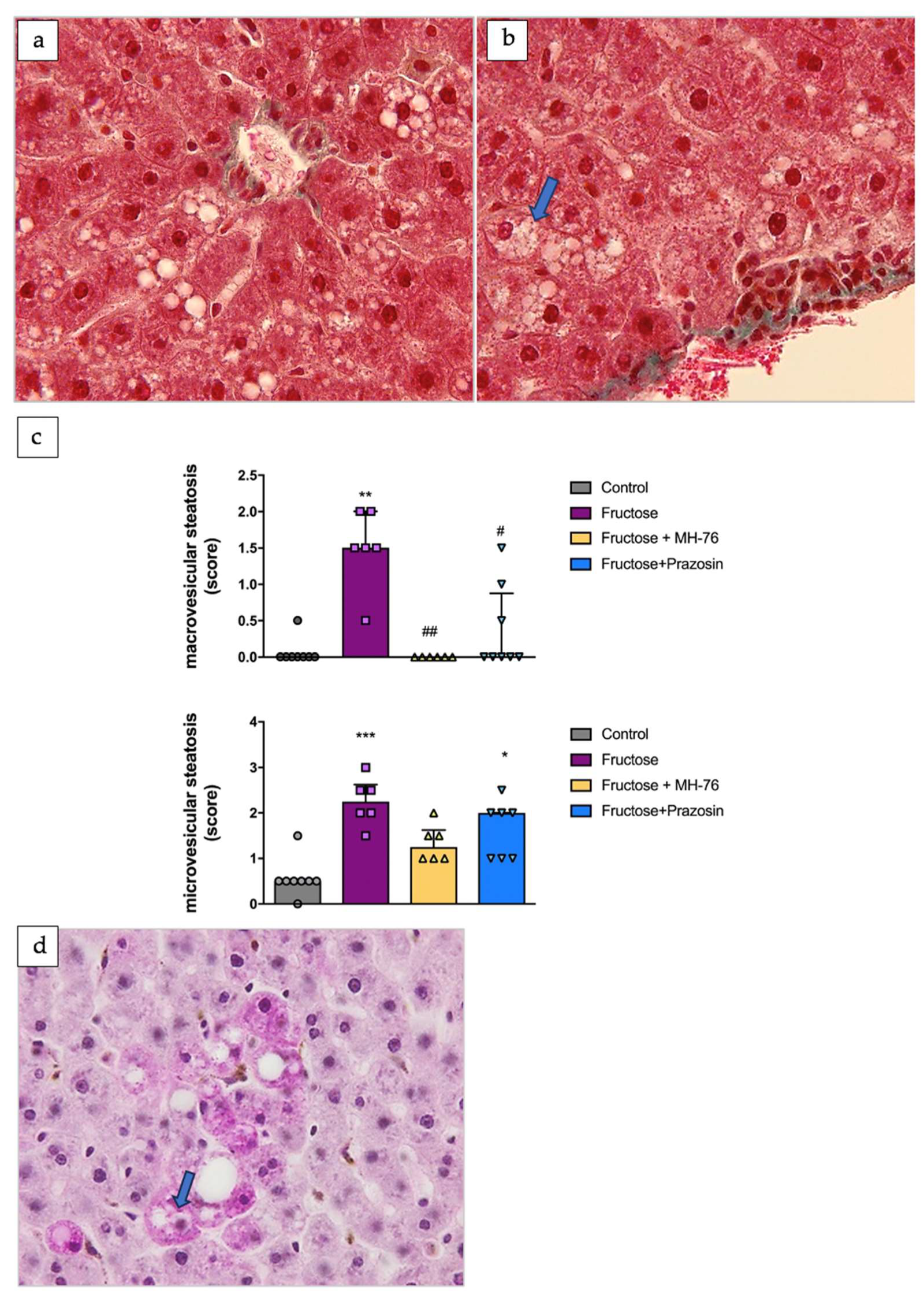
3.4.3. Steatosis
3.4.4. Periportal Fibrosis and Inflammation
3.4.5. Pericentral Fibrosis and Inflammation
3.4.6. Sinusoidal Changes and Sinusoidal Cellularity
3.4.7. Pigment Deposits
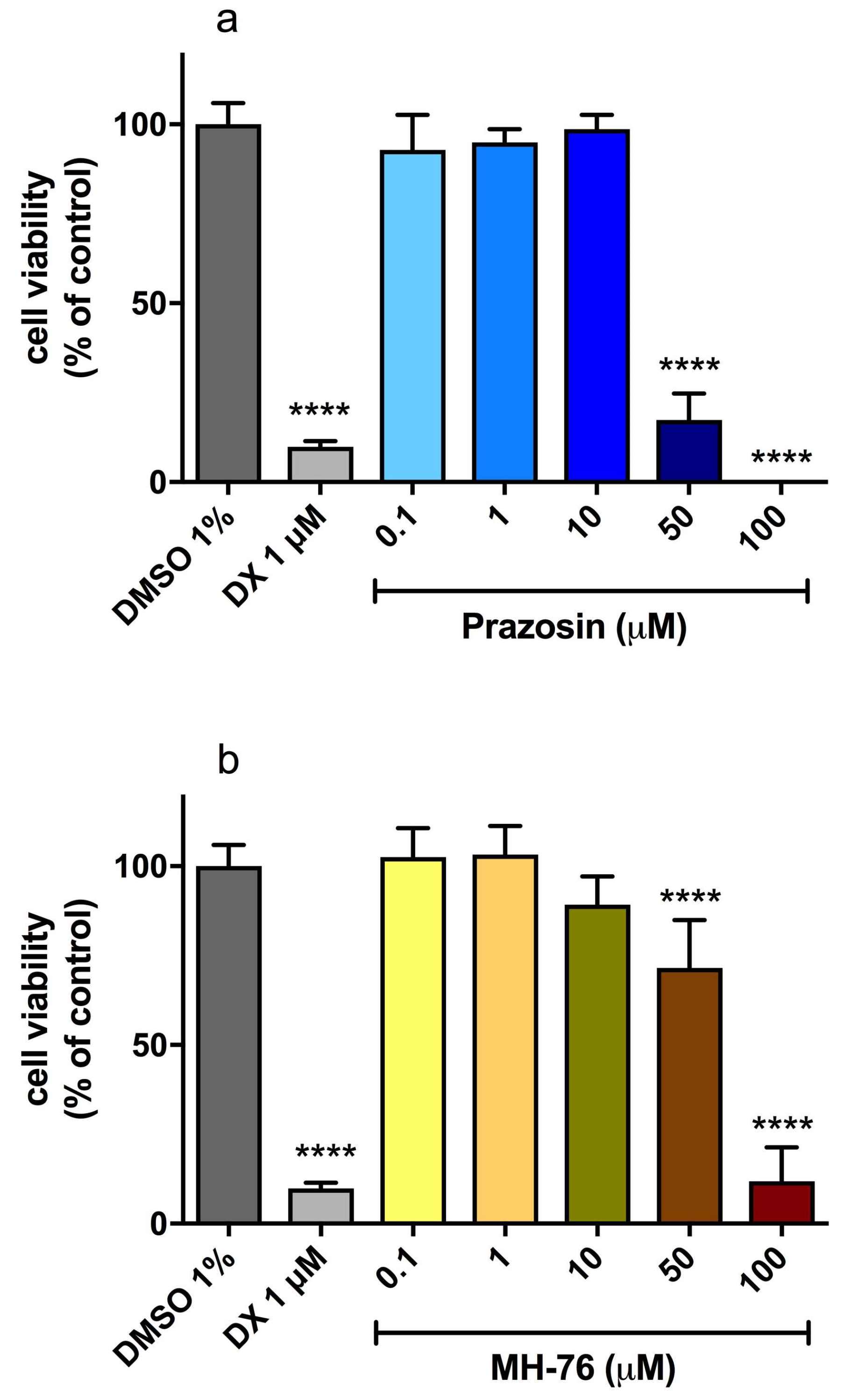
3.5. Hepatotoxicity of MH-76 and Prazosin
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alwahsh, S.M.; Xu, M.; Seyhan, H.A.; Ahmad, S.; Mihm, S.; Ramadori, G.; Schultze, F.C. Diet high in fructose leads to an overexpression of lipocalin-2 in rat fatty liver. World J. Gastroenterol. 2014, 20, 1807–1821. [Google Scholar] [CrossRef]
- Drożdż, K.; Nabrdalik, K.; Hajzler, W.; Kwiendacz, H.; Gumprecht, J.; Lip, G.Y.H. Metabolic-Associated Fatty Liver Disease (MAFLD), Diabetes, and Cardiovascular Disease: Associations with Fructose Metabolism and Gut Microbiota. Nutrients 2021, 14, 103. [Google Scholar] [CrossRef]
- Wieser, V.; Moschen, A.R.; Tilg, H. Inflammation, cytokines and insulin resistance: A clinical perspective. Arch. Immunol. Ther. Exp. 2013, 61, 119–125. [Google Scholar] [CrossRef]
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of NAFLD and NASH trends, predictors, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11–20. [Google Scholar] [CrossRef]
- Rinella, M.E.; Lazarus, J.V.; Ratziu, V.; Francque, S.M.; Sanyal, A.J.; Kanwal, F.; Romero, D.; Abdelmalek, M.F.; Anstee, Q.M.; Arab, J.P.; et al. NAFLD Nomenclature consensus group. A multi-society Delphi consensus statement on new fatty liver disease nomenclature. Hepatology 2023, 1, 1–15. [Google Scholar]
- Carnagarin, R.; Tan, K.; Adams, L.; Matthews, V.B.; Kiuchi, M.G.; Marisol Lugo Gavidia, L.; Lambert, G.W.; Lambert, E.A.; Herat, L.Y.; Schlaich, M.P. Metabolic Dysfunction-Associated Fatty Liver Disease (MAFLD)—A Condition Associated with Heightened Sympathetic Activation. Int. J. Mol. Sci. 2021, 22, 4241. [Google Scholar] [CrossRef] [PubMed]
- Kho, M.C.; Lee, Y.J.; Park, J.H.; Cha, J.D.; Choi, K.M.; Kang, D.G.; Lee, H.S. Combination with Red ginseng and Polygoni Multiflori ameliorates high fructose diet induced metabolic syndrome. BMC Complement. Altern. Med. 2016, 16, 98. [Google Scholar] [CrossRef]
- Jin, X.; Qiu, T.; Li, L.; Yu, R.; Chen, X.; Li, C.; Proud, C.G.; Jiang, T. Pathophysiology of obesity and its associated diseases. Acta Pharm. Sin. B 2023, 13, 2403–2424. [Google Scholar] [CrossRef]
- Marjot, T.; Moolla, A.; Cobbold, J.F.; Hodson, L.; Tomlinson, J.W. Nonalcoholic fatty liver disease in adults: Current concepts in etiology, outcomes, and management. Endocr. Rev. 2020, 41, 66–1172. [Google Scholar] [CrossRef]
- Duan, Y.; Pan, X.; Luo, J.; Xiao, X.; Li, J.; Bestman, P.L.; Luo, M. Association of Inflammatory Cytokines with Non-Alcoholic Fatty Liver Disease. Front. Immunol. 2022, 13, 880298. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, B.; Sultana, R.; Greene, M.W. Adipose tissue and insulin resistance in obese. Biomed. Pharmacother. 2021, 137, 111315. [Google Scholar] [CrossRef] [PubMed]
- Badmus, O.O.; Hinds, T.D., Jr.; Stec, D.E. Mechanisms Linking Metabolic-Associated Fatty Liver Disease (MAFLD) to Cardiovascular Disease. Curr. Hypertens. Rep. 2023, 8, 151–162. [Google Scholar] [CrossRef]
- Carnagarin, R.; Gregory, C.; Azzam, O.; Hillis, G.S.; Schultz, C.; Watts, G.F.; Bell, D.; Matthews, V.; Schlaich, M.P. The Role of Sympatho-Inhibition in Combination Treatment of Obesity-Related Hypertension. Curr. Hypertens. Rep. 2017, 19, 99. [Google Scholar] [CrossRef]
- Schwinghammer, U.A.; Melkonyan, M.M.; Hunanyan, L.; Tremmel, R.; Weiskirchen, R.; Borkham-Kamphorst, E.; Schaeffeler, E.; Seferyan, T.; Mikulits, W.; Yenkoyan, K.; et al. α2-Adrenergic Receptor in Liver Fibrosis: Implications for the Adrenoblocker Mesedin. Cells 2020, 18, 456. [Google Scholar] [CrossRef]
- Serna-Salas, S.A.; Arroyave-Ospina, J.C.; Zhang, M.; Damba, T.; Buist-Homan, M.; Muñoz-Ortega, M.H.; Ventura-Juárez, J.; Moshage, H. α-1 Adrenergic receptor antagonist doxazosin reverses hepatic stellate cells activation via induction of senescence. Mech. Ageing Dev. 2022, 201, 111617. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Ortega, M.H.; Llamas-Ramírez, R.W.; Romero-Delgadillo, N.I.; Elías-Flores, T.G.; Tavares-Rodríguez, E.d.J.; Campos-Esparza, M.d.R.; Cervantes-García, D.; Muñoz-Fernández, L.; Gerardo-Rodríguez, M.; Ventura-Juárez, J. Doxazosin Treatment Attenuates Carbon Tetrachloride-Induced Liver Fibrosis in Hamsters through a Decrease in Transforming Growth Factor β Secretion. Gut Liver 2016, 10, 101–108. [Google Scholar] [CrossRef]
- Chapman, N.; Chen, C.Y.; Fujita, T.; Hobbs, F.D.; Kim, S.J.; Staessen, J.A.; Tanomsup, S.; Wang, J.G.; Williams, B. Time to re-appraise the role of alpha-1 adrenoceptor antagonists in the management of hypertension? J. Hypertens. 2010, 28, 1796–1803. [Google Scholar] [CrossRef] [PubMed]
- Pessina, A.C.; Ciccariello, L.; Perrone, F.; Stoico, V.; Gussoni, G.; Scotti, A.; Muggeo, M. Clinical efficacy and tolerability of alpha-blocker doxazosin as add-on therapy in patients with hypertension and impaired glucose metabolism. Nutr. Metab. Cardiovasc. Dis. 2006, 16, 137–147. [Google Scholar] [CrossRef]
- Shannon, R.; Chaudhry, M. Effect of alpha1-adrenergic receptors in cardiac pathophysiology. Am. Heart J. 2006, 152, 842–850. [Google Scholar] [CrossRef] [PubMed]
- Eiras, S.; Fernández, P.; Piñeiro, R.; Iglesias, M.J.; González-Juanatey, J.R.; Lago, F. Doxazosin induces activation of GADD153 and cleavage of focal adhesion kinase in cardiomyocytes en route to apoptosis. Cardiovasc. Res. 2006, 71, 118–128. [Google Scholar] [CrossRef] [PubMed]
- González-Juanatey, J.R.; Iglesias, M.J.; Alcaide, C.; Piñeiro, R.; Lago, F. Doxazosin induces apoptosis in cardiomyocytes cultured in vitro by a mechanism that is independent of alpha1-adrenergic blockade. Circulation 2003, 107, 127–131. [Google Scholar] [CrossRef]
- Bilbro, J.; Mart, M.; Kyprianou, N. Therapeutic value of quinazoline-based compounds in prostate cancer. Anticancer. Res. 2013, 33, 4695–4700. [Google Scholar] [PubMed]
- King, L.; Christie, D.; Dare, W.; Bernaitis, N.; Chess-Williams, R.; McDermott, C.; Forbes, A.; Anoopkumar-Dukie, S. Quinazoline alpha-adrenoreceptor blockers as an adjunct cancer treatment: From bench to bedside. Eur. J. Pharmacol. 2021, 893, 173831. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Li, M.; Wagner, M.B.; Chen, G.; Song, X. Doxazosin Stimulates Galectin-3 Expression and Collagen Synthesis in HL-1 Cardiomyocytes Independent of Protein Kinase C Pathway. Front. Pharmacol. 2016, 7, 495. [Google Scholar] [CrossRef]
- Liu, J.; Takase, I.; Hakucho, A.; Okamura, N.; Fujimiya, T. Carvedilol attenuates the progression of alcohol fatty liver disease in rats. Alcohol. Clin. Exp. Res. 2012, 36, 1587–1599. [Google Scholar] [CrossRef]
- McKee, C.; Soeda, J.; Asilmaz, E.; Sigalla, B.; Morgan, M.; Sinelli, N.; Roskams, T.; Oben, J.A. Propranolol, a β-adrenoceptor antagonist, worsens liver injury in a model of non-alcoholic steatohepatitis. Biochem. Biophys. Res. Commun. 2013, 437, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Randle, L.E.; Sathish, J.G.; Kitteringham, N.R.; Macdonald, I.; Williams, D.P.; Park, B.K. alpha(1)-Adrenoceptor antagonists prevent paracetamol-induced hepatotoxicity in mice. Br. J. Pharmacol. 2008, 153, 820–830. [Google Scholar] [CrossRef]
- Rose, L.; Graham, L.; Koenecke, A.; Powell, M.; Xiong, R.; Shen, Z.; Mench, B.; Kinzler, K.W.; Bettegowda, C.; Vogelstein, B.; et al. The Association between Alpha-1 Adrenergic Receptor Antagonists and In-Hospital Mortality from COVID-19. Front. Med. 2021, 8, 637647. [Google Scholar] [CrossRef]
- Renna, N.F.; Diez, E.A.; Miatello, R.M. Effects of dipeptidyl-peptidase 4 inhibitor about vascular inflammation in a metabolic syndrome model. PLoS ONE 2014, 9, e106563, eCollection 2014. [Google Scholar] [CrossRef]
- Kubacka, M.; Kotańska, M.; Szafarz, M.; Pociecha, K.; Waszkielewicz, A.M.; Marona, H.; Filipek, B.; Mogilski, S. Beneficial effects of non-quinazoline α1-adrenolytics on hypertension and altered metabolism in fructose-fed rats. A comparison with prazosin. Nutr. Metab. Cardiovasc. Dis. 2019, 29, 751–760. [Google Scholar] [CrossRef]
- Kubacka, M.; Mogilski, S.; Zadrożna, M.; Nowak, B.; Szafarz, M.; Pomierny, B.; Marona, H.; Waszkielewicz, A.; Jawień, W.; Sapa, J.; et al. MH-76, a Novel Non-Quinazoline α1-Adrenoceptor Antagonist, but Not Prazosin Reduces Inflammation and Improves Insulin Signaling in Adipose Tissue of Fructose-Fed Rats. Pharmaceuticals 2021, 14, 477. [Google Scholar] [CrossRef]
- Marona, H.; Kubacka, M.; Filipek, B.; Siwek, A.; Dybała, M.; Szneler, E.; Pociecha, T.; Gunia, A.; Waszkielewicz, A.M. Synthesis, alpha-adrenoceptors affinity and alpha 1-adrenoceptor antagonistic properties of some 1,4-substituted piperazine derivatives. Pharmazie 2011, 66, 733–739. [Google Scholar] [PubMed]
- Mika, K.; Szafarz, M.; Bednarski, M.; Latacz, G.; Sudoł, S.; Handzlik, J.; Pociecha, K.; Knutelska, J.; Nicosia, N.; Szczepańska, K.; et al. Histamine H3 Receptor Ligands-KSK-59 and KSK-73-Reduce Body Weight Gain in a Rat Model of Excessive Eating. Pharmaceuticals 2021, 14, 1080. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound and non-protein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Benzie, I.; Strain, J. Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous mea- surement of total antioxidant power and ascorbic. Enzymology 1999, 299, 15–27. [Google Scholar]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Nonalcoholic Steatohepatitis Clinical Research Network. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Fukusato, T. Histopathology of nonalcoholic fatty liver disease/nonalcoholic steatohepatitis. World J. Gastroenterol. 2014, 20, 15539–15548. [Google Scholar] [CrossRef]
- Latacz, G.; Lubelska, A.; Jastrzębska-Więsek, M.; Partyka, A.; Marć, M.A.; Satała, G.; Wilczyńska, D.; Kotańska, M.; Więcek, M.; Kamińska, K.; et al. The 1,3,5-Triazine Derivatives as Innovative Chemical Family of 5-HT6 Serotonin Receptor Agents with Therapeutic Perspectives for Cognitive Impairment. Int. J. Mol. Sci. 2019, 20, 3420. [Google Scholar] [CrossRef]
- Mak, K.M.; Png, C.Y.M. The Hepatic Central Vein: Structure, Fibrosis, and Role in Liver Biology. Anat. Rec. 2020, 303, 1747–1767. [Google Scholar] [CrossRef] [PubMed]
- Ter Horst, K.W.; Serlie, M.J. Fructose Consumption, Lipogenesis, and Non-Alcoholic Fatty Liver Disease. Nutrients 2017, 9, 981. [Google Scholar] [CrossRef]
- Sakurai, Y.; Kubota, N.; Yamauchi, T.; Kadowaki, T. Role of Insulin Resistance in MAFLD. Int. J. Mol. Sci. 2021, 22, 4156. [Google Scholar] [CrossRef]
- Yu, S.; Li, C.; Ji, G.; Zhang, L. The Contribution of Dietary Fructose to Non-alcoholic Fatty Liver Disease. Front. Pharmacol. 2021, 12, 783393. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, T.; Li, J.; Wang, S.; Qiu, F.; Yu, H.; Zhang, Y.; Wang, T. Effects of Natural Products on Fructose-Induced Nonalcoholic Fatty Liver Disease (NAFLD). Nutrients 2017, 9, 96. [Google Scholar] [CrossRef]
- Pan, Y.; Kong, L.D. High fructose diet-induced metabolic syndrome: Pathophysiological mechanism and treatment by traditional Chinese medicine. Pharmacol. Res. 2018, 130, 438–450. [Google Scholar] [CrossRef]
- López Panqueva, R.D.P. Useful Algorithms for Histopathological Diagnosis of Liver Disease Based on Patterns of Liver Damage. Rev. Col. Gastroenterol. 2016, 31, 443–457. [Google Scholar]
- Korenblat, K.M.; Fabbrini, E.; Mohammed, B.S.; Klein, S. Liver, muscle and adipose tissue insulin action is directly related to intrahepatic triglyceride content in obese subjects. Gastroenterology 2008, 134, 1369–1375. [Google Scholar] [CrossRef]
- Watt, M.J.; Miotto, P.M.; De Nardo, W.; Montgomery, M.K. The liver as an endocrine organ-linking NAFLD and insulin resistance. Endocr. Rev. 2019, 40, 1367–1393. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, S.; Iwasaki, Y.; Matsumoto, S.; Satoh, T.; Ozawa, A.; Yamada, E.; Kakizaki, S.; Trejo, J.A.O.; Uchiyama, Y.; Yamada, M.; et al. Administration of small-molecule guanabenz acetate attenuates fatty liver and hyperglycemia associated with obesity. Sci. Rep. 2020, 10, 13671. [Google Scholar] [CrossRef]
- Benedict, M.; Zhang, X. Non-alcoholic fatty liver disease: An expanded review. World J. Hepatol. 2017, 9, 715–732. [Google Scholar] [CrossRef]
- Tandra, S.; Yeh, M.M.; Brunt, E.M.; Vuppalanchi, R.; Cummings, O.W.; Ünalp-Arida, A.; Wilson, L.A.; Chalasani, N. Presence and significance of microvesicular steatosis in nonalcoholic fatty liver disease. J. Hepatol. 2011, 55, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Zhong, F.; Zhou, X.; Xu, J.; Gao, L. Rodent Models of Nonalcoholic Fatty Liver Disease. Digestion 2020, 101, 522–535. [Google Scholar] [CrossRef] [PubMed]
- Taskinen, M.R.; Packard, C.J.; Borén, J. Dietary Fructose and the Metabolic Syndrome. Nutrients 2019, 11, 1987. [Google Scholar] [CrossRef] [PubMed]
- Vilà, L.; Roglans, N.; Alegret, M.; Sánchez, R.M.; Vázquez-Carrera, M.; Laguna, J.C. Suppressor of cytokine signaling-3 (SOCS-3) and a deficit of serine/threonine (Ser/Thr) phosphoproteins involved in leptin transduction mediate the effect of fructose on rat liver lipid metabolism. Hepatology 2008, 48, 1506–1516. [Google Scholar] [CrossRef]
- Bilska-Wilkosz, A.; Ochenduszka, M.; Iciek, M.; Sokołowska-Jezewicz, M.; Wiliński, B.; Góralska, M.; Srebro, Z.; Włodek, L. Effects of acetylsalicylic acid on the levels of sulfane sulfur and non-protein sulfhydryl groups in mouse tissues. Pharmacol. Rep. 2013, 65, 173–178. [Google Scholar] [CrossRef]
- Ulrich, K.; Jakob, U. The role of thiols in antioxidant systems. Free. Radic. Biol. Med. 2019, 140, 14–27. [Google Scholar] [CrossRef]
- Liu, T.; Sun, L.; Ahang, Y.; Wang, Y.; Zheng, J. Imbalanced GSH/ROS and sequential cell death. J. Biochem. Mol. Toxicol. 2022, 36, e22942. [Google Scholar] [CrossRef]
- Worner, T.M.; Lieber, C.S. Perivenular fibrosis as precursor lesion of cirrhosis. JAMA 1985, 254, 627–630. [Google Scholar] [CrossRef] [PubMed]
- Opal, S.M.; DePalo, V.A. Anti-inflammatory cytokines. Chest 2000, 117, 1162–1172. [Google Scholar] [CrossRef]
- Xing, Z.; Gauldie, J.; Cox, G.; Baumann, H.; Jordana, M.; Lei, X.F.; Achong, M.K. IL-6 is an anti-inflammatory cytokine required for controlling local or systemic acute inflammatory responses. J. Clin. Investig. 1998, 101, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Libert, C.; Takahashi, N.; Cauwels, A.; Brouckaert, P.; Bluethmann, H.; Fiers, W. Response of interleukin-6-deficient mice to tumor necrosis factor-induced metabolic changes and lethality. Eur. J. Immunol. 1994, 24, 2237–2242. [Google Scholar] [CrossRef]
- Sundaram, M.S.; Hemshekhar, M.; Thushara, R.M.; Santhosh, M.S.; Kumar, S.K.; Paul, M.; Devaraja, S.; Kemparaju, K.; Rangappa, K.S.; Girish, K.S. Tamarind seed extract mitigates the liver oxidative stress in arthritic rats. Food Funct. 2014, 5, 587–597. [Google Scholar] [CrossRef]
- Zhou, C.; Bai, W.; Chen, Q.; Xu, Z.; Zhu, X.; Wen, A.; Yang, X. Protective effect of crocetin against burn-induced intestinal injury. J. Surg. Res. 2015, 198, 99–107. [Google Scholar] [CrossRef]
- Thuy, S.; Ladurner, R.; Volynets, V.; Wagner, S.; Strahl, S.; Königsrainer, A.; Maier, K.P.; Bischoff, S.C.; Bergheim, I. Nonalcoholic fatty liver disease in humans is associated with increased plasma endotoxin and plasminogen activator inhibitor 1 concentrations and with fructose intake. J. Nutr. 2008, 138, 1452–1455. [Google Scholar] [CrossRef] [PubMed]
- Mota, M.; Banini, B.A.; Cazanave, S.C.; Sanyal, A.J. Molecular mechanisms of lipotoxicity and glucotoxicity in nonalcoholic fatty liver disease. Metabolism 2016, 65, 1049–1061. [Google Scholar] [CrossRef]
- Kubacka, M.; Zadrożna, M.; Nowak, B.; Kotańska, M.; Filipek, B.; Waszkielewicz, A.M.; Marona, H.; Mogilski, S. Reversal of cardiac, vascular, and renal dysfunction by non-quinazoline α1-adrenolytics in DOCA-salt hypertensive rats: A comparison with prazosin, a quinazoline-based α1-adrenoceptor antagonist. Hypertens. Res. 2019, 42, 1125–1141. [Google Scholar] [CrossRef]
- Erve, J.C.; Vashishtha, S.C.; Ojewoye, O.; Adedoyin, A.; Espina, R.; Demaio, W.; Talaat, R.E. Metabolism of prazosin in rat and characterization of metabolites in plasma, urine, faeces, brain and bile using liquid chromatography/mass spectrometry (LC/MS). Xenobiotica 2008, 38, 540–558. [Google Scholar] [CrossRef] [PubMed]
- Anglin, I.E.; Glassman, D.T.; Kyprianou, N. Induction of prostate apoptosis by alpha1-adrenoceptor antagonists: Mechanistic significance of the quinazoline component. Prostate Cancer Prostatic Dis. 2002, 5, 88–95. [Google Scholar] [CrossRef] [PubMed]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubacka, M.; Nowak, B.; Zadrożna, M.; Szafarz, M.; Latacz, G.; Marona, H.; Sapa, J.; Mogilski, S.; Bednarski, M.; Kotańska, M. Manifestations of Liver Impairment and the Effects of MH-76, a Non-Quinazoline α1-Adrenoceptor Antagonist, and Prazosin on Liver Tissue in Fructose-Induced Metabolic Syndrome. Metabolites 2023, 13, 1130. https://doi.org/10.3390/metabo13111130
Kubacka M, Nowak B, Zadrożna M, Szafarz M, Latacz G, Marona H, Sapa J, Mogilski S, Bednarski M, Kotańska M. Manifestations of Liver Impairment and the Effects of MH-76, a Non-Quinazoline α1-Adrenoceptor Antagonist, and Prazosin on Liver Tissue in Fructose-Induced Metabolic Syndrome. Metabolites. 2023; 13(11):1130. https://doi.org/10.3390/metabo13111130
Chicago/Turabian StyleKubacka, Monika, Barbara Nowak, Monika Zadrożna, Małgorzata Szafarz, Gniewomir Latacz, Henryk Marona, Jacek Sapa, Szczepan Mogilski, Marek Bednarski, and Magdalena Kotańska. 2023. "Manifestations of Liver Impairment and the Effects of MH-76, a Non-Quinazoline α1-Adrenoceptor Antagonist, and Prazosin on Liver Tissue in Fructose-Induced Metabolic Syndrome" Metabolites 13, no. 11: 1130. https://doi.org/10.3390/metabo13111130