

Comparative Insights into Four Major Legume Sprouts Efficacies for Diabetes Management and Its Complications: Untargeted versus Targeted NMR Biochemometrics Approach
 , ,
, ,  and
and 
Abstract
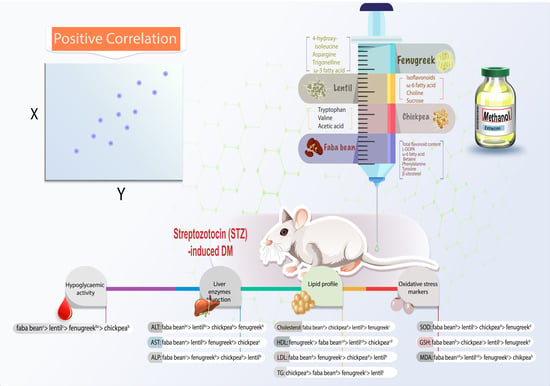
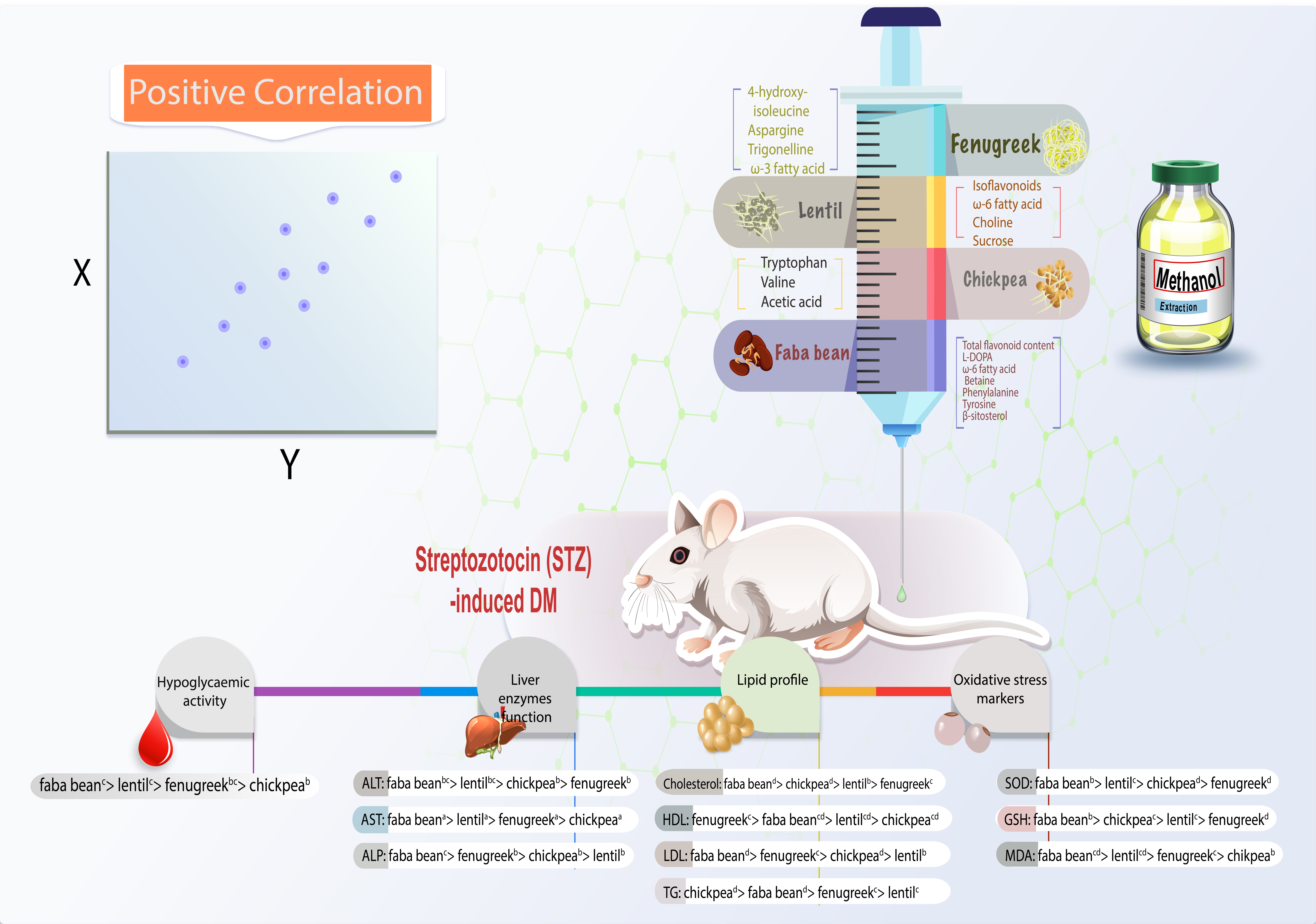
:
1. Introduction
2. Materials and Methods
2.1. Plant Material and Chemicals
2.2. Sprouting Method
2.3. Extraction, Metabolites Profiling, and Fingerprinting
2.4. Animals and Diabetes Induction
2.5. Hypoglycemic Activity Evaluation
2.5.1. Experimental Design
2.5.2. Sample Preparation
2.5.3. Determination of Blood Glucose Level
2.6. Effect of Different Treatments of Sprouts on Diabetes Complications
2.6.1. Assessment of Liver Functions
- Determination of liver function enzymes
2.6.2. Assessment on Lipid Metabolism
- Determination of Cholesterol Level
- Determination of high-density lipoprotein cholesterol (HDL-C)
- Determination of low-density lipoprotein cholesterol (LDL-C)
- Determination of serum triglycerides (TG)
2.6.3. Effect on Oxidative Stress Markers
- Determination of superoxide dismutase activity
- Determination of glutathione level
- Determination of malondialdehyde level
2.7. Determination of Total Protein in Tissue Homogenate
2.8. Histopathological Examination of Liver Tissues
2.9. Statistical and Multivariate Data Analysis
3. Results and Discussion
3.1. Antihyperglycemic Activity
3.2. Management of Diabetes Complications
3.2.1. Liver Enzymes
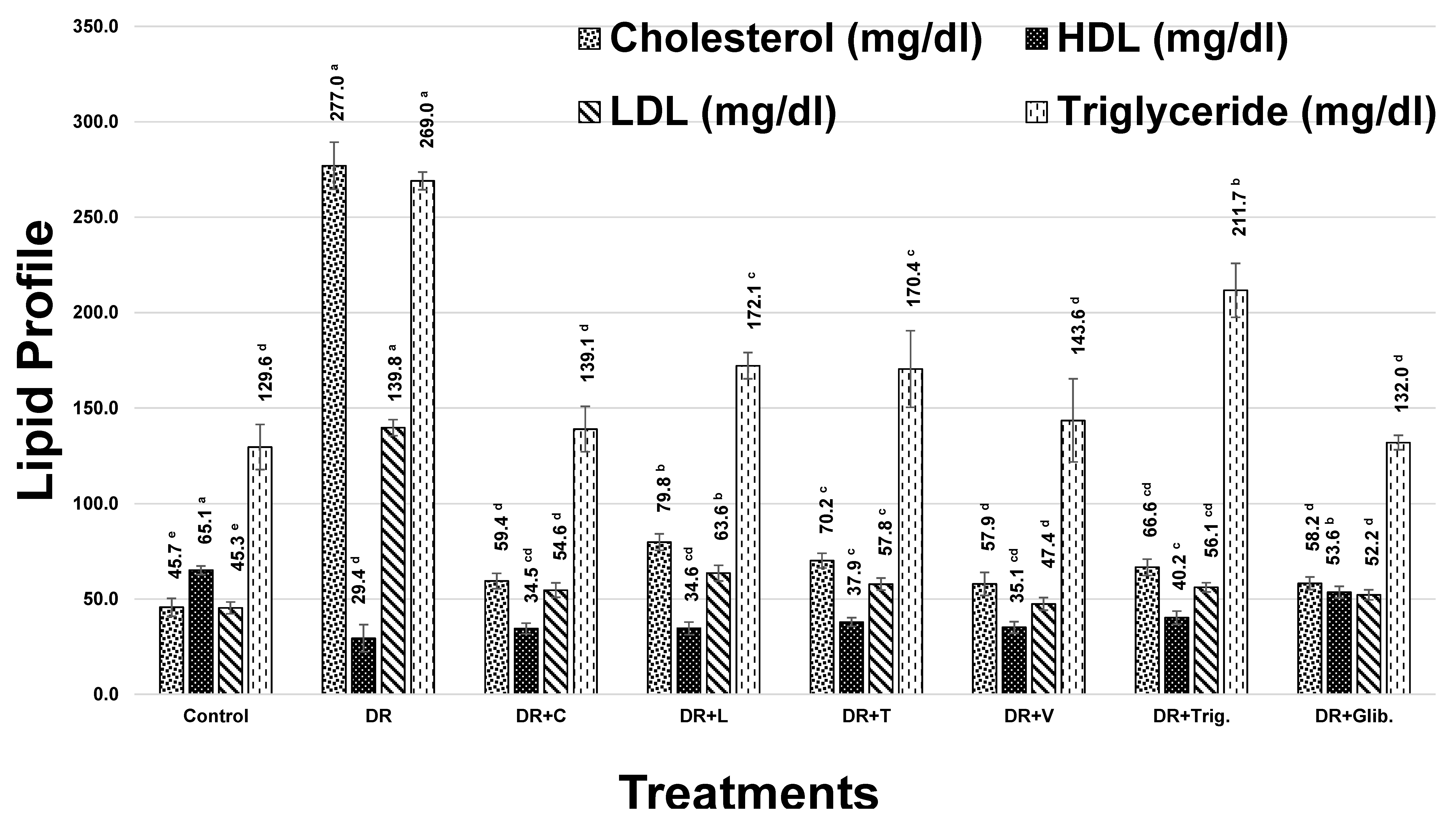
3.2.2. Lipid Profile
3.2.3. Liver Oxidative Stress Markers
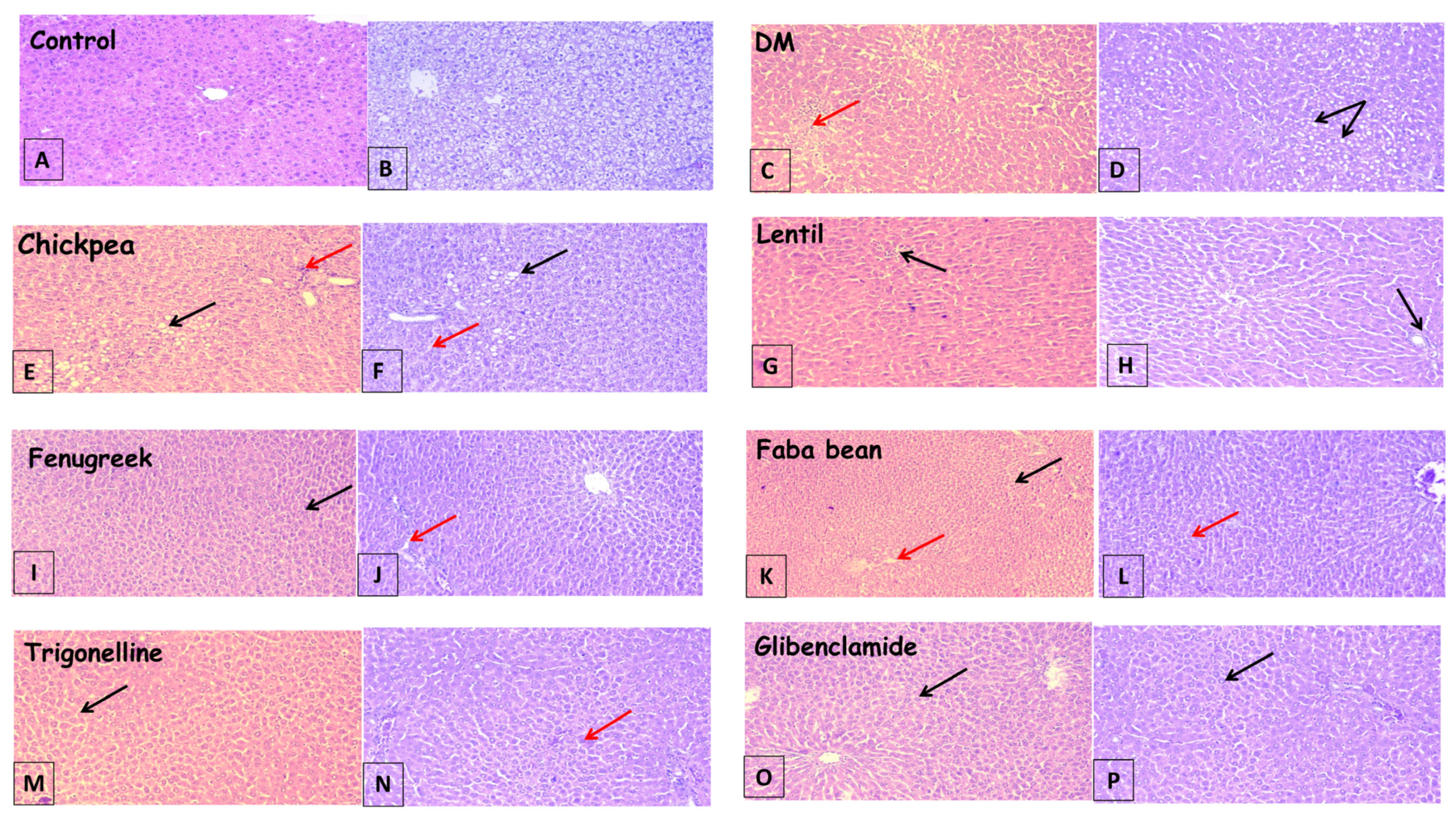
3.3. Histopathological Findings
3.4. Bioactivities in Relationship with Major Metabolites in Legume Sprouts
3.4.1. Regression Analysis
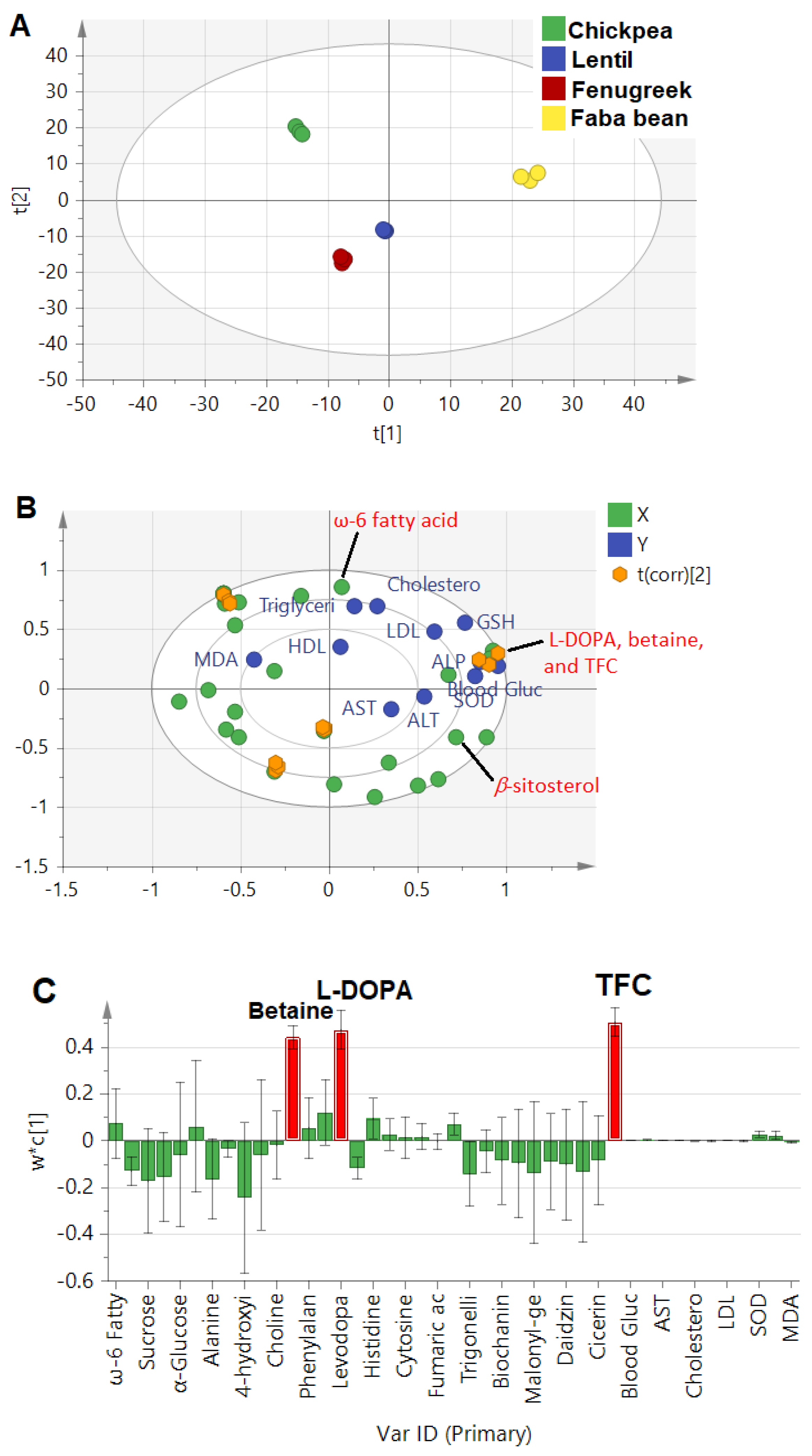
3.4.2. Partial Least Square Analysis (PLS)
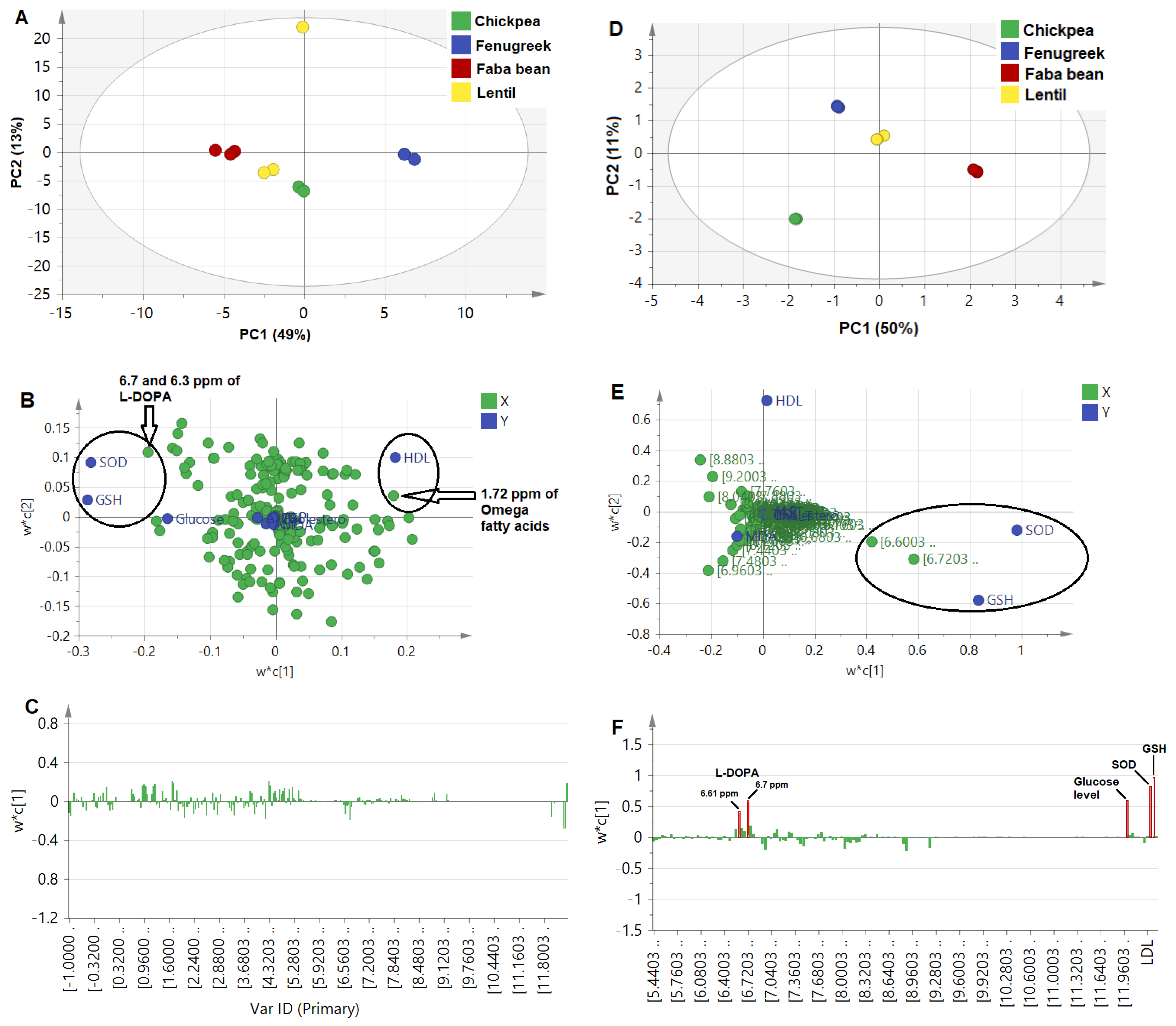
- Untargeted PLS approach
- Targeted PLS approach
4. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vatanparast, M.; Powell, A.; Doyle, J.J.; Egan, A.N. Targeting legume loci: A comparison of three methods for target enrichment bait design in Leguminosae phylogenomics. Appl. Plant Sci. 2018, 6, e1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutjaritjai, N.; Wangpakapattanawong, P.; Balslev, H.; Inta, A. Traditional uses of Leguminosae among the Karen in Thailand. Plants 2019, 8, 600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tungmunnithum, D.; Drouet, S.; Lorenzo, J.M.; Hano, C. Characterization of bioactive phenolics and antioxidant capacity of edible bean extracts of 50 Fabaceae populations grown in Thailand. Foods 2021, 10, 3118. [Google Scholar] [CrossRef] [PubMed]
- Khrisanapant, P.; Kebede, B.; Leong, S.Y.; Oey, I. A comprehensive characterisation of volatile and fatty acid profiles of legume seeds. Foods 2019, 8, 651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devi, C.; Kushwaha, A.; Kumar, A. Sprouting characteristics and associated changes in nutritional composition of cowpea (Vigna unguiculata). J. Food Sci. Technol. 2015, 52, 6821–6827. [Google Scholar] [CrossRef]
- Mohd Ali, N.; Mohd Yusof, H.; Long, K.; Yeap, S.K.; Ho, W.Y.; Beh, B.K.; Koh, S.P.; Abdullah, M.P.; Alitheen, N.B. Antioxidant and hepatoprotective effect of aqueous extract of germinated and fermented mung bean on ethanol-mediated liver damage. BioMed Res. Int. 2013, 2013, 693613. [Google Scholar] [CrossRef]
- Farag, M.; Sharaf El-Din, M.; Aboul-Fotouh Selim, M.; Owis, A.; Abouzid, S. Mass spectrometry-based metabolites profiling of nutrients and anti-nutrients in major legume sprouts. Food Biosci. 2021, 39, 100800. [Google Scholar] [CrossRef]
- Farag, M.A.; Sharaf El-Din, M.G.; Selim, M.A.; Owis, A.I.; Abouzid, S.F.; Porzel, A.; Wessjohann, L.A.; Otify, A. Nuclear magnetic resonance metabolomics approach for the analysis of major legume sprouts coupled to chemometrics. Molecules 2021, 26, 761. [Google Scholar] [CrossRef]
- Vijayakumar, M.V.; Singh, S.; Chhipa, R.R.; Bhat, M.K. The hypoglycaemic activity of fenugreek seed extract is mediated through the stimulation of an insulin signalling pathway. Br. J. Pharmacol. 2005, 146, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Alberti, K.G.M.M.; Zimmet, P.Z. Definition, diagnosis and classification of diabetes mellitus and its complications. Part 1: Diagnosis and classification of diabetes mellitus. Provisional report of a WHO consultation. Diabet. Med. 1998, 15, 539–553. [Google Scholar] [CrossRef]
- Yazdi, H.B.; Hojati, V.; Shiravi, A.; Hosseinian, S.; Vaezi, G.; Hadjzadeh, M.A. Liver Dysfunction and Oxidative Stress in Streptozotocin-Induced Diabetic Rats: Protective Role of Artemisia Turanica. J. Pharmacopuncture 2019, 22, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Mertens, J.; De Block, C.; Spinhoven, M.; Driessen, A.; Francque, S.M.; Kwanten, W.J. Hepatopathy associated with type 1 diabetes: Distinguishing non-alcoholic fatty liver disease From glycogenic hepatopathy. Front. Pharmacol. 2021, 12, 768576. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, A.K.; Sahana, C.; Zehra, A.; Madhusudana, K.; Kumar, D.A.; Agawane, S.B. Mitigation of starch-induced postprandial glycemic spikes in rats by antioxidants-rich extract of Cicer arietinum Linn. seeds and sprouts. J. Pharm. Bioallied Sci. 2013, 5, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Tefera, M.M.; Altaye, B.M.; Yimer, E.M.; Berhe, D.F.; Tadesse Bekele, S. Antidiabetic effect of germinated Lens culinaris Medik seed extract in streptozotocin-induced diabetic mice. J. Exp. Pharmacol. 2020, 12, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Zhou, S.; Zeng, S. Experimental diabetes treated with trigonelline: Effect on β cell and pancreatic oxidative parameters. Fundam. Clin. Pharmacol. 2013, 27, 279–287. [Google Scholar] [CrossRef]
- Lv, Q.; Yang, Y.; Zhao, Y.; Gu, D.; He, D.; Yili, A.; Ma, Q.; Cheng, Z.; Gao, Y.; Aisa, H.A. Comparative study on separation and purification of isoflavones from the seeds and sprouts of chickpea by high-speed countercurrent chromatography. J. Liq. Chrom. Relat. Tech. 2009, 32, 2879–2892. [Google Scholar] [CrossRef] [Green Version]
- Farag, M.A.; Porzel, A.; Wessjohann, L.A. Comparative metabolite profiling and fingerprinting of medicinal licorice roots using a multiplex approach of GC-MS, LC-MS and 1D NMR techniques. Phytochemistry 2012, 76, 60–72. [Google Scholar] [CrossRef]
- Farag, M.A.; Porzel, A.; Schmidt, J.; Wessjohann, L.A. Metabolite profiling and fingerprinting of commercial cultivars of Humulus lupulus L. (hop): A comparison of MS and NMR methods in metabolomics. Metabolomics 2012, 8, 492–507. [Google Scholar] [CrossRef]
- Punithavathi, V.; Anuthama, R.; Prince, P. Combined treatment with naringin and vitamin C ameliorates streptozotocin-induced diabetes in male Wistar rats. J. Appl. Toxicol. 2008, 28, 806–813. [Google Scholar] [CrossRef]
- Bhandari, U.; Pillai, K. Effect of ethanolic extract of Zingiber officinale on dyslipidaemia in diabetic rats. J. Ethnopharmacol. 2005, 97, 227–230. [Google Scholar] [CrossRef]
- Vijayakumar, M.V.; Bhat, M.K. Hypoglycemic effect of a novel dialysed fenugreek seeds extract is sustainable and is mediated, in part, by the activation of hepatic enzymes. Phytother. Res. 2008, 22, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Liang, B.; Li, Y. Antihyperglycemic effect of Ginkgo biloba extract in streptozotocin-induced diabetes in rats. BioMed Res. Int. 2012, 2013, 162724. [Google Scholar] [PubMed] [Green Version]
- Cronin, D.; Smith, S. A simple and rapid procedure for the analysis of reducing, total and individual sugars in potatoes. Potato Res. 1979, 22, 99–105. [Google Scholar] [CrossRef]
- Reitman, S.; Frankel, S. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Thomas, L. Clinical Laboratory Diagnostics: Use and Assessment of Clinical Laboratory Results; TH-Books Verlagsgesellschaft: Frankfurt, Germany, 1998. [Google Scholar]
- Meiattini, F.; Prencipe, L.; Bardelli, F.; Giannini, G.; Tarli, P. The 4-hydroxybenzoate/4-aminophenazone chromogenic system used in the enzymic determination of serum cholesterol. Clin. Chem. 1978, 24, 2161–2165. [Google Scholar] [CrossRef]
- Burstein, M.; Scholnick, H.; Morfin, R. Rapid method for the isolation of lipoproteins from human serum by precipitation with polyanions. J. Lipid Res. 1970, 11, 583–595. [Google Scholar] [CrossRef]
- Assmann, G.; Jabs, H.-U.; Kohnert, U.; Nolte, W.; Schriewer, H. LDL-cholesterol determination in blood serum following precipitation of LDL with polyvinylsulfate. Clin. Chim. Acta 1984, 140, 77–83. [Google Scholar] [CrossRef]
- Fossati, P.; Prencipe, L. Serum triglycerides determined colorimetrically with an enzyme that produces hydrogen peroxide. Clin. Chem. 1982, 28, 2077–2080. [Google Scholar] [CrossRef]
- Nishikimi, M.; Rao, N.A.; Yagi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [CrossRef]
- Moron, M.S.; Depierre, J.W.; Mannervik, B. Levels of glutathione, glutathione reductase and glutathione S-transferase activities in rat lung and liver. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1979, 582, 67–78. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, C.; Zouain, C.; Alves, J.; Goes, A. Induction of protective immunity and modulation of granulomatous hypersensitivity in mice using PIII, an anionic fraction of Schistosoma mansoni adult worm. Parasitology 1997, 115, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Motawi, T.K.; Darwish, H.A.; Hamed, M.A.; El-Rigal, N.S.; Naser, A.F.A. A Therapeutic insight of niacin and coenzyme Q10 against diabetic encephalopathy in rats. Mol. Neurobiol. 2017, 54, 1601–1611. [Google Scholar] [CrossRef]
- Farag, M.A.; Porzel, A.; Wessjohann, L.A. Unraveling the active hypoglycemic agent trigonelline in Balanites aegyptiaca date fruit using metabolite fingerprinting by NMR. J. Pharm. Biomed. Anal. 2015, 115, 383–387. [Google Scholar] [CrossRef]
- Subramanian, S.P.; Prasath, G.S. Antidiabetic and antidyslipidemic nature of trigonelline, a major alkaloid of fenugreek seeds studied in high-fat-fed and low-dose streptozotocin-induced experimental diabetic rats. Biomed. Prev. Nutr. 2014, 4, 475–480. [Google Scholar] [CrossRef]
- Yang, L.; Gao, Y.; Bajpai, V.K.; El-Kammar, H.A.; Simal-Gandara, J.; Cao, H.; Cheng, K.W.; Wang, M.; Arroo, R.R.J.; Zou, L.; et al. Advance toward isolation, extraction, metabolism and health benefits of kaempferol, a major dietary flavonoid with future perspectives. Crit. Rev. Food Sci. Nutr. 2021. [Google Scholar] [CrossRef]
- Gupta, R.K.; Patel, A.K.; Shah, N.; Chaudhary, A.K.; Jha, U.K.; Yadav, U.C.; Gupta, P.K.; Pakuwal, U. Oxidative stress and antioxidants in disease and cancer: A review. Asian Pac. J. Cancer Prev. 2014, 15, 4405–4409. [Google Scholar] [CrossRef] [Green Version]
- Safhi, M.M.; Alam, M.F.; Sivakumar, S.M.; Anwer, T. Hepatoprotective potential of Sargassum muticum against STZ-induced diabetic liver damage in wistar rats by inhibiting cytokines and the apoptosis pathway. Anal. Cell. Pathol. 2019, 2019, 7958701. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Tang, C.; Zhang, H. Hepatoprotective effects of kaempferol 3-O-rutinoside and kaempferol 3-O-glucoside from Carthamus tinctorius L. on CCl(4)-induced oxidative liver injury in mice. J. Food Drug Anal. 2015, 23, 310–317. [Google Scholar] [CrossRef]
- Bhowmik, B.; Siddiquee, T.; Mujumder, A.; Afsana, F.; Ahmed, T.; Mdala, I.A.; do V Moreira, N.C.; Khan, A.K.A.; Hussain, A.; Holmboe-Ottesen, G.; et al. Serum lipid profile and its association with diabetes and prediabetes in a rural Bangladeshi population. Int. J. Environ. Res. Public Health 2018, 15, 1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farid, M.M.; Aboul Naser, A.F.; Salem, M.M.; Ahmed, Y.R.; Emam, M.; Hamed, M.A. Chemical compositions of Commiphora opobalsamum stem bark to alleviate liver complications in streptozotocin-induced diabetes in rats: Role of oxidative stress and DNA damage. Biomarkers 2022, 27, 671–683. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-F.; Zhang, F.; Zhang, J.; Zhang, R.-M.; Li, R. Protection effect of trigonelline on liver of rats with non-alcoholic fatty liver diseases. Asian Pac. J. Trop. Med. 2015, 8, 651–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibarra, A.; He, K.; Bai, N.; Bily, A.; Roller, M.; Coussaert, A.; Provost, N.; Ripoll, C. Fenugreek extract rich in 4-hydroxyisoleucine and trigonelline activates PPARα and inhibits LDL oxidation: Key mechanisms in controlling the metabolic syndrome. Nat. Prod. Commun. 2008, 3, 1934578X0800300923. [Google Scholar] [CrossRef] [Green Version]
- Ochiai, A.; Miyata, S.; Iwase, M.; Shimizu, M.; Inoue, J.; Sato, R. Kaempferol stimulates gene expression of low-density lipoprotein receptor through activation of Sp1 in cultured hepatocytes. Sci. Rep. 2016, 6, 24940. [Google Scholar] [CrossRef] [Green Version]
- Skrapari, I.; Perrea, D.; Ioannidis, I.; Karabina, S.A.; Elisaf, M.; Tselepis, A.D.; Karagiannacos, P.; Katsilambros, N. Glibenclamide improves postprandial hypertriglyceridaemia in type 2 diabetic patients by reducing chylomicrons but not the very low-density lipoprotein subfraction levels. Diabet. Med. 2001, 18, 781–785. [Google Scholar] [CrossRef]
- Sarian, M.N.; Ahmed, Q.U.; Mat So’ad, S.Z.; Alhassan, A.M.; Murugesu, S.; Perumal, V.; Syed Mohamad, S.N.A.; Khatib, A.; Latip, J. Antioxidant and antidiabetic effects of flavonoids: A structure-activity relationship based study. BioMed Res. Int. 2017, 2017, 8386065. [Google Scholar] [CrossRef]
- Jubaidi, F.F.; Zainalabidin, S.; Taib, I.S.; Hamid, Z.A.; Budin, S.B. The potential role of flavonoids in ameliorating diabetic cardiomyopathy via alleviation of cardiac oxidative stress, inflammation and apoptosis. Int. J. Mol. Sci. 2021, 22, 5094. [Google Scholar] [CrossRef]
- Unuofin, J.O.; Lebelo, S.L. Antioxidant effects and mechanisms of medicinal plants and their bioactive compounds for the prevention and treatment of type 2 diabetes: An updated review. Oxid. Med. Cell. Longev. 2020, 2020, 1356893. [Google Scholar] [CrossRef] [Green Version]
- Erejuwa, O.O.; Sulaiman, S.A.; Wahab, M.S.; Salam, S.K.; Salleh, M.S.; Gurtu, S. Antioxidant protective effect of glibenclamide and metformin in combination with honey in pancreas of streptozotocin-induced diabetic rats. Int. J. Mol. Sci. 2010, 11, 2056–2066. [Google Scholar] [CrossRef]
- Rabbani, S.I.; Devi, K.; Khanam, S. Protective role of glibenclamide against nicotinamide-streptozotocin induced nuclear damage in diabetic Wistar rats. J. Pharmacol. Pharmacother. 2010, 1, 18–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zang, Y.; Zhang, D.; Yu, C.; Jin, C.; Igarashi, K. Antioxidant and hepatoprotective activity of kaempferol 3-O-β-d-(2,6-di-O-α-l-rhamnopyranosyl)galactopyronoside against carbon tetrachloride-induced liver injury in mice. Food Sci. Biotechnol. 2017, 26, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Randhir, R.; Shetty, K.J.P.B. Microwave-induced stimulation of L-DOPA, phenolics and antioxidant activity in fava bean (Vicia faba) for Parkinson’s diet. Process Biochem. 2004, 39, 1775–1784. [Google Scholar] [CrossRef]
- Farag, M.A.; Khaled, S.E.; El Gingeehy, Z.; Shamma, S.N.; Zayed, A. Comparative metabolite profiling and fingerprinting of medicinal cinnamon bark and its commercial preparations via a multiplex approach of GC-MS, UV, and NMR techniques. Metabolites 2022, 12, 614. [Google Scholar] [CrossRef]
- Mehring, A.; Zayed, A.; Salem, M.A.; Alseekh, S.; Fernie, A.R.; Ulber, R. Time-dependent behaviour of methyl jasmonate elicited cell suspension cultures of Ocimum species via untargeted mass spectrometry-based metabolomics. Ind. Crops Prod. 2022, 187, 115414. [Google Scholar] [CrossRef]
- Khaled, S.E.; Hashem, F.A.M.; Shabana, M.H.; Hammam, A.-M.M.; Madboli, A.N.A.; Al-Mahdy, D.A.; Farag, M.A. A biochemometric approach for the assessment of Phyllanthus emblica female fertility effects as determined via UPLC-ESI-qTOF-MS and GC-MS. Food Funct. 2019, 10, 4620–4635. [Google Scholar] [CrossRef]
- Coskun, O.; Kanter, M.; Korkmaz, A.; Oter, S. Quercetin, a flavonoid antioxidant, prevents and protects streptozotocin-induced oxidative stress and β-cell damage in rat pancreas. Pharmacol. Res. 2005, 51, 117–123. [Google Scholar] [CrossRef]
- Feng, R.N.; Niu, Y.C.; Sun, X.W.; Li, Q.; Zhao, C.; Wang, C.; Guo, F.C.; Sun, C.H.; Li, Y. Histidine supplementation improves insulin resistance through suppressed inflammation in obese women with the metabolic syndrome: A randomised controlled trial. Diabetologia 2013, 56, 985–994. [Google Scholar] [CrossRef]
- Aware, C.; Patil, R.; Gaikwad, S.; Yadav, S.; Bapat, V.; Jadhav, J. Evaluation of l-dopa, proximate composition with in vitro anti-inflammatory and antioxidant activity of Mucuna macrocarpa beans: A future drug for Parkinson treatment. Asian Pac. J. Trop. Biomed. 2017, 7, 1097–1106. [Google Scholar] [CrossRef]
- Arumugam, M.K.; Paal, M.C.; Donohue, T.M., Jr.; Ganesan, M.; Osna, N.A.; Kharbanda, K.K. Beneficial effects of betaine: A comprehensive review. Biology 2021, 10, 456. [Google Scholar] [CrossRef]
- Wang, L.; Chen, L.; Tan, Y.; Wei, J.; Chang, Y.; Jin, T.; Zhu, H. Betaine supplement alleviates hepatic triglyceride accumulation of apolipoprotein E deficient mice via reducing methylation of peroxisomal proliferator-activated receptor alpha promoter. Lipids Health Dis. 2013, 12, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holeček, M. Histidine in health and disease: Metabolism, physiological importance, and use as a supplement. Nutrients 2020, 12, 848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.-H.; Liu, T.-C.; Yin, M.-C. Beneficial effects of histidine and carnosine on ethanol-induced chronic liver injury. Food Chem. Toxicol. 2008, 46, 1503–1509. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.Y.; Chen, Y.X.; Fang, M.; Zhu, X.L.; Zhao, Y.X.; Liu, X.Y. Low-dose levodopa protects nerve cells from oxidative stress and up-regulates expression of pCREB and CD39. PLoS ONE 2014, 9, e95387. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Glucose Level | ALT | AST | ALP | ||||
|---|---|---|---|---|---|---|---|---|
| Glucose (mg/dL) | % Improvement | ALT (Unit/L) | % Improvement | AST (Unit/L) | % Improvement | ALP (Unit/L) | % Improvement | |
| Control | 74.6 e ± 2.4 | --- | 1.5 c ± 0.1 | --- | 2.2 b ± 0.2 | --- | 258.5 d ± 11.0 | --- |
| DR | 399.2 a ± 30.9 | --- | 2.3 a ± 0.3 | --- | 3.6 a ± 0.3 | --- | 433.2 a ± 16.8 | --- |
| DR + C | 304.9 b ± 36.4 | 126.5 | 1.9 b ± 0.1 | 24.3 | 3.5 a ± 0.4 | 6.3 | 323.8 b ± 5.5 | 42.3 |
| DR + L | 254.4 c ± 26.6 | 194.2 | 1.7 bc ± 0.1 | 35.1 | 3.2 a ±0.6 | 20.5 | 329.7 b ± 4.4 | 40.0 |
| DR + F | 292.6 bc ± 25.4 | 143.1 | 1.9 b ± 0.2 | 22.3 | 3.3 a ± 0.2 | 14.3 | 318.9 b ± 8.9 | 44.6 |
| DR + V | 139.6 d ± 11.4 | 348.0 | 1.7 bc±0.1 | 36.5 | 3.1 a ± 0.1 | 24.1 | 278.6 c ± 9.2 | 59.8 |
| DR + Trig. | 280.5 bc ± 32.0 | 159.1 | 1.8 b ± 0.2 | 31.8 | 3.5 a ± 0.4 | 4.5 | 317.1 b ± 3.3 | 44.9 |
| DR + Glib. | 127.4 d ± 2.5 | 364.3 | 1.6 bc ± 0.11 | 43.2 | 2.4 b ± 0.2 | 55.4 | 273.0 c ± 21.3 | 62.0 |
| Group | SOD | GSH | MDA | |||
|---|---|---|---|---|---|---|
| SOD Level (µg/mg Protein) | % Improvement | GSH Level (µg/g Tissue) | % Improvement | MDA (µmol/mg Protein) | % Improvement | |
| Control | 28.4 a ± 1.4 | --- | 26.0 a ± 2.0 | --- | 1.1 d ± 0.2 | --- |
| DR | 9.7 e ± 0.8 | --- | 9.2 d ± 0.7 | --- | 3.1 a ± 0.5 | --- |
| DR + C | 12.4 d ± 1.6 | 9.4 | 12.7 c ± 1.1 | 13.5 | 2.0 b ± 0.2 | 94.6 |
| DR + L | 15.5 c ± 1.1 | 20.5 | 12.3 c ± 0.7 | 11.7 | 1.6 cd ± 0.2 | 133.6 |
| DR + F | 11.8 d ± 1.0 | 7.5 | 10.3 d ± 0.6 | 4.2 | 1.7 c ± 0.7 | 123.0 |
| DR + V | 17.5 b ± 1.7 | 27.6 | 15.8 b ± 0.6 | 25.4 | 1.5 cd ± 0.2 | 144.2 |
| DR + Trig. | 11.2 de ± 1.0 | 5.3 | 13.8 c ± 0.6 | 17.7 | 1.5 cd ± 0.2 | 143.4 |
| DR + Glib. | 14.7 c ± 0.7 | 17.5 | 16.4 b ± 1.6 | 27.7 | 1.3 cd ± 0.2 | 156.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farag, M.A.; Aboul Naser, A.F.; Zayed, A.; Sharaf El-Dine, M.G. Comparative Insights into Four Major Legume Sprouts Efficacies for Diabetes Management and Its Complications: Untargeted versus Targeted NMR Biochemometrics Approach. Metabolites 2023, 13, 63. https://doi.org/10.3390/metabo13010063
Farag MA, Aboul Naser AF, Zayed A, Sharaf El-Dine MG. Comparative Insights into Four Major Legume Sprouts Efficacies for Diabetes Management and Its Complications: Untargeted versus Targeted NMR Biochemometrics Approach. Metabolites. 2023; 13(1):63. https://doi.org/10.3390/metabo13010063
Chicago/Turabian StyleFarag, Mohamed A., Asmaa F. Aboul Naser, Ahmed Zayed, and Mohamed G. Sharaf El-Dine. 2023. "Comparative Insights into Four Major Legume Sprouts Efficacies for Diabetes Management and Its Complications: Untargeted versus Targeted NMR Biochemometrics Approach" Metabolites 13, no. 1: 63. https://doi.org/10.3390/metabo13010063