
Obstructive Sleep Apnea, Circadian Clock Disruption, and Metabolic Consequences

 ,
,  , and
, and 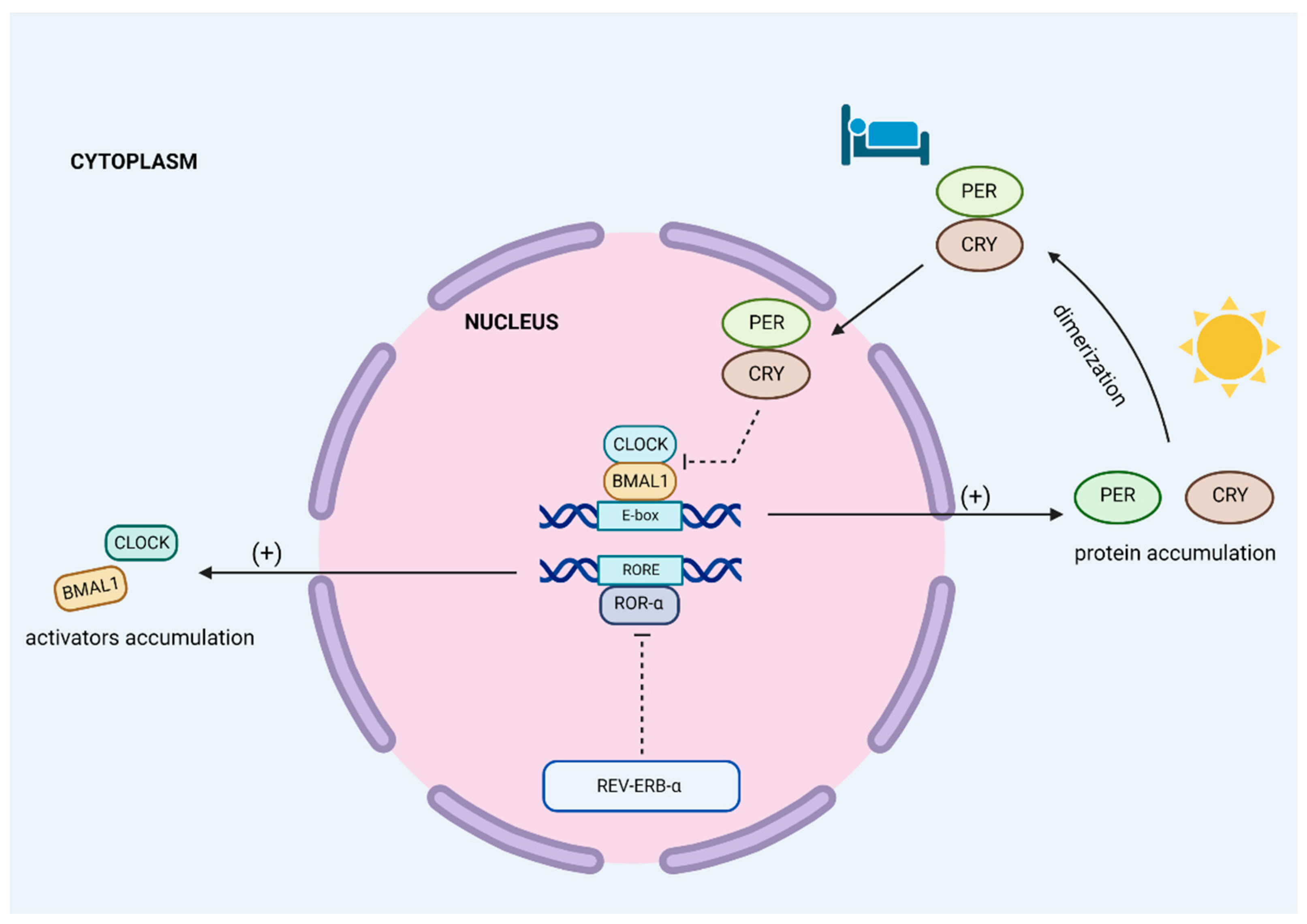{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
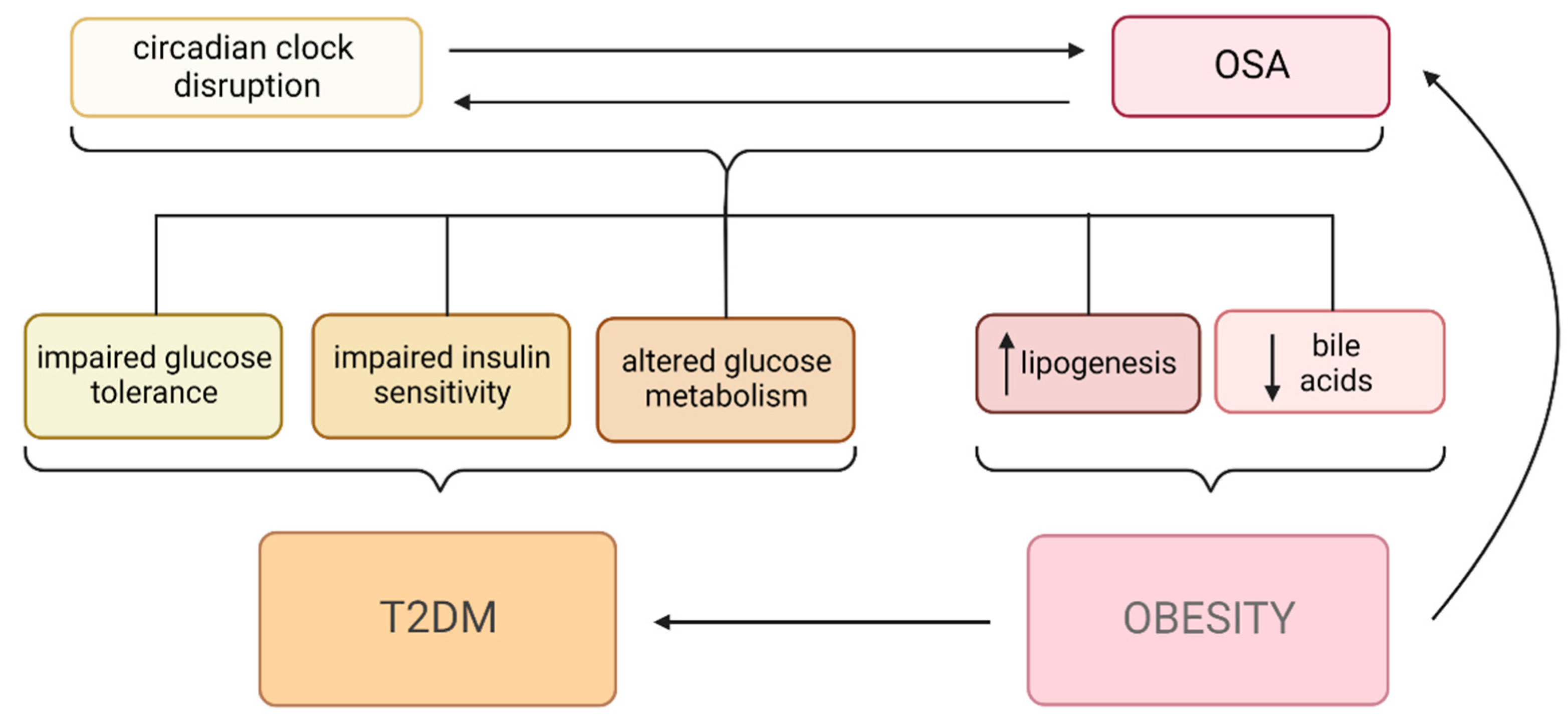
2. Metabolic Changes in OSA
2.1. Diabetes Mellitus and OSA
2.2. Obesity and OSA
3. Circadian Clock Organization
4. Circadian Clock Disruption in the Context of Metabolic Changes
4.1. General Impact of Circadian Clock Disruption on Metabolism
4.2. Glucose Metabolism
4.3. Lipid Metabolism
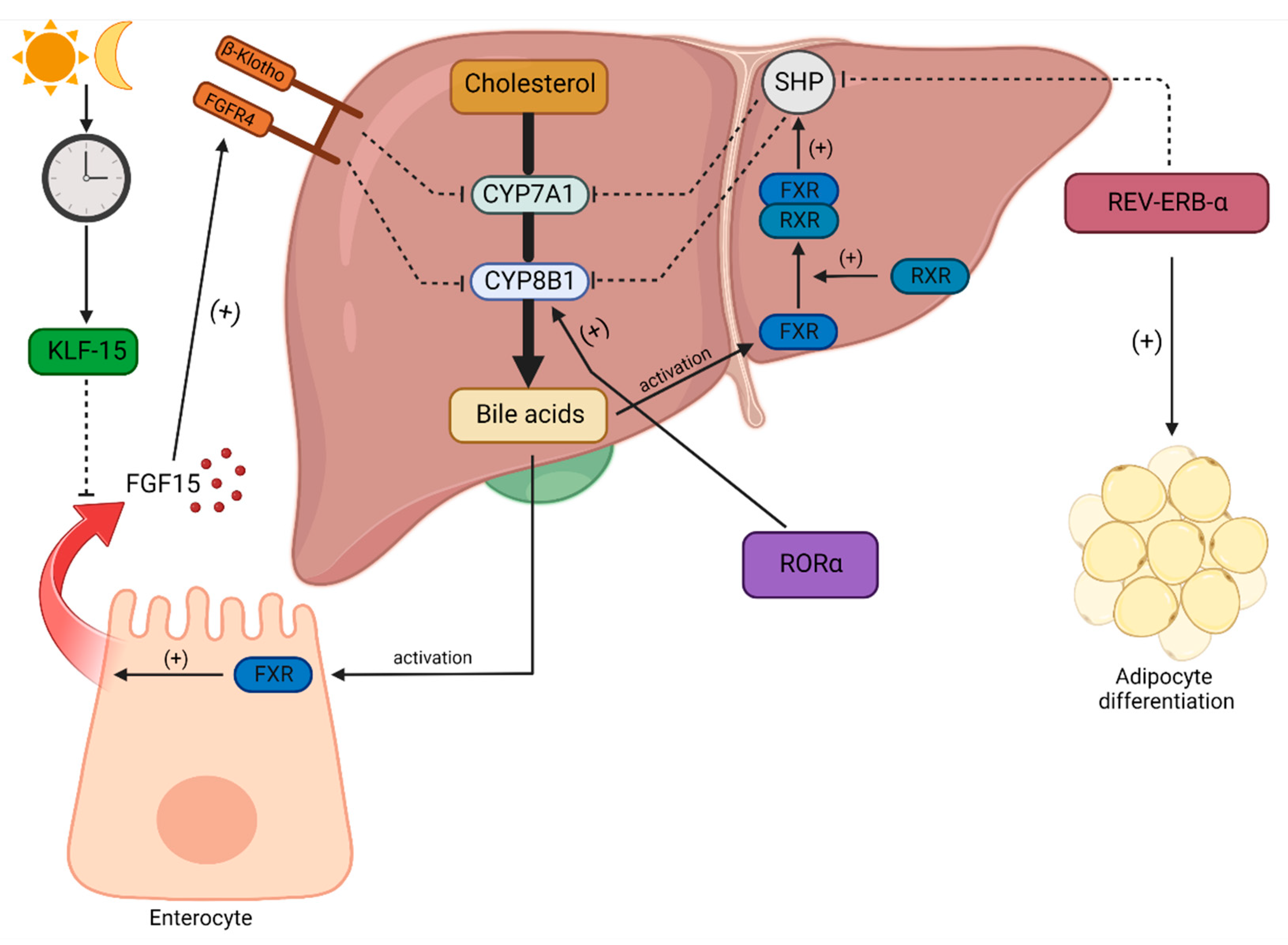
4.4. Bile Acids
4.5. NAD+
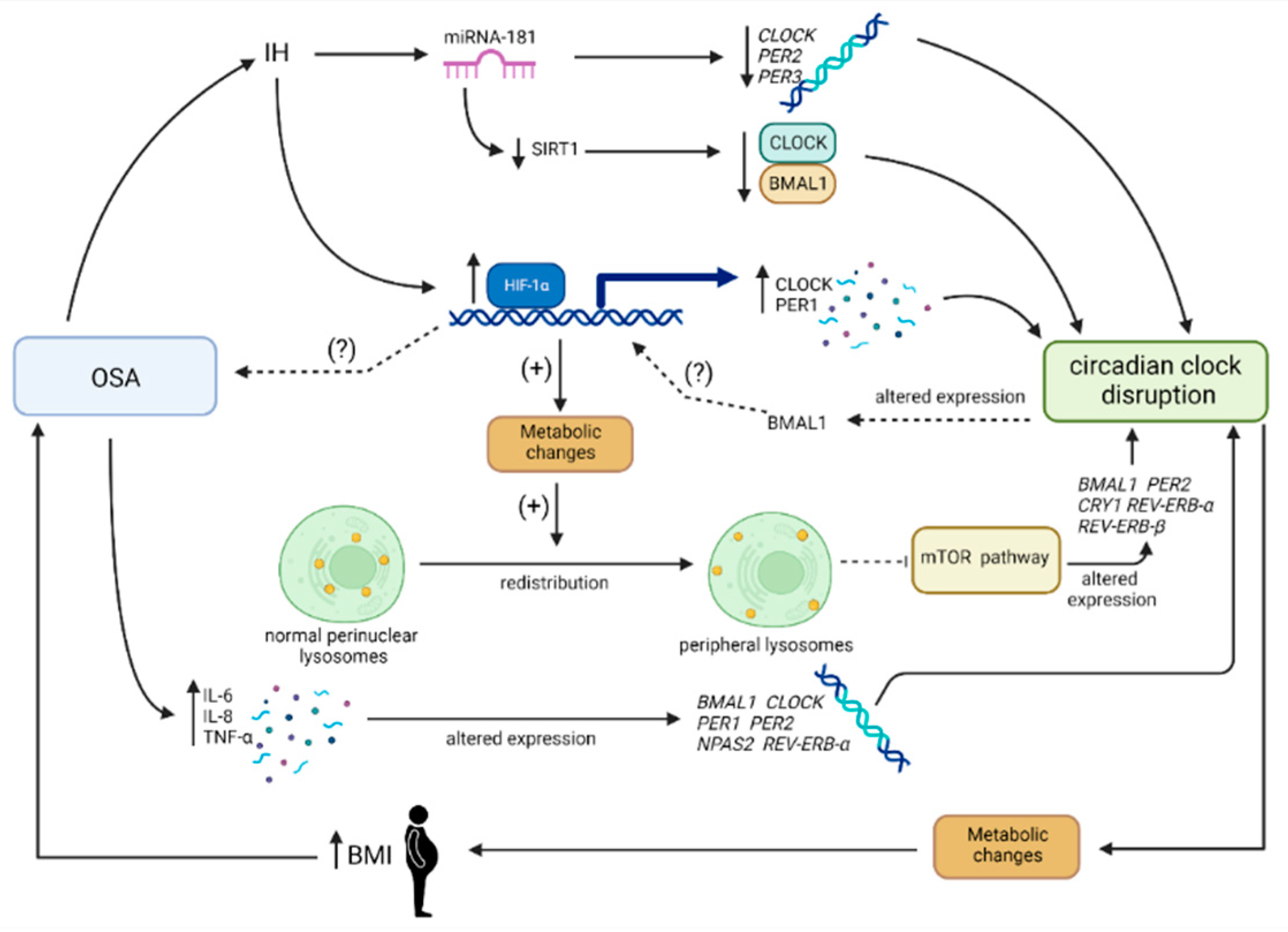
5. OSA and Circadian Clock Disruption
6. Limitation of the Study
7. Future Perspectives
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rundo, J.V. Obstructive Sleep Apnea Basics. Cleve. Clin. J. Med. 2019, 86, 2–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.R. Obstructive Sleep Apnea. Ann. Intern. Med. 2019, 171, ITC81–ITC96. [Google Scholar] [CrossRef] [PubMed]
- Semelka, M.; Wilson, J.; Floyd, R. Diagnosis and Treatment of Obstructive Sleep Apnea in Adults. Am. Fam. Physician 2016, 94, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Young, T.; Evans, L.; Finn, L.; Palta, M. Estimation of the Clinically Diagnosed Proportion of Sleep Apnea Syndrome in Middle-Aged Men and Women. Sleep 1997, 20, 705–706. [Google Scholar] [CrossRef]
- Punjabi, N.M. The Epidemiology of Adult Obstructive Sleep Apnea. Proc. Am. Thorac. Soc. 2008, 5, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Heinzer, R.; Vat, S.; Marques-Vidal, P.; Marti-Soler, H.; Andries, D.; Tobback, N.; Mooser, V.; Preisig, M.; Malhotra, A.; Waeber, G.; et al. Prevalence of Sleep-Disordered Breathing in the General Population: The HypnoLaus Study. Lancet. Respir. Med. 2015, 3, 310–318. [Google Scholar] [CrossRef] [Green Version]
- Senaratna, C.V.; Perret, J.L.; Lodge, C.J.; Lowe, A.J.; Campbell, B.E.; Matheson, M.C.; Hamilton, G.S.; Dharmage, S.C. Prevalence of Obstructive Sleep Apnea in the General Population: A Systematic Review. Sleep Med. Rev. 2017, 34, 70–81. [Google Scholar] [CrossRef]
- Monahan, K.; Redline, S. Role of Obstructive Sleep Apnea in Cardiovascular Disease. Curr. Opin. Cardiol. 2011, 26, 541–547. [Google Scholar] [CrossRef] [Green Version]
- Dredla, B.K.; Castillo, P.R. Cardiovascular Consequences of Obstructive Sleep Apnea. Curr. Cardiol. Rep. 2019, 21, 137. [Google Scholar] [CrossRef]
- Gonzaga, C.; Bertolami, A.; Bertolami, M.; Amodeo, C.; Calhoun, D. Obstructive Sleep Apnea, Hypertension and Cardiovascular Diseases. J. Hum. Hypertens. 2015, 29, 705–712. [Google Scholar] [CrossRef]
- Mckee, Z.; Auckley, D.H. A Sleeping Beast: Obstructive Sleep Apnea and Stroke. Cleve. Clin. J. Med. 2019, 86, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Yaggi, H.K.; Concato, J.; Kernan, W.N.; Lichtman, J.H.; Brass, L.M.; Mohsenin, V. Obstructive Sleep Apnea as a Risk Factor for Stroke and Death. N. Engl. J. Med. 2005, 353, 2034–2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, S.O.; He, K.; Narla, R.R.; Kang, H.G.; Ryu, H.U.; Boyko, E.J. Metabolic Consequences of Obstructive Sleep Apnea Especially Pertaining to Diabetes Mellitus and Insulin Sensitivity. Diabetes Metab. J. 2019, 43, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, X.; Lu, Y. Obstructive Sleep Apnea Syndrome and Metabolic Diseases. Endocrinology 2018, 159, 2670–2675. [Google Scholar] [CrossRef] [Green Version]
- Gabryelska, A.; Karuga, F.F.; Szmyd, B.; Białasiewicz, P. HIF-1α as a Mediator of Insulin Resistance, T2DM, and Its Complications: Potential Links with Obstructive Sleep Apnea. Front. Physiol. 2020, 11, 1035. [Google Scholar] [CrossRef]
- Gabryelska, A.; Chrzanowski, J.; Sochal, M.; Kaczmarski, P.; Turkiewicz, S.; Ditmer, M.; Karuga, F.F.; Czupryniak, L.; Białasiewicz, P. Nocturnal Oxygen Saturation Parameters as Independent Risk Factors for Type 2 Diabetes Mellitus among Obstructive Sleep Apnea Patients. J. Clin. Med. 2021, 10, 3770. [Google Scholar] [CrossRef]
- Siachpazidou, D.I.; Stavrou, V.T.; Astara, K.; Pastaka, C.; Gogou, E.; Hatzoglou, C.; Economou, N.-T.; Gourgoulianis, K.I. Alzheimer’s Disease in Patients with Obstructive Sleep Apnea Syndrome. Tanaffos 2020, 19, 176–185. [Google Scholar] [CrossRef]
- Andrade, A.G.; Bubu, O.M.; Varga, A.W.; Osorio, R.S. The Relationship between Obstructive Sleep Apnea and Alzheimer’s Disease. J. Alzheimers. Dis. 2018, 64, S255–S270. [Google Scholar] [CrossRef]
- Karuga, F.F.; Kaczmarski, P.; Szmyd, B.; Białasiewicz, P.; Sochal, M.; Gabryelska, A. The Association between Idiopathic Pulmonary Fibrosis and Obstructive Sleep Apnea: A Systematic Review and Meta-Analysis. J. Clin. Med. 2022, 11, 5008. [Google Scholar] [CrossRef]
- Mesarwi, O.A.; Loomba, R.; Malhotra, A. Obstructive Sleep Apnea, Hypoxia, and Nonalcoholic Fatty Liver Disease. Am. J. Respir. Crit. Care Med. 2019, 199, 830–841. [Google Scholar] [CrossRef]
- Kuna, K.; Szewczyk, K.; Gabryelska, A.; Białasiewicz, P.; Ditmer, M.; Strzelecki, D.; Sochal, M. Potential Role of Sleep Deficiency in Inducing Immune Dysfunction. Biomedicines 2022, 10, 2159. [Google Scholar] [CrossRef] [PubMed]
- Ditmer, M.; Gabryelska, A.; Turkiewicz, S.; Białasiewicz, P.; Małecka-wojciesko, E.; Sochal, M. Sleep Problems in Chronic Inflammatory Diseases: Prevalence, Treatment, and New Perspectives: A Narrative Review. J. Clin. Med. 2021, 11, 67. [Google Scholar] [CrossRef] [PubMed]
- Gabryelska, A.; Sochal, M.; Wasik, B.; Bialasiewicz, P. Patients with Obstructive Sleep Apnea Are Over Four Times More Likely to Suffer From Psoriasis Than the General Population. J. Clin. Sleep Med. 2018, 14, 153. [Google Scholar] [CrossRef] [PubMed]
- Athar, W.; Card, M.E.; Charokopos, A.; Akgü, K.M.A.; Derycke, E.C.; Haskell, S.G.; Yaggi, H.K.; Bastian, L.A. Obstructive Sleep Apnea and Pain Intensity in Young Adults. Ann. Am. Thorac. Soc. 2020, 17, 1273–1278. [Google Scholar] [CrossRef]
- Turkiewicz, S.; Ditmer, M.; Sochal, M.; Białasiewicz, P.; Strzelecki, D.; Gabryelska, A. Obstructive Sleep Apnea as an Acceleration Trigger of Cellular Senescence Processes through Telomere Shortening. Int. J. Mol. Sci. 2021, 22, 12536. [Google Scholar] [CrossRef]
- Kaczmarski, P.; Karuga, F.F.; Szmyd, B.; Sochal, M.; Białasiewicz, P.; Strzelecki, D.; Gabryelska, A. The Role of Inflammation, Hypoxia, and Opioid Receptor Expression in Pain Modulation in Patients Suffering from Obstructive Sleep Apnea. Int. J. Mol. Sci. 2022, 23, 9080. [Google Scholar] [CrossRef]
- Koritala, B.S.C.; Conroy, Z.; Smith, D.F. Circadian Biology in Obstructive Sleep Apnea. Diagnostics 2021, 11, 1082. [Google Scholar] [CrossRef]
- Soreca, I. The Role of Circadian Rhythms in Obstructive Sleep Apnea Symptoms and Novel Targets for Treatment. Chronobiol. Int. 2021, 38, 1274–1282. [Google Scholar] [CrossRef]
- Uddin, M.S.; Sumsuzzman, D.M.; Jeandet, P.; Behl, T.; Rauf, A.; Amran, M.S.; Ashraf, G.M. Deciphering the Interacting Mechanisms of Circadian Disruption and Alzheimer’s Disease. Neurochem. Res. 2021, 46, 1603–1617. [Google Scholar] [CrossRef]
- Szmyd, B.; Rogut, M.; Białasiewicz, P.; Gabryelska, A. The Impact of Glucocorticoids and Statins on Sleep Quality. Sleep Med. Rev. 2021, 55, 101380. [Google Scholar] [CrossRef]
- Lemmer, B.; Oster, H. The Role of Circadian Rhythms in the Hypertension of Diabetes Mellitus and the Metabolic Syndrome. Curr. Hypertens. Rep. 2018, 20, 43. [Google Scholar] [CrossRef] [PubMed]
- McHill, A.W.; Wright, K.P. Role of Sleep and Circadian Disruption on Energy Expenditure and in Metabolic Predisposition to Human Obesity and Metabolic Disease. Obes. Rev. 2017, 18 (Suppl. S1), 15–24. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, R.A.; Guthrie, D.W. Pathophysiology of Diabetes Mellitus. Crit. Care Nurs. Q. 2004, 27, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef]
- Fletcher, B.; Gulanick, M.; Lamendola, C. Risk Factors for Type 2 Diabetes Mellitus. J. Cardiovasc. Nurs. 2002, 16, 486. [Google Scholar] [CrossRef]
- Khan, M.A.B.; Hashim, M.J.; King, J.K.; Govender, R.D.; Mustafa, H.; Kaabi, J. Al Epidemiology of Type 2 Diabetes—Global Burden of Disease and Forecasted Trends. J. Epidemiol. Glob. Health 2020, 10, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Punjabi, N.M.; Shahar, E.; Redline, S.; Gottlieb, D.J.; Givelber, R.; Resnick, H.E. Sleep-Disordered Breathing, Glucose Intolerance, and Insulin Resistance: The Sleep Heart Health Study. Am. J. Epidemiol. 2004, 160, 521–530. [Google Scholar] [CrossRef] [Green Version]
- Tanno, S.; Tanigawa, T.; Saito, I.; Nishida, W.; Maruyama, K.; Eguchi, E.; Sakurai, S.; Osawa, H.; Punjabi, N.M. Sleep-Related Intermittent Hypoxemia and Glucose Intolerance: A Community-Based Study. Sleep Med. 2014, 15, 1212–1218. [Google Scholar] [CrossRef]
- Weykamp, C. HbA1c: A Review of Analytical and Clinical Aspects. Ann. Lab. Med. 2013, 33, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Aronsohn, R.S.; Whitmore, H.; Van Cauter, E.; Tasali, E. Impact of Untreated Obstructive Sleep Apnea on Glucose Control in Type 2 Diabetes. Am. J. Respir. Crit. Care Med. 2010, 181, 507–513. [Google Scholar] [CrossRef]
- Kendzerska, T.; Gershon, A.S.; Hawker, G.; Tomlinson, G.; Leung, R.S. Obstructive Sleep Apnea and Incident Diabetes. A Historical Cohort Study. Am. J. Respir. Crit. Care Med. 2014, 190, 218–225. [Google Scholar] [CrossRef] [Green Version]
- Ryan, S. Sleep and Diabetes. Curr. Opin. Pulm. Med. 2018, 24, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Prabhakar, N.R.; Peng, Y.J.; Nanduri, J. Hypoxia-Inducible Factors and Obstructive Sleep Apnea. J. Clin. Investig. 2020, 130, 5042–5051. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, Regional, and National Prevalence of Overweight and Obesity in Children and Adults during 1980–2013: A Systematic Analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef] [Green Version]
- Bray, G.A. Evaluation of Obesity. Who Are the Obese? Postgrad. Med. 2003, 114, 19–38. [Google Scholar] [CrossRef]
- Romero-Corral, A.; Somers, V.K.; Sierra-Johnson, J.; Thomas, R.J.; Collazo-Clavell, M.L.; Korinek, J.; Allison, T.G.; Batsis, J.A.; Sert-Kuniyoshi, F.H.; Lopez-Jimenez, F. Accuracy of Body Mass Index in Diagnosing Obesity in the Adult General Population. Int. J. Obes. 2008, 32, 959–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayoral, L.P.C.; Andrade, G.M.; Mayoral, E.P.C.; Huerta, T.H.; Canseco, S.P.; Rodal Canales, F.J.; Cabrera-Fuentes, H.A.; Cruz, M.M.; Pérez Santiago, A.D.; Alpuche, J.J.; et al. Obesity Subtypes, Related Biomarkers & Heterogeneity. Indian J. Med. Res. 2020, 151, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Geva, N.; Pinhas-Hamiel, O.; Frenkel, H.; Shina, A.; Derazne, E.; Tzur, D.; Afek, A.; Goldbart, A.D.; Twig, G. Obesity and Sleep Disorders: A Nationwide Study of 1.3 Million Israeli Adolescents. Obes. Res. Clin. Pract. 2020, 14, 542–547. [Google Scholar] [CrossRef]
- Grewal, G.; Joshi, G.P. Obesity and Obstructive Sleep Apnea in the Ambulatory Patient. Anesthesiol. Clin. 2019, 37, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, G.D.; Fiorelli, E.M.; Furlan, L.; Montano, N.; Tobaldini, E. Obesity and Sleep Disturbances: The “Chicken or the Egg” Question. Eur. J. Intern. Med. 2021, 92, 11–16. [Google Scholar] [CrossRef]
- Broussard, J.L.; Cauter, E. Van Disturbances of Sleep and Circadian Rhythms: Novel Risk Factors for Obesity. Curr. Opin. Endocrinol. Diabetes Obes. 2016, 23, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Canuto, R.; Pattussi, M.P.; Macagnan, J.B.A.; Henn, R.L.; Olinto, M.T.A. Sleep Deprivation and Obesity in Shift Workers in Southern Brazil. Public Health Nutr. 2014, 17, 2619–2623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, T.; Oyama, I.; Nakamura, T.; Shirane, K.; Otsuka, H.; Kunimoto, M.; Kadowaki, K.; Maruyama, T.; Otomo, H.; Fujino, Y.; et al. Retrospective Cohort Study of the Risk of Obesity among Shift Workers: Findings from the Industry-Based Shift Workers’ Health Study, Japan. Occup. Environ. Med. 2011, 68, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Alodhayani, A.A.; Alshaikh, O.M.; Ghomraoui, F.A.; AlShaibani, T.J.; Algadheeb, A.S.; Bendahmash, A.W.; Baqays, A.A. Correlation between Obesity and Sleep Disturbance in Saudi Medical Students. J. Phys. Ther. Sci. 2017, 29, 181–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, C.M.; Melanson, E.L.; Frydendall, E.J.; Perreault, L.; Eckel, R.H.; Wright, K.P. Energy Expenditure during Sleep, Sleep Deprivation and Sleep Following Sleep Deprivation in Adult Humans. J. Physiol. 2011, 589, 235–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, T.F.; de Bruin, P.F.C.; de Bruin, V.M.S.; Lopes, P.M.; Lemos, F.N. Obesity, Hypersomnolence, and Quality of Sleep: The Impact of Bariatric Surgery. Obes. Surg. 2017, 27, 1775–1779. [Google Scholar] [CrossRef]
- Beccuti, G.; Pannain, S. Sleep and Obesity. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 402–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.J.; Sundar, K.M. Evaluation and Management of Adults with Obstructive Sleep Apnea Syndrome. Lung 2021, 199, 87–101. [Google Scholar] [CrossRef]
- Borel, A.L. Sleep Apnea and Sleep Habits: Relationships with Metabolic Syndrome. Nutrients 2019, 11, 2628. [Google Scholar] [CrossRef] [Green Version]
- Pavlova, M. Circadian Rhythm Sleep-Wake Disorders. Continuum 2017, 23, 1051–1063. [Google Scholar] [CrossRef]
- Lange, T.; Dimitrov, S.; Born, J. Effects of Sleep and Circadian Rhythm on the Human Immune System. Ann. N. Y. Acad. Sci. 2010, 1193, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Quante, M.; Mariani, S.; Weng, J.; Marinac, C.R.; Kaplan, E.R.; Rueschman, M.; Mitchell, J.A.; James, P.; Hipp, J.A.; Cespedes Feliciano, E.M.; et al. Zeitgebers and Their Association with Rest-Activity Patterns. Chronobiol. Int. 2019, 36, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Potter, G.D.M.; Skene, D.J.; Arendt, J.; Cade, J.E.; Grant, P.J.; Hardie, L.J. Circadian Rhythm and Sleep Disruption: Causes, Metabolic Consequences, and Countermeasures. Endocr. Rev. 2016, 37, 584–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panda, S. Circadian Physiology of Metabolism. Science 2016, 354, 1008–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenwasser, A.M.; Turek, F.W. Neurobiology of Circadian Rhythm Regulation. Sleep Med. Clin. 2015, 10, 403–412. [Google Scholar] [CrossRef]
- Hastings, M.H.; Maywood, E.S.; Brancaccio, M. Generation of Circadian Rhythms in the Suprachiasmatic Nucleus. Nat. Rev. Neurosci. 2018, 19, 453–469. [Google Scholar] [CrossRef]
- Patke, A.; Young, M.W.; Axelrod, S. Molecular Mechanisms and Physiological Importance of Circadian Rhythms. Nat. Rev. Mol. Cell Biol. 2020, 21, 67–84. [Google Scholar] [CrossRef]
- Reddy, P.; Zehring, W.A.; Wheeler, D.A.; Pirrotta, V.; Hadfield, C.; Hall, J.C.; Rosbash, M. Molecular Analysis of the Period Locus in Drosophila Melanogaster and Identification of a Transcript Involved in Biological Rhythms. Cell 1984, 38, 701–710. [Google Scholar] [CrossRef]
- Konopka, R.J.; Benzer, S. Clock Mutants of Drosophila Melanogaster. Proc. Natl. Acad. Sci. USA 1971, 68, 2112–2116. [Google Scholar] [CrossRef] [Green Version]
- Maury, E. Off the Clock: From Circadian Disruption to Metabolic Disease. Int. J. Mol. Sci. 2019, 20, 1597. [Google Scholar] [CrossRef]
- Cox, K.H.; Takahashi, J.S. Circadian Clock Genes and the Transcriptional Architecture of the Clock Mechanism. J. Mol. Endocrinol. 2019, 63, R93–R102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, L.U.; Bai, G.; Pang, Y. Roles of NPAS2 in Circadian Rhythm and Disease. Acta Biochim. Biophys. Sin. 2021, 53, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Guillaumond, F.; Dardente, H.; Giguère, V.; Cermakian, N. Differential Control of Bmal1 Circadian Transcription by REV-ERB and ROR Nuclear Receptors. J. Biol. Rhythms 2005, 20, 391–403. [Google Scholar] [CrossRef]
- Zhang, R.; Lahens, N.F.; Ballance, H.I.; Hughes, M.E.; Hogenesch, J.B. A Circadian Gene Expression Atlas in Mammals: Implications for Biology and Medicine. Proc. Natl. Acad. Sci. USA 2014, 111, 16219–16224. [Google Scholar] [CrossRef] [Green Version]
- Van Cauter, E.; Polonsky, K.S.; Scheen, A.J. Roles of Circadian Rhythmicity and Sleep in Human Glucose Regulation. Endocr. Rev. 1997, 18, 716–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson, B.; Knutsson, A.; Lindahl, B. Is There an Association between Shift Work and Having a Metabolic Syndrome? Results from a Population Based Study of 27,485 People. Occup. Environ. Med. 2001, 58, 747–752. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.S.; Armstrong, M.E.G.; Cairns, B.J.; Key, T.J.; Travis, R.C. Shift Work and Chronic Disease: The Epidemiological Evidence. Occup. Med. 2011, 61, 78–89. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.J.; Purvis, T.E.; Mistretta, J.; Scheer, F.A.J.L. Effects of the Internal Circadian System and Circadian Misalignment on Glucose Tolerance in Chronic Shift Workers. J. Clin. Endocrinol. Metab. 2016, 101, 1066–1074. [Google Scholar] [CrossRef]
- Turek, F.W.; Joshu, C.; Kohsaka, A.; Lin, E.; Ivanova, G.; McDearmon, E.; Laposky, A.; Losee-Olson, S.; Easton, A.; Jensen, D.R.; et al. Obesity and Metabolic Syndrome in Circadian Clock Mutant Mice. Science 2005, 308, 1043–1045. [Google Scholar] [CrossRef] [Green Version]
- Marcheva, B.; Ramsey, K.M.; Buhr, E.D.; Kobayashi, Y.; Su, H.; Ko, C.H.; Ivanova, G.; Omura, C.; Mo, S.; Vitaterna, M.H.; et al. Disruption of the Clock Components CLOCK and BMAL1 Leads to Hypoinsulinaemia and Diabetes. Nature 2010, 466, 627–631. [Google Scholar] [CrossRef]
- Barclay, J.L.; Shostak, A.; Leliavski, A.; Tsang, A.H.; Jöhren, O.; Müller-Fielitz, H.; Landgraf, D.; Naujokat, N.; van der Horst, G.T.J.; Oster, H. High-Fat Diet-Induced Hyperinsulinemia and Tissue-Specific Insulin Resistance in Cry-Deficient Mice. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E1053–E1063. [Google Scholar] [CrossRef] [Green Version]
- Lamia, K.A.; Storch, K.F.; Weitz, C.J. Physiological Significance of a Peripheral Tissue Circadian Clock. Proc. Natl. Acad. Sci. USA 2008, 105, 15172–15177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, S.Q.; Ansari, T.S.; McGuinness, O.P.; Wasserman, D.H.; Johnson, C.H. Circadian Disruption Leads to Insulin Resistance and Obesity. Curr. Biol. 2013, 23, 372–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Liu, A.; Weidenhammer, A.; Cooksey, R.C.; McClain, D.; Kim, M.K.; Aguilera, G.; Abel, E.D.; Chung, J.H. The Role of MPer2 Clock Gene in Glucocorticoid and Feeding Rhythms. Endocrinology 2009, 150, 2153–2160. [Google Scholar] [CrossRef] [PubMed]
- Scott, E.M.; Carter, A.M.; Grant, P.J. Association between Polymorphisms in the Clock Gene, Obesity and the Metabolic Syndrome in Man. Int. J. Obes. 2008, 32, 658–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sookoian, S.; Gemma, C.; Gianotti, T.F.; Burgueño, A.; Castaño, G.; Pirola, C.J. Genetic Variants of Clock Transcription Factor Are Associated with Individual Susceptibility to Obesity. Am. J. Clin. Nutr. 2008, 87, 1606–1615. [Google Scholar] [CrossRef] [Green Version]
- Woon, P.Y.; Kaisaki, P.J.; Bragança, J.; Bihoreau, M.T.; Levy, J.C.; Farrall, M.; Gauguier, D. Aryl Hydrocarbon Receptor Nuclear Translocator-like (BMAL1) Is Associated with Susceptibility to Hypertension and Type 2 Diabetes. Proc. Natl. Acad. Sci. USA 2007, 104, 14412–14417. [Google Scholar] [CrossRef] [Green Version]
- Dupuis, J.; Langenberg, C.; Prokopenko, I.; Saxena, R.; Soranzo, N.; Jackson, A.U.; Wheeler, E.; Glazer, N.L.; Bouatia-Naji, N.; Gloyn, A.L.; et al. New Genetic Loci Implicated in Fasting Glucose Homeostasis and Their Impact on Type 2 Diabetes Risk. Nat. Genet. 2010, 42, 105–116. [Google Scholar] [CrossRef]
- Ruano, E.G.; Canivell, S.; Vieira, E. REV-ERB ALPHA Polymorphism Is Associated with Obesity in the Spanish Obese Male Population. PLoS ONE 2014, 9, e104065. [Google Scholar] [CrossRef] [Green Version]
- Titchenell, P.M.; Lazar, M.A.; Birnbaum, M.J. Unraveling the Regulation of Hepatic Metabolism by Insulin. Trends Endocrinol. Metab. 2017, 28, 497–505. [Google Scholar] [CrossRef]
- Challet, E.; Malan, A.; Turek, F.W.; Van Reeth, O. Daily Variations of Blood Glucose, Acid-Base State and PCO2 in Rats: Effect of Light Exposure. Neurosci. Lett. 2004, 355, 131–135. [Google Scholar] [CrossRef]
- Cailotto, C.; La Fleur, S.E.; Van Heijningen, C.; Wortel, J.; Kalsbeek, A.; Feenstra, M.; Pévet, P.; Buijs, R.M. The Suprachiasmatic Nucleus Controls the Daily Variation of Plasma Glucose via the Autonomic Output to the Liver: Are the Clock Genes Involved? Eur. J. Neurosci. 2005, 22, 2531–2540. [Google Scholar] [CrossRef] [PubMed]
- Kalsbeek, A.; Strubbe, J.H. Circadian Control of Insulin Secretion Is Independent of the Temporal Distribution of Feeding. Physiol. Behav. 1998, 63, 553–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, K.F.; Nestel, P.J. Diurnal Variation in Glucose Tolerance and in Insulin Secretion in Man. Diabetes 1973, 22, 333–348. [Google Scholar] [CrossRef]
- Baker, I.A.; Jarrett, R.J. Diurnal Variation in the Blood-Sugar and Plasma-Insulin Response to Tolbutamide. Lancet 1972, 2, 945–947. [Google Scholar] [CrossRef]
- Vollmers, C.; Gill, S.; DiTacchio, L.; Pulivarthy, S.R.; Le, H.D.; Panda, S. Time of Feeding and the Intrinsic Circadian Clock Drive Rhythms in Hepatic Gene Expression. Proc. Natl. Acad. Sci. USA 2009, 106, 21453–21458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Fleur, S.E.; Kalsbeek, A.; Wortel, J.; Fekkes, M.L.; Buijs, R.M. A Daily Rhythm in Glucose Tolerance: A Role for the Suprachiasmatic Nucleus. Diabetes 2001, 50, 1237–1243. [Google Scholar] [CrossRef] [Green Version]
- Hogenboom, R.; Kalsbeek, M.J.; Korpel, N.L.; de Goede, P.; Koenen, M.; Buijs, R.M.; Romijn, J.A.; Swaab, D.F.; Kalsbeek, A.; Yi, C.X. Loss of Arginine Vasopressin- and Vasoactive Intestinal Polypeptide-Containing Neurons and Glial Cells in the Suprachiasmatic Nucleus of Individuals with Type 2 Diabetes. Diabetologia 2019, 62, 2088–2093. [Google Scholar] [CrossRef] [Green Version]
- Dickmeis, T.; Weger, B.D.; Weger, M. The Circadian Clock and Glucocorticoids--Interactions across Many Time Scales. Mol. Cell. Endocrinol. 2013, 380, 2–15. [Google Scholar] [CrossRef]
- Kuo, T.; McQueen, A.; Chen, T.C.; Wang, J.C. Regulation of Glucose Homeostasis by Glucocorticoids. Adv. Exp. Med. Biol. 2015, 872, 99–126. [Google Scholar] [CrossRef]
- Buijs, R.M.; Wortel, J.; Van Heerikhuize, J.J.; Feenstra, M.G.; Ter Horst, G.J.; Romijn, H.J.; Kalsbeek, A. Anatomical and Functional Demonstration of a Multisynaptic Suprachiasmatic Nucleus Adrenal (Cortex) Pathway. Eur. J. Neurosci. 1999, 11, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Li, M.D.; Xin, H.; Yuan, Y.; Yang, X.; Li, H.; Tian, D.; Zhang, H.; Zhang, Z.; Han, T.L.; Chen, Q.; et al. Circadian Clock-Controlled Checkpoints in the Pathogenesis of Complex Disease. Front. Genet. 2021, 12, 721231. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Ueno, T.; Tsunemi, A.; Nagura, C.; Tahira, K.; Fukuda, N.; Soma, M.; Abe, M. The Adrenal Gland Circadian Clock Exhibits a Distinct Phase Advance in Spontaneously Hypertensive Rats. Hypertens. Res. 2019, 42, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Bright, G.M.; Melton, T.W.; Rogol, A.D.; Clarke, W.L. Failure of Cortisol Blockade to Inhibit Early Morning Increases in Basal Insulin Requirements in Fasting Insulin-Dependent Diabetics. Diabetes 1980, 29, 662–664. [Google Scholar] [CrossRef] [Green Version]
- Moøller, N.; Joørgensen, J.O.L. Effects of Growth Hormone on Glucose, Lipid, and Protein Metabolism in Human Subjects. Endocr. Rev. 2009, 30, 152–177. [Google Scholar] [CrossRef] [Green Version]
- Linkowski, P.; Kerkhofs, M.; van Onderbergen, A.; Hubain, P.; Copinschi, G.; L’Hermite-Balériaux, M.; Leclercq, R.; Brasseur, M.; Mendlewicz, J.; van Cauter, E. The 24-Hour Profiles of Cortisol, Prolactin, and Growth Hormone Secretion in Mania. Arch. Gen. Psychiatry 1994, 51, 616–624. [Google Scholar] [CrossRef]
- Linkowski, P.; Mendlewicz, J.; Kerkhofs, M.; Leclercq, R.; Golstein, J.; Brasseur, M.; Copinschi, G.; Cauter, E. Van 24-Hour Profiles of Adrenocorticotropin, Cortisol, and Growth Hormone in Major Depressive Illness: Effect of Antidepressant Treatment. J. Clin. Endocrinol. Metab. 1987, 65, 141–152. [Google Scholar] [CrossRef]
- Brandenberger, G.; Weibel, L. The 24-h Growth Hormone Rhythm in Men: Sleep and Circadian Influences Questioned. J. Sleep Res. 2004, 13, 251–255. [Google Scholar] [CrossRef]
- Weibel, L.; Follenius, M.; Spiegel, K.; Gronfier, C.; Brandenberger, G. Growth Hormone Secretion in Night Workers. Chronobiol. Int. 1997, 14, 49–60. [Google Scholar] [CrossRef]
- Weibel, L.; Brandenberger, G. Disturbances in Hormonal Profiles of Night Workers during Their Usual Sleep and Work Times. J. Biol. Rhythms 1998, 13, 202–208. [Google Scholar] [CrossRef]
- Livingstone, C. Insulin-like Growth Factor-I (IGF-I) and Clinical Nutrition. Clin. Sci. 2013, 125, 265–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livingstone, C. The Insulin-like Growth Factor System and Nutritional Assessment. Scientifica 2012, 2012, 768731. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Cordoba-Chacon, J.; Kineman, R.D.; Cronstein, B.N.; Muzumdar, R.; Gong, Z.; Werner, H.; Yakar, S. Growth Hormone Control of Hepatic Lipid Metabolism. Diabetes 2016, 65, 3598–3609. [Google Scholar] [CrossRef] [Green Version]
- Chaudhari, A.; Gupta, R.; Patel, S.; Velingkaar, N.; Kondratov, R. Cryptochromes Regulate IGF-1 Production and Signaling through Control of JAK2-Dependent STAT5B Phosphorylation. Mol. Biol. Cell 2017, 28, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Bertani, S.; Carboni, L.; Criado, A.; Michielin, F.; Mangiarini, L.; Vicentini, E. Circadian Profile of Peripheral Hormone Levels in Sprague-Dawley Rats and in Common Marmosets (Callithrix jacchus). In Vivo 2010, 24, 827–836. [Google Scholar] [CrossRef]
- Heuck, C.; SkjrbK, C.; Rskov, H.; Wolthers, O.D. Circadian Variation in Serum Free Ultrafiltrable Insulin-like Growth Factor I Concentrations in Healthy Children. Pediatr. Res. 1999, 45, 733–736. [Google Scholar] [CrossRef] [Green Version]
- HULBERT, A.J. Thyroid Hormones and Their Effects: A New Perspective. Biol. Rev. Camb. Philos. Soc. 2000, 75, 519–631. [Google Scholar] [CrossRef] [Green Version]
- Mullur, R.; Liu, Y.Y.; Brent, G.A. Thyroid Hormone Regulation of Metabolism. Physiol. Rev. 2014, 94, 355–382. [Google Scholar] [CrossRef] [Green Version]
- Ortiga-Carvalho, T.M.; Chiamolera, M.I.; Pazos-Moura, C.C.; Wondisford, F.E. Hypothalamus-Pituitary-Thyroid Axis. Compr. Physiol. 2016, 6, 1387–1428. [Google Scholar] [CrossRef]
- Dimitriadis, G.D.; Raptis, S.A. Thyroid Hormone Excess and Glucose Intolerance. Exp. Clin. Endocrinol. Diabetes 2001, 109 (Suppl. S2), S225–S239. [Google Scholar] [CrossRef]
- Maxon, H.R.; Kreines, K.W.; Goldsmith, R.E.; Knowles, H.C. Long-Term Observations of Glucose Tolerance in Thyrotoxic Patients. Arch. Intern. Med. 1975, 135, 1477–1480. [Google Scholar] [CrossRef] [PubMed]
- Kreines, K.; Jett, M.; Knowles, H.C. Observations in Hyperthyroidism of Abnormal Glucose Tolerance and Other Traits Related to Diabetes Mellitus. Diabetes 1965, 14, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Comte, B.; Vidal, H.; Laville, M.; Riou, J.P. Influence of Thyroid Hormones on Gluconeogenesis from Glycerol in Rat Hepatocytes: A Dose-Response Study. Metabolism 1990, 39, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Kim, Y.; Shin, J.; Lee, S.A.; Choi, Y.; Park, E.C. Association between Thyroid Hormones and the Components of Metabolic Syndrome. BMC Endocr. Disord. 2018, 18, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzoccoli, G.; Giuliani, A.; Carughi, S.; De Cata, A.; Puzzolante, F.; La Viola, M.; Urbano, N.; Perfetto, F.; Tarquini, R. The Hypothalamic-Pituitary-Thyroid Axis and Melatonin in Humans: Possible Interactions in the Control of Body Temperature. Neuro Endocrinol. Lett. 2004, 25, 368–372. [Google Scholar] [CrossRef] [Green Version]
- Roelfsema, F.; Pijl, H.; Kok, P.; Endert, E.; Fliers, E.; Biermasz, N.R.; Pereira, A.M.; Veldhuis, J.D. Thyrotropin Secretion in Healthy Subjects Is Robust and Independent of Age and Gender, and Only Weakly Dependent on Body Mass Index. J. Clin. Endocrinol. Metab. 2014, 99, 570–578. [Google Scholar] [CrossRef] [Green Version]
- Fahrenkrug, J.; Georg, B.; Hannibal, J.; Jørgensen, H.L. Hypophysectomy Abolishes Rhythms in Rat Thyroid Hormones but Not in the Thyroid Clock. J. Endocrinol. 2017, 233, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Karamitri, A.; Jockers, R. Melatonin in Type 2 Diabetes Mellitus and Obesity. Nat. Rev. Endocrinol. 2019, 15, 105–125. [Google Scholar] [CrossRef]
- Jockers, R.; Delagrange, P.; Dubocovich, M.L.; Markus, R.P.; Renault, N.; Tosini, G.; Cecon, E.; Zlotos, D.P. Update on Melatonin Receptors: IUPHAR Review 20. Br. J. Pharmacol. 2016, 173, 2702–2725. [Google Scholar] [CrossRef] [Green Version]
- Cipolla-Neto, J.; Do Amaral, F.G. Melatonin as a Hormone: New Physiological and Clinical Insights. Endocr. Rev. 2018, 39, 990–1028. [Google Scholar] [CrossRef]
- Bouatia-Naji, N.; Bonnefond, A.; Cavalcanti-Proença, C.; Sparsø, T.; Holmkvist, J.; Marchand, M.; Delplanque, J.; Lobbens, S.; Rocheleau, G.; Durand, E.; et al. A Variant near MTNR1B Is Associated with Increased Fasting Plasma Glucose Levels and Type 2 Diabetes Risk. Nat. Genet. 2009, 41, 89–94. [Google Scholar] [CrossRef]
- Lyssenko, V.; Nagorny, C.L.F.; Erdos, M.R.; Wierup, N.; Jonsson, A.; Spégel, P.; Bugliani, M.; Saxena, R.; Fex, M.; Pulizzi, N.; et al. Common Variant in MTNR1B Associated with Increased Risk of Type 2 Diabetes and Impaired Early Insulin Secretion. Nat. Genet. 2009, 41, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Prokopenko, I.; Langenberg, C.; Florez, J.C.; Saxena, R.; Soranzo, N.; Thorleifsson, G.; Loos, R.J.F.; Manning, A.K.; Jackson, A.U.; Aulchenko, Y.; et al. Variants in MTNR1B Influence Fasting Glucose Levels. Nat. Genet. 2009, 41, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Tam, C.H.T.; Ho, J.S.K.; Wang, Y.; Lee, H.M.; Lam, V.K.L.; Germer, S.; Martin, M.; So, W.Y.; Ma, R.C.W.; Chan, J.C.N.; et al. Common Polymorphisms in MTNR1B, G6PC2 and GCK Are Associated with Increased Fasting Plasma Glucose and Impaired Beta-Cell Function in Chinese Subjects. PLoS ONE 2010, 5, e11428. [Google Scholar] [CrossRef] [PubMed]
- La Fleur, S.E.; Kalsbeek, A.; Wortel, J.; Van Der Vliet, J.; Buijs, R.M. Role for the Pineal and Melatonin in Glucose Homeostasis: Pinealectomy Increases Night-Time Glucose Concentrations. J. Neuroendocrinol. 2001, 13, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Alcantara, S.; Baba, K.; Tosini, G. Removal of Melatonin Receptor Type 1 Induces Insulin Resistance in the Mouse. Obesity 2010, 18, 1861–1863. [Google Scholar] [CrossRef] [Green Version]
- Mühlbauer, E.; Gross, E.; Labucay, K.; Wolgast, S.; Peschke, E. Loss of Melatonin Signalling and Its Impact on Circadian Rhythms in Mouse Organs Regulating Blood Glucose. Eur. J. Pharmacol. 2009, 606, 61–71. [Google Scholar] [CrossRef]
- Bazwinsky-Wutschke, I.; Bieseke, L.; Mühlbauer, E.; Peschke, E. Influence of Melatonin Receptor Signalling on Parameters Involved in Blood Glucose Regulation. J. Pineal Res. 2014, 56, 82–96. [Google Scholar] [CrossRef]
- Kukkonen, J.P.; Turunen, P.M. Cellular Signaling Mechanisms of Hypocretin/Orexin. Front. Neurol. Neurosci. 2021, 45, 91–102. [Google Scholar] [CrossRef]
- Sakurai, T. The Neural Circuit of Orexin (Hypocretin): Maintaining Sleep and Wakefulness. Nat. Rev. Neurosci. 2007, 8, 171–181. [Google Scholar] [CrossRef]
- Chieffi, S.; Carotenuto, M.; Monda, V.; Valenzano, A.; Villano, I.; Precenzano, F.; Tafuri, D.; Salerno, M.; Filippi, N.; Nuccio, F.; et al. Orexin System: The Key for a Healthy Life. Front. Physiol. 2017, 8, 357. [Google Scholar] [CrossRef] [Green Version]
- Zeitzer, J.M.; Buckmaster, C.L.; Parker, K.J.; Hauck, C.M.; Lyons, D.M.; Mignot, E. Circadian and Homeostatic Regulation of Hypocretin in a Primate Model: Implications for the Consolidation of Wakefulness. J. Neurosci. 2003, 23, 3555–3560. [Google Scholar] [CrossRef] [Green Version]
- Salomon, R.M.; Ripley, B.; Kennedy, J.S.; Johnson, B.; Schmidt, D.; Zeitzer, J.M.; Nishino, S.; Mignot, E. Diurnal Variation of Cerebrospinal Fluid Hypocretin-1 (Orexin-A) Levels in Control and Depressed Subjects. Biol. Psychiatry 2003, 54, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Gabryelska, A.; Szmyd, B.; Maschauer, E.L.; Roguski, A.; Canham, R.; Morrison, I.; Białasiewicz, P.; Riha, R.L. Utility of Measuring CSF Hypocretin-1 Level in Patients with Suspected Narcolepsy. Sleep Med. 2020, 71, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Tsuneki, H.; Tokai, E.; Nakamura, Y.; Takahashi, K.; Fujita, M.; Asaoka, T.; Kon, K.; Anzawa, Y.; Wada, T.; Takasaki, I.; et al. Hypothalamic Orexin Prevents Hepatic Insulin Resistance via Daily Bidirectional Regulation of Autonomic Nervous System in Mice. Diabetes 2015, 64, 459–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeghate, E.; Fernandez-Cabezudo, M.; Hameed, R.; El-Hasasna, H.; El Wasila, M.; Abbas, T.; Al-Ramadi, B. Orexin-1 Receptor Co-Localizes with Pancreatic Hormones in Islet Cells and Modulates the Outcome of Streptozotocin-Induced Diabetes Mellitus. PLoS ONE 2010, 5, e8587. [Google Scholar] [CrossRef] [Green Version]
- Adeghate, E.; Hameed, R. Mechanism of Orexin B-Stimulated Insulin and Glucagon Release from the Pancreas of Normal and Diabetic Rats. Pancreas 2011, 40, 131–136. [Google Scholar] [CrossRef]
- Nowak, K.W.; Strowski, M.Z.; Switonska, M.M.; Kaczmarek, P.; Singh, V.; Fabis, M.; Mackowiak, P.; Nowak, M.; Malendowicz, L.K. Evidence That Orexins A and B Stimulate Insulin Secretion from Rat Pancreatic Islets via Both Receptor Subtypes. Int. J. Mol. Med. 2005, 15, 969–972. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Sun, C.; Wang, B.; Yan, P.; Wu, A.; Yang, G.; Li, W. Orexin-A Stimulates the Expression of GLUT4 in a Glucose Dependent Manner in the Liver of Orange-Spotted Grouper (Epinephelus coioides). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2016, 199, 95–104. [Google Scholar] [CrossRef]
- Kaczmarek, P.; Skrzypski, M.; Pruszynska-Oszmalek, E.; Sassek, M.; Kolodziejski, P.A.; Billert, M.; Szczepankiewicz, D.; Wojciechowicz, T.; Maechler, P.; Nowak, K.W.; et al. Chronic Orexin-A (Hypocretin-1) Treatment of Type 2 Diabetic Rats Improves Glucose Control and Beta-Cell Functions. J. Physiol. Pharmacol. 2017, 68, 669–681. [Google Scholar] [CrossRef]
- Joshi, D.; Sarkar, D.; Singh, S.K. Decreased Expression of Orexin 1 Receptor in Adult Mice Testes during Alloxan-Induced Diabetes Mellitus Perturbs Testicular Steroidogenesis and Glucose Homeostasis. Biochem. Biophys. Res. Commun. 2017, 490, 1346–1354. [Google Scholar] [CrossRef] [PubMed]
- Colldén, G.; Tschöp, M.H.; Müller, T.D. Therapeutic Potential of Targeting the Ghrelin Pathway. Int. J. Mol. Sci. 2017, 18, 798. [Google Scholar] [CrossRef] [Green Version]
- Perchard, R.; Clayton, P.E. Ghrelin and Growth. Endocr. Dev. 2017, 32, 74–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szentirmai, É.; Kapás, L.; Krueger, J.M. Ghrelin Microinjection into Forebrain Sites Induces Wakefulness and Feeding in Rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R575–R585. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Morris, C.J.; Caputo, R.; Garaulet, M.; Scheer, F.A.J.L. Ghrelin Is Impacted by the Endogenous Circadian System and by Circadian Misalignment in Humans. Int. J. Obes. 2019, 43, 1644–1649. [Google Scholar] [CrossRef] [PubMed]
- Van Der Lely, A.J.; Tschöp, M.; Heiman, M.L.; Ghigo, E. Biological, Physiological, Pathophysiological, and Pharmacological Aspects of Ghrelin. Endocr. Rev. 2004, 25, 426–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heppner, K.M.; Tong, J.; Kirchner, H.; Nass, R.; Tschöp, M.H. The Ghrelin O-Acyltransferase-Ghrelin System: A Novel Regulator of Glucose Metabolism. Curr. Opin. Endocrinol. Diabetes Obes. 2011, 18, 50–55. [Google Scholar] [CrossRef]
- Su, M.; Yan, M.; Yao, J.; Fang, Y.; Jin, H.; Gong, Y. Unacylated Ghrelin Regulates Glucose-Sensitive Neurons Activity and Glycolipid Metabolism via Orexin-A Neurons in the Lateral Hypothalamic Area. Horm. Metab. Res. 2020, 52, 747–754. [Google Scholar] [CrossRef]
- Verhulst, P.J.; Depoortere, I. Ghrelin’s Second Life: From Appetite Stimulator to Glucose Regulator. World J. Gastroenterol. 2012, 18, 3183–3195. [Google Scholar] [CrossRef]
- Sun, Y.; Asnicar, M.; Saha, P.K.; Chan, L.; Smith, R.G. Ablation of Ghrelin Improves the Diabetic but Not Obese Phenotype of Ob/Ob Mice. Cell Metab. 2006, 3, 379–386. [Google Scholar] [CrossRef]
- Tibaldi, J.M. Intensifying Insulin Therapy in Type 2 Diabetes Mellitus: Dosing Options for Insulin Analogue Premixes. Clin. Ther. 2011, 33, 1630–1642. [Google Scholar] [CrossRef] [PubMed]
- Carroll, M.F.; Schade, D.S. The Dawn Phenomenon Revisited: Implications for Diabetes Therapy. Endocr. Pract. 2005, 11, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Monnier, L.; Colette, C.; Dejager, S.; Owens, D. Magnitude of the Dawn Phenomenon and Its Impact on the Overall Glucose Exposure in Type 2 Diabetes: Is This of Concern? Diabetes Care 2013, 36, 4057–4062. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Wang, H.; Li, Y.; Tao, X.; Sun, J. Poor Sleep Quality Is Associated with Dawn Phenomenon and Impaired Circadian Clock Gene Expression in Subjects with Type 2 Diabetes Mellitus. Int. J. Endocrinol. 2017, 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, G.; Li, X.; Hou, X.; Zhou, W.; Gong, Y.; Liu, F.; He, Y.; Song, J.; Wang, J.; Basil, P.; et al. REV-ERB in GABAergic Neurons Controls Diurnal Hepatic Insulin Sensitivity. Nature 2021, 592, 763–767. [Google Scholar] [CrossRef]
- Feng, D.; Liu, T.; Sun, Z.; Bugge, A.; Mullican, S.E.; Alenghat, T.; Liu, X.S.; Lazar, M.A. A Circadian Rhythm Orchestrated by Histone Deacetylase 3 Controls Hepatic Lipid Metabolism. Science 2011, 331, 1315–1319. [Google Scholar] [CrossRef] [Green Version]
- Garbarino-Pico, E.; Carpentieri, A.R.; Castagnet, P.I.; Pasquaré, S.J.; Giusto, N.M.; Caputto, B.L.; Guido, M.E. Synthesis of Retinal Ganglion Cell Phospholipids Is under Control of an Endogenous Circadian Clock: Daily Variations in Phospholipid-Synthesizing Enzyme Activities. J. Neurosci. Res. 2004, 76, 642–652. [Google Scholar] [CrossRef]
- Ikemoto, A.; Fukuma, A.; Fujii, Y.; Okuyama, H. Diurnal Rhythms of Retinal Phospholipid Synthetic Enzymes Are Retained but Their Activities Are Decreased in Rats under Alpha-Linolenic Acid Deficiency. Arch. Biochem. Biophys. 2000, 383, 108–113. [Google Scholar] [CrossRef]
- Minami, Y.; Kasukawa, T.; Kakazu, Y.; Iigo, M.; Sugimoto, M.; Ikeda, S.; Yasui, A.; Van Der Horst, G.T.J.; Soga, T.; Ueda, H.R. Measurement of Internal Body Time by Blood Metabolomics. Proc. Natl. Acad. Sci. USA 2009, 106, 9890–9895. [Google Scholar] [CrossRef] [Green Version]
- Ruf, T.; Arnold, W. Daily and Seasonal Rhythms in Human Mucosa Phospholipid Fatty Acid Composition. J. Biol. Rhythms 2015, 30, 331–341. [Google Scholar] [CrossRef]
- Singh, R.; Sharma, S.; Singh, R.K.; Mahdi, A.A.; Singh, R.K.; Lee Gierke, C.; Cornelissen, G. Effect of Gender, Age, Diet and Smoking Status on Chronomics of Circulating Plasma Lipid Components in Healthy Indians. Clin. Chim. Acta. 2016, 459, 10–18. [Google Scholar] [CrossRef]
- Zhou, H.; Hylemon, P.B. Bile Acids Are Nutrient Signaling Hormones. Steroids 2014, 86, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Russell, D.W. The Enzymes, Regulation, and Genetics of Bile Acid Synthesis. Annu. Rev. Biochem. 2003, 72, 137–174. [Google Scholar] [CrossRef] [Green Version]
- Chiang, J.Y.L.; Ferrell, J.M. Bile Acid Metabolism in Liver Pathobiology. Gene Expr. 2018, 18, 71–87. [Google Scholar] [CrossRef] [Green Version]
- Qi, L.; Tian, Y.; Chen, Y. Circulating Bile Acid Profiles: A Need for Further Examination. J. Clin. Endocrinol. Metab. 2021, 106, 3093–3112. [Google Scholar] [CrossRef]
- Gälman, C.; Angelin, B.; Rudling, M. Bile Acid Synthesis in Humans Has a Rapid Diurnal Variation That Is Asynchronous with Cholesterol Synthesis. Gastroenterology 2005, 129, 1445–1453. [Google Scholar] [CrossRef]
- Eggink, H.M.; Oosterman, J.E.; de Goede, P.; de Vries, E.M.; Foppen, E.; Koehorst, M.; Groen, A.K.; Boelen, A.; Romijn, J.A.; la Fleur, S.E.; et al. Complex Interaction between Circadian Rhythm and Diet on Bile Acid Homeostasis in Male Rats. Chronobiol. Int. 2017, 34, 1339–1353. [Google Scholar] [CrossRef]
- Pathak, P.; Li, T.; Chiang, J.Y.L. Retinoic Acid-Related Orphan Receptor α Regulates Diurnal Rhythm and Fasting Induction of Sterol 12α-Hydroxylase in Bile Acid Synthesis. J. Biol. Chem. 2013, 288, 37154–37165. [Google Scholar] [CrossRef] [Green Version]
- Duez, H.; van der Veen, J.N.; Duhem, C.; Pourcet, B.; Touvier, T.; Fontaine, C.; Derudas, B.; Baugé, E.; Havinga, R.; Bloks, V.W.; et al. Regulation of Bile Acid Synthesis by the Nuclear Receptor Rev-Erbalpha. Gastroenterology 2008, 135, 689–698. [Google Scholar] [CrossRef]
- Kerr, T.A.; Saeki, S.; Schneider, M.; Schaefer, K.; Berdy, S.; Redder, T.; Shan, B.; Russell, D.W.; Schwarz, M. Loss of Nuclear Receptor SHP Impairs but Does Not Eliminate Negative Feedback Regulation of Bile Acid Synthesis. Dev. Cell 2002, 2, 713–720. [Google Scholar] [CrossRef]
- Chiang, J.Y.L.; Ferrell, J.M. Bile Acid Receptors FXR and TGR5 Signaling in Fatty Liver Diseases and Therapy. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 318, G554–G573. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Zhang, R.; Jain, R.; Shi, H.; Zhang, L.; Zhou, G.; Sangwung, P.; Tugal, D.; Atkins, G.B.; Prosdocimo, D.A.; et al. Circadian Control of Bile Acid Synthesis by a KLF15-Fgf15 Axis. Nat. Commun. 2015, 6, 7231. [Google Scholar] [CrossRef] [Green Version]
- Ma, K.; Xiao, R.; Tseng, H.T.; Shan, L.; Fu, L.; Moore, D.D. Circadian Dysregulation Disrupts Bile Acid Homeostasis. PLoS ONE 2009, 4, e6843. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Yang, J.; Xiang, D.; Li, G.; Liu, D.; Zhang, C. Circadian Rhythms and Bile Acid Homeostasis: A Comprehensive Review. Chronobiol. Int. 2020, 37, 618–628. [Google Scholar] [CrossRef]
- Sultani, G.; Samsudeen, A.F.; Osborne, B.; Turner, N. NAD+: A Key Metabolic Regulator with Great Therapeutic Potential. J. Neuroendocrinol. 2017, 29, e12508. [Google Scholar] [CrossRef]
- Cantó, C.; Gerhart-Hines, Z.; Feige, J.N.; Lagouge, M.; Noriega, L.; Milne, J.C.; Elliott, P.J.; Puigserver, P.; Auwerx, J. AMPK Regulates Energy Expenditure by Modulating NAD+ Metabolism and SIRT1 Activity. Nature 2009, 458, 1056–1060. [Google Scholar] [CrossRef] [Green Version]
- Ramsey, K.M.; Yoshino, J.; Brace, C.S.; Abrassart, D.; Kobayashi, Y.; Marcheva, B.; Hong, H.K.; Chong, J.L.; Buhr, E.D.; Lee, C.; et al. Circadian Clock Feedback Cycle through NAMPT-Mediated NAD+ Biosynthesis. Science 2009, 324, 651–654. [Google Scholar] [CrossRef] [Green Version]
- Peek, C.B.; Affinati, A.H.; Ramsey, K.M.; Kuo, H.Y.; Yu, W.; Sena, L.A.; Ilkayeva, O.; Marcheva, B.; Kobayashi, Y.; Omura, C.; et al. Circadian Clock NAD+ Cycle Drives Mitochondrial Oxidative Metabolism in Mice. Science 2013, 342, 1243417. [Google Scholar] [CrossRef] [Green Version]
- Storch, K.F.; Lipan, O.; Leykin, I.; Viswanathan, N.; Davis, F.C.; Wong, W.H.; Weitz, C.J. Extensive and Divergent Circadian Gene Expression in Liver and Heart. Nature 2002, 417, 78–83. [Google Scholar] [CrossRef]
- Chang, H.C.; Guarente, L. SIRT1 and Other Sirtuins in Metabolism. Trends Endocrinol. Metab. 2014, 25, 138–145. [Google Scholar] [CrossRef]
- Bellet, M.M.; Orozco-Solis, R.; Sahar, S.; Eckel-Mahan, K.; Sassone-Corsi, P. The Time of Metabolism: NAD+, SIRT1, and the Circadian Clock. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 31–38. [Google Scholar] [CrossRef]
- Gottlieb, D.J.; Punjabi, N.M. Diagnosis and Management of Obstructive Sleep Apnea: A Review. JAMA 2020, 323, 1380–1400. [Google Scholar] [CrossRef] [PubMed]
- Marrone, O.; Bonsignore, M.R. Blood-Pressure Variability in Patients with Obstructive Sleep Apnea: Current Perspectives. Nat. Sci. Sleep 2018, 10, 229–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, R.; Zhang, Y.; Yang, L.; Somers, V.K.; Covassin, N.; Tang, X. Association between Arousals during Sleep and Hypertension among Patients with Obstructive Sleep Apnea. J. Am. Heart Assoc. 2022, 11, e022141. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; Seo, B.S.; Kim, J.H. Effect of Arousal on Sympathetic Overactivity in Patients with Obstructive Sleep Apnea. Sleep Med. 2019, 62, 86–91. [Google Scholar] [CrossRef]
- Taylor, K.S.; Murai, H.; Millar, P.J.; Haruki, N.; Kimmerly, D.S.; Morris, B.L.; Tomlinson, G.; Bradley, T.D.; Floras, J.S. Arousal From Sleep and Sympathetic Excitation During Wakefulness. Hypertension 2016, 68, 1467–1474. [Google Scholar] [CrossRef]
- Gabryelska, A.; Turkiewicz, S.; Karuga, F.F.; Sochal, M.; Strzelecki, D.; Białasiewicz, P. Disruption of Circadian Rhythm Genes in Obstructive Sleep Apnea Patients-Possible Mechanisms Involved and Clinical Implication. Int. J. Mol. Sci. 2022, 23, 709. [Google Scholar] [CrossRef]
- von Allmen, D.C.; Francey, L.J.; Rogers, G.M.; Ruben, M.D.; Cohen, A.P.; Wu, G.; Schmidt, R.E.; Ishman, S.L.; Amin, R.S.; Hogenesch, J.B.; et al. Circadian Dysregulation: The Next Frontier in Obstructive Sleep Apnea Research. Otolaryngol.–Head Neck Surg. 2018, 159, 948–955. [Google Scholar] [CrossRef]
- Tirpe, A.A.; Gulei, D.; Ciortea, S.M.; Crivii, C.; Berindan-Neagoe, I. Hypoxia: Overview on Hypoxia-Mediated Mechanisms with a Focus on the Role of HIF Genes. Int. J. Mol. Sci. 2019, 20, 6140. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. Hypoxia-Inducible Factors in Physiology and Medicine. Cell 2012, 148, 399–408. [Google Scholar] [CrossRef]
- Kaelin, W.G.; Ratcliffe, P.J. Oxygen Sensing by Metazoans: The Central Role of the HIF Hydroxylase Pathway. Mol. Cell 2008, 30, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Ko, J.; Ju, C.; Eltzschig, H.K. Hypoxia Signaling in Human Diseases and Therapeutic Targets. Exp. Mol. Med. 2019, 51, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Ke, Q.; Costa, M. Hypoxia-Inducible Factor-1 (HIF-1). Mol. Pharmacol. 2006, 70, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Adamovich, Y.; Ladeuix, B.; Golik, M.; Koeners, M.P.; Asher, G. Rhythmic Oxygen Levels Reset Circadian Clocks through HIF1α. Cell Metab. 2017, 25, 93–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozek, K.; Kiełbasa, S.M.; Kramer, A.; Herzel, H. Promoter Analysis of Mammalian Clock Controlled Genes. Genome Inform. 2007, 18, 65–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koike, N.; Yoo, S.H.; Huang, H.C.; Kumar, V.; Lee, C.; Kim, T.K.; Takahashi, J.S. Transcriptional Architecture and Chromatin Landscape of the Core Circadian Clock in Mammals. Science 2012, 338, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Chilov, D.; Hofer, T.; Bauer, C.; Wenger, R.H.; Gassmann, M. Hypoxia Affects Expression of Circadian Genes PER1 and CLOCK in Mouse Brain. FASEB J. 2001, 15, 2613–2622. [Google Scholar] [CrossRef]
- Gabryelska, A.; Szmyd, B.; Panek, M.; Szemraj, J.; Kuna, P.; Białasiewicz, P. Serum Hypoxia-Inducible Factor-1α Protein Level as a Diagnostic Marker of Obstructive Sleep Apnea. Polish Arch. Intern. Med. 2020, 130, 158–160. [Google Scholar] [CrossRef] [Green Version]
- Gabryelska, A.; Sochal, M.; Turkiewicz, S.; Białasiewicz, P. Relationship between HIF-1 and Circadian Clock Proteins in Obstructive Sleep Apnea Patients-Preliminary Study. J. Clin. Med. 2020, 9, 1599. [Google Scholar] [CrossRef]
- Okabe, T.; Kumagai, M.; Nakajima, Y.; Shirotake, S.; Kodaira, K.; Oyama, M.; Ueno, M.; Ikeda, M. The Impact of HIF1α on the Per2 Circadian Rhythm in Renal Cancer Cell Lines. PLoS ONE 2014, 9, e109693. [Google Scholar] [CrossRef]
- Peek, C.B.; Levine, D.C.; Cedernaes, J.; Taguchi, A.; Kobayashi, Y.; Tsai, S.J.; Bonar, N.A.; McNulty, M.R.; Ramsey, K.M.; Bass, J. Circadian Clock Interaction with HIF1α Mediates Oxygenic Metabolism and Anaerobic Glycolysis in Skeletal Muscle. Cell Metab. 2017, 25, 86–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutkowska, A.; Szmyd, B.; Kaszkowiak, M.; Domańska-Senderowska, D.; Pastuszak-Lewandoska, D.; Brzeziańska-Lasota, E.; Kordiak, J.; Antczak, A. Expression of Inflammatory Interleukins and Selected MiRNAs in Non-Small Cell Lung Cancer. Sci. Rep. 2021, 11, 5092. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef] [Green Version]
- Czarnecka, K.H.; Szmyd, B.; Barańska, M.; Kaszkowiak, M.; Kordiak, J.; Antczak, A.; Pastuszak-Lewandoska, D.; Brzeziańska-Lasota, E. A Strong Decrease in TIMP3 Expression Mediated by the Presence of MiR-17 and 20a Enables Extracellular Matrix Remodeling in the NSCLC Lesion Surroundings. Front. Oncol. 2019, 9, 1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Li, J.; Cairns, M.J. Identifying MiRNAs, Targets and Functions. Brief. Bioinform. 2014, 15, 1–19. [Google Scholar] [CrossRef]
- Vishnoi, A.; Rani, S. MiRNA Biogenesis and Regulation of Diseases: An Overview. Methods Mol. Biol. 2017, 1509, 1–10. [Google Scholar] [CrossRef]
- Kulshreshtha, R.; Ferracin, M.; Wojcik, S.E.; Garzon, R.; Alder, H.; Agosto-Perez, F.J.; Davuluri, R.; Liu, C.-G.; Croce, C.M.; Negrini, M.; et al. A MicroRNA Signature of Hypoxia. Mol. Cell Biol. 2007, 27, 1859–1867. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Zhao, L.; Yan, J.; Ren, X.; Zhu, K.; Gao, T.; Du, X.; Luo, H.; Li, Z.; Xu, M. MicroRNA Expression Profile Is Altered in the Upper Airway Skeletal Muscle Tissue of Patients with Obstructive Sleep Apnea-Hypopnea Syndrome. J. Int. Med. Res. 2019, 47, 4163–4182. [Google Scholar] [CrossRef] [Green Version]
- MiRDB—MicroRNA Target Prediction Database. Available online: http://www.mirdb.org/ (accessed on 6 December 2022).
- Yang, M.Y.; Lin, P.W.; Lin, H.C.; Lin, P.M.; Chen, I.Y.; Friedman, M.; Hung, C.F.; Salapatas, A.M.; Lin, M.C.; Lin, S.F. Alternations of Circadian Clock Genes Expression and Oscillation in Obstructive Sleep Apnea. J. Clin. Med. 2019, 8, 1634. [Google Scholar] [CrossRef] [Green Version]
- Masri, S. Sirtuin-Dependent Clock Control: New Advances in Metabolism, Aging and Cancer. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 521–527. [Google Scholar] [CrossRef]
- Wang, R.H.; Zhao, T.; Cui, K.; Hu, G.; Chen, Q.; Chen, W.; Wang, X.W.; Soto-Gutierrez, A.; Zhao, K.; Deng, C.X. Negative Reciprocal Regulation between Sirt1 and Per2 Modulates the Circadian Clock and Aging. Sci. Rep. 2016, 6, 28633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakahata, Y.; Kaluzova, M.; Grimaldi, B.; Sahar, S.; Hirayama, J.; Chen, D.; Guarente, L.P.; Sassone-Corsi, P. The NAD+-Dependent Deacetylase SIRT1 Modulates CLOCK-Mediated Chromatin Remodeling and Circadian Control. Cell 2008, 134, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.J.; Liaw, S.F.; Lin, C.C.; Chiu, C.H.; Lin, M.W.; Chang, F.T. Effect of Nasal CPAP on SIRT1 and Endothelial Function in Obstructive Sleep Apnea Syndrome. Lung 2015, 193, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Tam, C.S.; Wong, M.; McBain, R.; Bailey, S.; Waters, K.A. Inflammatory Measures in Children with Obstructive Sleep Apnoea. J. Paediatr. Child Health 2006, 42, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Zheng, X. Tumor Necrosis Factor Alpha Is a Promising Circulating Biomarker for the Development of Obstructive Sleep Apnea Syndrome: A Meta-Analysis. Oncotarget 2017, 8, 27616–27626. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.M.; Pan, L.; Ren, D.Q.; Du, C.J.; Guo, Y.Z. Effects of Continuous Positive Airway Pressure Therapy on Systemic Inflammation in Obstructive Sleep Apnea: A Meta-Analysis. Sleep Med. 2013, 14, 1139–1150. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hu, R.; Ren, X.; He, J. Interleukin-8 Concentrations in Obstructive Sleep Apnea Syndrome: A Systematic Review and Meta-Analysis. Bioengineered 2021, 12, 10666–10681. [Google Scholar] [CrossRef]
- Kheirandish-Gozal, L.; Gozal, D. Obstructive Sleep Apnea and Inflammation: Proof of Concept Based on Two Illustrative Cytokines. Int. J. Mol. Sci. 2019, 20, 459. [Google Scholar] [CrossRef] [Green Version]
- Keller, M.; Mazuch, J.; Abraham, U.; Eom, G.D.; Herzog, E.D.; Volk, H.D.; Kramer, A.; Maier, B. A Circadian Clock in Macrophages Controls Inflammatory Immune Responses. Proc. Natl. Acad. Sci. USA 2009, 106, 21407–21412. [Google Scholar] [CrossRef] [Green Version]
- Entzian, P.; Linnemann, K.; Schlaak, M.; Zabel, P. Obstructive Sleep Apnea Syndrome and Circadian Rhythms of Hormones and Cytokines. Am. J. Respir. Crit. Care Med. 1996, 153, 1080–1086. [Google Scholar] [CrossRef]
- Smith, D.F.; Hossain, M.M.; Hura, A.; Huang, G.; McConnell, K.; Ishman, S.L.; Amin, R.S. Inflammatory Milieu and Cardiovascular Homeostasis in Children with Obstructive Sleep Apnea. Sleep 2017, 40, zsx022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Song, S.; Wang, Y.; Huang, C.; Zhang, F.; Liu, J.; Hong, J.S. Low-Grade Inflammation Aggravates Rotenone Neurotoxicity and Disrupts Circadian Clock Gene Expression in Rats. Neurotox. Res. 2019, 35, 421–431. [Google Scholar] [CrossRef]
- Walton, Z.E.; Patel, C.H.; Brooks, R.C.; Yu, Y.; Ibrahim-Hashim, A.; Riddle, M.; Porcu, A.; Jiang, T.; Ecker, B.L.; Tameire, F.; et al. Acid Suspends the Circadian Clock in Hypoxia through Inhibition of MTOR. Cell 2018, 174, 72–87.e32. [Google Scholar] [CrossRef] [Green Version]
- Roenneberg, T.; Allebrandt, K.V.; Merrow, M.; Vetter, C. Social Jetlag and Obesity. Curr. Biol. 2012, 22, 939–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, M.J.; Moffitt, T.E.; Gregory, A.M.; Goldman-Mellor, S.; Nolan, P.M.; Poulton, R.; Caspi, A. Social Jetlag, Obesity and Metabolic Disorder: Investigation in a Cohort Study. Int. J. Obes. 2015, 39, 842–848. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malicki, M.; Karuga, F.F.; Szmyd, B.; Sochal, M.; Gabryelska, A. Obstructive Sleep Apnea, Circadian Clock Disruption, and Metabolic Consequences. Metabolites 2023, 13, 60. https://doi.org/10.3390/metabo13010060
Malicki M, Karuga FF, Szmyd B, Sochal M, Gabryelska A. Obstructive Sleep Apnea, Circadian Clock Disruption, and Metabolic Consequences. Metabolites. 2023; 13(1):60. https://doi.org/10.3390/metabo13010060
Chicago/Turabian StyleMalicki, Mikołaj, Filip Franciszek Karuga, Bartosz Szmyd, Marcin Sochal, and Agata Gabryelska. 2023. "Obstructive Sleep Apnea, Circadian Clock Disruption, and Metabolic Consequences" Metabolites 13, no. 1: 60. https://doi.org/10.3390/metabo13010060