
Plant Metabolomics as a Tool for Detecting Adulterants in Edible Plant: A Case Study of Allium ursinum
 , , , and
, , , and
Abstract
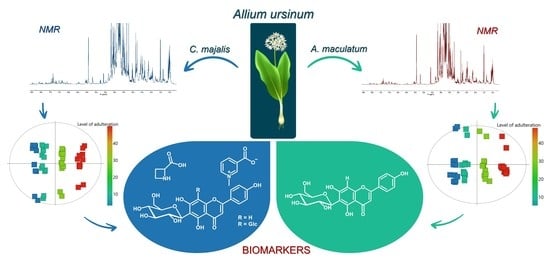
:
1. Introduction
2. Materials and Methods
2.1. Plant Sample Procurement
2.2. Sample Preparation for NMR Fingerprinting
2.3. NMR Analysis
2.4. Multivariate Data Analysis
2.5. Purification of Biomarkers from Convallaria majalis
2.6. Purification of Biomarkers from Arum maculatum
2.7. LC-MS Analysis of Biomarkers
3. Results and Discussion
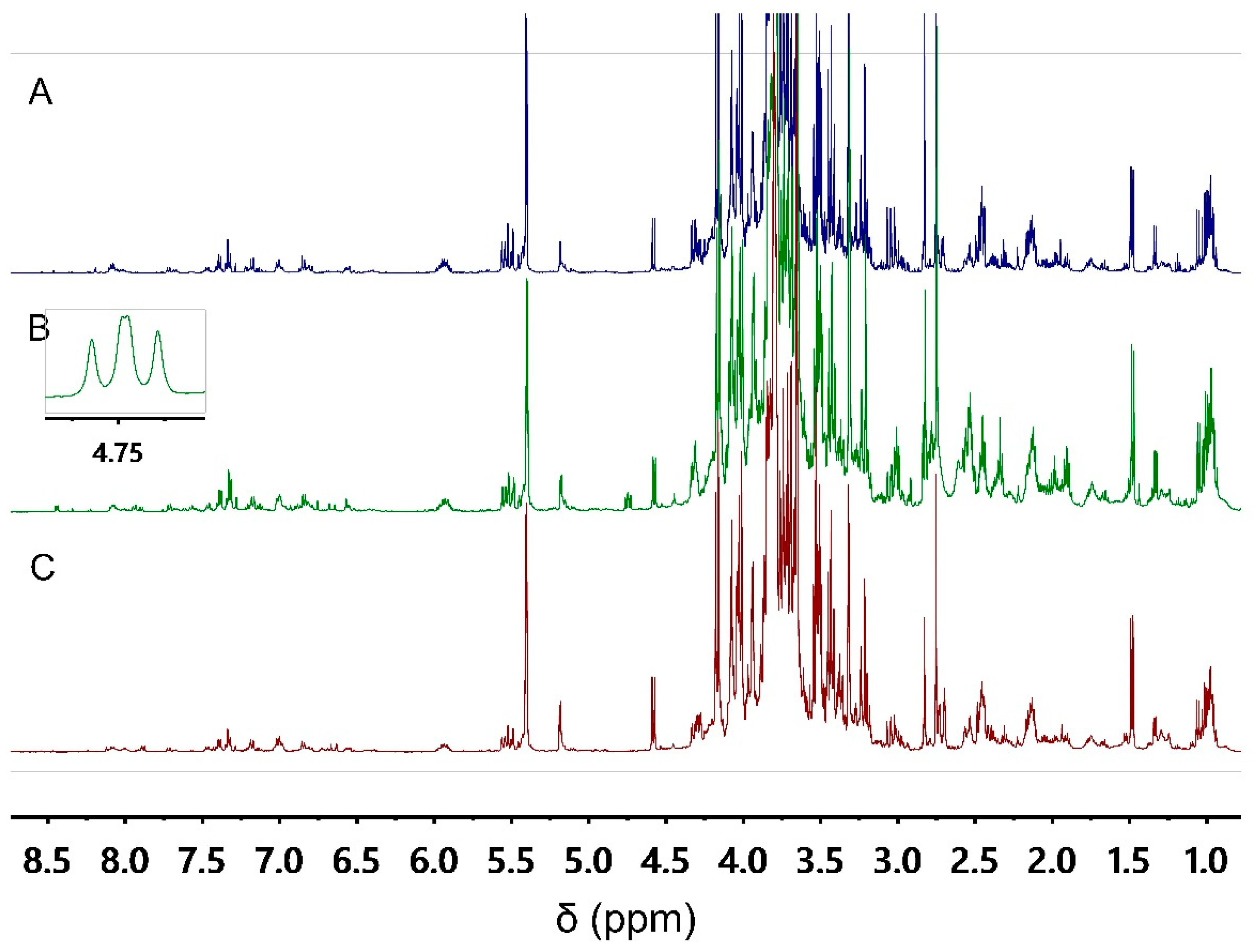
3.1. Metabolomics Fingerprinting
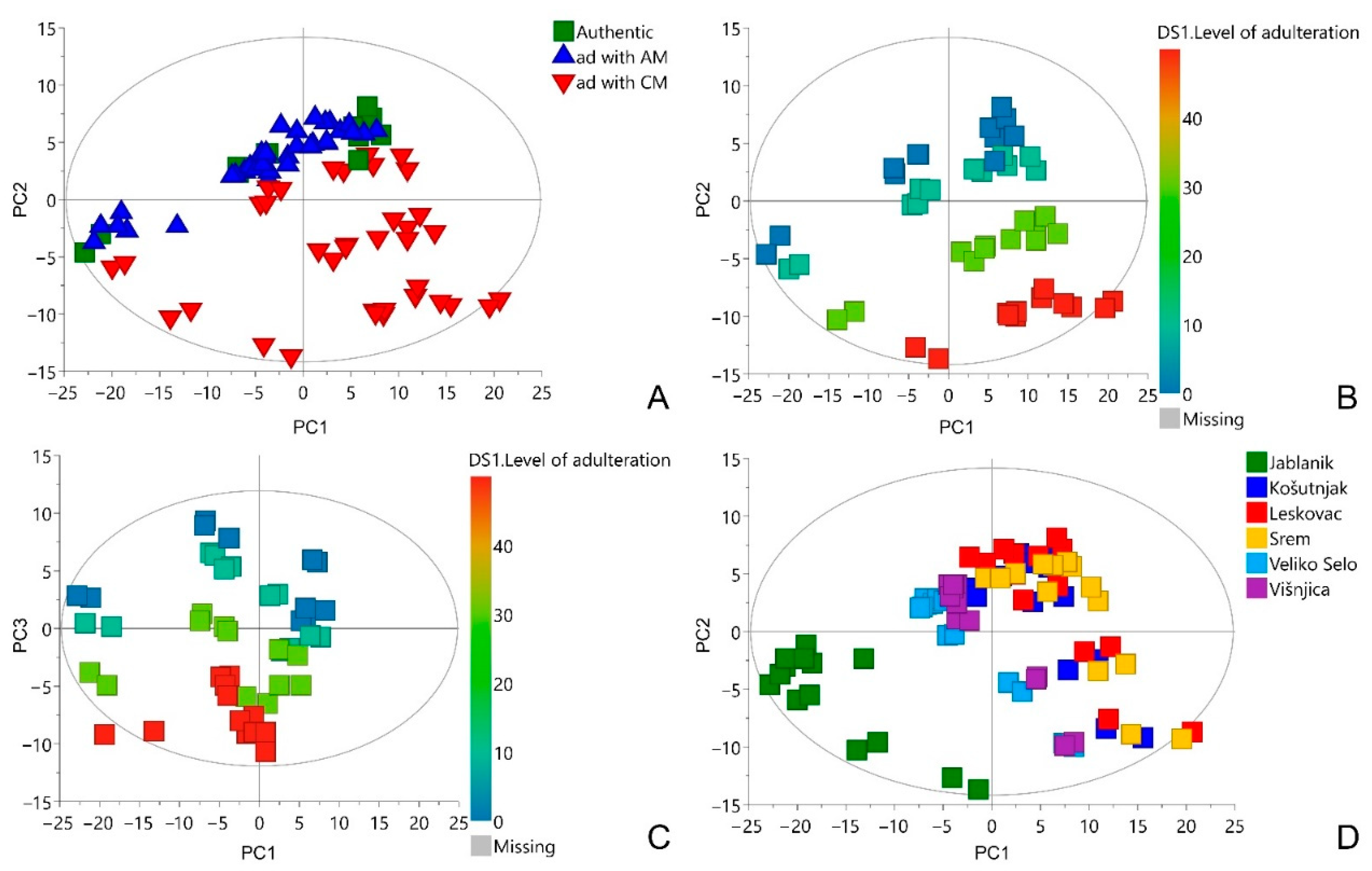
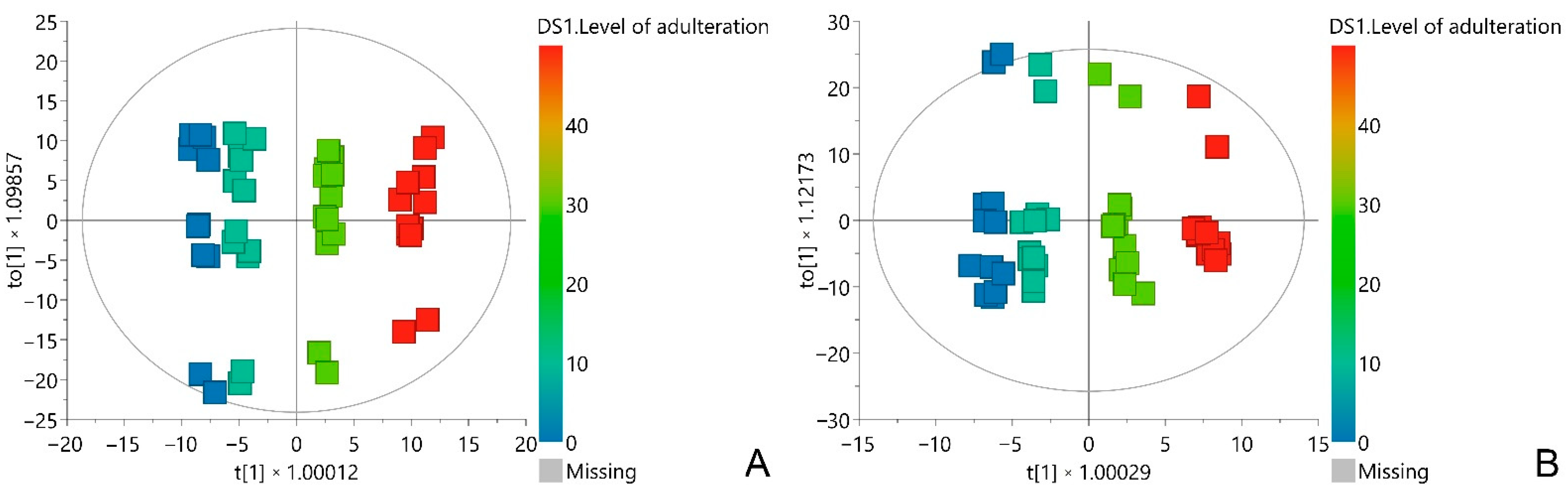
3.2. Multivariate Data Analysis
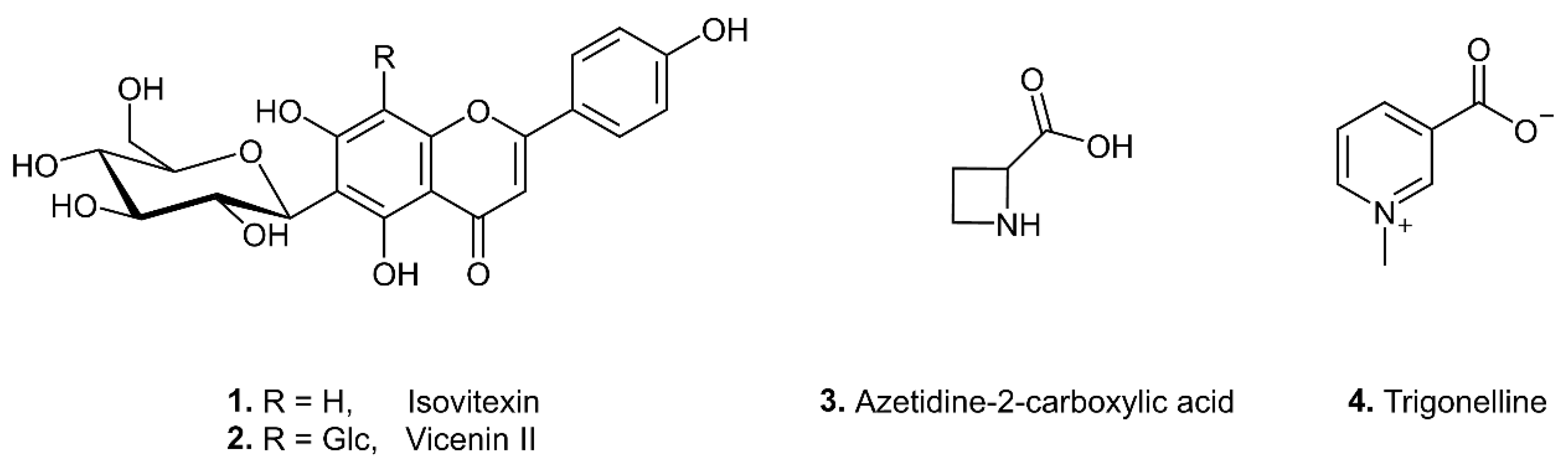
3.3. Structure Determination of Biomarkers of Adulteration
3.4. Relationships between Biomarkers and Toxic principles in the Adulterants
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gođevac, D.; Vujisić, L.; Mojović, M.; Ignjatović, A.; Spasojević, I.; Vajs, V. Evaluation of Antioxidant Capacity of Allium ursinum L. Volatile Oil and Its Effect on Membrane Fluidity. Food Chem. 2008, 107, 1692–1700. [Google Scholar] [CrossRef]
- Rather, I.A.; Koh, W.Y.; Paek, W.K.; Lim, J. The Sources of Chemical Contaminants in Food and Their Health Implications. Front. Pharmacol. 2017, 8, 830. [Google Scholar] [CrossRef] [PubMed]
- Genualdi, S.; MacMahon, S.; Robbins, K.; Farris, S.; Shyong, N.; DeJager, L. Method Development and Survey of Sudan I–IV in Palm Oil and Chilli Spices in the Washington, DC, Area. Food Addit. Contam. Part A 2016, 33, 583–591. [Google Scholar] [CrossRef]
- Maeda, K.; Idehara, R.; Kusaka, S. Mistaken Identity: Severe Vomiting, Bradycardia and Hypotension after Eating a Wild Herb. Clin. Toxicol. 2012, 50, 532–533. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A. Bitter Pill to Swallow: A Case of Accidental Poisoning with Digitalis Purpurea. Case Rep. 2010, 2010, bcr0120102633. [Google Scholar] [CrossRef] [PubMed]
- Mezzasalma, V.; Ganopoulos, I.; Galimberti, A.; Cornara, L.; Ferri, E.; Labra, M. Poisonous or Non-Poisonous Plants? DNA-Based Tools and Applications for Accurate Identification. Int. J. Legal Med. 2017, 131, 1–19. [Google Scholar] [CrossRef]
- American Botanical Council (ABC). Botanical Adulterants Program Content. Available online: http://abc.herbalgram.org/site/PageServer (accessed on 11 July 2022).
- Govindaraghavan, S. Pharmacopeial HPLC Identification Methods Are Not Sufficient to Detect Adulterations in Commercial Bilberry (Vaccinium myrtillus) Extracts. Anthocyanin Profile Provides Additional Clues. Fitoterapia 2014, 99, 124–138. [Google Scholar] [CrossRef]
- Villani, T.S.; Reichert, W.; Ferruzzi, M.G.; Pasinetti, G.M.; Simon, J.E.; Wu, Q. Chemical Investigation of Commercial Grape Seed Derived Products to Assess Quality and Detect Adulteration. Food Chem. 2015, 170, 271–280. [Google Scholar] [CrossRef]
- Wielogorska, E.; Chevallier, O.; Black, C.; Galvin-King, P.; Delêtre, M.; Kelleher, C.T.; Haughey, S.A.; Elliott, C.T. Development of a Comprehensive Analytical Platform for the Detection and Quantitation of Food Fraud Using a Biomarker Approach. The Oregano Adulteration Case Study. Food Chem. 2018, 239, 32–39. [Google Scholar] [CrossRef]
- Wolfender, J.-L.; Rudaz, S.; Hae Choi, Y.; Kyong Kim, H. Plant Metabolomics: From Holistic Data to Relevant Biomarkers. Curr. Med. Chem. 2013, 20, 1056–1090. [Google Scholar] [CrossRef]
- Alonso, A.; Marsal, S.; JuliÃ, A. Analytical Methods in Untargeted Metabolomics: State of the Art in 2015. Front. Bioeng. Biotechnol. 2015, 3, 23. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Choi, Y.H.; Verpoorte, R. NMR-Based Plant Metabolomics: Where do we Stand, where do we Go? Trends Biotechnol. 2011, 29, 267–275. [Google Scholar] [CrossRef]
- Sobolewska, D.; Podolak, I.; Makowska-Wąs, J. Allium ursinum: Botanical, Phytochemical and Pharmacological Overview. Phytochem. Rev. 2015, 14, 81–97. [Google Scholar] [CrossRef]
- Senizza, B.; Rocchetti, G.; Okur, M.A.; Zengin, G.; Yıldıztugay, E.; Ak, G.; Montesano, D.; Lucini, L. Phytochemical Profile and Biological Properties of Colchicum triphyllum (Meadow Saffron). Foods 2020, 9, 457. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, G.; Clément, J.; Fezard, J.B.; Laribi, S. Intoxication Au Colchique Par Confusion Avec l’ail Des Ours. Rev. Med. Interne 2022, 43, 559–561. [Google Scholar] [CrossRef] [PubMed]
- Touiti, N.; Houssaini, T.S.; Achour, S. Overview on Pharmacovigilance of Nephrotoxic Herbal Medicines Used Worldwide. Clin. Phytosci. 2021, 7, 9. [Google Scholar] [CrossRef]
- Focke, F.; Haase, I.; Fischer, M. DNA-Based Identification of Spices: DNA Isolation, Whole Genome Amplification, and Polymerase Chain Reaction. J. Agric. Food Chem. 2011, 59, 513–520. [Google Scholar] [CrossRef]
- Zoltani, C.K. Cardiovascular Toxicity. In Veterinary and Human Toxicology; Elsevier: Amsterdam, The Netherlands, 2012; pp. 235–245. [Google Scholar]
- Ceylan, F.; Akar Sahingoz, S. Using Ethnobotanical Plants in Food Preparation: Cuckoo Pint (Arum maculatum L.). Int. J. Gastron. Food Sci. 2022, 29, 100529. [Google Scholar] [CrossRef]
- Prakas Raju, K.N.J.; Goel, K.; Anandhi, D.; Pandit, V.; Surendar, R.; Sasikumar, M. Wild Tuber Poisoning: Arum maculatum—A Rare Case Report. Int. J. Crit. Illn. Inj. Sci. 2018, 8, 111–114. [Google Scholar] [CrossRef]
- Erbil, N.; Arslan, M.; Murathan, Z. Antioxidant, antimicrobial, and antimutagenic effects and biochemical contents of Arum maculatum L. That is a medical plant from turkish flora. Fresenius Environ. Bull. 2018, 27, 8709. [Google Scholar]
- Mandrone, M.; Marincich, L.; Chiocchio, I.; Petroli, A.; Gođevac, D.; Maresca, I.; Poli, F. NMR-Based Metabolomics for Frauds Detection and Quality Control of Oregano Samples. Food Control 2021, 127, 108141. [Google Scholar] [CrossRef]
- Zhu, C.; Vitali, B.; Donders, G.; Parolin, C.; Li, Y.; Laghi, L. Univariate Statistical Analysis as a Guide to 1H-NMR Spectra Signal Assignment by Visual Inspection. Metabolites 2019, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Owuor, P.O.; Obaga, S.O.; Othieno, C.O. The Effects of Altitude on the Chemical Composition of Black Tea. J. Sci. Food Agric. 1990, 50, 9–17. [Google Scholar] [CrossRef]
- Anđelković, B.; Vujisić, L.; Vučković, I.; Tešević, V.; Vajs, V.; Gođevac, D. Metabolomics Study of Populus Type Propolis. J. Pharm. Biomed. Anal. J. 2017, 135, 217–226. [Google Scholar] [CrossRef]
- Mansour, G.; Ghanem, C.; Mercenaro, L.; Nassif, N.; Hassoun, G.; del Caro, A. Effects of Altitude on the Chemical Composition of Grapes and Wine: A Review. OENO One 2022, 56, 227–239. [Google Scholar] [CrossRef]
- Wiklund, S.; Johansson, E.; Sjöström, L.; Mellerowicz, E.J.; Edlund, U.; Shockcor, J.P.; Gottfries, J.; Moritz, T.; Trygg, J. Visualization of GC/TOF-MS-Based Metabolomics Data for Identification of Biochemically Interesting Compounds Using OPLS Class Models. Anal. Chem. 2008, 80, 115–122. [Google Scholar] [CrossRef]
- Triba, M.N.; le Moyec, L.; Amathieu, R.; Goossens, C.; Bouchemal, N.; Nahon, P.; Rutledge, D.N.; Savarin, P. PLS/OPLS Models in Metabolomics: The Impact of Permutation of Dataset Rows on the K-Fold Cross-Validation Quality Parameters. Mol. Biosyst. 2015, 11, 13–19. [Google Scholar] [CrossRef]
- Tchoumtchoua, J.; Mathiron, D.; Pontarin, N.; Gagneul, D.; van Bohemen, A.-I.; Otogo N’nang, E.; Mesnard, F.; Petit, E.; Fontaine, J.-X.; Molinié, R.; et al. Phenolic Profiling of Flax Highlights Contrasting Patterns in Winter and Spring Varieties. Molecules 2019, 24, 4303. [Google Scholar] [CrossRef]
- Shie, J.-J.; Chen, C.-A.; Lin, C.-C.; Ku, A.F.; Cheng, T.-J.R.; Fang, J.-M.; Wong, C.-H. Regioselective Synthesis of Di-C-Glycosylflavones Possessing Anti-Inflammation Activities. Org. Biomol. Chem. 2010, 8, 4451. [Google Scholar] [CrossRef]
- Mabry, T.J.; Markham, K.R.; Thomas, M.B. The Systematic Identification of Flavonoids; Springer: Berlin/Heidelberg, Germany, 1970. [Google Scholar]
- Abad-García, B.; Garmón-Lobato, S.; Berrueta, L.A.; Gallo, B.; Vicente, F. On Line Characterization of 58 Phenolic Compounds in Citrus Fruit Juices from Spanish Cultivars by High-Performance Liquid Chromatography with Photodiode-Array Detection Coupled to Electrospray Ionization Triple Quadrupole Mass Spectrometry. Talanta 2012, 99, 213–224. [Google Scholar] [CrossRef]
- Abu-Reidah, I.M.; Ali-Shtayeh, M.S.; Jamous, R.M.; Arráez-Román, D.; Segura-Carretero, A. Comprehensive Metabolite Profiling of Arum palaestinum (Araceae) Leaves by Using Liquid Chromatography–Tandem Mass Spectrometry. Int. Food Res. J. 2015, 70, 74–86. [Google Scholar] [CrossRef]
- Brito, A.; Ramirez, J.; Areche, C.; Sepúlveda, B.; Simirgiotis, M. HPLC-UV-MS Profiles of Phenolic Compounds and Antioxidant Activity of Fruits from Three Citrus Species Consumed in Northern Chile. Molecules 2014, 19, 17400–17421. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.B.; Turatti, I.C.C.; Gouveia, D.R.; Ernst, M.; Teixeira, S.P.; Lopes, N.P. Mass Spectrometry of Flavonoid Vicenin-2, Based Sunlight Barriers in Lychnophora Species. Sci. Rep. 2014, 4, 4309. [Google Scholar] [CrossRef] [PubMed]
- Meng, C.; Liu, R.; Wang, W.; Guo, W.; Ma, H.; Xie, S.; Liu, Y.; Wang, C. Metabolic Profiling Comparison of Isovitexin in Normal and Kidney Stone Model Rats by Ultra-high-performance Liquid Chromatography Coupled to Quadrupole Time-of-flight Mass Spectrometry. J. Sep. Sci. 2020, 43, 2363–2379. [Google Scholar] [CrossRef]
- Futamura, Y.; Kurokawa, M.; Obata, R.; Nishiyama, S.; Sugai, T. Efficient Route to (S)-Azetidine-2-Carboxylic Acid. Biosci. Biotechnol. Biochem. 2014, 69, 1892–1897. [Google Scholar] [CrossRef]
- Farag, M.A.; Porzel, A.; Wessjohann, L.A. Unraveling the Active Hypoglycemic Agent Trigonelline in Balanites aegyptiaca Date Fruit Using Metabolite Fingerprinting by NMR. J. Pharm. Biomed. Anal. 2015, 115, 383–387. [Google Scholar] [CrossRef]
- He, M.; Min, J.W.; Kong, W.L.; He, X.H.; Li, J.X.; Peng, B.W. A review on the pharmacological effects of vitexin and isovitexin. Fitoterapia 2016, 115, 74–85. [Google Scholar] [CrossRef]
- Mohamadi, N.; Sharififar, F.; Pournamdari, M.; Ansari, M. A Review on Biosynthesis, Analytical Techniques, and Pharmacological Activities of Trigonelline as a Plant Alkaloid. J. Diet. Suppl. 2018, 15, 207–222. [Google Scholar] [CrossRef]
- Couty, F.; Evano, G. Azetidine-2-carboxylic acid. From lily of the valley to key pharmaceuticals. A jubilee review. Org. Prep. Proced. Int. 2006, 38, 427–465. [Google Scholar] [CrossRef]
- The European Agency for the Evaluation of Medicinal Products. Available online: https://ema.europa.eu/documents/mrl-report/convallaria-majalis-summary-report-committee-veterinary-medicinal-products_en.pdf (accessed on 16 August 2022).
- Vučković, I.; Rapinoja, M.L.; Vaismaa, M.; Vanninen, P.; Koskela, H. Application of comprehensive NMR-based analysis strategy in annotation, isolation and structure elucidation of low molecular weight metabolites of Ricinus communis seeds. Phytochem. Anal. 2016, 27, 64–72. [Google Scholar] [CrossRef]
- Nagana Gowda, G.A.; Raftery, D. Can NMR Solve Some Significant Challenges in Metabolomics? J. Magn. Reson. 2015, 260, 144–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Name | No. of Components | R2 | Q2 | p (CV-ANOVA) | F (CV-ANOVA) |
|---|---|---|---|---|---|
| PCA | 10 | 0.925 | 0.834 | - | - |
| OPLS, adulteration with CM | 1 + 2 | 0.992 | 0.988 | 2 × 10−37 | 543 |
| OPLS, adulteration with AM | 1 + 2 | 0.989 | 0.981 | 9 × 10−34 | 357 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivanović, S.; Simić, K.; Lekić, S.; Jadranin, M.; Vujisić, L.; Gođevac, D. Plant Metabolomics as a Tool for Detecting Adulterants in Edible Plant: A Case Study of Allium ursinum. Metabolites 2022, 12, 849. https://doi.org/10.3390/metabo12090849
Ivanović S, Simić K, Lekić S, Jadranin M, Vujisić L, Gođevac D. Plant Metabolomics as a Tool for Detecting Adulterants in Edible Plant: A Case Study of Allium ursinum. Metabolites. 2022; 12(9):849. https://doi.org/10.3390/metabo12090849
Chicago/Turabian StyleIvanović, Stefan, Katarina Simić, Stefan Lekić, Milka Jadranin, Ljubodrag Vujisić, and Dejan Gođevac. 2022. "Plant Metabolomics as a Tool for Detecting Adulterants in Edible Plant: A Case Study of Allium ursinum" Metabolites 12, no. 9: 849. https://doi.org/10.3390/metabo12090849