
Evaluation of the Links between Lamb Feed Efficiency and Rumen and Plasma Metabolomic Data

 , ,
, ,
Abstract
:1. Introduction
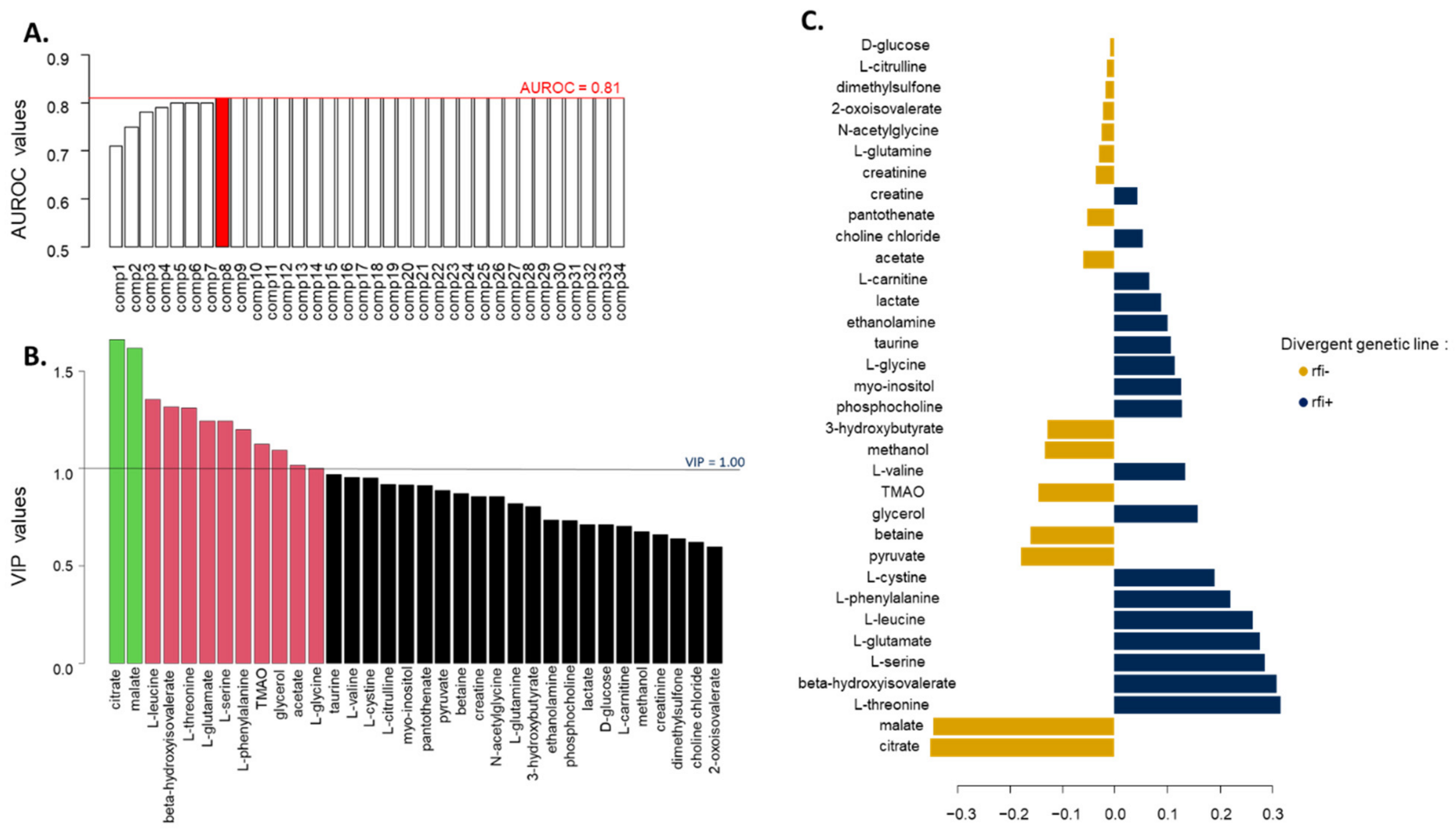
2. Results
2.1. Zootechnical Parameters
2.1.1. Growth Phase under a 100% Concentrate Diet
2.1.2. Finishing Phase under a Mixed Diet
2.2. Plasma Metabolites
2.2.1. Growth Phase under a 100% Concentrate Diet
2.2.2. Finishing Phase under a Mixed Diet
2.3. Rumen Metabolites
2.3.1. Growth Phase with a 100% Concentrate Diet
2.3.2. Finishing Phase with a Mixed Diet
2.4. Correlations between Rumen and Plasma Metabolites
3. Discussion
3.1. Phenotypes
3.2. Plasma Metabolites
3.2.1. Amino Acids
3.2.2. Organic Acids
3.2.3. Integration
3.3. Rumen Metabolites
4. Materials and Methods
4.1. Animals
4.2. Diets
4.3. Traits
4.4. Sampling
4.5. NMR Analysis
4.5.1. Chemical Analysis
4.5.2. Bioinformatic Analysis
4.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raineri, C.; Stivari, T.S.S.; Gameiro, A.H. Lamb Production Costs: Analyses of Composition and Elasticities Analysis of Lamb Production Costs. Asian Australas. J. Anim. Sci 2015, 28, 1209–1215. [Google Scholar] [CrossRef] [Green Version]
- Dumont, B.; Groot, J.C.J.; Tichit, M. Review: Make Ruminants Green Again—How Can Sustainable Intensification and Agroecology Converge for a Better Future? Animal 2018, 12, s210–s219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paganoni, B.; Rose, G.; Macleay, C.; Jones, C.; Brown, D.J.; Kearney, G.; Ferguson, M.; Thompson, A.N. More Feed Efficient Sheep Produce Less Methane and Carbon Dioxide When Eating High-Quality Pellets. J. Anim. Sci. 2017, 95, 3839–3850. [Google Scholar] [CrossRef] [PubMed]
- Løvendahl, P.; Difford, G.F.; Li, B.; Chagunda, M.G.G.; Huhtanen, P.; Lidauer, M.H.; Lassen, J.; Lund, P. Review: Selecting for Improved Feed Efficiency and Reduced Methane Emissions in Dairy Cattle. Animal 2018, 12, s336–s349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthur, P.F.; Archer, J.A.; Johnston, D.J.; Herd, R.M.; Richardson, E.C.; Parnell, P.F. Genetic and Phenotypic Variance and Covariance Components for Feed Intake, Feed Efficiency, and Other Postweaning Traits in Angus Cattle. J. Anim. Sci. 2001, 79, 2805. [Google Scholar] [CrossRef] [Green Version]
- Berry, D.P.; Crowley, J.J. Residual Intake and Body Weight Gain: A New Measure of Efficiency in Growing Cattle. J. Anim. Sci. 2012, 90, 109–115. [Google Scholar] [CrossRef]
- Cammack, K.M.; Leymaster, K.A.; Jenkins, T.G.; Nielsen, M.K. Estimates of Genetic Parameters for Feed Intake, Feeding Behavior, and Daily Gain in Composite Ram Lambs. J. Anim. Sci. 2005, 83, 777–785. [Google Scholar] [CrossRef] [Green Version]
- Johnson, P.; Miller, S.; Knowler, K. Preliminary Investigations into the Trait of Residual Energy Intake in Sheep. Proc. N. Zldn. Soc. Anim. Prod. 2016, 76, 5. [Google Scholar]
- Marie-Etancelin, C.; Francois, D.; Weisbecker, J.; Marcon, D.; Moreno-Romieux, C.; Bouvier, F.; Tortereau, F. Detailed Genetic Analysis of Feeding Behaviour in Romane Lambs and Links with Residual Feed Intake. J. Anim. Breed. Genet. 2019, 136, 174–182. [Google Scholar] [CrossRef]
- Tortereau, F.; Marie-Etancelin, C.; Weisbecker, J.-L.; Marcon, D.; Bouvier, F.; Moreno-Romieux, C.; François, D. Genetic Parameters for Feed Efficiency in Romane Rams and Responses to Single-Generation Selection. Animal 2020, 14, 681–687. [Google Scholar] [CrossRef]
- Gunsett, F.C. Linear Index Selection to Improve Traits Defined as Ratios. J. Anim. Sci. 1984, 59, 1185–1193. [Google Scholar] [CrossRef]
- Meale, S.J.; Morgavi, D.P.; Cassar-Malek, I.; Andueza, D.; Ortigues-Marty, I.; Robins, R.J.; Schiphorst, A.-M.; Laverroux, S.; Graulet, B.; Boudra, H.; et al. Exploration of Biological Markers of Feed Efficiency in Young Bulls. J. Agric. Food Chem. 2017, 65, 9817–9827. [Google Scholar] [CrossRef]
- De Paula, E.F.E.; de Souza, D.F.; Monteiro, A.L.G.; de Santana, M.H.A.; Gilaverte, S.; Rossi Junior, P.; Locatelli Dittrich, R. Residual Feed Intake and Hematological and Metabolic Blood Profiles of Lle de France Lambs. R. Bras. Zootec. 2013, 42, 806–812. [Google Scholar] [CrossRef] [Green Version]
- Perea, K.; Perz, K.; Olivo, S.K.; Williams, A.; Lachman, M.; Ishaq, S.L.; Thomson, J.; Yeoman, C.J. Feed Efficiency Phenotypes in Lambs Involve Changes in Ruminal, Colonic, and Small-Intestine-Located Microbiota. J. Anim. Sci. 2017, 95, 2585. [Google Scholar] [CrossRef] [Green Version]
- Muir, S.K.; Linden, N.; Knight, M.; Behrendt, R.; Kearney, G. Sheep Residual Feed Intake and Feeding Behaviour: Are ‘Nibblers’ or ‘Binge Eaters’ More Efficient? Anim. Prod. Sci. 2018, 58, 1459. [Google Scholar] [CrossRef]
- Thompson, S.; Schaefer, A.L.; Crow, G.H.; Basarab, J.; Colyn, J.; Ominski, K. Relationship between Residual Feed Intake and Radiated Heat Loss Using Infrared Thermography in Young Beef Bulls. J. Therm. Biol. 2018, 78, 304–311. [Google Scholar] [CrossRef]
- Montanholi, Y.R.; Swanson, K.C.; Palme, R.; Schenkel, F.S.; McBride, B.W.; Lu, D.; Miller, S.P. Assessing Feed Efficiency in Beef Steers through Feeding Behavior, Infrared Thermography and Glucocorticoids. Animal 2010, 4, 692–701. [Google Scholar] [CrossRef]
- Cantalapiedra-Hijar, G.; Abo-Ismail, M.; Carstens, G.E.; Guan, L.L.; Hegarty, R.; Kenny, D.A.; McGee, M.; Plastow, G.; Relling, A.; Ortigues-Marty, I. Review: Biological Determinants of between-Animal Variation in Feed Efficiency of Growing Beef Cattle. Animal 2018, 12, s321–s335. [Google Scholar] [CrossRef] [Green Version]
- Herd, R.M.; Oddy, V.H.; Richardson, E.C. Biological Basis for Variation in Residual Feed Intake in Beef Cattle. 1. Review of Potential Mechanisms. Aust. J. Exp. Agric. 2004, 44, 423. [Google Scholar] [CrossRef]
- Bica, R.; Palarea-Albaladejo, J.; Kew, W.; Uhrin, D.; Pacheco, D.; Macrae, A.; Dewhurst, R.J. Nuclear Magnetic Resonance to Detect Rumen Metabolites Associated with Enteric Methane Emissions from Beef Cattle. Sci. Rep. 2020, 10, 5578. [Google Scholar] [CrossRef] [Green Version]
- Goldansaz, S.A.; Markus, S.; Berjanskii, M.; Rout, M.; Guo, A.C.; Wang, Z.; Plastow, G.; Wishart, D.S. Candidate Serum Metabolite Biomarkers of Residual Feed Intake and Carcass Merit in Sheep. J. Anim. Sci. 2020, 98, skaa298. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Greenwood, P.L.; Cockett, N.E.; Hadfield, T.S.; Vuocolo, T.; Byrne, K.; White, J.D.; Tellam, R.L.; Schirra, H.J. Impacts of the Callipyge Mutation on Ovine Plasma Metabolites and Muscle Fibre Type. PLoS ONE 2014, 9, e99726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karisa, B.K.; Thomson, J.; Wang, Z.; Li, C.; Montanholi, Y.R.; Miller, S.P.; Moore, S.S.; Plastow, G.S. Plasma Metabolites Associated with Residual Feed Intake and Other Productivity Performance Traits in Beef Cattle. Livest. Sci. 2014, 165, 200–211. [Google Scholar] [CrossRef]
- Redden, R.R.; Surber, L.M.M.; Roeder, B.L.; Nichols, B.M.; Paterson, J.A.; Kott, R.W. Residual Feed Efficiency Established in a Post-Weaning Growth Test May Not Result in More Efficient Ewes on the Range. Small Rumin. Res. 2011, 96, 155–159. [Google Scholar] [CrossRef]
- Oliveira, L.F.; Ruggieri, A.C.; Branco, R.H.; Cota, O.L.; Canesin, R.C.; Costa, H.J.U.; Mercadante, M.E.Z. Feed Efficiency and Enteric Methane Production of Nellore Cattle in the Feedlot and on Pasture. Anim. Prod. Sci. 2018, 58, 886. [Google Scholar] [CrossRef]
- Owens, F.N.; Dubeski, P.; Hanson, C.F. Factors That Alter the Growth and Development of Ruminants. J. Anim. Sci. 1993, 71, 3138–3150. [Google Scholar] [CrossRef]
- Beauclercq, S.; Nadal-Desbarats, L.; Hennequet-Antier, C.; Gabriel, I.; Tesseraud, S.; Calenge, F.; Le Bihan-Duval, E.; Mignon-Grasteau, S. Relationships between Digestive Efficiency and Metabolomic Profiles of Serum and Intestinal Contents in Chickens. Sci. Rep. 2018, 8, 6678. [Google Scholar] [CrossRef] [Green Version]
- Foroutan, A.; Fitzsimmons, C.; Mandal, R.; Berjanskii, M.V.; Wishart, D.S. Serum Metabolite Biomarkers for Predicting Residual Feed Intake (RFI) of Young Angus Bulls. Metabolites 2020, 10, 491. [Google Scholar] [CrossRef]
- Jorge-Smeding, E.; Renand, G.; Centeno, D.; Pétéra, M.; Durand, S.; Polakof, S.; Cantalapiedra-Hijar, G. Metabolomics Reveals Changes in Urea Cycle Associated to Residual Feed Intake in Growing Heifers. In Proceedings of the Energy and Protein Metabolism and Nutrition, Belo Horizonte, Brazil, 9–12 September 2019; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019; pp. 231–232. [Google Scholar]
- Clemmons, B.A.; Martino, C.; Powers, J.B.; Campagna, S.R.; Voy, B.H.; Donohoe, D.R.; Gaffney, J.; Embree, M.M.; Myer, P.R. Rumen Bacteria and Serum Metabolites Predictive of Feed Efficiency Phenotypes in Beef Cattle. Sci. Rep. 2019, 9, 19265. [Google Scholar] [CrossRef] [Green Version]
- Tardivel, P.J.C.; Canlet, C.; Lefort, G.; Tremblay-Franco, M.; Debrauwer, L.; Concordet, D.; Servien, R. ASICS: An Automatic Method for Identification and Quantification of Metabolites in Complex 1D 1H NMR Spectra. Metabolomics 2017, 13, 109. [Google Scholar] [CrossRef] [Green Version]
- Lobley, G.E. Protein Turnover—What Does It Mean for Animal Production? Can. J. Anim. Sci. 2003, 83, 327–340. [Google Scholar] [CrossRef]
- Maeda, Y.; Yamanouchi, N.; Okamoto, S.; Hashiguchi, T. Genetic Parameters of Muscle Protein Turnover Rate in the Japanese Quail, Coturnix Coturnix Japonica. Nihon Chikusan Gakkaiho 1989, 60, 484–490. [Google Scholar] [CrossRef]
- McCarthy, I.D.; Houlihan, D.F.; Carter, C.G. Individual Variation in Protein Turnover and Growth Efficiency in Rainbow Trout, Oncorhynchus Mykiss (Walbaum). Proc. R. Soc. Lond. B 1994, 257, 141–147. [Google Scholar] [CrossRef]
- Wessels, R.H.; Titgemeyer, E.C.; St Jean, G. Effect of Amino Acid Supplementation on Whole-Body Protein Turnover in Holstein Steers. J. Anim. Sci. 1997, 75, 3066. [Google Scholar] [CrossRef] [Green Version]
- INRA. Alimentation Des Ruminants; Éditions Quæ: Versailles, France, 2018. [Google Scholar]
- Hamm, L.L. Renal Handling of Citrate. Kidney Int. 1990, 38, 728–735. [Google Scholar] [CrossRef] [Green Version]
- Krebs, H.A.; Johnson, W.A. The Role of Citric Acid in Intermediate Metabolism in Animal Tissues. FEBS Lett. 1980, 117, 9. [Google Scholar] [CrossRef] [Green Version]
- Watson, J.A.; Lowenstein, J.M. Citrate and the Conversion of Carbohydrate into Fat. J. Biol. Chem. 1970, 245, 5993–6002. [Google Scholar] [CrossRef]
- MacDonald, M.J.; Smith, A.D.; Hasan, N.M.; Sabat, G.; Fahien, L.A. Feasibility of Pathways for Transfer of Acyl Groups from Mitochondria to the Cytosol to Form Short Chain Acyl-CoAs in the Pancreatic Beta Cell. J. Biol. Chem. 2007, 282, 30596–30606. [Google Scholar] [CrossRef] [Green Version]
- Kelly, A.K.; McGee, M.; Crews, D.H.; Sweeney, T.; Boland, T.M.; Kenny, D.A. Repeatability of Feed Efficiency, Carcass Ultrasound, Feeding Behavior, and Blood Metabolic Variables in Finishing Heifers Divergently Selected for Residual Feed Intake1. J. Anim. Sci. 2010, 88, 3214–3225. [Google Scholar] [CrossRef]
- Leighton, B.; Nicholas, A.R.; Pogson, C.I. The Pathway of Ketogenesis in Rumen Epithelium of the Sheep. Biochem. J. 1983, 216, 769–772. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, M.J.; Fahien, L.A.; Brown, L.J.; Hasan, N.M.; Buss, J.D.; Kendrick, M.A. Perspective: Emerging Evidence for Signaling Roles of Mitochondrial Anaplerotic Products in Insulin Secretion. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E1–E15. [Google Scholar] [CrossRef] [Green Version]
- Mosaoa, R.; Kasprzyk-Pawelec, A.; Fernandez, H.R.; Avantaggiati, M.L. The Mitochondrial Citrate Carrier SLC25A1/CIC and the Fundamental Role of Citrate in Cancer, Inflammation and Beyond. Biomolecules 2021, 11, 141. [Google Scholar] [CrossRef]
- Mycielska, M.E.; Djamgoz, M.B.A. Citrate Transport in the Human Prostate Epithelial PNT2-C2 Cell Line: Electrophysiological Analyses: Electrophysiology of Prostatic Citrate. J. Physiol. 2004, 559, 821–833. [Google Scholar] [CrossRef] [Green Version]
- Sharifabadi, H.R.; Zamiri, M.J.; Rowghani, E.; Bottje, W.G. Relationship between the Activity of Mitochondrial Respiratory Chain Complexes and Feed Efficiency in Fat-Tailed Ghezel Lambs. J. Anim. Sci. 2012, 90, 1807–1815. [Google Scholar] [CrossRef]
- Grubbs, J.K.; Fritchen, A.N.; Huff-Lonergan, E.; Dekkers, J.C.M.; Gabler, N.K.; Lonergan, S.M. Divergent Genetic Selection for Residual Feed Intake Impacts Mitochondria Reactive Oxygen Species Production in Pigs. J. Anim. Sci. 2013, 91, 2133–2140. [Google Scholar] [CrossRef] [Green Version]
- Bottje, W.G.; Carstens, G.E. Association of Mitochondrial Function and Feed Efficiency in Poultry and Livestock Species. J. Anim. Sci. 2009, 87, E48–E63. [Google Scholar] [CrossRef] [Green Version]
- Kolath, W.H.; Kerley, M.S.; Golden, J.W.; Keisler, D.H. The Relationship between Mitochondrial Function and Residual Feed Intake in Angus Steers. J. Anim. Sci. 2006, 84, 861–865. [Google Scholar] [CrossRef] [Green Version]
- Yanibada, B.; Hohenester, U.; Pétéra, M.; Canlet, C.; Durand, S.; Jourdan, F.; Boccard, J.; Martin, C.; Eugène, M.; Morgavi, D.P.; et al. Inhibition of Enteric Methanogenesis in Dairy Cows Induces Changes in Plasma Metabolome Highlighting Metabolic Shifts and Potential Markers of Emission. Sci. Rep. 2020, 10, 15591. [Google Scholar] [CrossRef]
- Ungerfeld, E.M. Metabolic Hydrogen Flows in Rumen Fermentation: Principles and Possibilities of Interventions. Front. Microbiol. 2020, 11, 589. [Google Scholar] [CrossRef] [Green Version]
- Clemmons, B.A.; Powers, J.B.; Campagna, S.R.; Seay, T.B.; Embree, M.M.; Myer, P.R. Rumen Fluid Metabolomics of Beef Steers Differing in Feed Efficiency. Metabolomics 2020, 16, 23. [Google Scholar] [CrossRef]
- Artegoitia, V.M.; Foote, A.P.; Lewis, R.M.; Freetly, H.C. Rumen Fluid Metabolomics Analysis Associated with Feed Efficiency on Crossbred Steers. Sci. Rep. 2017, 7, 2864. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.; Munro, J.C.; Zhou, M.; Guan, L.L.; Schenkel, F.S.; Steele, M.A.; Miller, S.P.; Montanholi, Y.R. Associations of Rumen Parameters with Feed Efficiency and Sampling Routine in Beef Cattle. Animal 2018, 12, 1442–1450. [Google Scholar] [CrossRef] [PubMed]
- Giráldez, F.J.; Santos, N.; Santos, A.; Valdés, C.; López, S.; Andrés, S. Fattening Lambs with Divergent Residual Feed Intakes and Weight Gains: Unravelling Mechanisms Driving Feed Efficiency. Anim. Feed. Sci. Technol. 2021, 273, 114821. [Google Scholar] [CrossRef]
- Myer, P.R.; Freetly, H.C.; Wells, J.E.; Smith, T.P.L.; Kuehn, L.A. Analysis of the Gut Bacterial Communities in Beef Cattle and Their Association with Feed Intake, Growth, and Efficiency. J. Anim. Sci. 2017, 95, 3215. [Google Scholar] [CrossRef]
- Spillane, C.; Waters, S.M. Rumen Microbiome Composition Is Altered in Sheep Divergent in Feed Efficiency. Front. Microbiol. 2020, 11, 16. [Google Scholar]
- Lopes, D.R.G.; de Souza Duarte, M.; La Reau, A.J.; Chaves, I.Z.; de Oliveira Mendes, T.A.; Detmann, E.; Bento, C.B.P.; Mercadante, M.E.Z.; Bonilha, S.F.M.; Suen, G.; et al. Assessing the Relationship between the Rumen Microbiota and Feed Efficiency in Nellore Steers. J. Anim. Sci. Biotechnol. 2021, 12, 79. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists. Animal Feed Official Methods Analysis, 16th ed.; AOAC: Gaithersburg, MA, USA, 1998. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Lefort, G.; Liaubet, L.; Canlet, C.; Tardivel, P.; Père, M.-C.; Quesnel, H.; Paris, A.; Iannuccelli, N.; Vialaneix, N.; Servien, R. ASICS: An R Package for a Whole Analysis Workflow of 1D 1H NMR Spectra. Bioinformatics 2019, 35, 4356–4363. [Google Scholar] [CrossRef]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.-A. MixOmics: An R Package for ‘omics Feature Selection and Multiple Data Integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [Green Version]
- Hervé, M.R.; Nicolè, F.; Lê Cao, K.-A. Multivariate Analysis of Multiple Datasets: A Practical Guide for Chemical Ecology. J. Chem. Ecol. 2018, 44, 215–234. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | Phase CONC (N = 277) | |||||||
| Traits * | Mean (SD) | rfi− | rfi+ | p-Value | ||||
| ADFIC 1 (g/day) | 2098 (273) | 1991 | 2173 | 5 × 10−12 | ||||
| E-MWC 2 (kg) | 20.6 (1.9) | 20.3 | 20.7 | 0.0014 | ||||
| ADGC 3 (g/day) | 327.6 (62.4) | 326.5 | 326.5 | 0.93 | ||||
| BFTC 4 (mm) | 5.73 (0.87) | 5.81 | 5.74 | 0.41 | ||||
| MDC 5 (mm) | 28.1 (2.4) | 27.8 | 28.3 | 0.02 | ||||
| RFIC 6 (g/day) | 0 (151.4) | −71.4 | 66.6 | <2.2 × 10−16 | ||||
| B | Phase MIX (N = 167) | |||||||
| Traits * | Mean (SD) | rfi− | rfi+ | rfi− | rfi+ | p-Value | ||
| Summer | Fall | Summer | Fall | |||||
| ADFIM 1 (g/day) | 1857 (280) | 1804 | 1856 | 1817 | 1869 | 1830 | 1843 | 0.80 |
| E-MWM 2 (kg) | 22.8 (1.5) | 22.7 | 22.6 | 22.9 | 22.8 | 22.7 | 22.8 | 0.38 |
| ADGM 3 (g/day) | 124.4 (64.3) | 147 | 113 | 137 | 104 | 130 | 121 | 0.40 |
| BFTM 4 (mm) | 4.54 (0.78) | 4.79 | 4.55 | 4.54 | 4.30 | 4.67 | 4.42 | 0.02 |
| MDM 5 (mm) | 27.1 (2.4) | 26.8 | 18.5 | 27.1 | 18.9 | 22.6 | 23.0 | 0.20 |
| RFIM 6(g/day) | 0 (183.1) | −3.2 | −3.8 | 4.1 | 3.5 | −3.5 | 3.8 | 0.81 |
| Phase MIX | (N = 109 #) | |||||||
| ADForIM 7 (g/day) | 1279 (224) | 1276 | 1290 | 1268 | 1282 | 1283 | 1275 | 0.75 |
| RForIM 8(g/day) | 0 (175.7) | −3.7 | −3.0 | 3.3 | 4.0 | −3.3 | 3.6 | 0.83 |
| Phenotypic RFIC (g/Day) 1 | |||||
|---|---|---|---|---|---|
| Genetic Line | ≤−75 | ]−75,0] | ]0,75] | >75 | |
| rfi− | 62 | 38 | 17 | 18 | |
| rfi+ | 20 | 19 | 40 | 63 | |
| Phenotypic RFIM 1 (g/Day) | |||||
|---|---|---|---|---|---|
| Genetic Line | ≤−90 | ]−90,0] | ]0,90] | >90 | |
| rfi− | 28 | 17 | 17 | 21 | |
| rfi+ | 25 | 18 | 13 | 28 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Touitou, F.; Tortereau, F.; Bret, L.; Marty-Gasset, N.; Marcon, D.; Meynadier, A. Evaluation of the Links between Lamb Feed Efficiency and Rumen and Plasma Metabolomic Data. Metabolites 2022, 12, 304. https://doi.org/10.3390/metabo12040304
Touitou F, Tortereau F, Bret L, Marty-Gasset N, Marcon D, Meynadier A. Evaluation of the Links between Lamb Feed Efficiency and Rumen and Plasma Metabolomic Data. Metabolites. 2022; 12(4):304. https://doi.org/10.3390/metabo12040304
Chicago/Turabian StyleTouitou, Florian, Flavie Tortereau, Lydie Bret, Nathalie Marty-Gasset, Didier Marcon, and Annabelle Meynadier. 2022. "Evaluation of the Links between Lamb Feed Efficiency and Rumen and Plasma Metabolomic Data" Metabolites 12, no. 4: 304. https://doi.org/10.3390/metabo12040304