


Maternal Nutrition Affects Nitrogen Isotopic Signature in Blood Plasma of Beef Cattle Dams and Their Offspring
 , , ,
, , , 
Abstract
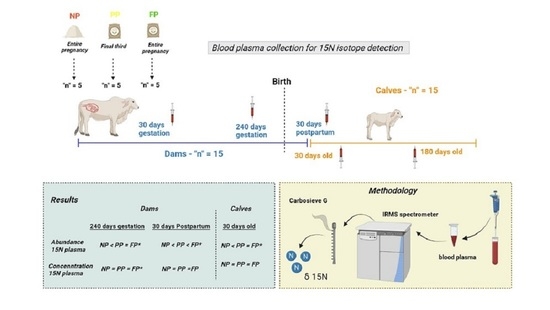
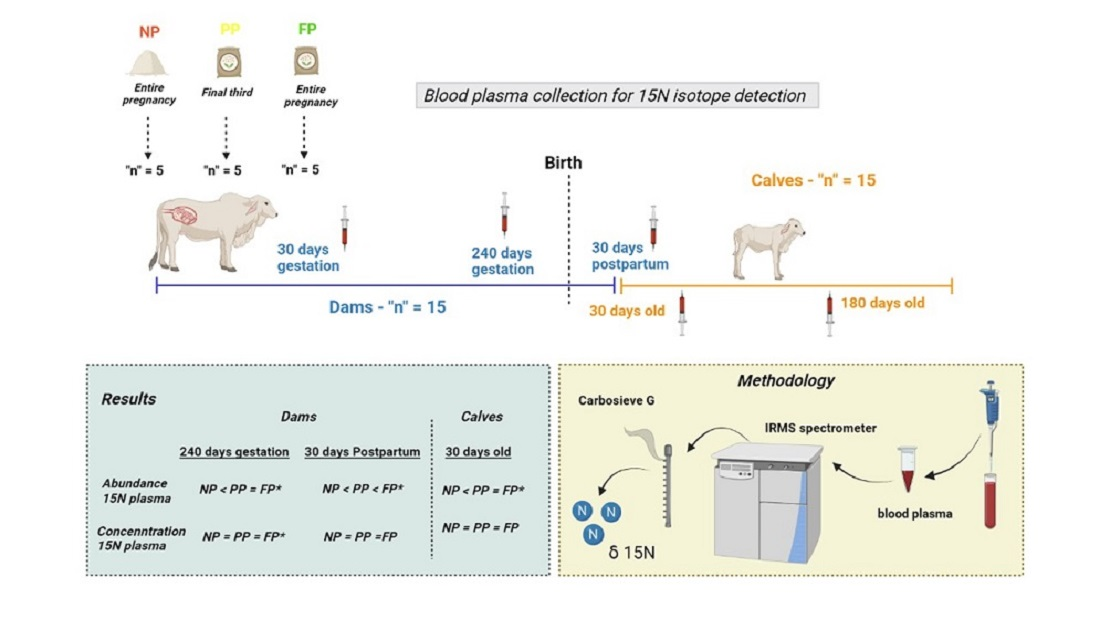
:
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Management
2.2. Bromatological Analysis of Supplements and Pastures
2.3. Processing of Forage and Supplements for the Isotopic Analysis
2.4. Blood Plasma Collection and Processing
2.5. Isotopic Analysis of Delta 15 N (15 N/14 N)
2.6. Statistical Analysis
3. Results
3.1. Isotopic Evaluation of Forages and Supplements
3.2. Isotopic Evaluation of Blood Plasma of Cows
3.3. Isotopic Evaluation of Blood Plasma from Calves
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Euclides Filho, K.; Euclides, V.; Pires, A. Desenvolvimento Recente Da Pecuária de Corte Brasileira e Suas Perspectivas. In Bovinocultura de Corte; FEALQ: Piracicaba, Brazil, 2010; Volume 1, pp. 11–40. [Google Scholar]
- Robinson, J.J.; McDonald, I.; Fraser, C.; Crofts, R.M.J. Studies on Reproduction in Prolific Ewes, I. Growth of the Products of Conception. J. Agric. Sci. 1977, 88, 539–552. [Google Scholar] [CrossRef]
- Du, M.; Tong, J.; Zhao, J.; Underwood, K.R.; Zhu, M.; Ford, S.P.; Nathanielsz, P.W. Fetal Programming of Skeletal Muscle Development in Ruminant Animals. J. Anim. Sci. 2010, 88, E51–E61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Bazer, F.W.; Wallace, J.M.; Spencer, T.E. Board-Invited Review: Intrauterine Growth Retardation: Implications for the Animal Sciences. J. Anim. Sci. 2006, 84, 2316–2337. [Google Scholar] [CrossRef] [PubMed]
- Duarte Júnior, M.; Zervoudakis, K.; Zervoudakis, J.; Tsuneda, P.; Silva, J.; Silva, R.; Koscheck, J.; Fioravanti Filho, R.; Almeida, R. Suplementação de Fêmeas Bovinas Em Pastejo: Aspectos Nutricionais e Reprodutivos. PubVet 2016, 9, 321–336. [Google Scholar] [CrossRef] [Green Version]
- Barker, D.J.P. The Fetal and Infant Origins of Adult Disease. Br. Med. J. 1990, 301, 1111. [Google Scholar] [CrossRef] [Green Version]
- Funston, R.N.; Summers, A.F. Effect of Prenatal Programming on Heifer Development. Vet. Clin. N. Am. Food Anim. Pract. 2013, 29, 517–536. [Google Scholar] [CrossRef]
- Wilkinson, D.J. Historical and Contemporary Stable Isotope Tracer Approaches to Studying Mammalian Protein Metabolism. Mass Spectrom. Rev. 2018, 37, 57–80. [Google Scholar] [CrossRef]
- Robbins, C.T.; Felicetti, L.A.; Sponheimer, M. The Effect of Dietary Protein Quality on Nitrogen Isotope Discrimination in Mammals and Birds. Oecologia 2005, 144, 534–540. [Google Scholar] [CrossRef]
- Sutoh, M.; Obara, Y.; Yoneyamat, T. The Effects of Feed- Ing Regimen and Dietary Sucrose Supplementation on Natural Abundance of 15N in Some Components of Ruminal Fluid and Plasma of Sheep. J. Anim. Sci. 1992, 71, 226–231. [Google Scholar] [CrossRef] [Green Version]
- Wattiaux, M.A.; Reed, J.D. Fractionation of Nitrogen Isotopes by Mixed Ruminal Bacterial. J. Anim. Sci. 1995, 73, 257–266. [Google Scholar] [CrossRef]
- Cheng, L.; Sheahan, A.J.; Gibbs, S.J. Technical Note: Nitrogen Isotopic Fractionation Can Be Used to Predict Nitrogen-Use Efficiency in Dairy Cows Fed Temperate Pasture. J. Anim. Sci. 2013, 91, 5785–5788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheadon, N.M.; Mcgee, M.; Edwards, G.R.; Dewhurst, R.J. Plasma Nitrogen Isotopic Fractionation and Feed Efficiency in Growing Beef Heifers. Br. J. Nutr. 2014, 111, 1705–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahar, B.; Monahan, F.J.; Moloney, A.P.; O’Kiely, P.; Scrimgeour, C.M.; Schmidt, O. Alteration of the Carbon and Nitrogen Stable Isotope Composition of Beef by Substitution of Grass Silage with Maize Silage. Rapid Commun. Mass Spectrom. 2005, 19, 1937–1942. [Google Scholar] [CrossRef] [PubMed]
- Tieszen, L.L. Natural Variations in the Carbon Isotope Values of Plants: Implications for Archaeology, Ecology, and Paleoecology. J. Archaeol. Sci. 1991, 18, 227–248. [Google Scholar] [CrossRef]
- Vieira Júnior, L.C.; Jorge, A.M.; Factori, M.A.; Martins, M.B.; Ribeiro, F.A.; Queiroz, E.O. Exemplos Práticos Do Processo de Autenticação de Carnes e Seus Derivados. PubVet 2012, 6, 1387–1392. [Google Scholar] [CrossRef] [Green Version]
- Schalch Junior, F.J.; Polizel, G.H.G.; Cançado, F.A.C.Q.; Fernandes, A.C.; Mortari, I.; Pires, P.R.L.; Fukumasu, H.; Santana, M.H.d.A.; Saran Netto, A. Prenatal Supplementation in Beef Cattle and Its Effects on Plasma Metabolome of Dams and Calves. Metabolites 2022, 12, 347. [Google Scholar] [CrossRef]
- Silva, D.; Queiroz, A. Análise de Alimentos: Métodos Químicos e Biológicos, 3rd ed.; UFV: Viçosa, Brazil, 2009. [Google Scholar]
- Cracco, R.C.; de Oliveira Bussiman, F.; Polizel, G.H.G.; Furlan, É.; Garcia, N.P.; Poit, D.A.S.; Pugliesi, G.; de Santana, M.H.A. Effects of Maternal Nutrition on Female Offspring Weight Gain and Sexual Development. Front. Genet. 2021, 12, 2059. [Google Scholar] [CrossRef]
- Van Soest, P. Nutritional Ecology of the Ruminant, 4th ed.; Cornell University Press: Ithaca, NY, USA, 1995. [Google Scholar]
- Sindirações Métodos Analíticos—Minerais Por Espectrometria de Emissão Atômica Por Plasma Indutivamente Acoplado (ICP-OES). In Compêndio Brasileiro de Alimentação Animal; Sindirações: São Paulo, Brazil, 2013; pp. 175–180.
- Milagres, J.J.d.M. Emissões de N2 e N2O Provenientes Do Fertilizante Aplicado a Solos Cultivados Com Cana-de-Açúcar Pelo Método Do Traçador 15N; CENA/USP: Piracicaba, SP, Brazil, 2014. [Google Scholar]
- Cantalapiedra-Hijar, G.; Fouillet, H.; Chantelauze, C.; Khodorova, N.; Bahloul, L.; Ortigues-Marty, I. The Isotopic Nitrogen Turnover Rate as a Proxy to Evaluate in the Long-Term the Protein Turnover in Growing Ruminants. J. Agric. Sci. 2020, 157, 701–710. [Google Scholar] [CrossRef]
- Jenkins, S.G.; Partridge, S.T.; Stephenson, T.R.; Farley, S.D.; Robbins, C.T. Nitrogen and Carbon Isotope Fractionation between Mothers, Neonates, and Nursing Offspring. Oecologia 2001, 129, 336–341. [Google Scholar] [CrossRef]
- Aurioles, D.; Koch, P.L.; Le Boeuf, B.J. Differences in Foraging Location of Mexican and California Elephant Seals: Evidence from Stable Isotopes in Pups. Mar. Mammal Sci. 2006, 22, 326–338. [Google Scholar] [CrossRef]
- Aurioles-Gamboa, D.; Newsome, S.D.; Salazar-Pico, S.; Koch, P.L. Stable Isotope Differences between Sea Lions (Zalophus) from the Gulf of California and Gaĺpagos Islands. J. Mammal. 2009, 90, 1410–1420. [Google Scholar] [CrossRef]
- Newsome, S.D.; Koch, P.L.; Etnier, M.A.; Aurioles-Gamboa, D. Using Carbon and Nitrogen Iso-Tope Values to Investigate Maternal Strategies in Northeast Pacific Otariids. Mar. Mammal Sci. 2006, 22, 556–572. [Google Scholar] [CrossRef]
- Elorriaga-Verplancken, F.; Aurioles-Gamboa, D.; Newsome, S.D.; Martínez-Díaz, S.F. Δ15N and Δ13C Values in Dental Collagen as a Proxy for Age- and Sex-Related Variation in Foraging Strategies of California Sea Lions. Mar. Biol. 2013, 160, 641–652. [Google Scholar] [CrossRef]
- Gannes, L.Z.; Del Rio, C.M.; Koch, P. Natural Abundance Variations in Stable Isotopes and Their Potential Uses in Animal Physiological Ecology. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 1998, 119, 725–737. [Google Scholar] [CrossRef]
- Hoffman, M.L.; Reed, S.A.; Pillai, S.M.; Jones, A.K.; McFadden, K.K.; Zinn, S.A.; Govoni, K.E. Physiology and Endocrinology Symposium: The Effects of Poor Maternal Nutrition during Gestation on Offspring Postnatal Growth and Metabolism. J. Anim. Sci. 2017, 95, 2222–2232. [Google Scholar] [CrossRef] [Green Version]
- Zago, D.; Canozzi, M.E.A.; Barcellos, J.O.J. Pregnant Beef Cow’s Nutrition and Its Effects on Postnatal Weight and Carcass Quality of Their Progeny. PLoS ONE 2020, 15, e0237941. [Google Scholar] [CrossRef]
- Rodrigues, L.M.; Schoonmaker, J.P.; Resende, F.D.; Siqueira, G.R.; Neto, O.R.M.; Gionbelli, M.P.; Gionbelli, T.R.S.; Ladeira, M.M.; Rodrigues, L.M.; Schoonmaker, J.P.; et al. Effects of Protein Supplementation on Nellore Cows’ Reproductive Performance, Growth, Myogenesis, Lipogenesis and Intestine Development of the Progeny. Anim. Prod. Sci. 2020, 61, 371–380. [Google Scholar] [CrossRef]
- Cantalapiedra-Hijar, G.; Ortigues-Marty, I.; Sepchat, B.; Agabriel, J.; Huneau, J.F.; Fouillet, H. Diet-Animal Fractionation of Nitrogen Stable Isotopes Reflects the Efficiency of Nitrogen Assimilation in Ruminants. Br. J. Nutr. 2015, 113, 1158–1169. [Google Scholar] [CrossRef] [Green Version]
- Tudor, G.D.; O’Rourke, P.K. The Effect of Pre- and Post-Natal Nutrition on the Growth of Beef Cattle. 2. The Effect of Severe Restriction in Early Post-Natal Life on Growth and Feed Efficiency during Recovery. Aust. J. Agric. Res. 1980, 31, 179–189. [Google Scholar] [CrossRef]
- Barboza, P.S.; Parker, K.L. Body Protein Stores and Isotopic Indicators of N Balance in Female Reindeer (Rangifer Tarandus) during Winter. Physiol. Biochem. Zool. 2006, 79, 628–644. [Google Scholar] [CrossRef]

{kind=link}
| Ingredients | Mineral Supplement | Energetic–Proteic Supplement |
|---|---|---|
| Ground corn (%) Soybean meal (%) Dicalcium phosphate (%) Urea 45% (%) Salt (%) Minerthal 160 MD (%) * Total digestible nutrients (%) Crude protein (%) Non-protein nitrogen (%) Acid detergent fiber (%) Neutral detergent fiber (%) Fat (%) Calcium (g/kg) Phosphate (g/kg) | 35 - 10 - 30 25 26.76 2.79 - 1.25 4.29 1.26 74.11 59.38 | 60 30 - 2.5 5 2.5 67.55 24.78 7.03 4.76 11.24 2.61 6.2 7.24 |
| Forage Availability | Diets | ||
|---|---|---|---|
| NP | PP | FP | |
| Pasture Availability (kg DM/ha) | 3476.24 ± 1594.40 | 4597.35 ± 1189.80 | 5578.03 ± 2049.37 |
| Leaf Availability (kg DM/ha) | 573.59 ± 340.56 | 569.13 ± 485.76 | 727.49 ± 643.30 |
| Thatch Availability (kg DM/ha) | 562.46 ± 396.97 | 799.05 ± 545.80 | 1347.44 ± 1038.42 |
| Dead material availability (kg DM/ha) | 2340.07 ± 1275.00 | 3229.43 ± 973.34 | 3503.07 ± 1410.43 |
| Stocking Rate (AU/ha) | 2.19 ± 1.02 | 1.74 ± 0.56 | 2.26 ± 0.99 |
| Leaf supply for animal unit (KG/DM) | 316.64 ± 216.96 | 359.93 ± 245.10 | 366.59 ± 306.58 |
| Forage Nutrientes (%) | Diets | ||
|---|---|---|---|
| NP | PP | FP | |
| Crude protein | 7.38 ± 1.72 | 7.82 ± 2.28 | 7.40 ± 2.30 |
| Total digestible nutrients | 63.07 ± 1.45 | 64.10 ± 2.33 | 61.43 ± 2.12 |
| Neutral detergent fiber | 59.03 ± 3.67 | 61.43 ± 5.05 | 58.49 ± 4.11 |
| Calcium | 0.38 ± 0.11 | 0.35 ± 0.05 | 0.39 ± 0.08 |
| Phosphor | 0.19 ± 0.03 | 0.19 ± 0.03 | 0.17 ± 0.03 |
| Traits | Period | Diets | p-Value | ||
|---|---|---|---|---|---|
| NP | PP | FP | |||
| Abundance | Initial | 2.002 ± 0.086 A | 2.107 ± 0.151 A | 2.160 ± 0.078 A | 0.341 |
| Pre | 2.412 ± 0.104 B a | 3.390 ± 0.135 B b | 3.796 ± 0.157 B b | <0.001 | |
| Post | 2.156 ± 0.073 A B a | 2.722 ± 0.087 C b | 3.094 ± 0.090 C c | <0.001 | |
| p-value | 0.033 | <0.001 | <0.001 | ||
| Concentration | Initial | 1.204 ± 0.034 A B | 1.192 ± 0.027 | 1.190 ± 0.073 | 0.986 |
| Pre | 1.120 ± 0.022 A | 1.270 ± 0.080 | 1.246 ± 0.060 | 0.064 | |
| Post | 1.254 ± 0.038 B | 1.280 ± 0.049 | 1.338 ± 0.056 | 0.207 | |
| p-value | 0.037 | 0.518 | 0.275 | ||
| Traits | Age | Diets | p-Value | ||
|---|---|---|---|---|---|
| NP | PP | FP | |||
| Abundance | 30 days | 3.022 ± 0.149 a | 3.910 ± 0.095 b | 3.954 ± 0.150 b | 0.002 |
| 180 days | 3.028 ± 0.228 | 3.385 ± 0.221 | 3.134 ± 0.223 | 0.878 | |
| p-value | 1.000 | 0.368 | 0.023 | ||
| Concentration | 30 days | 1.082 ± 0.048 | 1.105 ± 0.022 | 1.180 ± 0.042 | 0.479 |
| 180 days | 1.090 ± 0.022 | 1.137 ± 0.049 | 1.178 ± 0.025 | 0.386 | |
| p-value | 1.000 | 0.991 | 1.000 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meciano, G.d.P.; Schalch Junior, F.J.; Polizel, G.H.G.; Fernandes, A.C.; Cracco, R.C.; Saran Netto, A.; Gomes, R.d.C.; Cônsolo, N.R.B.; Santana, M.H.d.A. Maternal Nutrition Affects Nitrogen Isotopic Signature in Blood Plasma of Beef Cattle Dams and Their Offspring. Metabolites 2022, 12, 1249. https://doi.org/10.3390/metabo12121249
Meciano GdP, Schalch Junior FJ, Polizel GHG, Fernandes AC, Cracco RC, Saran Netto A, Gomes RdC, Cônsolo NRB, Santana MHdA. Maternal Nutrition Affects Nitrogen Isotopic Signature in Blood Plasma of Beef Cattle Dams and Their Offspring. Metabolites. 2022; 12(12):1249. https://doi.org/10.3390/metabo12121249
Chicago/Turabian StyleMeciano, Gabriela de Pauli, Fernando José Schalch Junior, Guilherme Henrique Gebim Polizel, Arícia Christofaro Fernandes, Roberta Cavalcante Cracco, Arlindo Saran Netto, Rodrigo da Costa Gomes, Nara Regina Brandão Cônsolo, and Miguel Henrique de Almeida Santana. 2022. "Maternal Nutrition Affects Nitrogen Isotopic Signature in Blood Plasma of Beef Cattle Dams and Their Offspring" Metabolites 12, no. 12: 1249. https://doi.org/10.3390/metabo12121249