

Butin Mitigates Memory Impairment in Streptozotocin-Induced Diabetic Rats by Inhibiting Oxidative Stress and Inflammatory Responses


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Chemicals and Reagents
2.2. Animals
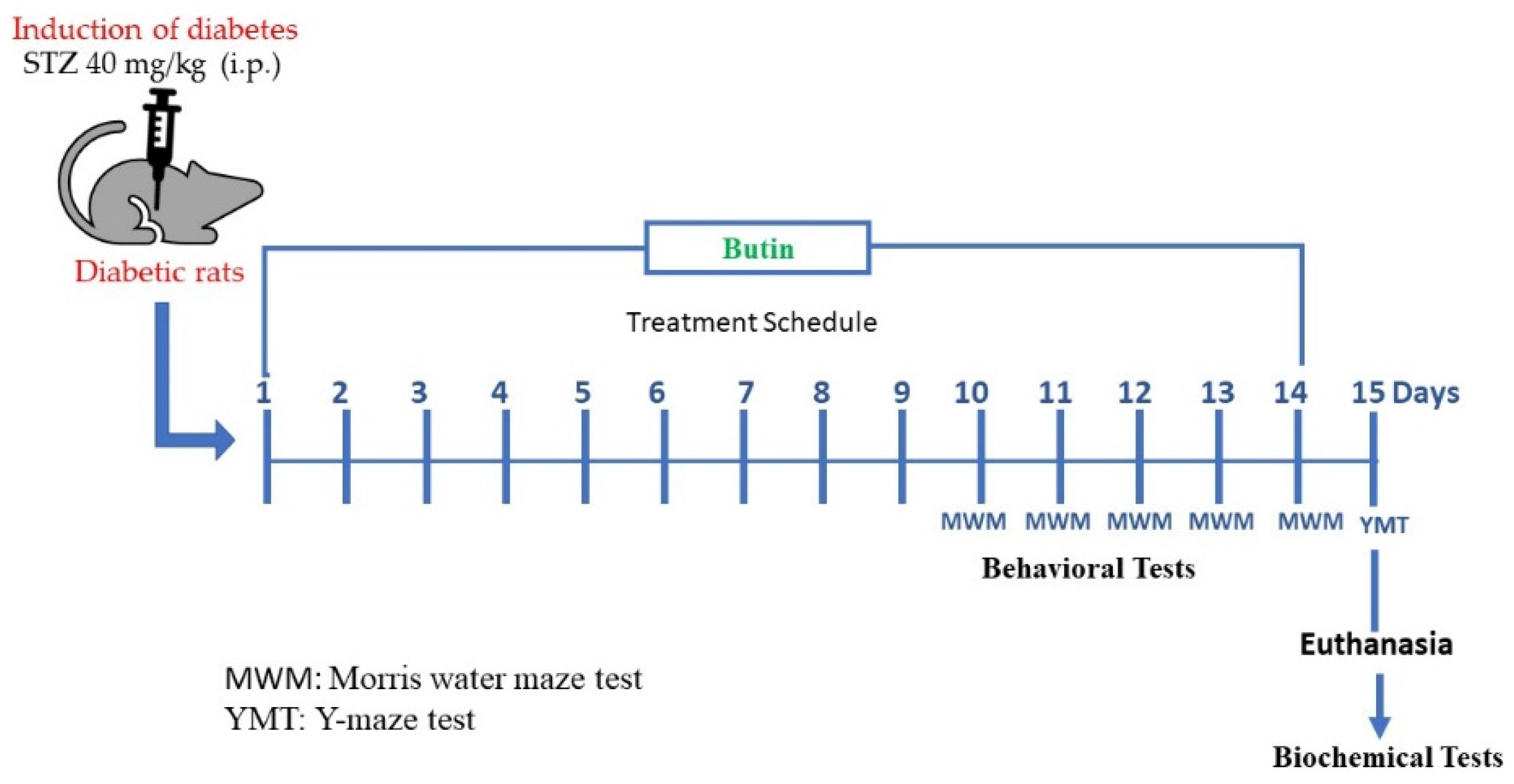
2.3. Experimental Design
2.3.1. Induction of Diabetes Mellitus
2.3.2. Evaluation of Butin against STZ-Induced Memory Deficit in Diabetic Rats
2.4. Behavioural Tests
2.4.1. Morris Water Maze (MWM) Test
2.4.2. Y-Maze Test
2.5. Biochemical Tests
2.5.1. Brain Tissue Homogenisation
2.5.2. Acetylcholinesterase (AchE) Activity
2.5.3. Endogenous Antioxidants
2.5.4. Markers of Oxidative and Nitrative Stress
2.5.5. Pro-Inflammatory Cytokines
2.5.6. Estimation of Mitochondrial Complex I, II and III
2.6. Statistical Analysis
3. Results
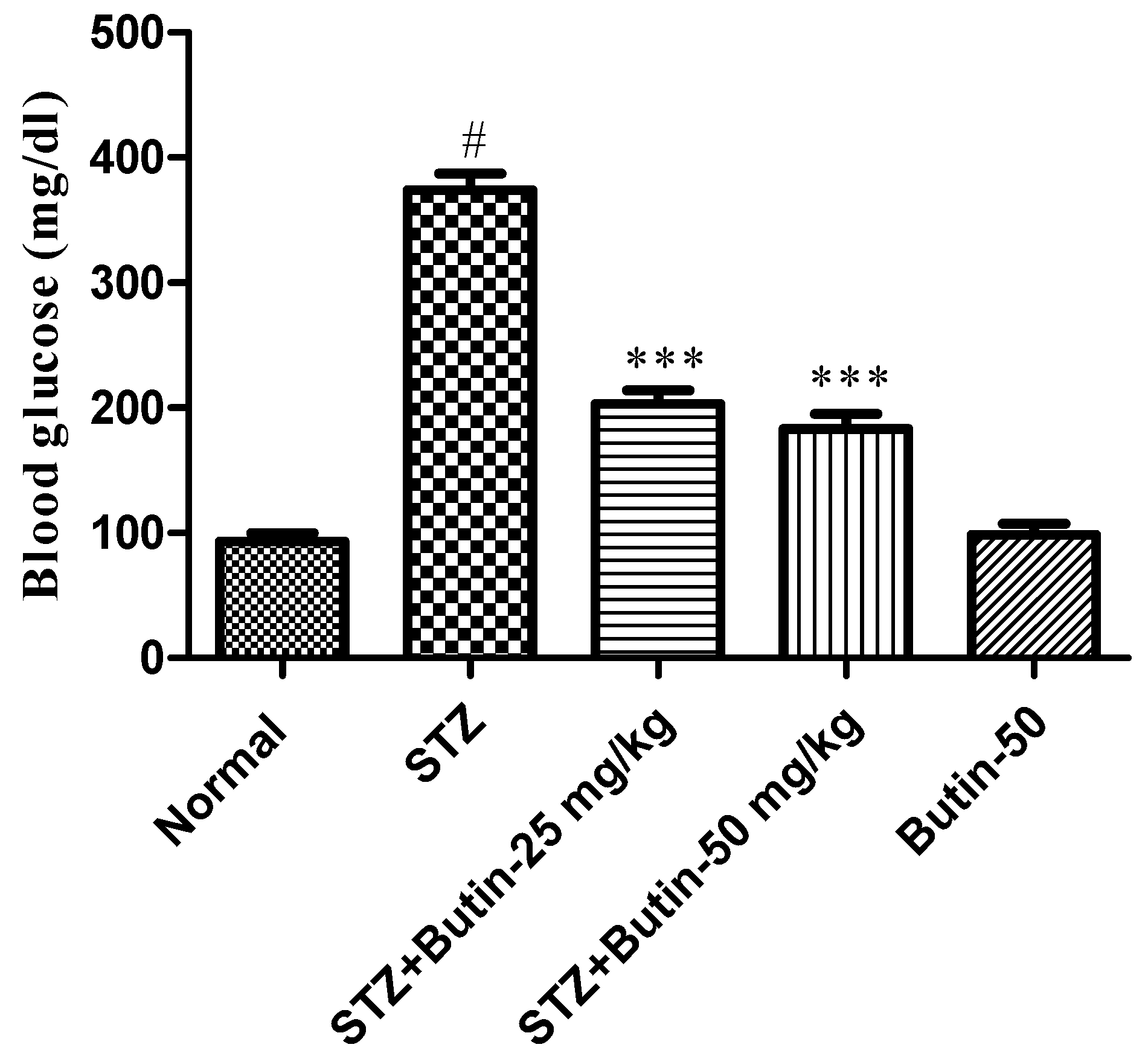
3.1. Blood Glucose Levels
3.2. Behavioural Parameters
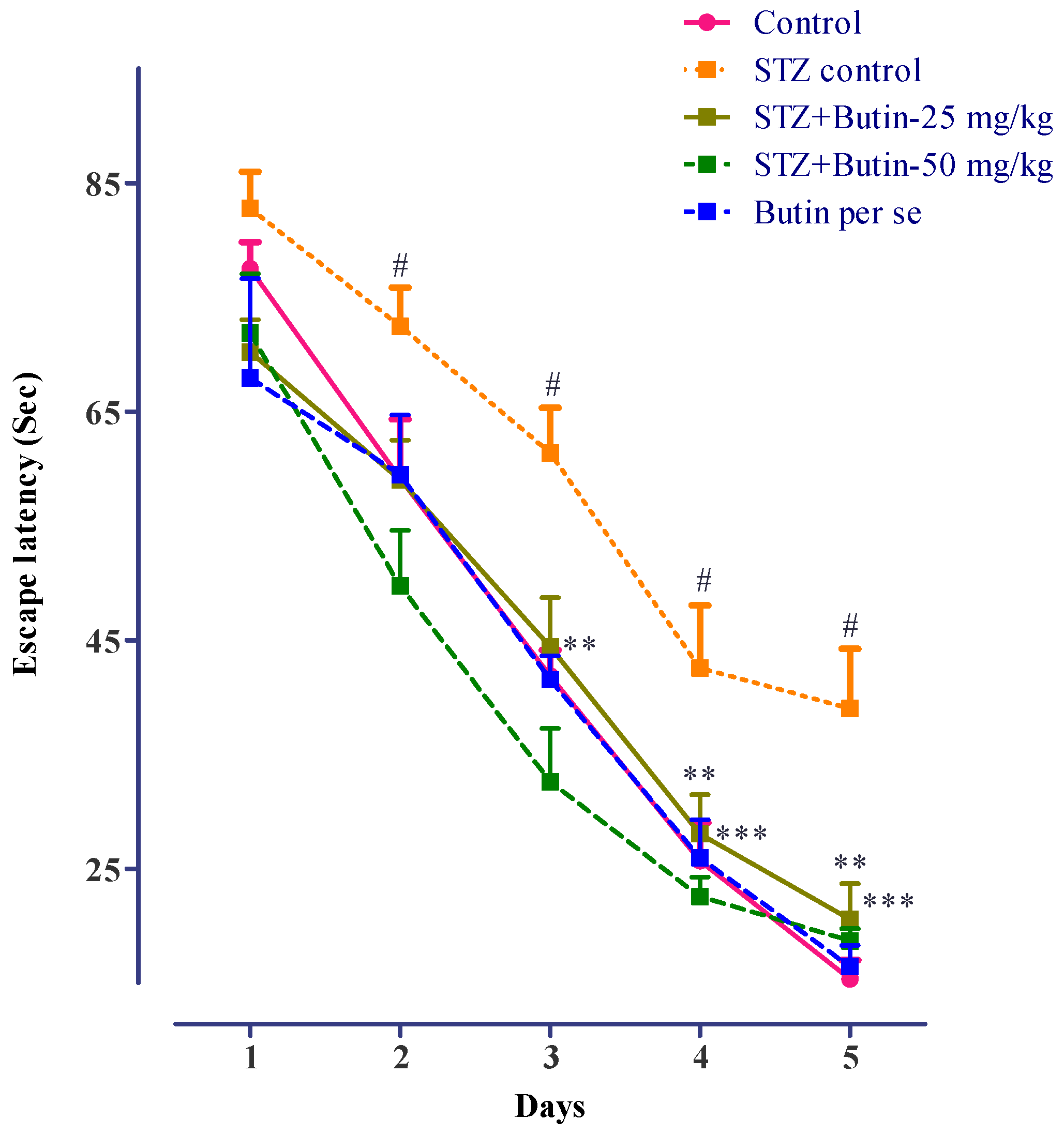
3.2.1. MWM
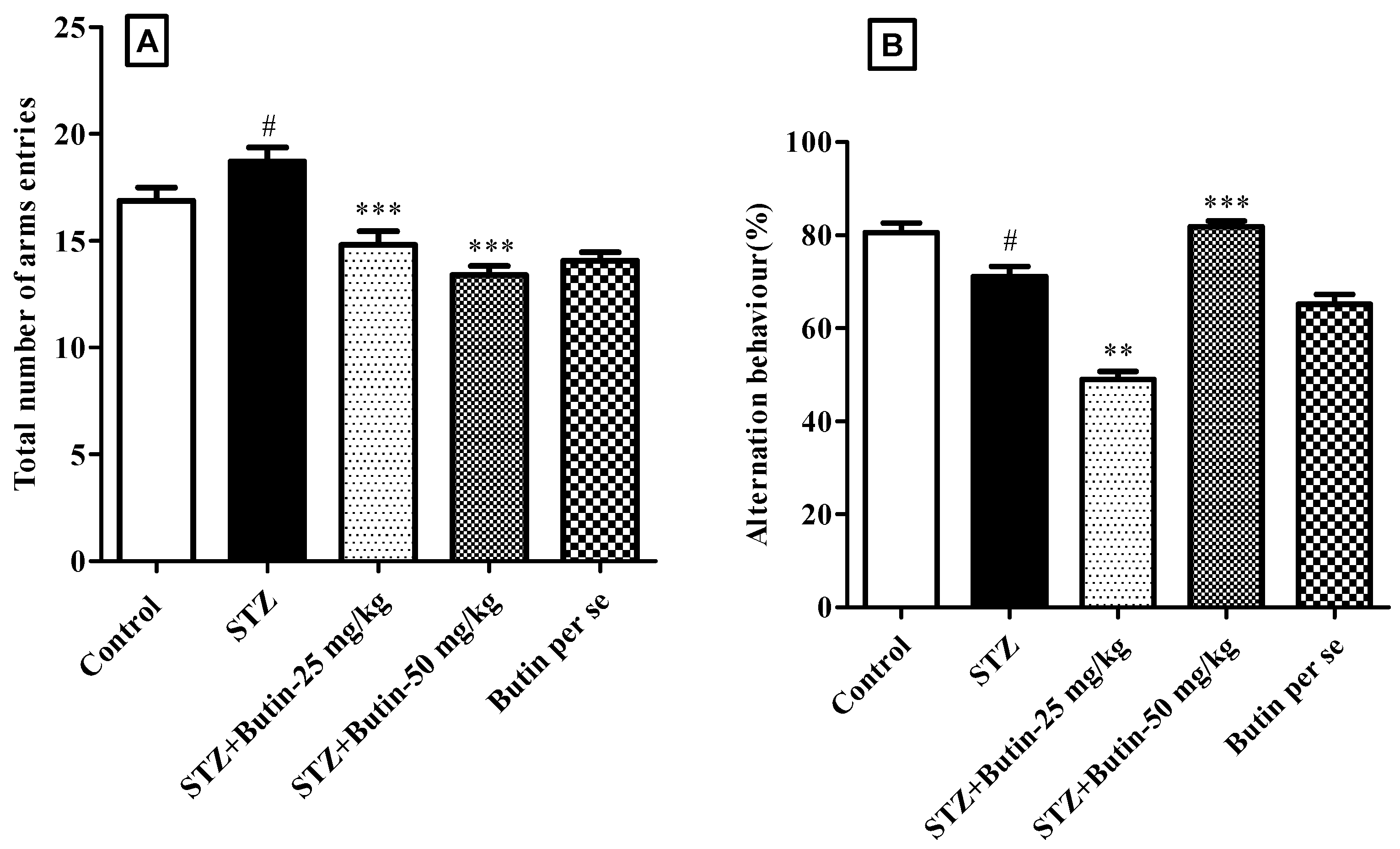
3.2.2. Y-Maze Test
3.3. Biochemical Parameters
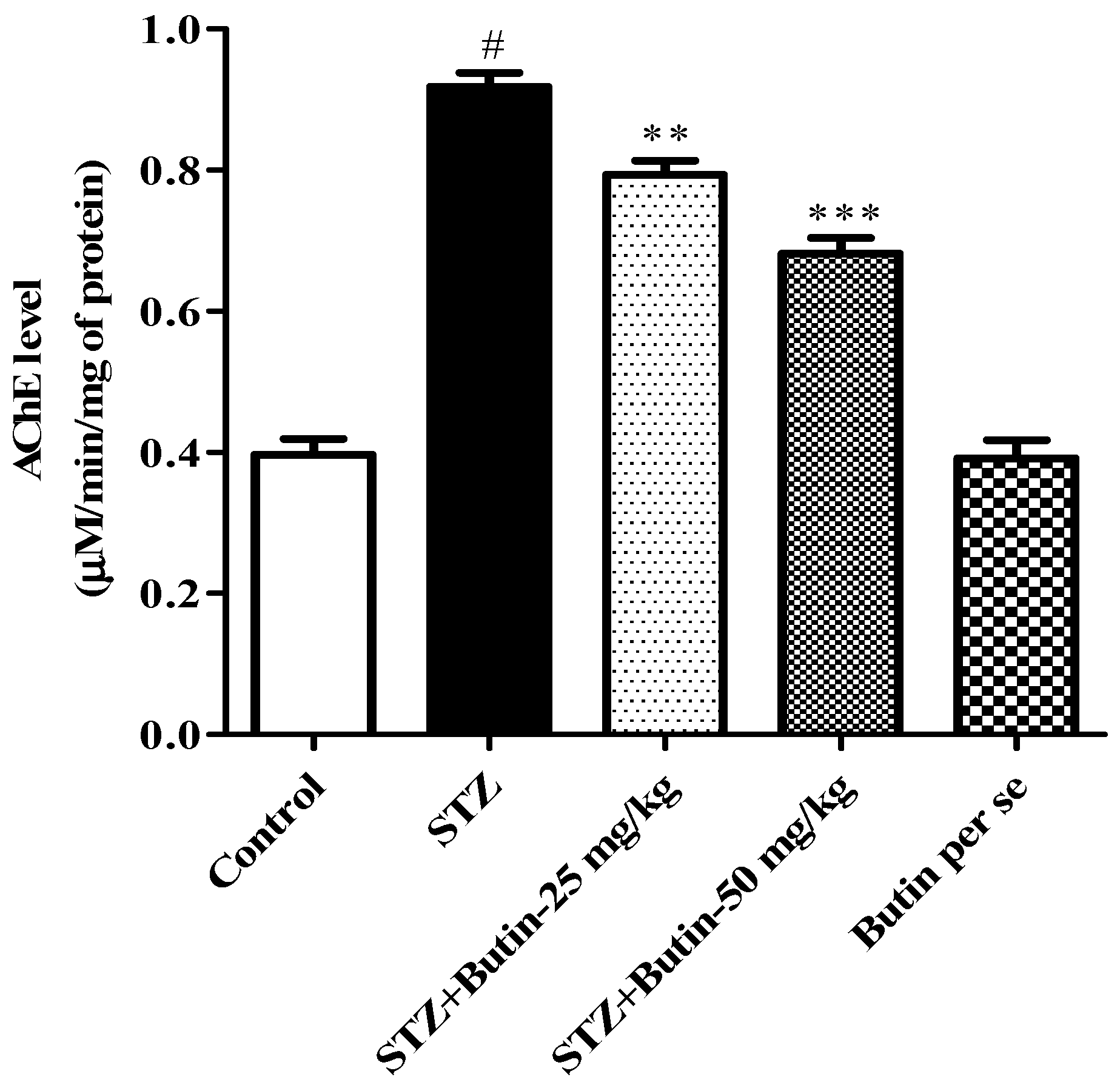
3.3.1. AchE Activity
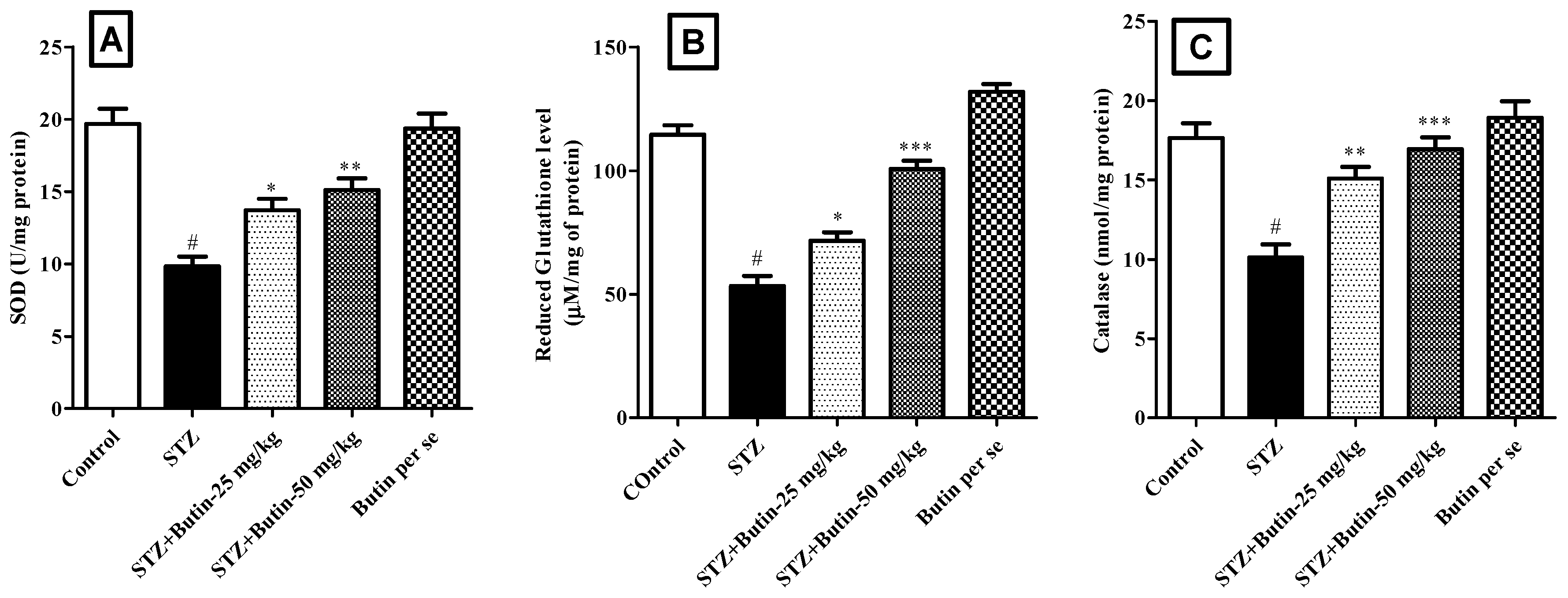
3.3.2. Endogenous Antioxidants
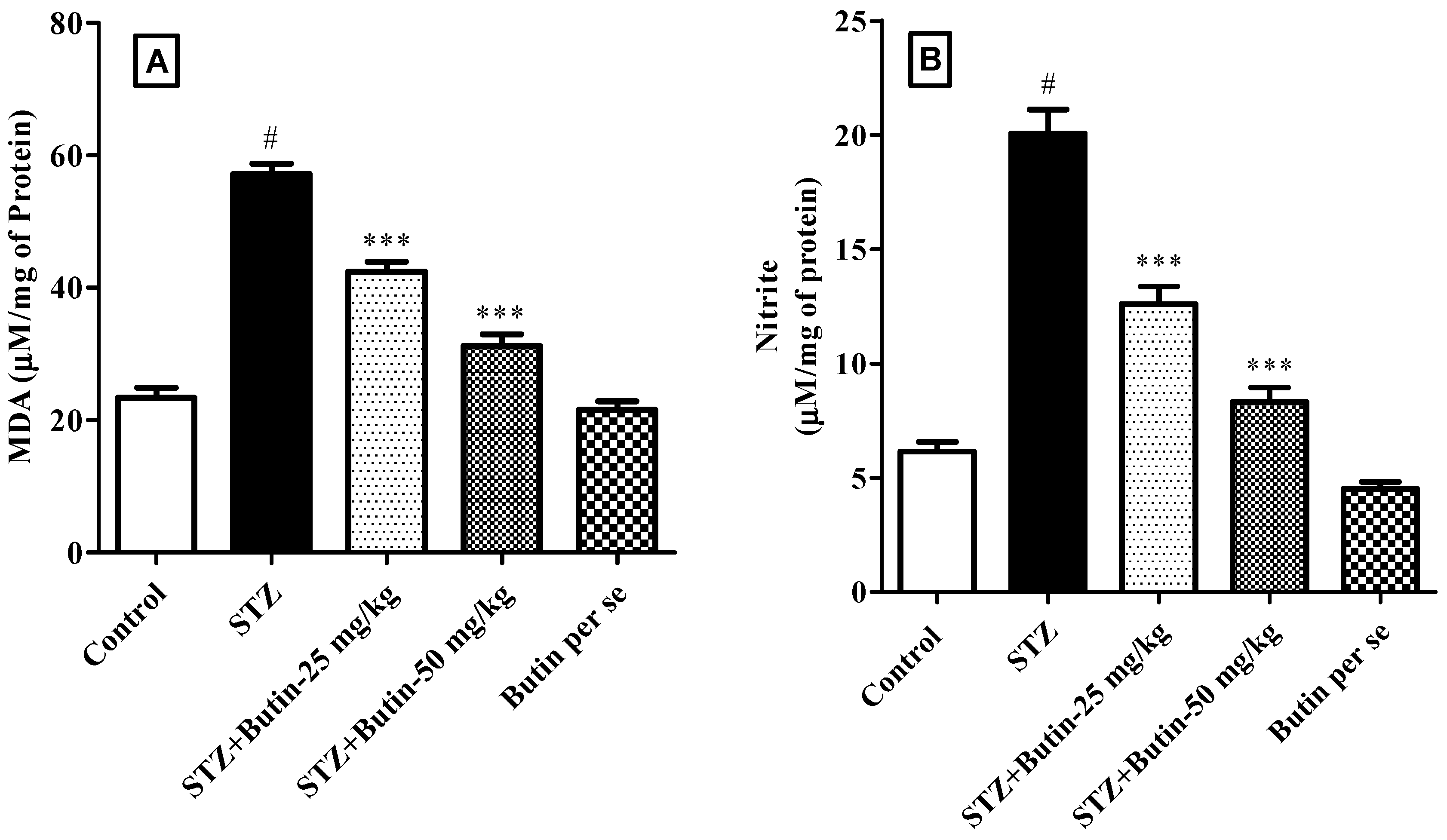
3.3.3. Markers of Oxidative and Nitrative Stress
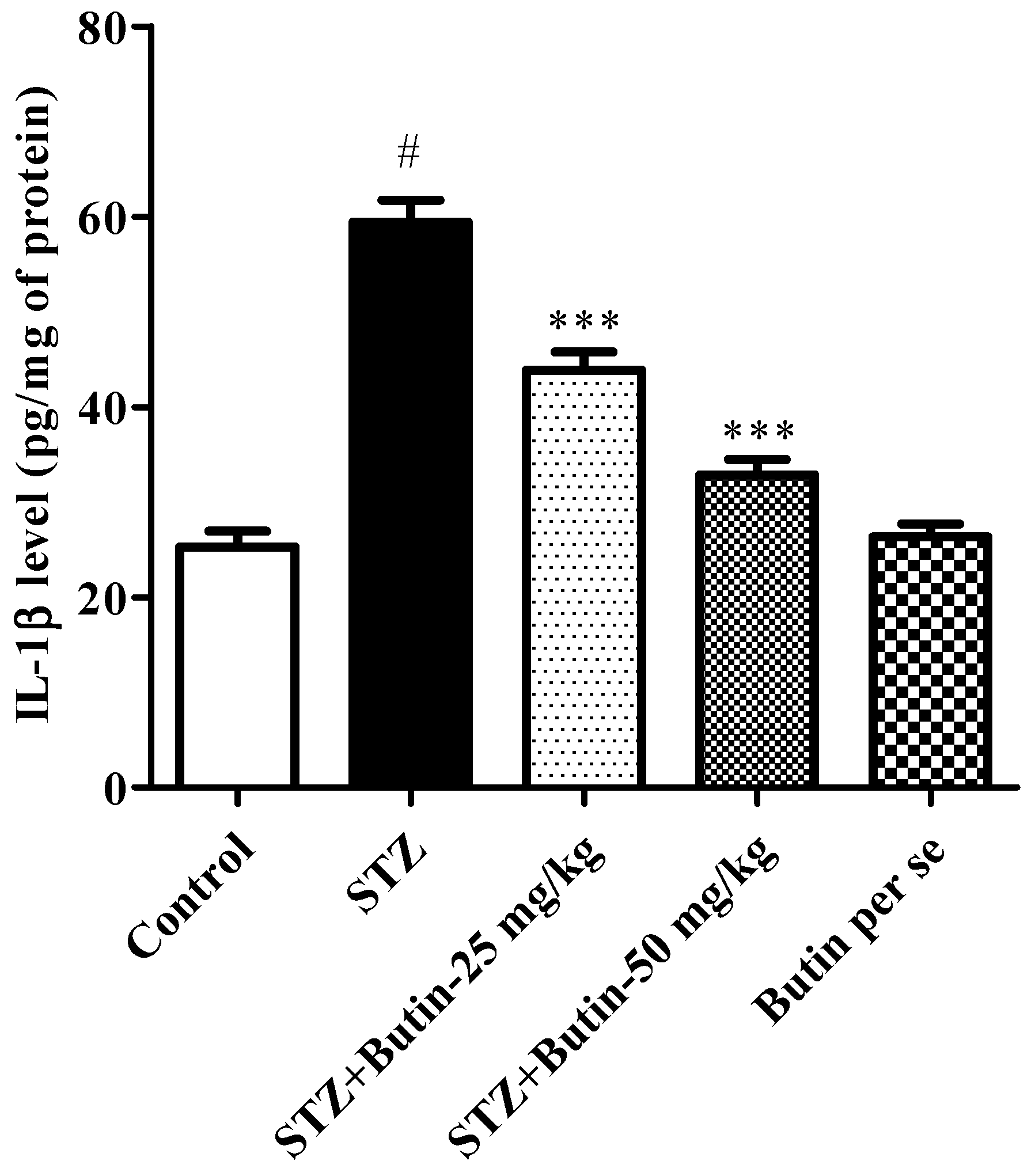
3.3.4. Pro-Inflammatory Cytokines
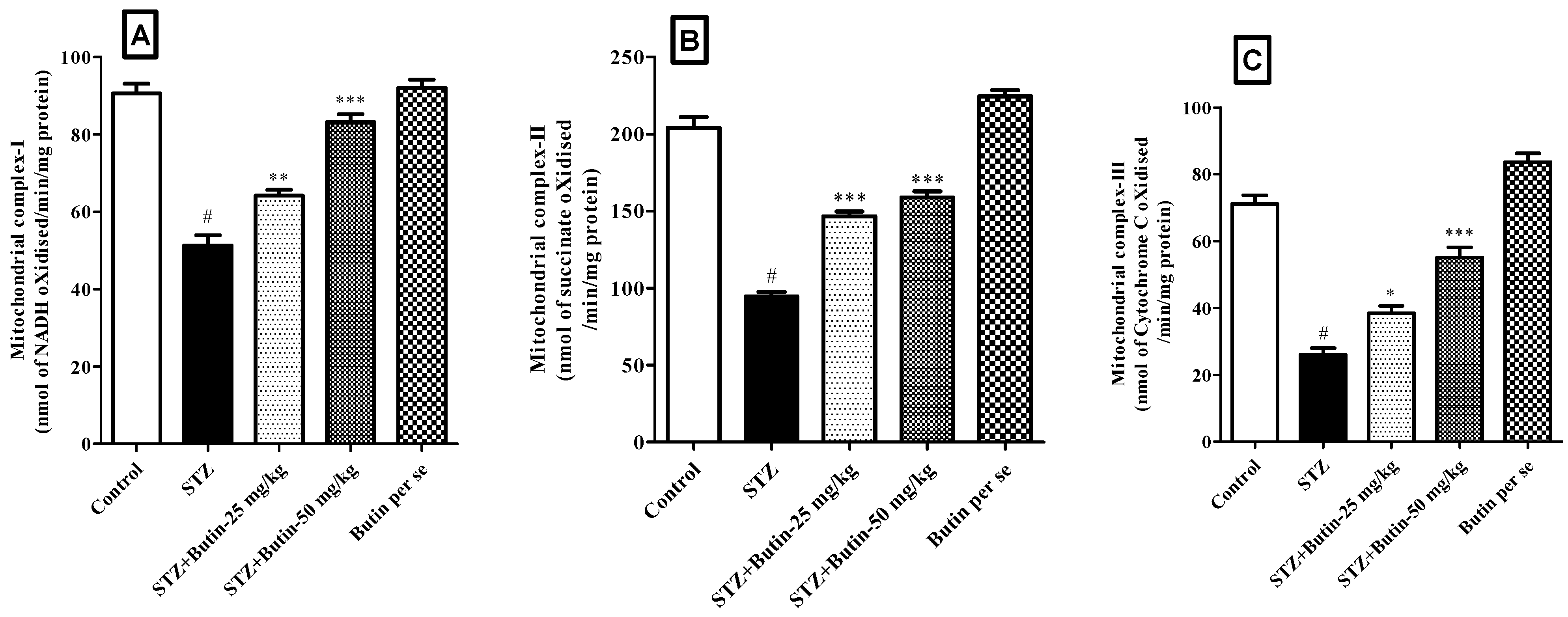
3.3.5. Estimation of Mitochondrial Complex I, II and III
4. Discussion
5. Conclusion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Velmurugan, B.K.; Rathinasamy, B.; Lohanathan, B.P.; Thiyagarajan, V.; Weng, C.-F. Neuroprotective Role of Phytochemicals. Molecules 2018, 23, 2485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Li, N.; Pu, Y.; Zhang, T.; Wang, B. Neuroprotective Effects of Ginseng Phytochemicals: Recent Perspectives. Molecules 2019, 24, 2939. [Google Scholar] [CrossRef] [Green Version]
- Khan, H.; Ullah, H.; Aschner, M.; Cheang, W.S.; Akkol, E.K. Neuroprotective Effects of Quercetin in Alzheimer’s Disease. Biomolecules 2020, 10, 59. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Dong, Y.; Tucker, D.; Wang, R.; Ahmed, M.E.; Brann, D.; Zhang, Q. Treadmill Exercise Exerts Neuroprotection and Regulates Microglial Polarization and Oxidative Stress in a Streptozotocin-Induced Rat Model of Sporadic Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 56, 1469–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.-P.; Yan, T.-H.; Jiang, L.-Y.; Hu, W.; Hu, M.; Wang, C.; Zhang, Q.; Long, Y.; Wang, J.-Q.; Li, Y.-Q.; et al. Pioglitazone ameliorates memory deficits in streptozotocin-induced diabetic mice by reducing brain β-amyloid through PPARγ activation. Acta Pharmacol. Sin. 2013, 34, 455–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfaris, N.A.; Alshammari, G.M.; Altamimi, J.Z.; Aljabryn, D.H.; Alagal, R.I.; Aldera, H.; Alkhateeb, M.A.; Yahya, M.A. Ellagic acid prevents streptozotocin-induced hippocampal damage and memory loss in rats by stimulating Nrf2 and nuclear factor-κB, and activating insulin receptor substrate/PI3K/Akt axis. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2021, 72. [Google Scholar] [CrossRef]
- Moon, Y.; Sung, J.; An, R.; Hernandez, M.E.; Sosnoff, J.J. Gait variability in people with neurological disorders: A systematic review and meta-analysis. Hum. Mov. Sci. 2016, 47, 197–208. [Google Scholar] [CrossRef]
- Chen, Y.; Liang, Z.; Tian, Z.; Blanchard, J.; Dai, C.L.; Chalbot, S.; Iqbal, K.; Liu, F.; Gong, C.X. Intracerebroventricular streptozotocin exacerbates Alzheimer-like changes of 3xTg-AD mice. Mol. Neurobiol. 2014, 49, 547–562. [Google Scholar] [CrossRef] [Green Version]
- Khazdair, M.R.; Anaeigoudari, A.; Hashemzehi, M.; Mohebbati, R. Neuroprotective potency of some spice herbs, a literature review. J. Tradit. Complement. Med. 2019, 9, 98–105. [Google Scholar] [CrossRef]
- Rehman, M.U.; Wali, A.F.; Ahmad, A.; Shakeel, S.; Rasool, S.; Ali, R.; Rashid, S.M.; Madkhali, H.; Ganaie, M.A.; Khan, R. Neuroprotective Strategies for Neurological Disorders by Natural Products: An update. Curr. Neuropharmacol. 2019, 17, 247–267. [Google Scholar] [CrossRef]
- Thippeswamy, B.S.; Nagakannan, P.; Shivasharan, B.D.; Mahendran, S.; Veerapur, V.P.; Badami, S. Protective Effect of Embelin from Embelia ribes Burm. Against Transient Global Ischemia-Induced Brain Damage in Rats. Neurotox. Res. 2011, 20, 379–386. [Google Scholar] [CrossRef]
- Afzal, M.; Al-Abbasi, F.A.; Kazmi, I.; Imam, S.S.; Alshehri, S.; Ghoneim, M.M.; Almalki, W.H.; Nadeem, M.S.; Sayyed, N. Fustin Inhibits Oxidative Free Radicals and Inflammatory Cytokines in Cerebral Cortex and Hippocampus and Protects Cognitive Impairment in Streptozotocin-Induced Diabetic Rats. ACS Chem. Neurosci. 2021, 12, 4587–4597. [Google Scholar] [CrossRef] [PubMed]
- Alharbi, K.S.; Afzal, M.; Alzarea, S.I.; Khan, S.A.; Alomar, F.A.; Kazmi, I. Rosinidin Protects Streptozotocin-Induced Memory Impairment-Activated Neurotoxicity by Suppressing Oxidative Stress and Inflammatory Mediators in Rats. Medicina 2022, 58, 993. [Google Scholar] [CrossRef]
- Shaikh, A.; Dhadde, S.B.; Durg, S.; Veerapur, V.P.; Badami, S.; Thippeswamy, B.S.; Patil, J.S. Effect of Embelin Against Lipopolysaccharide-induced Sickness Behaviour in Mice. Phytother. Res. 2016, 30, 815–822. [Google Scholar] [CrossRef]
- Nagakannan, P.; Shivasharan, B.; Thippeswamy, B.; Veerapur, V. Restoration of Brain Antioxidant Status by Hydroalcoholic Extract of Mimusops elengi Flowers in Rats Treated with Monosodium Glutamate. J. Environ. Pathol. Toxicol. Oncol. 2012, 31, 213–221. [Google Scholar] [CrossRef]
- Ayaz, M.; Sadiq, A.; Junaid, M.; Ullah, F.; Ovais, M.; Ullah, I.; Ahmed, J.; Shahid, M. Flavonoids as Prospective Neuroprotectants and Their Therapeutic Propensity in Aging Associated Neurological Disorders. Front. Aging Neurosci. 2019, 11, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frandsen, J.R.; Narayanasamy, P. Neuroprotection through flavonoid: Enhancement of the glyoxalase pathway. Redox Biol. 2018, 14, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Kang, K.A.; Piao, M.J.; Chang, W.Y.; Maeng, Y.H.; Chae, S.; Lee, I.K.; Kim, B.J.; Hyun, J.W. Butin reduces oxidative stress-induced mitochondrial dysfunction via scavenging of reactive oxygen species. Food Chem. Toxicol. 2010, 48, 922–927. [Google Scholar] [CrossRef]
- Zhang, R.; Lee, I.K.; Piao, M.J.; Kim, K.C.; Kim, A.D.; Kim, H.S.; Chae, S.; Hyun, J.W. Butin (7,3′,4′-Trihydroxydihydroflavone) Reduces Oxidative Stress-Induced Cell Death via Inhibition of the Mitochondria-Dependent Apoptotic Pathway. Int. J. Mol. Sci. 2011, 12, 3871–3887. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.A.; Lee, J.H.; Chae, S.; Zhang, R.; Piao, M.J.; Kim, H.S.; You, H.J.; Hyun, J.W. Butin decreases oxidative stress-induced 8-hydroxy-2′-deoxyguanosine levels via activation of oxoguanine glycosylase 1. Chem. Interact. 2009, 181, 338–342. [Google Scholar] [CrossRef]
- Li, P.; Jiwu, C. Butin Attenuates Brain Edema in a Rat Model of Intracerebral Hemorrhage by Anti Inflammatory Pathway. Transl. Neurosci. 2018, 9, 7–12. [Google Scholar] [CrossRef]
- Cho, N.; Choi, J.H.; Yang, H.; Jeong, E.J.; Lee, K.Y.; Kim, Y.C.; Sung, S.H. Neuroprotective and anti-inflammatory effects of flavonoids isolated from Rhus verniciflua in neuronal HT22 and microglial BV2 cell lines. Food Chem. Toxicol. 2012, 50, 1940–1945. [Google Scholar] [CrossRef]
- Duan, J.; Guan, Y.; Mu, F.; Guo, C.; Zhang, E.; Yin, Y.; Wei, G.; Zhu, Y.; Cui, J.; Cao, J.; et al. Protective effect of butin against ischemia/reperfusion-induced myocardial injury in diabetic mice: Involvement of the AMPK/GSK-3β/Nrf2 signaling pathway. Sci. Rep. 2017, 7, srep41491. [Google Scholar] [CrossRef] [Green Version]
- Alshehri, S.; Al-Abbasi, F.A.; Ghoneim, M.M.; Imam, S.S.; Afzal, M.; Alharbi, K.S.; Nadeem, M.S.; Sayyed, N.; Kazmi, I. Anti-Huntington’s Effect of Butin in 3-Nitropropionic Acid-Treated Rats: Possible Mechanism of Action. Neurotox. Res. 2022, 40, 66–77. [Google Scholar] [CrossRef]
- Deshmukh, R.; Sharma, V.; Mehan, S.; Sharma, N.; Bedi, K. Amelioration of intracerebroventricular streptozotocin induced cognitive dysfunction and oxidative stress by vinpocetine—A PDE1 inhibitor. Eur. J. Pharmacol. 2009, 620, 49–56. [Google Scholar] [CrossRef]
- Binawade, Y.; Jagtap, A. Neuroprotective Effect of Lutein Against 3-Nitropropionic Acid–Induced Huntington’s Disease–Like Symptoms: Possible Behavioral, Biochemical, and Cellular Alterations. J. Med. Food 2013, 16, 934–943. [Google Scholar] [CrossRef]
- Galeano, P.; Martino Adami, P.V.; Do Carmo, S.; Blanco, E.; Rotondaro, C.; Capani, F.; Castaño, E.M.; Cuello, A.C.; Morelli, L. Longitudinal analysis of the behavioral phenotype in a novel transgenic rat model of early stages of Alzheimer’s disease. Front. Behav. Neurosci. 2014, 8, 321. [Google Scholar] [CrossRef] [Green Version]
- Jadiswami, C.; Megha, H.M.; Dhadde, S.B.; Durg, S.; Potadar, P.P.; Thippeswamy, B.S.; Veerapur, V.P. Piroxicam attenuates 3-nitropropionic acid-induced brain oxidative stress and behavioral alteration in mice. Toxicol. Mech. Methods 2014, 24, 672–678. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Misra, H.P.; Fridovich, I. The Role of Superoxide Anion in the Autoxidation of Epinephrine and a Simple Assay for Superoxide Dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Aebi, H.; Wyss, S.R.; Scherz, B.; Skvaril, F. Heterogeneity of Erythrocyte Catalase II. Isolation and Characterization of Normal and Variant Erythrocyte Catalase and Their Subunits. Eur. J. Biochem. 1974, 48, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Wills, E.D. Mechanisms of lipid peroxide formation in animal tissues. Biochem. J. 1966, 99, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Spinazzi, M.; Casarin, A.; Pertegato, V.; Salviati, L.; Angelini, C. Assessment of mitochondrial respiratory chain enzymatic activities on tissues and cultured cells. Nat. Protoc. 2012, 7, 1235–1246. [Google Scholar] [CrossRef]
- Lee, G.-Y.; Lee, C.; Park, G.H.; Jang, J.-H. Amelioration of Scopolamine-Induced Learning and Memory Impairment by α-Pinene in C57BL/6 Mice. Evid.-Based Complement. Altern. Med. 2017, 2017, 4926815. [Google Scholar] [CrossRef] [Green Version]
- Liao, J.; Nai, Y.; Feng, L.; Chen, Y.; Li, M.; Xu, H. Walnut Oil Prevents Scopolamine-Induced Memory Dysfunction in a Mouse Model. Molecules 2020, 25, 1630. [Google Scholar] [CrossRef] [Green Version]
- Rabiei, Z.; Rafieian-Kopaei, M.; Heidarian, E.; Saghaei, E.; Mokhtari, S. Effects of Zizyphus jujube Extract on Memory and Learning Impairment Induced by Bilateral Electric Lesions of the Nucleus Basalis of Meynert in Rat. Neurochem. Res. 2014, 39, 353–360. [Google Scholar] [CrossRef]
- Yildiz, M.; Kiliç, D.; Ünver, Y.; Şentürk, M.; Askin, H.; Küfrevioğlu, Ö.İ. Acetylcholinesterase inhibitory properties of some benzoic acid derivatives. AIP Conf. Proc. 2016, 1726, 20102. [Google Scholar] [CrossRef]
- Janeczek, M.; Gefen, T.; Samimi, M.; Kim, G.; Weintraub, S.; Bigio, E.; Rogalski, E.; Mesulam, M.M.; Geula, C. Variations in Acetylcholinesterase Activity within Human Cortical Pyramidal Neurons Across Age and Cognitive Trajectories. Cereb. Cortex 2018, 28, 1329–1337. [Google Scholar] [CrossRef]
- Georgy, G.S.; Nassar, N.N.; Mansour, H.A.; Abdallah, D.M. Cerebrolysin Ameloriates Cognitive Deficits in Type III Diabetic Rats. PLoS ONE 2013, 8, e64847. [Google Scholar] [CrossRef] [Green Version]
- Karthivashan, G.; Park, S.-Y.; Kweon, M.-H.; Kim, J.; Haque, E.; Cho, D.-Y.; Kim, I.-S.; Cho, E.-A.; Ganesan, P.; Choi, D.-K. Ameliorative potential of desalted Salicornia europaea L. extract in multifaceted Alzheimer’s-like scopolamine-induced amnesic mice model. Sci. Rep. 2018, 8, 7174. [Google Scholar] [CrossRef] [PubMed]
- Yadang, F.S.A.; Nguezeye, Y.; Kom, C.W.; Betote, P.H.D.; Mamat, A.; Tchokouaha, L.R.Y.; Taiwé, G.S.; Agbor, G.A.; Bum, E.N. Scopolamine-Induced Memory Impairment in Mice: Neuroprotective Effects of Carissa edulis (Forssk.) Valh (Apocynaceae) Aqueous Extract. Int. J. Alzheimer’s Dis. 2020, 2020, 6372059. [Google Scholar] [CrossRef]
- Luo, H.; Xiang, Y.; Qu, X.; Liu, H.; Liu, C.; Li, G.; Han, L.; Qin, X. Apelin-13 Suppresses Neuroinflammation Against Cognitive Deficit in a Streptozotocin-Induced Rat Model of Alzheimer’s Disease Through Activation of BDNF-TrkB Signaling Pathway. Front. Pharmacol. 2019, 10, 395. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.M.; Barnes, K.; De Marco, M.; Shaw, P.J.; Ferraiuolo, L.; Blackburn, D.J.; Venneri, A.; Mortiboys, H. Mitochondrial Dysfunction in Alzheimer’s Disease: A Biomarker of the Future? Biomedicines 2021, 9, 63. [Google Scholar] [CrossRef]
- Morgen, K.; Frölich, L. The metabolism hypothesis of Alzheimer’s disease: From the concept of central insulin resistance and associated consequences to insulin therapy. J. Neural Transm. 2015, 122, 499–504. [Google Scholar] [CrossRef]
- Johnson, E.C.B.; Dammer, E.B.; Duong, D.; Ping, L.; Zhou, M.; Yin, L.; Higginbotham, L.A.; Guajardo, A.; White, B.; Troncoso, J.C.; et al. Large-scale proteomic analysis of Alzheimer’s disease brain and cerebrospinal fluid reveals early changes in energy metabolism associated with microglia and astrocyte activation. Nat. Med. 2020, 26, 769–780. [Google Scholar] [CrossRef]
- Cadonic, C.; Sabbir, M.G.; Albensi, B.C. Mechanisms of Mitochondrial Dysfunction in Alzheimer’s Disease. Mol. Neurobiol. 2016, 53, 6078–6090. [Google Scholar] [CrossRef]
- van Groen, T.; Miettinen, P.; Kadish, I. Transgenic AD model mice, effects of potential anti-AD treatments on inflammation, and pathology. J. Alzheimer’s Dis. 2011, 24, 301–313. [Google Scholar] [CrossRef]
- Mishra, A.; Brinton, R.D. Inflammation: Bridging Age, Menopause and APOEε4 Genotype to Alzheimer’s Disease. Front. Aging Neurosci. 2018, 10, 312. [Google Scholar] [CrossRef]
- McGeer, P.L.; Rogers, J.; McGeer, E.G. Inflammation, Antiinflammatory Agents, and Alzheimer’s Disease: The Last 22 Years. J. Alzheimer’s Dis. 2016, 54, 853–857. [Google Scholar] [CrossRef] [PubMed]
- Chu, S.; Gu, J.; Feng, L.; Liu, J.; Zhang, M.; Jia, X.; Liu, M.; Yao, D. Ginsenoside Rg5 improves cognitive dysfunction and beta-amyloid deposition in STZ-induced memory impaired rats via attenuating neuroinflammatory responses. Int. Immunopharmacol. 2014, 19, 317–326. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omer, A.B.; Dalhat, M.H.; Khan, M.K.; Afzal, O.; Altamimi, A.S.A.; Alzarea, S.I.; Almalki, W.H.; Kazmi, I. Butin Mitigates Memory Impairment in Streptozotocin-Induced Diabetic Rats by Inhibiting Oxidative Stress and Inflammatory Responses. Metabolites 2022, 12, 1050. https://doi.org/10.3390/metabo12111050
Omer AB, Dalhat MH, Khan MK, Afzal O, Altamimi ASA, Alzarea SI, Almalki WH, Kazmi I. Butin Mitigates Memory Impairment in Streptozotocin-Induced Diabetic Rats by Inhibiting Oxidative Stress and Inflammatory Responses. Metabolites. 2022; 12(11):1050. https://doi.org/10.3390/metabo12111050
Chicago/Turabian StyleOmer, Asma B., Mahmood Hassan Dalhat, Mohammad Kaleem Khan, Obaid Afzal, Abdulmalik S. A. Altamimi, Sami I. Alzarea, Waleed Hassan Almalki, and Imran Kazmi. 2022. "Butin Mitigates Memory Impairment in Streptozotocin-Induced Diabetic Rats by Inhibiting Oxidative Stress and Inflammatory Responses" Metabolites 12, no. 11: 1050. https://doi.org/10.3390/metabo12111050