
Physiological and Proteomic Responses of Dairy Buffalo to Heat Stress Induced by Different Altitudes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Management
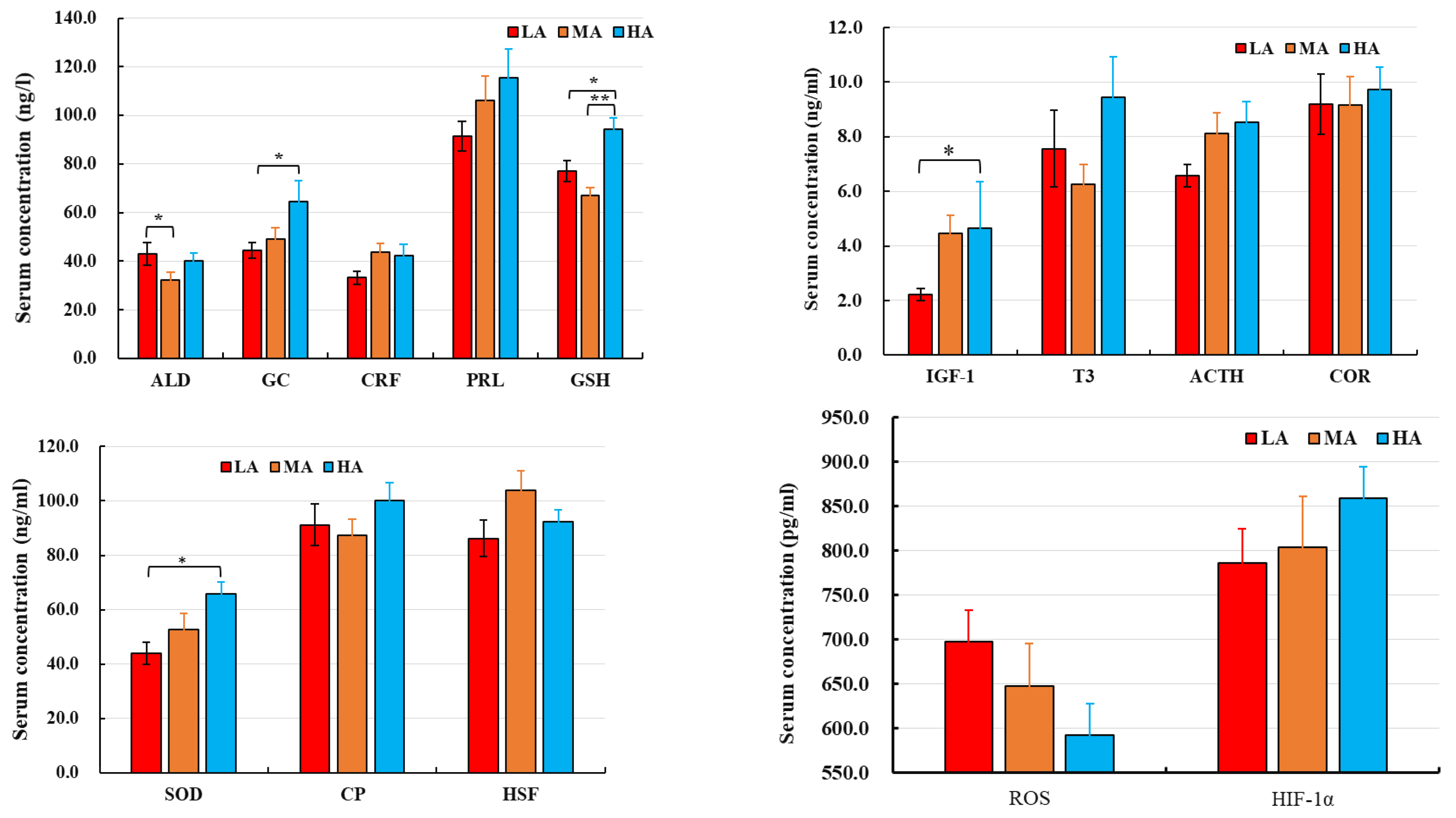
2.2. ELISA Analysis of Serum Parameters
2.3. Sample Preparation and Fractionation for DDA Library Generation
2.4. Data-Dependent Acquisition Mass Spectrometry Assay
2.5. Mass Spectrometry Assay for Data-Independent Acquisition
2.6. Mass Spectrometry Data Analysis
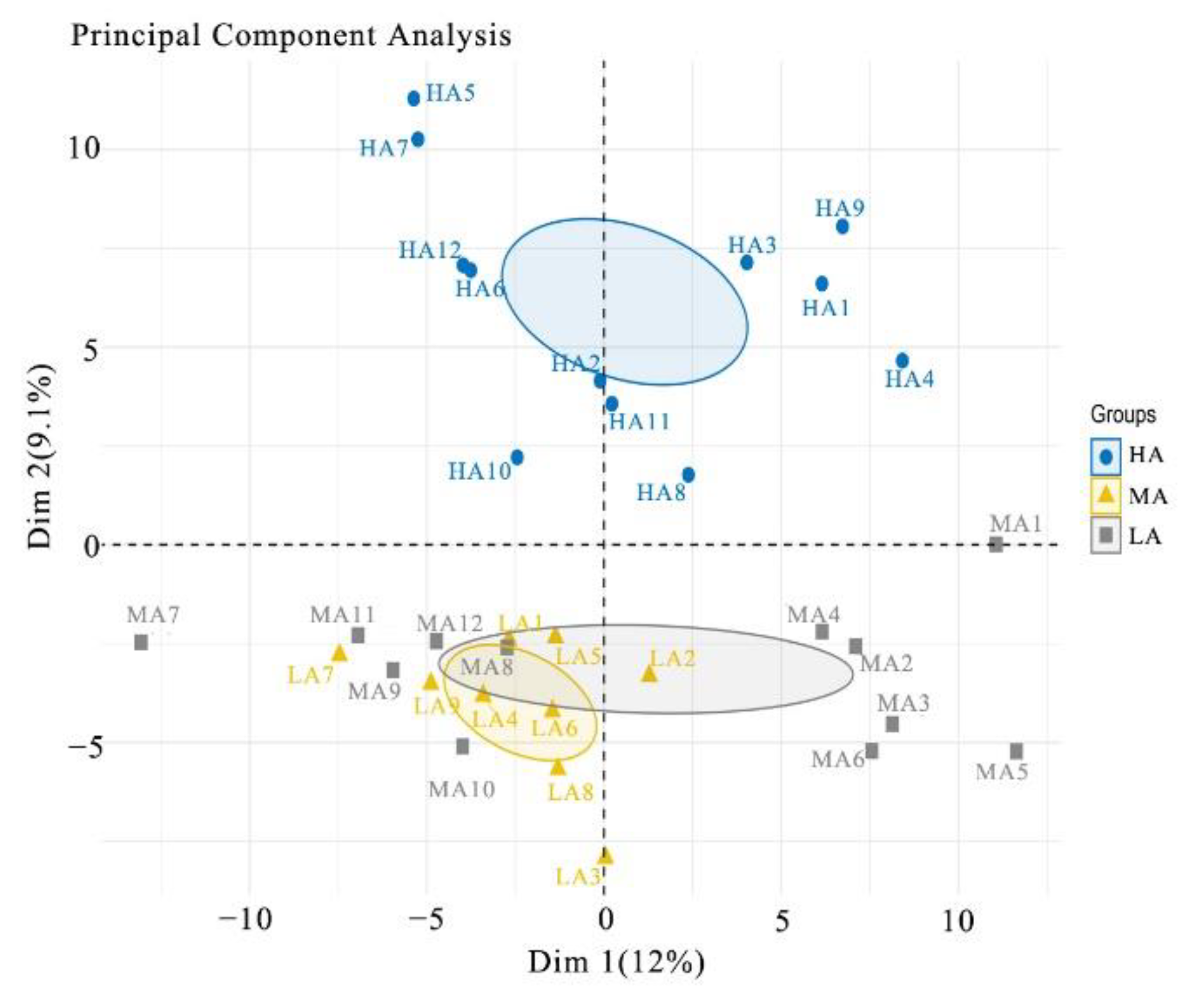
2.7. Principal Component Analysis, GO and KEGG Pathway Annotation
2.8. LC-PRM/MS Quantitative Validation of Targeted Proteins
3. Results
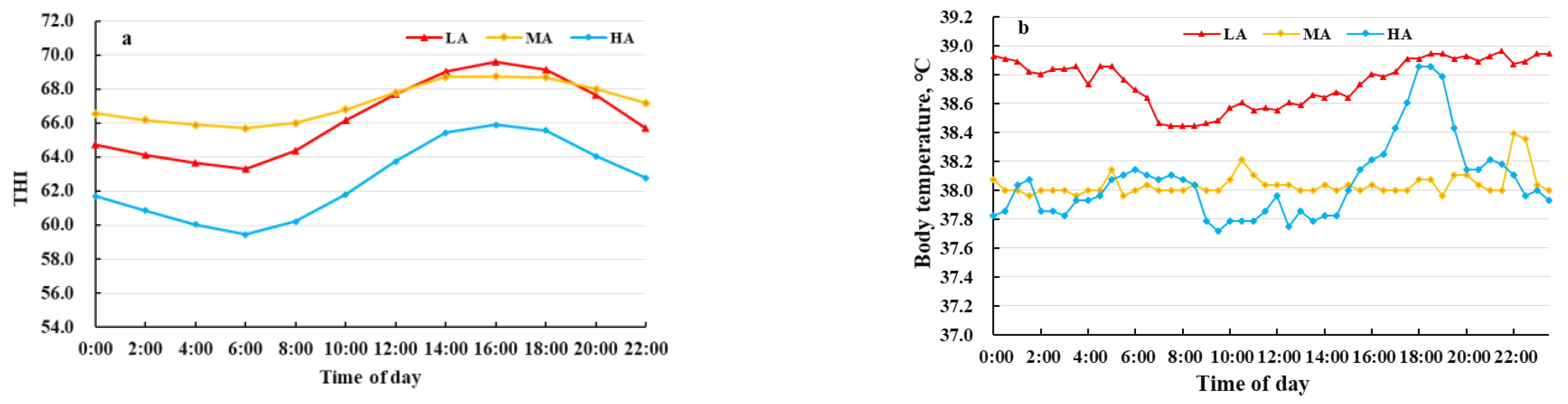
3.1. Physiological Parameters of Buffalo Raised at Different Altitudes
3.2. Spectral Library Construction of DIA Proteomics
3.3. Data Reliability and Principal Component Analysis
3.4. Differential Protein Identification
3.5. Gene Ontology Analysis
3.6. Quantitative Analysis of Targeted Proteins with Parallel Reaction Monitoring
4. Discussion
4.1. Blood Physiological Parameters of Buffalo
4.2. Blood Immunoglobulins of Buffalo
4.3. Blood Oxygen–Delivery Proteins of Buffalo
4.4. Serum Apolipoproteins of Buffalo
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ménard, O.; Ahmad, S.; Rousseau, F.; Briard-Bion, V.; Gaucheron, F.; Lopez, C. Buffalo vs. cow milk fat globules: Size distribution, zeta-potential, compositions in total fatty acids and in polar lipids from the milk fat globule membrane. Food Chem. 2010, 120, 544–551. [Google Scholar] [CrossRef]
- Khedkar, C.D.; Kalyankar, S.D.; Deosarkar, S.S. Buffalo milk. In Encyclopedia of Food and Health; Caballero, B., Finglas, P., Toldrá, F., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 1, pp. 522–528. [Google Scholar]
- Lallo, C.H.O.; Cohen, J.; Rankine, D.; Taylor, M.; Cambell, J.; Stephenson, T. Characterizing heat stress on livestock using the temperature humidity index (THI)—Prospects for a warmer Caribbean. Reg. Environ. Chang. 2018, 18, 2329–2340. [Google Scholar] [CrossRef]
- Desalegn, K. The climate change impacts on livestock production: A Review. Glob. Vet. 2016, 16, 206–212. [Google Scholar]
- Chanda, T.; Debnath, G.K.; Khan, K.I.; Rahman, M.M.; Chanda, G.C. Impact of heat stress on milk yield and composition in early lactation of Holstein Friesian crossbred cattle. Bangladesh J. Anim. Sci. 2017, 46, 192–197. [Google Scholar] [CrossRef]
- Marai, I.F.M.; Haeeb, A.A.M. Buffalo’s biological functions as affected by heat stress-a review. Livest. Sci. 2010, 127, 89–109. [Google Scholar] [CrossRef]
- Payne, W.J.A. Cattle and Buffalo Meat Production in the Tropic; Intermediate Tropical Agriculture Series; Longman Scientific & Technical: Harlow, UK, 1990. [Google Scholar]
- De Rensis, F.; Scaramuzzi, R.J. Heat stress and seasonal effects on reproduction in the dairy cow—A review. Theriogenology 2003, 60, 1139–1151. [Google Scholar] [CrossRef]
- Chauhan, S.S.; Celi, P.; Leury, B.J.; Clarke, I.J.; Dunshea, F.R. Dietary antioxidants at supranutritional doses improve oxidative status and reduce the negative effects of heat stress in sheep. J. Anim Sci. 2014, 92, 4897–4908. [Google Scholar] [CrossRef]
- Dosek, A.; Ohno, H.; Acs, Z.; Taylor, A.W.; Radak, Z. High altitude and oxidative stress. Respir. Physiol. Neurobiol. 2017, 158, 128–131. [Google Scholar] [CrossRef]
- Gaur, P.; Prasad, S.; Kumar, B.; Sharma, S.K.; Vats, P. High-altitude hypoxia induced reactive oxygen species generation, signaling, and mitigation approaches. Int. J. Biometeorol. 2021, 65, 601–615. [Google Scholar] [CrossRef]
- Qiao, G.H.; Yu, C.Q.; Li, J.H.; Yang, X.; Zhu, X.Q.; Zhou, X.H. Effect of high altitude on nutrient digestibility, rumen fermentation and basal metabolism rate in Chinese Holstein cows on the Tibetan plateau. Anim. Prod. Sci. 2013, 53, 240–246. [Google Scholar] [CrossRef]
- Agarwal, S.; Ganesh, S. Perinuclear mitochondrial clustering, increased ROS levels, and HIF1 are required for the activation of HSF1 by heat stress. J. Cell Sci. 2020, 133, 245589. [Google Scholar] [CrossRef]
- Muriithi, N.J.; Maina, G.S.; Maina, M.B.; Kiambi, M.J.; Eliud, N.M. Determination of hematological effects of methanolic leaf extract of Vernonia lasiopus in normal mice. J. Blood. Lymph. 2015, 5, 139. [Google Scholar]
- Sidorenko, S.V.; Ziganshin, R.H.; Luneva, O.G.; Deev, L.I.; Alekseeva, N.V.; Maksimov, G.V.; Grygorczyk, R.; Orlovabe, S.N. Proteomics-based identification of hypoxia-sensitive membrane-bound proteins in rat erythrocytes. J. Proteomics. 2018, 30, 25–33. [Google Scholar] [CrossRef]
- Searle, B.C.; Pino, L.K.; Egertson, J.D.; Ting, Y.S.; Lawrence, R.T.; MacLean, B.X.; Villen, J.; MacCross, M.J. Chromatogram libraries improve peptide detection and quantification by data independent acquisition mass spectrometry. Nat. Commun. 2018, 9, 5128. [Google Scholar] [CrossRef]
- Tucker, C.B.; Rogers, A.R.; Schütz, K.E. Effect of solar radiation on dairy cattle behaviour: Use of shade and body temperature in a pasture-based system. Appl. Anim. Behav. Sci. 2008, 109, 141–154. [Google Scholar] [CrossRef]
- Morignat, E.; Gay, E.; Vinard, J.L.; Calavas, D.; Hénaux, V. Quantifying the influence of ambient temperature on dairy and beef cattle mortality in France from a time-series analysis. Environ Res. 2015, 140, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Gorniak, T.; Meyer, U.; Südekum, K.H.; Dänicke, S. Impact of mild heat stress on dry matter intake, milk yield and milk composition in mid-lactation Holstein dairy cows in a temperate climate. Arch. Anim. Nutr. 2014, 5, 358–369. [Google Scholar] [CrossRef] [PubMed]
- Giraffa, G.; Rossetti, L.; Mucchetti, G.; Addeo, F.; Neviani, E. Influence of the temperature gradient on the growth of thermophilic lactobacilli used as natural starters in grana cheese. J. Dairy Sci. 1998, 81, 31–36. [Google Scholar] [CrossRef]
- Lefaki, M.; Papaevgeniou, N.; Chondrogianni, N. Redox regulation of proteasome function. Redox Biol. 2017, 13, 452–458. [Google Scholar] [CrossRef]
- Ray, P.D.; Huang, B.W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell. Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef]
- Akbarian, A.; Michiels, J.; Degroote, J.; Majdeddin, M.; Golian, A.; De Smet, S. Association between heat stress and oxidative stress in poultry; mitochondrial dysfunction and dietary interventions with phytochemicals. J. Anim. Sci. Biotechnol. 2016, 7, 37. [Google Scholar] [CrossRef] [PubMed]
- Holmström, K.M.; Finkel, T. Cellular mechanisms and physiological consequences of redox-dependent signalling. Nat. Rev. Mol. Cell Biol. 2014, 15, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.R. Critical role of zinc as either an antioxidant or a prooxidant in cellular systems. Oxid. Med. Cell. Longev. 2018, 15, 9156285. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Xu, Y.; Mao, C.; Wang, Z.; Shi, B. Effects of heat stress on antioxidant status and immune function and expression of related genes in lambs. Int. J. Biometeorol. 2020, 64, 2093–2104. [Google Scholar] [CrossRef]
- Peli, A.; Scagliarini, L.; Bergamini, P.F.; Prosperi, A.; Pietra, M. Influence of heat stress on the immunity in growing beef cattle. Large Anim. Rev. 2013, 19, 215–218. [Google Scholar]
- Berman, A. Invited review: Are adaptations present to support dairy cattle productivity in warm climates? J. Dairy Sci. 2011, 94, 2147–2158. [Google Scholar] [CrossRef]
- Dahlbäck, B. C4b-binding protein: A forgotten factor in thrombosis and hemostasis. Semin. Thromb. Hemos. 2011, 37, 355–361. [Google Scholar] [CrossRef]
- Blom, A.M.; Villoutreix, B.O.; Dahlback, B. Functions of human complement inhibitor C4b-binding protein in relation to its structure. Arch. Immunol. Ther. Exp. Engl. Ed. 2004, 2, 83–95. [Google Scholar]
- Huang, X.F.; Wang, Y.Q.; Li, F.F.; Lin, D.; Dai, M.L.; Wang, Q.F.; Jin, Z.B. CFHR2-rs2986127 as a genetic protective marker for acute anterior uveitis in Chinese. J. Gene Med. 2016, 18, 193–198. [Google Scholar] [CrossRef]
- Ceciliani, F.; Lecchi, C. The immune functions of α1 acid glycoprotein. Curr. Protein. Pept. Sci. 2019, 20, 505–524. [Google Scholar] [CrossRef]
- Wong, C.W.; Seow, H.F.; Ai, H.L.; Husband, A.J.; Watson, D.L. Modulation of immune responses by bovine beta-casein. Immunol. Cell. Biol. 1996, 74, 323–329. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Dai, J.; Wei, H.; Tian, C.; Damron, F.; Zhou, J.; Qiu, D. An extracytoplasmic function sigma factor-dependent periplasmic glutathione peroxidase is involved in oxidative stress response of Shewanella Oneidensis. BMC Microbiol. 2015, 15, 1–12. [Google Scholar] [CrossRef]
- Higuchi, A.; Takahashi, K.; Hirashima, M.; Kawakita, T.; Tsubota, K.; Chakravarti, S. Selenoprotein p controls oxidative stress in cornea. PLoS ONE 2010, 5, e9911. [Google Scholar] [CrossRef]
- Lubos, E.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 in health and disease: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox. Sign. 2011, 15, 1957–1997. [Google Scholar] [CrossRef]
- Haffor, A.S.; Al-Johany, A.M. Effect of heat stress, hypoxia and hypoxia-hyperoxia on free radical production in mice mus musculus. J. Med. Sci. 2005, 5, 89–94. [Google Scholar] [CrossRef]
- Liu, P.; Guo, L.L.; Mao, H.M.; Gu, Z.B. Serum proteomics analysis reveals the thermal fitness of dairy buffalo to chronic heat stress. J. Therm. Biol. 2020, 89, 1–5. [Google Scholar] [CrossRef]
- Srikandakumar, A.; Johnson, E.H. Effect of heat stress on milk production, rectal temperature, respiratory rate and blood chemistry in Holstein, Jersey and Australian Milking Zebu cows. Trop. Anim. Health Prod. 2004, 36, 685–692. [Google Scholar] [CrossRef]
- Derkx, F.H.M.; Bouma, B.N.; Tan-Tjiong, H.L.; Veld, A.J.M.I.; Schalekamp, M.A.D.H. Plasma Kallikrein as an Activator of Inactive Renin (“Prorenin”)—Studies In-Vitro and In-Vivo. In Hypertension: Mechanisms and Management. International Boehringer Mannheim Symposia; Philipp, T., Distler, A., Eds.; Springer: Berlin/Heidelberg, Germany, 1980; pp. 149–162. [Google Scholar]
- Dahlbäck, B.; Wiedmer, T.; Sims, P.J. Binding of anticoagulant vitamin K-dependent proteins to platelet-derived microparticles. Biochemistry 1992, 31, 12769–12777. [Google Scholar] [CrossRef]
- Baker, M.E.; French, F.S.; Joseph, D.R. Vitamin K-dependent protein S is similar to rat androgen-binding protein. Biochemistry 1987, 243, 293–296. [Google Scholar] [CrossRef]
- Dahlbäck, B. Vitamin K-dependent protein S: Beyond the protein C pathway. Semin, Thromb. Hemost. 2017, 44, 176–184. [Google Scholar] [CrossRef]
- Yin, Z.F.; Huang, Z.F.; Cui, J. Prothrombotic phenotype of protein Z deficiency. Proc. Natl. Acad. Sci. USA 2000, 97, 6734–6738. [Google Scholar] [CrossRef]
- Zhou, Y.; Luo, G. Apolipoproteins, as the carrier proteins for lipids, are involved in the development of breast cancer. Clin. Transl. Oncol. 2022, 22, 1952–1962. [Google Scholar] [CrossRef]
- Greevenbroek, V.; Marleen, M.J. Lipid metabolism: Pleiotropic effects of high-density lipoproteins. Curr. Opin. Lipidol. 2006, 17, 363–365. [Google Scholar] [CrossRef] [PubMed]
- Ganfornina, M.D.; Carmo, S.D.; Lora, J.M.; Torres-Schumann, S.; Vogel, M.; Allhorn, M.; González, C.; Bastiani, M.J.; Rassart, E.; Sanchez, D. Apolipoprotein D is involved in the mechanisms regulating protection from oxidative stress. Aging Cell 2008, 7, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Bouillon, R.; Pauwels, S. The Vitamin D-Binding Protein, Vitamin D, 4th ed.; Academic Press: Cambridge, MA, USA, 2018; Volume 1, pp. 97–115. [Google Scholar]
- Chou, C.H.; Chuang, L.Y.; Lu, C.Y.; Guh, J.Y. Vitamin D-binding protein is required for the protective effects of vitamin D in renal fibroblasts and is phosphorylated in diabetic rats. Mol. Cell. Endocrinol. 2015, 411, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.E.; Kong, J.; Zhang, W.; Szeto, F.L.; Ye, H.; Deb, D.K.; Matthew, J.B.; Yan, C.L. Targeted expression of human vitamin D receptor in adipocytes decreases energy expenditure and induces obesity in mice. J. Biol. Chem. 2011, 286, 33804–33810. [Google Scholar] [CrossRef]
- Ravid, A. Vitamin D inhibits the activation of stress-activated protein kinases by physiological and environmental stresses in keratinocytes. J. Endocrinol. 2002, 173, 525. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein ID | Protein Name | LA | MA | HA |
|---|---|---|---|---|
| Q28085 | Complement factor H | 5,911,034.4 b | 5,846,954.6 b | 8,003,723.2 a |
| L8I2Z1 | Complement component C8 alpha chain | 822,025.5 b | 916,592.3 b | 1,185,262.4 a |
| L8IHJ7 | Complement factor I | 5,073,044.5 b | 5,423,548.3 b | 6,754,342.4 a |
| L8IE64 | Properdin (Fragment) | 520,308.3 b | 518,567.5 b | 669,974.8 a |
| L8IGE2 | Complement factor H-related protein 3 (Fragment) | 43,900.4 b | 38,432.8 b | 67,400.8 a |
| Q2UVX4 | Complement C3 | 23,107,853.3 ab | 21,185,578.8 b | 26,465,504.0 a |
| F1MM86 | Complement component C6 | 704,760.6 ab | 642,334.4 b | 911,150.4 a |
| P81187 | Complement factor B | 9,508,441.3 b | 10,024,967.0 ab | 11,490,473.8 a |
| L8J0E1 | Complement C5 | 1,908,360.9 b | 2,204,806.2 a | 2,240,134.3 a |
| P17697 | Clusterin regulation of complement activation | 997,871.7 b | 1,381,123.9 a | 1,311,446.6 a |
| L8HKR7 | Ig gamma-3 chain C region (Fragment) | 122,983,076.8 b | 272,235,860.9 ab | 287,926,333.6 a |
| L8HP94 | Ig alpha-1 chain C region (Fragment) | 63,903.4 ab | 78,224.1 a | 55,975.5 b |
| Q7M3F3 | Alpha-lactalbumin | 10,190.8 a | 3491.5 b | 9000.8 ab |
| V5RKP7 | Beta lactoglobulin (Fragment) | 23,405,971.6 ab | 47,755,768.4 a | 8,275,413.5 b |
| C3W955 | Beta-lactoglobulin | 184,142.0 a | 66,021.5 b | 118,728.6 ab |
| F1MHC3 | CD44 antigen | 70,788.3 a | 57,362.6 ab | 44,820.5 b |
| B7VGH4 | Beta-casein | 310,429.6 a | 113,776.7 b | 147,863.8 ab |
| K9ZS80 | K-casein (Fragment) | 24,306.4 a | 11,099.1 ab | 7056.9 b |
| A0A1L6BP00 | Kappa-casein | 24,079.1 ab | 50,215.8 a | 19,399.3 b |
| Q3YJI8 | MHC class I antigen (Fragment) | 459,090.4 b | 1,910,361.4 a | 1,304,512.7 ab |
| D2U6V0 | Alpha-1 acid glycoprotein | 6,132,936.6 a | 4,486,339.8 ab | 3,510,769.9 b |
| Q2KJF1 | Alpha-1B-glycoprotein | 6073654.1 a | 3,382,706.8 b | 5,714,935.8 ab |
| I6SJ59 | Selenoprotein P | 39,427.8 a | 16,819.0 b | 25,330.5 a |
| F8UTU9 | Glutathione peroxidase (Fragment) | 152,540.5 a | 76,484.9 b | 117,009.8 ab |
| A2VE03 | ITCH protein | 13,480,011.6 b | 10,037,581.9 b | 70,279,200.0 a |
| A7MBB0 | VCAM1 protein | 53,086.1 ab | 34,630.1 b | 70,988.3 a |
| L8J552 | Microfibril-associated glycoprotein 4 (Fragment) | 6731.8 b | 19,358.0 a | 12,106.7 b |
| F1MKS5 | Histidine-rich glycoprotein | 3,266,560.6 a | 2,577,096.9 b | 2,840,214.9 ab |
| L8IJH2 | Leucine-rich alpha-2-glycoprotein | 3,057,520.8 a | 2,452,175.9 b | 2,764,774.9 ab |
| G1K1X9 | Vitamin K-dependent protein Z | 445,107.1 a | 421,009.4 b | 351,618.1 a |
| L8IQM7 | Vitamin K-dependent protein S | 795,398.2 b | 792,676.4 b | 921,083.0 a |
| Q3MHN5 | Vitamin D-binding protein | 7,839,167 ab | 7,718,252.2 b | 9,062,119.1 a |
| E1BJN3 | Amine oxidase | 83,957.7 ab | 78,938.2 b | 134,389.1 a |
| L8HRL7 | Amine oxidase (Fragment) | 179,222.3 ab | 150,927.4 b | 243,629.0 a |
| L8HTY4 | Amine oxidase (Fragment) | 2,291,499.2 b | 2,849,165.4 a | 2,660,405 ab |
| Q29437 | “Primary amine oxidase, liver isozyme” | 757,429.0 b | 1,052,047.6 a | 973,569.0 a |
| L8IJ46 | Serum amyloid A protein | 264,612.5 b | 484,433.4 a | 411,107.2 ab |
| L8IP29 | Hepatocyte growth factor activator | 44,423.2 b | 48,712.5 b | 59,188.6 a |
| L8IUV1 | Hepatocyte growth factor-like protein | 82,650.3 b | 73,035.3 b | 100,062.1 a |
| F1MMK9 | Protein AMBP | 3,690,957.5 ab | 3,457,769.0 b | 4,323,694.5 a |
| A8E196 | Adult beta-globin | 7,859,566.5 a | 1,420,398.8 b | 5,154,006.6 ab |
| F1MDA4 | Hemogen | 153,398.1 b | 145,803.7 b | 210,144.2 a |
| D4QBF4 | Haemoglobin beta | 373,158.7 a | 36,534.2 b | 132,260.7 b |
| P04346 | Haemoglobin subunit beta-A | 638,496.4 a | 159,891.8 b | 544,179.7 a |
| P09423 | Haemoglobin subunit alpha-I/II | 8,726,628.5 a | 1,434,904.3 b | 4,482,615.4 ab |
| Q9TSN7 | Haemoglobin subunit alpha-1 | 1,123,791.6 a | 193,289.1 b | 878,525.5 ab |
| Q9TSN8 | Haemoglobin subunit alpha-2 | 7,920,238.3 a | 1,293,871.4 b | 5,010,340.8 a |
| L8HVY9 | Hemopexin | 17,375,065.6 ab | 17,231,204.0 b | 22,307,675.4 a |
| G9DAR3 | Lactoferrin | 44,437,656.0 b | 51,004,339.7 b | 62,055,141.0 a |
| L8HWR0 | Serotransferrin | 78,140,829.8 b | 90,942,537.7 ab | 104,291,760.7 a |
| L8IA26 | Plasminogen (Fragment) | 2,852,684.3 ab | 2,415,660.0 b | 3,391,240.3 a |
| Q2KJ63 | Plasma kallikrein | 472,927.9 b | 461,005.8 b | 609,854.1 a |
| F1MSZ6 | Antithrombin-III | 3,855,682.9 a | 3,194,585.3 ab | 2,450,037.3 b |
| L8I612 | Coagulation factor V | 79,479.8 b | 97,967.3 a | 93,141.4 ab |
| L8IH10 | Coagulation factor IX | 402,115.3 b | 477,317.8 a | 469,924.1 a |
| L8IA55 | Apolipoprotein C-IV | 498,024.1 ab | 644,042.3 a | 361,942.8 b |
| L8I3W2 | Apolipoprotein F (Fragment) | 160,853.6 ab | 238,038.9 a | 105,477.5 b |
| F1MS32 | Apolipoprotein D | 845,238.8 b | 877,878.2 b | 1,551,254.0 a |
| Q58DL9 | Phospholipid transfer protein | 60,817.3 ab | 71,999.2 a | 53,606.6 b |
| L8IXW5 | Proactivator polypeptide | 10,898.0 b | 36,197.7 a | 20,660.7ab |
| F1MNM2 | Phosphatidylinositol-glycan-specific phospholipase D | 46,791.9 ab | 59,825.0 a | 44,146.0 b |
| GO_ID | Gene Ontology Terms | Protein ID | p-Value |
|---|---|---|---|
| LA vs.MA | |||
| 0005344 | Oxygen carrier activity | A8E196, D4QBF4, P04346, P09423, Q9TSN7, Q9TSN8 | 3.25 × 10−7 |
| 0019825 | Oxygen binding | ||
| 0140104 | Molecular carrier activity | 9.39 × 10−7 | |
| 0020037 | Heme binding | 2.78 × 10−5 | |
| 0046906 | Tetrapyrrole binding | 6.87 × 10−5 | |
| 0005833 | Haemoglobin complex | 3.25 × 10−7 | |
| 0005215 | Transporter activity | 0.003 | |
| 0005506 | Iron ion binding | P09423, Q9TSN7, Q9TSN8 | 0.038 |
| 0042612 | MHC class I protein complex | Q3YJI8 | 0.046 |
| 0042611 | MHC protein complex | ||
| MA vs. HA | |||
| 0019825 | Oxygen binding | A8E196, D4QBF4, P04346, P09423, Q9TSN7, Q9TSN8 | 6.38 × 10−9 |
| 0005344 | Oxygen carrier activity | ||
| 0140104 | Molecular carrier activity | 2.76 × 10−8 | |
| 0020037 | Heme binding | 2.15 × 10−6 | |
| 0046906 | Tetrapyrrole binding | 6.62 × 10−6 | |
| 0005215 | Transporter activity | 0.005 | |
| 0005833 | Haemoglobin complex | 6.38 × 10−9 | |
| 0005506 | Iron ion binding | P09423, Q9TSN7, Q9TSN8 | 0.044 |
| LA vs. HA | |||
| 0032682 | Negative regulation of chemokine production | F1MS32 | 0.038 |
| 0032642 | Regulation of chemokine production | ||
| 0071637 | Regulation of monocyte chemotactic protein-1 production | ||
| 0032602 | Chemokine production | ||
| 0071605 | Monocyte chemotactic protein-1 production | ||
| 0002686 | Negative regulation of leukocyte migration | ||
| 2000402 | Negative regulation of lymphocyte migration | ||
| 2000405 | Negative regulation of T cell migration | ||
| 0034439 | Lipoprotein lipid oxidation | ||
| 0060588 | Negative regulation of lipoprotein lipid oxidation | ||
| 0060587 | Regulation of lipoprotein lipid oxidation | ||
| 0006629 | Lipid metabolic process | L8IXW5, A6QLW1, F1MS32 | 0.036 |
| 0033554 | Cellular response to stress | A6QLW1, A2VE03 | 0.042 |
| 0003796 | Lysozyme activity | A0A0C5ADR0 | 0.038 |
| 2000266 | Regulation of blood coagulation | F1MSZ6 | |
| 0019787 | Ubiquitin-like protein transferase activity | A2VE03 | 0.038 |
| 0061659 | Ubiquitin-like protein ligase activity | ||
| 0090085 | Regulation of protein deubiquitination | ||
| DIA Results | PRM Results | ||||||
|---|---|---|---|---|---|---|---|
| Protein ID | Protein Name | LA | MA | HA | LA | MA | HA |
| D2U6V0 | Alpha-1 acid glycoprotein | 6,132,936.6 a | 4,486,339.8 a | 3,510,769.9 b | 0.01275 a | 0.00619 b | 0.00515 b |
| F1MSZ6 | Antithrombin-III | 3,855,682.9 a | 3,194,585.3 a | 2,450,037.3 b | 0.07293 | 0.11303 | 0.08698 |
| E1ACW2 | Lipoprotein lipase | 172,129.6 ab | 271,667.7 a | 79,035.1 b | 0.1019 a | 0.03868 b | 0.08046 a |
| B7VGH4 | Beta-casein | 310,429.6 a | 113,776.7 b | 147,863.8 b | 7.50464 a | 4.04234 b | 6.80145 a |
| L8IHQ3 | Apolipoprotein C-III (Fragment) | 10,570,906.6 a | 14,212,911.3 a | 8,912,108.0 b | 0.07345 a | 0.04655 b | 0.05991 ab |
| A0A1L6BP00 | Kappa-casein | 24,079.1 b | 50,215.8 a | 19,399.3 b | 9.73609 a | 5.85045 b | 7.12996 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lan, Q.; Cao, Z.; Yang, X.; Gu, Z. Physiological and Proteomic Responses of Dairy Buffalo to Heat Stress Induced by Different Altitudes. Metabolites 2022, 12, 909. https://doi.org/10.3390/metabo12100909
Lan Q, Cao Z, Yang X, Gu Z. Physiological and Proteomic Responses of Dairy Buffalo to Heat Stress Induced by Different Altitudes. Metabolites. 2022; 12(10):909. https://doi.org/10.3390/metabo12100909
Chicago/Turabian StyleLan, Qin, Zhiyong Cao, Xiujuan Yang, and Zhaobing Gu. 2022. "Physiological and Proteomic Responses of Dairy Buffalo to Heat Stress Induced by Different Altitudes" Metabolites 12, no. 10: 909. https://doi.org/10.3390/metabo12100909