
Metabolic Influences Modulating Erythrocyte Deformability and Eryptosis

 ,
,
Abstract
:1. Introduction
2. The Main Classical Physicochemical Modifiers of RBC Deformability
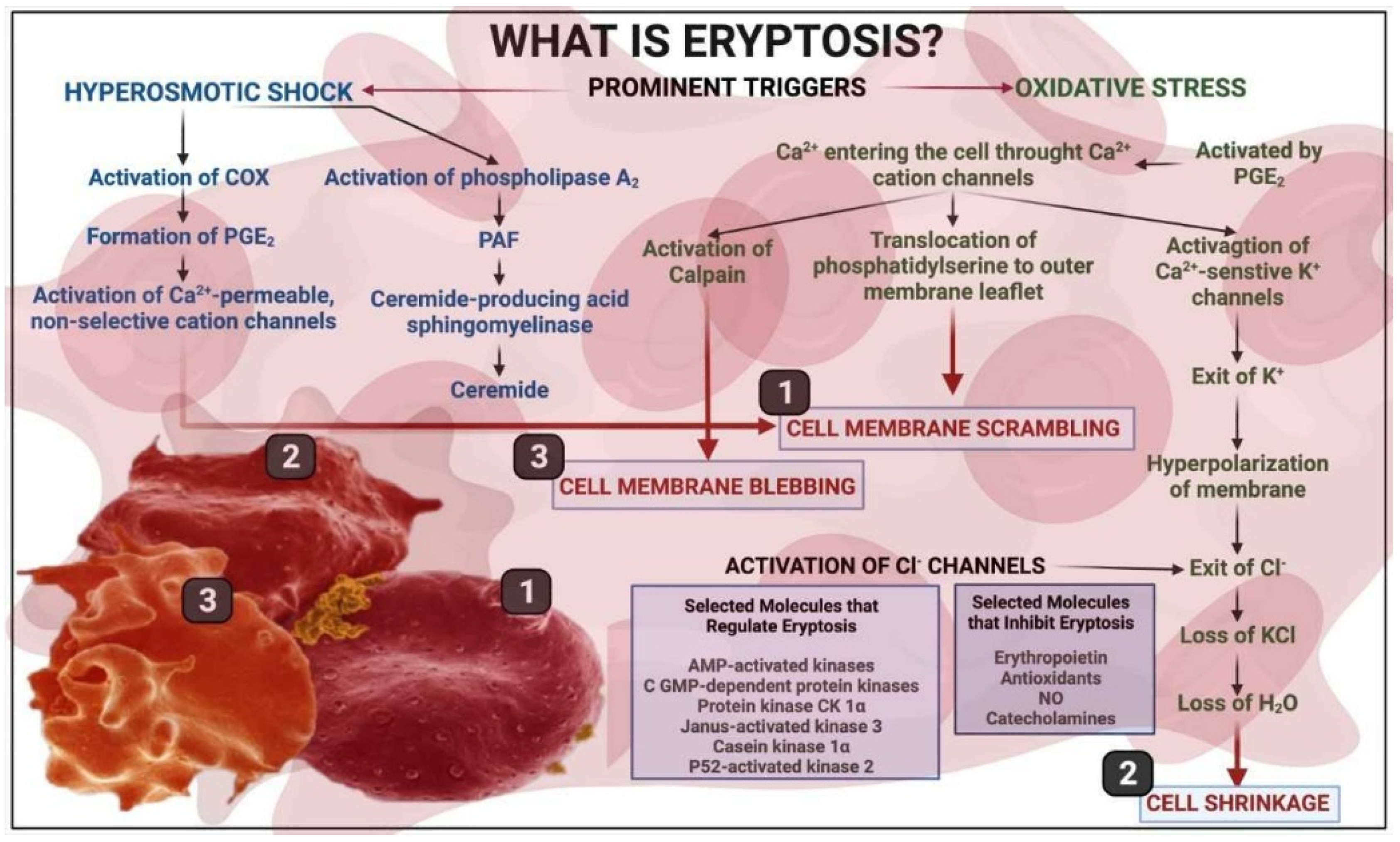
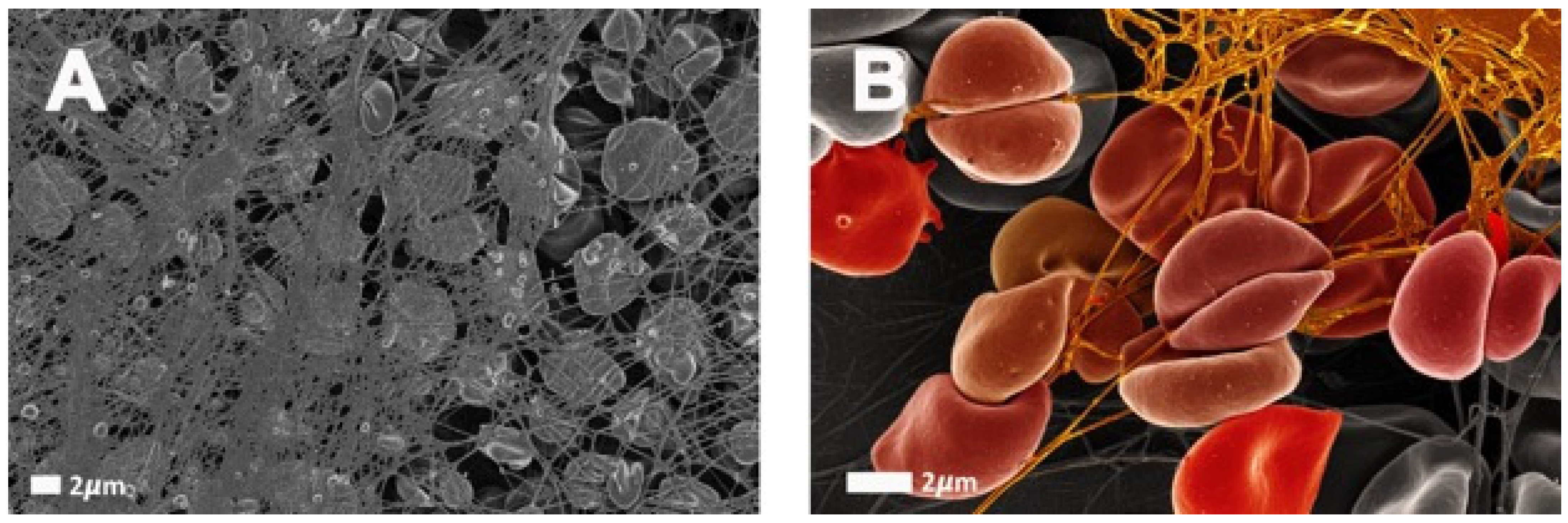
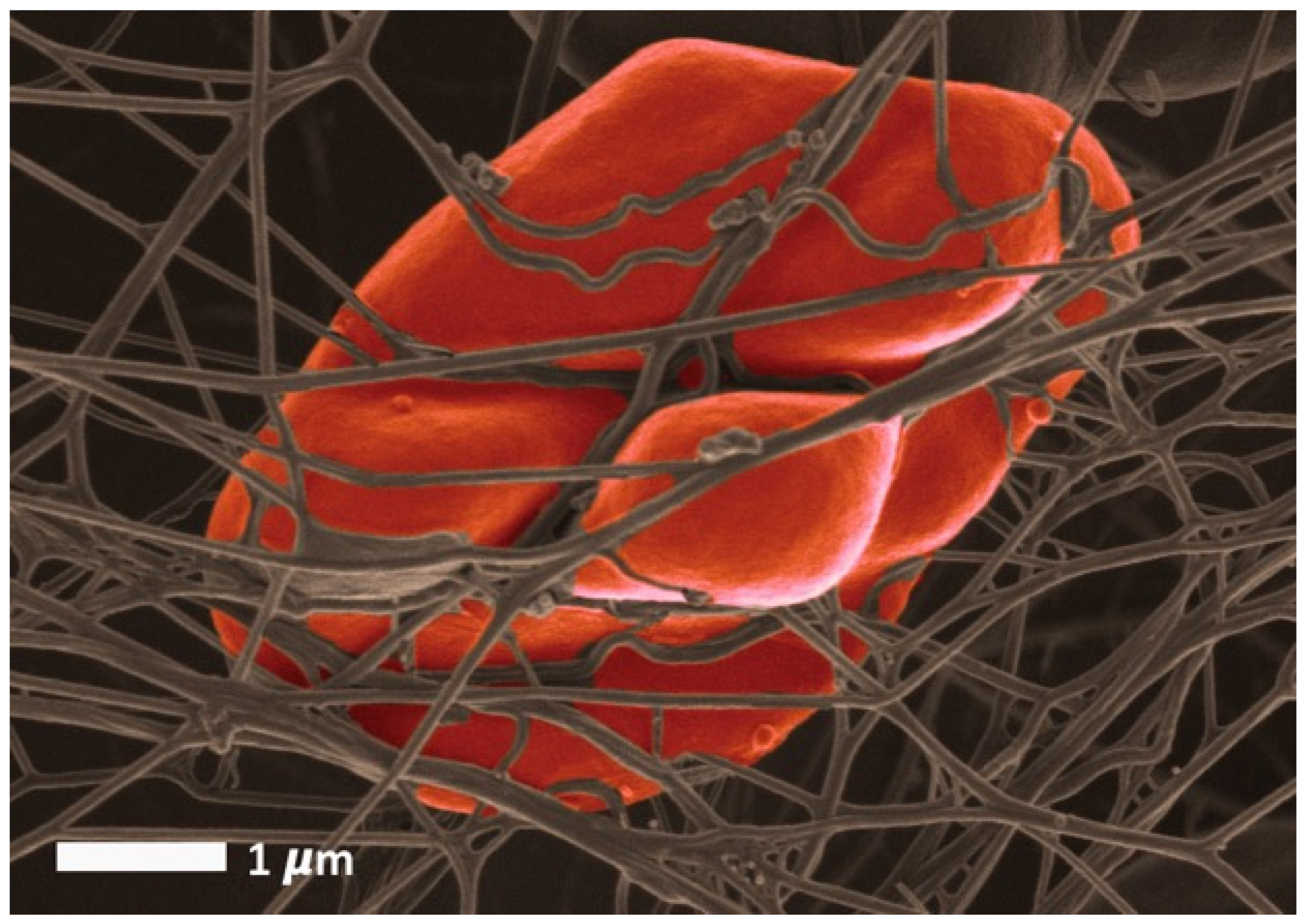
3. A Brief Overview of Eryptosis
4. RBC Receptors
5. Iron and Oxidative Stress as Drivers of RBC Deformability
6. Antioxidants
7. Zinc
8. RBCs and Their Energy Needs
9. RBCs and Circulating Lipids
10. The Effect of Lactate and Ketones on RBCs
11. Nitric Oxide and RBC Function
12. Hormones and Circulating Chemical Messengers
12.1. Insulin and IGF-I
12.2. Glucagon and RBCs
12.3. Thyroid Hormones
12.4. Leptin
12.5. Erythropoietin
12.6. Somatostatin
12.7. Melatonin
12.8. Leukotrienes and Prostaglandins
12.9. Sex Hormones
12.10. Dehydroepiandrosterone
12.11. Apelin
12.12. Catecholamines
12.13. Cortisol
12.14. Endocannabinoids
12.15. Other Hormones
13. RBCs in Various Pathophysiological Situations
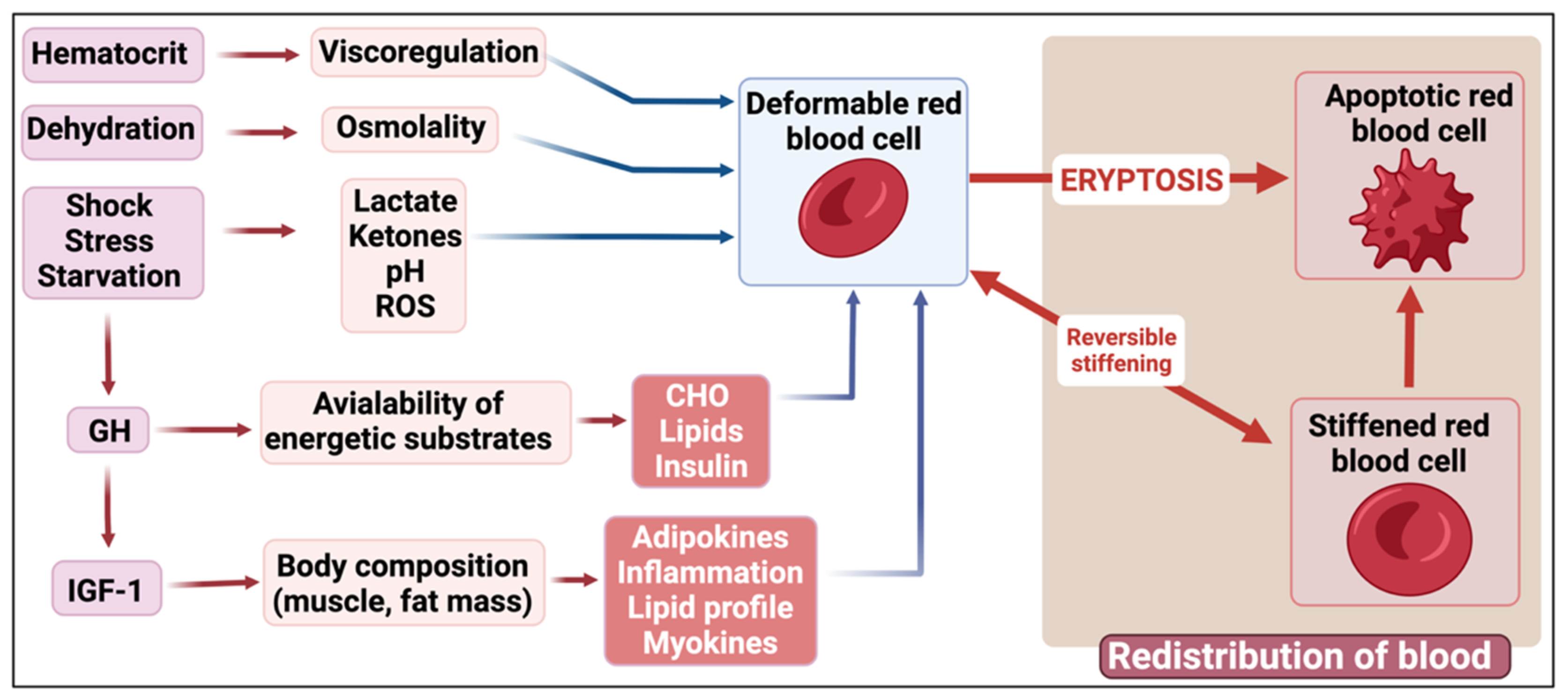
13.1. Stress
13.2. Chronic Fatigue Syndrome
13.3. Septic Shock
13.4. Sleep Apnea
13.5. COVID-19
14. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Huisjes, R.; Bogdanova, A.; Van Solinge, W.W.; Schiffelers, R.M.; Kaestner, L.; Van Wijk, R. Squeezing for Life—Properties of Red Blood Cell Deformability. Front. Physiol. 2018, 9, 656. [Google Scholar] [CrossRef] [PubMed]
- Munter, W.A.; Stein, P.D. Newtonian behavior of blood at high rates of shear. Biorheology 1973, 10, 501–508. [Google Scholar] [CrossRef]
- Skalak, R.; Chien, S. Rheology of blood cells as soft tissues. Biorheology 1982, 19, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Schmid-Schönbein, H.; Gaehtgens, P. What is red cell deformability? Scand. J. Clin. Lab. Investig. 1981, 156, 13–26. [Google Scholar] [CrossRef]
- Nader, E.; Skinner, S.; Romana, M.; Fort, R.; Lemonne, N.; Guillot, N.; Gauthier, A.; Antoine-Jonville, S.; Renoux, C.; Hardy-Dessources, M.D.; et al. Blood Rheology: Key Parameters, Impact on Blood Flow, Role in Sickle Cell Disease and Effects of Exercise. Front. Physiol. 2019, 10, 1329. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Cachia, M.A.; Ge, J.; Xu, Z.; Wang, C.; Sun, Y. Mechanical differences of sickle cell trait (SCT) and normal red blood cells. Lab Chip 2015, 15, 3138–3146. [Google Scholar] [CrossRef] [PubMed]
- Man, Y.; Kucukal, E.; An, R.; Watson, Q.D.; Bosch, J.; Zimmerman, P.A.; Little, J.A.; Gurkan, U.A. Microfluidic assessment of red blood cell mediated microvascular occlusion. Lab Chip 2020, 20, 2086–2099. [Google Scholar] [CrossRef] [PubMed]
- Man, Y.; Kucukal, E.; An, R.; Bode, A.; Little, J.A.; Gurkan, U.A. Standardized microfluidic assessment of red blood cell-mediated microcapillary oc-clusion: Association with clinical phenotype and hydroxyurea responsiveness in sickle cell disease. Microcirculation 2021, 28, e12662. [Google Scholar] [CrossRef]
- Man, Y.; Maji, D.; An, R.; Ahuja, S.P.; Little, J.A.; Suster, M.A.; Mohseni, P.; Gurkan, U.A. Microfluidic electrical impedance assessment of red blood cell-mediated microvascular occlusion. Lab Chip 2021, 21, 1036–1048. [Google Scholar] [CrossRef] [PubMed]
- Lanotte, L.; Mauer, J.; Mendez, S.; Fedosov, D.A.; Fromental, J.M.; Claveria, V.; Nicoud, F.; Gompper, G.; Abkarian, M. Red cells’ dynamic morphologies govern blood shear thinning under microcirculatory flow conditions. Proc. Natl. Acad. Sci. USA 2016, 113, 13289–13294. [Google Scholar] [CrossRef] [Green Version]
- Mauer, J.; Mendez, S.; Lanotte, L.; Nicoud, F.; Abkarian, M.; Gompper, G.; Fedosov, D.A. Flow-Induced Transitions of Red Blood Cell Shapes under Shear. Phys. Rev. Lett. 2018, 121, 118103. [Google Scholar] [CrossRef] [Green Version]
- Chien, S. Red cell deformability and its relevance to blood flow. Annu. Rev. Physiol. 1987, 49, 177–192. [Google Scholar] [CrossRef]
- Connes, P.; Alexy, T.; Detterich, J.; Romana, M.; Hardy-Dessources, M.D.; Ballas, S.K. The role of blood rheology in sickle cell disease. Blood Rev. 2016, 30, 111–118. [Google Scholar] [CrossRef]
- Barshtein, G.; Ben-Ami, R.; Yedgar, S. Role of red blood cell flow behavior in hemodynamics and hemostasis. Expert Rev. Cardiovasc. Ther. 2007, 5, 743–752. [Google Scholar] [CrossRef]
- Le Devehat, C.; Khodabandehlou, T.; Vimeux, M. Relationship between hemorheological and microcirculatory abnormalities in diabetes mellitus. Diabete Metab. 1994, 20, 401–404. [Google Scholar]
- Lang, F.; Abed, M.; Lang, E.; Föller, M. Oxidative stress and suicidal erythrocyte death. Antioxid. Redox Signal. 2014, 21, 138–153. [Google Scholar] [CrossRef]
- Lang, F.; Gulbins, E.; Lang, P.A.; Zappulla, D.; Föller, M. Ceramide in suicidal death of erythrocytes. Cell. Physiol. Biochem. 2010, 26, 21–28. [Google Scholar] [CrossRef]
- Lang, F.; Lang, E.; Foller, M. Physiology and pathophysiology of eryptosis. Transfus. Med. Hemother. 2012, 39, 308–314. [Google Scholar] [CrossRef]
- Lang, E.; Qadri, S.M.; Lang, F. Killing me softly–suicidal erythrocyte death. Int. J. Biochem. Cell Biol. 2012, 44, 1236–1243. [Google Scholar] [CrossRef]
- Pretorius, E.; Du Plooy, J.N.; Bester, J. A Comprehensive Review on Eryptosis. Cell. Physiol. Biochem. 2016, 39, 1977–2000. [Google Scholar] [CrossRef] [PubMed]
- Qadri, S.M.; Bauer, J.; Zelenak, C.; Mahmud, H.; Kucherenko, Y.; Lee, S.H.; Ferlinz, K.; Lang, F. Sphingosine but not sphingosine-1-phosphate stimulates suicidal erythrocyte death. Cell. Physiol. Biochem. 2011, 28, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Qadri, S.M.; Bissinger, R.; Solh, Z.; Oldenborg, P.A. Eryptosis in health and disease: A paradigm shift towards understanding the (patho)physiological implications of programmed cell death of erythrocytes. Blood Rev. 2017, 31, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Qadri, S.M.; Donkor, D.A.; Bhakta, V.; Eltringham-Smith, L.J.; Dwivedi, D.J.; Moore, J.C.; Pepler, L.; Ivetic, N.; Nazi, I.; Fox-Robichaud, A.E.; et al. Phosphatidylserine externalization and procoagulant activation of erythrocytes induced by Pseudomonas aeruginosa virulence factor pyocyanin. J. Cell. Mol. Med. 2016, 20, 710–720. [Google Scholar] [CrossRef] [Green Version]
- Lang, E.; Lang, F. Triggers, inhibitors, mechanisms, and significance of eryptosis: The suicidal erythrocyte death. BioMed Res. Int. 2015, 2015, 513518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duez, J.; Holleran, J.P.; Ndour, P.A.; Pionneau, C.; Diakite, S.; Roussel, C.; Dussiot, M.; Amireault, P.; Avery, V.M.; Buffet, P.A. Mechanical clearance of red blood cells by the human spleen: Potential therapeutic applications of a biomimetic RBC filtration method. Transfus. Clin. Biol. 2015, 22, 151–157. [Google Scholar] [CrossRef]
- Klei, T.R.; Dalimot, J.; Nota, B.; Veldthuis, M.; Mul, F.P.; Rademakers, T.; Hoogenboezem, M.; Nagelkerke, S.Q.; van Ijcken, W.F.; Oole, E.; et al. Hemolysis in the spleen drives erythrocyte turnover. Blood 2020, 136, 1579–1589. [Google Scholar] [CrossRef] [PubMed]
- Miko, I.; Nemeth, N.; Sogor, V.; Kiss, F.; Toth, E.; Peto, K.; Furka, A.; Vanyolos, E.; Toth, L.; Varga, J.; et al. Comparative erythrocyte deformability investigations by filtrometry, slit-flow and rotational ektacytometry in a long-term follow-up animal study on splenectomy and different spleen preserving operative techniques: Partial or subtotal spleen resection and spleen autotransplantation. Clin. Hemorheol. Microcirc. 2017, 66, 83–96. [Google Scholar] [CrossRef] [Green Version]
- Juhan, I.; Vague, P.; Buonocore, M.; Moulin, J.P.; Jouve, R.; Vialettes, B. Abnormalities of erythrocyte deformability and platelet aggregation in insulin-dependent diabetics corrected by insulin in vivo and in vitro. Lancet 1982, 1, 535–537. [Google Scholar] [CrossRef]
- Vague, P.; Juhan, I. Red cell deformability, platelet aggregation, and insulin action. Diabetes 1983, 32 (Suppl. 2), 88–91. [Google Scholar] [CrossRef] [PubMed]
- Brun, J.F.; Varlet-Marie, E.; Connes, P.; Aloulou, I. Hemorheological alterations related to training and overtraining. Biorheology 2010, 47, 95–115. [Google Scholar] [CrossRef] [PubMed]
- Nader, E.; Guillot, N.; Lavorel, L.; Hancco, I.; Fort, R.; Stauffer, E.; Renoux, C.; Joly, P.; Germain, M.; Connes, P. Eryptosis and hemorheological responses to maximal exercise in athletes: Comparison between running and cycling. Scand. J. Med. Sci. Sports 2018, 28, 1532–1540. [Google Scholar] [CrossRef] [PubMed]
- Nader, E.; Monedero, D.; Robert, M.; Skinner, S.; Stauffer, E.; Cibiel, A.; Germain, M.; Hugonnet, J.; Scheer, A.; Joly, P.; et al. Impact of a 10 km running trial on eryptosis, red blood cell rheology, and electrophysiology in endurance trained athletes: A pilot study. Eur. J. Appl. Physiol. 2020, 120, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Nader, E.; Romana, M.; Guillot, N.; Fort, R.; Stauffer, E.; Lemonne, N.; Garnier, Y.; Skinner, S.C.; Etienne-Julan, M.; Robert, M.; et al. Association Between Nitric Oxide, Oxidative Stress, Eryptosis, Red Blood Cell Microparticles, and Vascular Function in Sickle Cell Anemia. Front. Immunol. 2020, 11, 551441. [Google Scholar] [CrossRef]
- Koutsouris, D.; Delatour-Hanss, E.; Hanss, M. Physico-chemical factors of erythrocyte deformability. Biorheology 1985, 22, 119–132. [Google Scholar] [CrossRef]
- Engström, K.G.; Meiselman, H.J. Analysis of red blood cell membrane area and volume regulation using micropipette aspiration and perfusion. Biorheology 1995, 32, 115–116. [Google Scholar] [CrossRef]
- Reinhart, W.H.; Piety, N.Z.; Deuel, J.W.; Makhro, A.; Schulzki, T.; Bogdanov, N.; Goede, J.S.; Bogdanova, A.; Abidi, R.; Shevkoplyas, S.S. Washing stored red blood cells in an albumin solution improves their morphologic and hemorheologic properties. Transfusion 2015, 55, 1872–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugie, J.; Intaglietta, M.; Sung, L.A. Water transport and homeostasis as a major function of erythrocytes. Am. J. Physiol. Heart Circ. Physiol. 2018, 314, H1098–H1107. [Google Scholar] [CrossRef] [Green Version]
- Dupuy-Fons, C.; Brun, J.F.; Mallart, C.; Carvajal, J.; Fussellier, M.; Bardet, L.; Orsetti, A. In vitro influence of zinc and magnesium on the deformability of red blood cells artificially hardened by heating. Biol. Trace Elem. Res. 1995, 47, 247–255. [Google Scholar] [CrossRef]
- Semenov, A.N.; Shirshin, E.A.; Muravyov, A.V.; Priezzhev, A.V. The Effects of Different Signaling Pathways in Adenylyl Cyclase Stimulation on Red Blood Cells Deformability. Front. Physiol. 2019, 10, 923. [Google Scholar] [CrossRef] [Green Version]
- Bentzen, P.J.; Lang, F. Effect of anandamide on erythrocyte survival. Cell. Physiol. Biochem. 2007, 20, 1033–1042. [Google Scholar] [CrossRef]
- Jemaà, M.; Fezai, M.; Lang, F. Inhibition of Suicidal Erythrocyte Death by Reversine. Cell. Physiol. Biochem. 2017, 41, 2363–2373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiedaisch, V.; Akel, A.; Niemoeller, O.M.; Wieder, T.; Lang, F. Zinc-induced suicidal erythrocyte death. Am. J. Clin. Nutr. 2008, 87, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, P.A.; Kempe, D.S.; Akel, A.; Klarl, B.A.; Eisele, K.; Podolski, M.; Hermle, T.; Niemoeller, O.M.; Attanasio, P.; Huber, S.M.; et al. Inhibition of erythrocyte “apoptosis” by catecholamines. Naunyn. Schmiedeberg’s Arch. Pharmacol. 2005, 372, 228–235. [Google Scholar] [CrossRef]
- Nicolay, J.P.; Liebig, G.; Niemoeller, O.M.; Koka, S.; Ghashghaeinia, M.; Wieder, T.; Haendeler, J.; Busse, R.; Lang, F. Inhibition of suicidal erythrocyte death by nitric oxide. Pflugers Arch. 2008, 456, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Niemoeller, O.M.; Bentzen, P.J.; Lang, E.; Lang, F. Adenosine protects against suicidal erythrocyte death. Pflugers Arch. 2007, 454, 427–439. [Google Scholar] [CrossRef]
- Qadri, S.M.; Mahmud, H.; Lang, E.; Gu, S.; Bobbala, D.; Zelenak, C.; Jilani, K.; Siegfried, A.; Foller, M.; Lang, F. Enhanced suicidal erythrocyte death in mice carrying a loss-of-function mutation of the adenomatous polyposis coli gene. J. Cell. Mol. Med. 2012, 16, 1085–1093. [Google Scholar] [CrossRef]
- Pretorius, E.; Swanepoel, A.C.; Buys, A.V.; Vermeulen, N.; Duim, W.; Kell, D.B. Eryptosis as a marker of Parkinson’s disease. Aging 2014, 6, 788–819. [Google Scholar] [CrossRef] [Green Version]
- Makhro, A.; Hänggi, P.; Goede, J.S.; Wang, J.; Brüggemann, A.; Gassmann, M.; Schmugge, M.; Kaestner, L.; Speer, O.; Bogdanova, A. N-methyl-D-aspartate receptors in human erythroid precursor cells and in circulating red blood cells contribute to the intracellular calcium regulation. Am. J. Physiol. Cell Physiol. 2013, 305, C1123–C1138. [Google Scholar] [CrossRef] [Green Version]
- Reinhart, W.H.; Geissmann-Ott, C.; Bogdanova, A. Activation of N-methyl D-aspartate (NMDA) receptors has no influence on rheological properties of erythrocytes. Clin. Hemorheol. Microcirc. 2011, 49, 307–313. [Google Scholar] [CrossRef]
- Hänggi, P.; Makhro, A.; Gassmann, M.; Schmugge, M.; Goede, J.S.; Speer, O.; Bogdanova, A. Red blood cells of sickle cell disease patients exhibit abnormally high abundance of N-methyl D-aspartate receptors mediating excessive calcium uptake. Br. J. Haematol. 2014, 167, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Unal, Y.; Comu, F.M.; Emik, U.; Alkan, M.; Pampal, H.K.; Arslan, M. The effects of propofol and memantine on erythrocyte deformability. Bratisl. Lek. Listy 2014, 115, 253–255. [Google Scholar] [CrossRef] [Green Version]
- Sluyter, R. P2X and P2Y receptor signaling in red blood cells. Front. Mol. Biosci. 2015, 2, 60. [Google Scholar] [CrossRef] [Green Version]
- Aursnes, I.; Gjesdal, K.; Abildgaard, U. Platelet aggregation induced by ADP from unsheared erythrocytes at physiological Ca++-concentration. Br. J. Haematol. 1981, 47, 149–152. [Google Scholar] [CrossRef]
- Sprague, R.S.; Ellsworth, M.L.; Stephenson, A.H.; Lonigro, A.J. ATP: The red blood cell link to NO and local control of the pulmonary circulation. Am. J. Physiol. 1996, 271, H2717–H2722. [Google Scholar] [CrossRef]
- Vulpis, V. Endothelin, microcirculation and hemorheology. Clin. Hemorheol. Microcirc. 1999, 21, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Piskuric, N.A.; Zhang, M.; Vollmer, C.; Nurse, C.A. Potential roles of ATP and local neurons in the monitoring of blood O2 content by rat aortic bodies. Exp. Physiol. 2014, 99, 248–261. [Google Scholar] [CrossRef]
- Wang, L.; Olivecrona, G.; Gotberg, M.; Olsson, M.L.; Winzell, M.S.; Erlinge, D. ADP acting on P2Y13 receptors is a negative feedback pathway for ATP release from human red blood cells. Circ. Res. 2005, 96, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.; Hamazaki, T.; Sawazaki, S.; Nakamura, H. Reduction in the ADP release from shear-stressed red blood cells by fish oil administration. Thromb. Res. 1992, 65, 353–364. [Google Scholar] [CrossRef]
- Burnstock, G. Blood cells: An historical account of the roles of purinergic signalling. Purinergic Signal. 2015, 11, 411–434. [Google Scholar] [CrossRef] [Green Version]
- Lockwood, S.Y.; Erkal, J.L.; Spence, D.M. Endothelium-derived nitric oxide production is increased by ATP released from red blood cells incubated with hydroxyurea. Nitric Oxide 2014, 38, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.C. Identification and characterization of human erythrocyte muscarinic receptors. Gen. Pharmacol. 1986, 17, 281–285. [Google Scholar] [CrossRef]
- Kersh, G.J.; Tomich, J.M.; Montal, M. The M2 delta transmembrane domain of the nicotinic cholinergic receptor forms ion channels in human erythrocyte membranes. Biochem. Biophys. Res. Commun. 1989, 162, 352–356. [Google Scholar] [CrossRef]
- Mesquita, R.; Pires, I.; Saldanha, C.; Martins-Silva, J. Effects of acetylcholine and spermineNONOate on erythrocyte hemorheologic and oxygen carrying properties. Clin. Hemorheol. Microcirc. 2001, 25, 153–163. [Google Scholar]
- Muravyov, A.; Tikhomirova, I. Signaling pathways regulating red blood cell aggregation. Biorheology 2014, 51, 135–145. [Google Scholar] [CrossRef]
- Olson, J.M.; Ciliax, B.J.; Mancini, W.R.; Young, A.B. Presence of peripheral-type benzodiazepine binding sites on human erythrocyte membranes. Eur. J. Pharmacol. 1988, 152, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Dave, J.R.; Eskay, R.L. Demonstration of corticotropin-releasing factor binding sites on human and rat erythrocyte membranes and their modulation by chronic ethanol treatment in rats. Biochem. Biophys. Res. Commun. 1986, 136, 137–144. [Google Scholar] [CrossRef]
- Gopalakrishnan, V.; Ramaswamy, S.; Pillai, N.P.; Ranganathan, S.; Ghosh, M.N. Effect of prolactin on human red cell sodium transport. Experientia 1980, 36, 1423–1425. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.D.; Finch, C.A. Assessing iron status of a population. Am. J. Clin. Nutr. 1979, 32, 2115–2119. [Google Scholar] [CrossRef]
- Kell, D.B. Iron behaving badly: Inappropriate iron chelation as a major contributor to the aetiology of vascular and other progressive inflammatory and degenerative diseases. BMC Med. Genom. 2009, 2, 2. [Google Scholar] [CrossRef]
- Kell, D.B.; Pretorius, E. Serum ferritin is an important inflammatory disease marker, as it is mainly a leakage product from damaged cells. Metallomics 2014, 6, 748–773. [Google Scholar] [CrossRef] [Green Version]
- Powell, R.J.; Machiedo, G.W.; Rush, B.F.; Dikdan, G. Oxygen free radicals: Effect on red cell deformability in sepsis. Crit. Care Med. 1991, 19, 732–735. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, E.; Bester, J.; Vermeulen, N.; Lipinski, B.; Gericke, G.S.; Kell, D.B. Profound morphological changes in the erythrocytes and fibrin networks of patients with hemochromatosis or with hyperferritinemia, and their normalization by iron chelators and other agents. PLoS ONE 2014, 9, e8527. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, E.; Kell, D.B. Diagnostic morphology: Biophysical indicators for iron-driven inflammatory diseases. Integr. Biol. 2014, 6, 486–510. [Google Scholar] [CrossRef] [Green Version]
- Du Plooy, J.N.; Bester, J.; Pretorius, E. Eryptosis in Haemochromatosis: Implications for rheology. Clin. Hemorheol. Microcirc. 2018, 69, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Bekyarova, G.; Yankova, T.; Kozarev, I.; Yankov, D. Reduced erythrocyte deformability related to activated lipid peroxidation during the early postburn period. Burns 1996, 22, 291–294. [Google Scholar] [CrossRef]
- Ham, T.H.; Shen, S.C. Studies on the destruction of red blood cells; thermal injury; action of heat in causing increased spheroidicity, osmotic and mechanical fragilities and hemolysis of erythrocytes; observations on the mechanisms of destruction of such erythrocytes in dogs and in a patient with a fatal thermal burn. Blood 1948, 3, 373–403. [Google Scholar] [PubMed]
- Kaukonen, K.M.; Vaara, S.T.; Pettila, V.; Bellomo, R.; Tuimala, J.; Cooper, D.J.; Krusius, T.; Kuitunen, A.; Reinikainen, M.; Koskenkari, J.; et al. Age of red blood cells and outcome in acute kidney injury. Crit. Care 2013, 17, R222. [Google Scholar] [CrossRef] [Green Version]
- Pettilä, V.; Westbrook, A.J.; Nichol, A.D.; Bailey, M.J.; Wood, E.M.; Syres, G.; Phillips, L.E.; Street, A.; French, C.; Murray, L.; et al. Age of red blood cells and mortality in the critically ill. Crit. Care 2011, 15, R116. [Google Scholar] [CrossRef] [Green Version]
- Bester, J.; Buys, A.V.; Lipinski, B.; Kell, D.B.; Pretorius, E. High ferritin levels have major effects on the morphology of erythrocytes in Alzheimer’s disease. Front. Aging Neurosci. 2013, 5, 88. [Google Scholar] [CrossRef]
- Lipinski, B.; Pretorius, E.; Oberholzer, H.M.; Van Der Spuy, W.J. Interaction of fibrin with red blood cells: The role of iron. Ultrastruct. Pathol. 2012, 36, 79–84. [Google Scholar] [CrossRef]
- Pretorius, E. The adaptability of red blood cells. Cardiovasc. Diabetol. 2013, 12, 63. [Google Scholar] [CrossRef] [Green Version]
- Pretorius, E. Erythrocyte deformability and eryptosis during inflammation, and impaired blood rheology. Clin. Hemorheol. Microcirc. 2018, 69, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, E.; Bester, J.; Vermeulen, N.; Alummoottil, S.; Soma, P.; Buys, A.V.; Kell, D.B. Poorly controlled type 2 diabetes is accompanied by significant morphological and ultrastructural changes in both erythrocytes and in thrombin-generated fibrin: Implications for diagnostics. Cardiovasc. Diabetol. 2015, 14, 30. [Google Scholar] [CrossRef] [Green Version]
- Mcnamee, A.P.; Sabapathy, S.; Singh, I.; Horobin, J.; Guerrero, J.; Simmonds, M.J. Acute Free-Iron Exposure Does Not Explain the Impaired Haemorheology Associated with Haemochromatosis. PLoS ONE 2016, 11, e0146448. [Google Scholar] [CrossRef] [Green Version]
- Pretorius, E.; Vermeulen, N.; Bester, J.; Du Plooy, J.L.; Gericke, G.S. The effect of iron overload on red blood cell morphology. Lancet 2014, 383, 722. [Google Scholar] [CrossRef]
- Gelmini, G.; Quaini, F.; Ricci, R.; Schianchi, L.; Zanichelli, P.; Delsignore, R. Effects of iron-copper free diet on whole blood filtrability in rats. Clin. Hemorheol. Microcirc. 1989, 9, 469. [Google Scholar]
- Wen, Z.Y.; Ma, W.Y.; Sun, D.G.; Chen, J.D. The study on RBC deformability of iron deficiency anemia. Clin. Hemorheol. Microcirc. 1995, 15, 81–87. [Google Scholar] [CrossRef]
- Khaled, S.; Brun, J.F.; Wagner, A.; Mercier, J.; Bringer, J.; Prefaut, C. Increased blood viscosity in iron-depleted elite athletes. Clin. Hemorheol. Microcirc. 1998, 18, 309–318. [Google Scholar]
- Cicha, I.; Suzuki, Y.; Tateishi, N.; Maeda, N. Iron-induced oxidative damage in human red blood cells and the effect of thiol-containing antioxidants. Biorheology 1999, 36, 48. [Google Scholar]
- Kuçukatay, V.; Bor-Kucukatay, M.; Gundogdu, G.; Erken, G.; Ozcan, T.O.; Miloglu, F.D.; Kadioglu, Y. Vitamin E treatment enhances erythrocyte deformability in aged rats. Folia Biol. 2012, 58, 157–165. [Google Scholar]
- Oostenbrug, G.S.; Mensink, R.P.; Hardeman, M.R.; De Vries, T.; Brouns, F.; Hornstra, G. Exercise performance, red blood cell deformability, and lipid peroxidation: Effects of fish oil and vitamin E. J. Appl. Physiol. 1997, 83, 746–752. [Google Scholar] [CrossRef]
- Sentürk, U.K.; Gündüz, F.; Kuru, O.; Aktekin, M.R.; Kipmen, D.; Yalcin, O.; Bor-Kucukatay, M.; Yesilkaya, A.; Baskurt, O.K. Exercise-induced oxidative stress affects erythrocytes in sedentary rats but not exercise-trained rats. J. Appl. Physiol. 2001, 91, 1999–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yerer, M.B.; Aydogan, S. The in vivo antioxidant effectiveness of alpha-tocopherol in oxidative 8stress induced by sodium nitroprusside in rat red blood cells. Clin. Hemorheol. Microcirc. 2004, 30, 323–329. [Google Scholar] [PubMed]
- Begum, A.N.; Terao, J. Protective effect of alpha-tocotrienol against free radical-induced impairment of erythrocyte deformability. Biosci. Biotechnol. Biochem. 2002, 66, 398–403. [Google Scholar] [CrossRef]
- Nanjo, F.; Honda, M.; Okushio, K.; Matsumoto, N.; Ishigaki, F.; Ishigami, T.; Hara, Y. Effects of dietary tea catechins on alpha-tocopherol levels, lipid peroxidation, and erythrocyte deformability in rats fed on high palm oil and perilla oil diets. Biol. Pharm. Bull. 1993, 16, 1156–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sentürk, U.K.; Gündüz, F.; Kuru, O.; Koçer, G.; Ozkaya, Y.G.; Yesilkaya, A.; Bor-Küçükatay, M.; Uyüklü, M.; Yalcin, O.; Baskurt, O.K. Exercise-induced oxidative stress leads hemolysis in sedentary but not trained humans. J. Appl. Physiol. 2005, 99, 1434–1441. [Google Scholar] [CrossRef] [Green Version]
- Brun, J.F.; Fons, C.; Fussellier, M.; Bardet, L.; Orsetti, A. Zinc salts improve in vitro erythrocyte flexibility. Rev. Port. Hemorreol. 1991, 5, 231–238. [Google Scholar]
- Khaled, S.; Brun, J.F.; Micallef, J.P.; Bardet, L.; Cassanas, G.; Monnier, J.F.; Orsetti, A. Serum zinc and blood rheology in sportsmen (football players). Clin. Hemorheol. Microcirc. 1997, 17, 47–58. [Google Scholar]
- Khaled, S.; Brun, J.F.; Cassanas, G.; Bardet, L.; Orsetti, A. Effects of zinc supplementation on blood rheology during exercise. Clin. Hemorheol. Microcirc. 1999, 20, 1–10. [Google Scholar]
- Khaled, S.; Brun, J.F.; Cassanas, G.; Bardet, L.; Mercier, J.; Préfaut, C. In vitro effects of zinc gluconate and acetate on red cell aggregability. Clin. Hemorheol. Microcirc. 2000, 22, 325–329. [Google Scholar] [PubMed]
- Reinhart, S.A.; Schulzki, T.; Bonetti, P.O.; Reinhart, W.H. Studies on metabolically depleted erythrocytes. Clin. Hemorheol. Microcirc. 2014, 56, 161–173. [Google Scholar] [CrossRef]
- Jain, R.K.; Traykov, T.T. Effect of glucose and galactose on RBC deformability. Biorheology 1986, 23, 292. [Google Scholar]
- Robey, C.; Dasmahapatra, A.; Cohen, M.P.; Suarez, S. Sorbinil partially prevents decreased erythrocyte deformability in experimental diabetes mellitus. Diabetes 1987, 36, 1010–1013. [Google Scholar] [CrossRef] [PubMed]
- Schmid-Schönbein, H.; Volger, E. Red-cell aggregation and red-cell deformability in diabetes. Diabetes 1976, 25, 897–902. [Google Scholar] [PubMed]
- Fashing, P.; Kurzemann, S.; Wagner, B.; Wagner, O.; Waldhäusl, W.; Ehringer, H.; Koppensteiner, R. Only marginal influence of metabolic control on blood rheology in insulin-dependent diabetic patient without manifest angiopathy. Clin. Hemorheol. Microcirc. 1995, 15, 3–11. [Google Scholar] [CrossRef]
- Stuart, J.; Juhan-Vague, I. Erythrocyte rheology in diabetes. Biorheolology 1986, 23, 216. [Google Scholar]
- Schmid-Schönbein, H.; Teitel, P. In vitro assessment of “covertly” abnormal blood rheology: Critical appraisal of presently available microrheological methodology. A review focusing on diabetic retinopathy as a possible consequence of rheological occlusion. Clin. Hemorheol. Microcirc. 1987, 7, 203–238. [Google Scholar] [CrossRef]
- Li, Q.; Yang, L.Z. Hemoglobin A1c level higher than 9.05% causes a significant impairment of erythrocyte deformability in diabetes mellitus. Acta Endocrinol. 2018, 14, 66–75. [Google Scholar] [CrossRef]
- Blaslov, K.; Kruljac, I.; Mirošević, G.; Gaćina, P.; Kolonić, S.O.; Vrkljan, M. The prognostic value of red blood cell characteristics on diabetic retinopathy development and progression in type 2 diabetes mellitus. Clin. Hemorheol. Microcirc. 2019, 71, 475–481. [Google Scholar] [CrossRef]
- Calderón-Salinas, J.V.; Muñoz-Reyes, E.G.; Guerrero-Romero, J.F.; Rodríguez-Morán, M.; Bracho-Riquelme, R.L.; Carrera-Gracia, M.A.; Quintanar-Escorza, M.A. Eryptosis and oxidative damage in type 2 diabetic mellitus patients with chronic kidney disease. Mol. Cell. Biochem. 2011, 357, 171–179. [Google Scholar] [CrossRef]
- Restivo, I.; Attanzio, A.; Tesoriere, L.; Allegra, M. Suicidal Erythrocyte Death in Metabolic Syndrome. Antioxidants 2021, 10, 154. [Google Scholar] [CrossRef]
- Khodabandehlou, T.; Zhao, H.; Vimeux, M.; Aouane, F.; Ledévéhat, C. Haemorheological consequences of hyperglycaemic spike in healthy volunteers and insulin-dependent diabetics. Clin. Hemorheol. Microcirc. 1998, 19, 105–114. [Google Scholar]
- Babu, N.; Singh, M. Influence of hyperglycemia on aggregation, deformability and shape parameters of erythrocytes. Clin. Hemorheol. Microcirc. 2004, 31, 273–280. [Google Scholar]
- Shin, S.; Ku, Y.H.; Suh, J.S.; Singh, M. Rheological characteristics of erythrocytes incubated in glucose media. Clin. Hemorheol. Microcirc. 2008, 38, 153–161. [Google Scholar] [PubMed]
- Bigdelou, P.; Farnoud, A.M. Induction of eryptosis in red blood cells using a calcium ionophore. J. Vis. Exp. 2020, 155. [Google Scholar] [CrossRef]
- Wong, P. The basis of echinocytosis of the erythrocyte by glucose depletion. Cell Biochem. Funct. 2011, 29, 708–711. [Google Scholar] [CrossRef]
- Varlet-Marie, E.; Guiraudou, M.; Fédou, C.; Raynaud De Mauverger, E.; Durand, F.; Brun, J.F. Nutritional and metabolic determinants of blood rheology differ between trained and sedentary individuals. Clin. Hemorheol. Microcirc. 2013, 55, 39–54. [Google Scholar] [CrossRef]
- Brun, J.F.; Fons, C.; Supparo, I.; Mallard, C.; Orsetti, A. Relationships between metabolic and hemorheologic modifications associated with overweight. Clin. Hemorheol. Microcirc. 1993, 13, 201–213. [Google Scholar] [CrossRef]
- Forsyth, A.M.; Braunmuller, S.; Wan, J.; Franke, T.; Stone, H.A. The effects of membrane cholesterol and simvastatin on red blood cell deformability and ATP release. Microvasc. Res. 2012, 83, 347–351. [Google Scholar] [CrossRef]
- Guézennec, C.Y.; Nadaud, J.F.; Satabin, P.; Leger, F.; Lafargue, P. Influence of polyunsaturated fatty acid diet on the hemorheological response to physical exercise in hypoxia. Int. J. Sports Med. 1989, 10, 286–291. [Google Scholar] [CrossRef]
- Léger, C.L.; Guézennec, C.Y.; Kadri-Hassani, N.; Satabin, P. Les acides gras phospholipidiques membranaires au cours de l’effort physique de longue durée avec ou sans apport nutritionnel d’huiles de poissons. Cah. Nutr. Diététique 1992, 27, 82–89. [Google Scholar]
- Bakker, N.; Schoorl, M.; Demirkiran, A.; Cense, H.A.; Houdijk, A.P. Erythrocyte deformability and aggregation in morbidly obese women undergoing laparoscopic gastric bypass surgery and effects of oral omega-3 fatty acid supplementation. Clin. Hemorheol. Microcirc. 2020, 75, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Bakker, N.; Schoorl, M.; Stoutjesdijk, E.; Houdijk, A.P. Erythrocyte deformability and aggregability in patients undergoing colon cancer surgery and effects of two infusions with omega-3 fatty acids. Clin. Hemorheol. Microcirc. 2020, 74, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Thoth, K.; Ernst, E.; Habon, T.; Horvath, I.; Juricskay, I.; Mozsic, G. Hemorheological and hemodynamical effects of fish oil (Ameu) in patients with ischemic heart disease and hyperlipoproteinemia. Clin. Hemorheol. Microcirc. 1995, 15, 867–875. [Google Scholar] [CrossRef]
- Schütz, E.; Schuff-Werner, P.; Armstrong, V.W.; Senger, I.; Güttner, Y. Haemorheological changes during postprandial lipemia. In Proceedings of the VIIth European Conference on Clinical Haemorheology, Southampton, UK, 16–19 July 1991; Abstract book. p. 91. [Google Scholar]
- Varlet-Marie, E.; Brun, J.F. Reciprocal relationships between blood lactate and hemorheology in athletes: Another hemorheologic paradox? Clin. Hemorheol. Microcirc. 2004, 30, 331–337. [Google Scholar]
- Brun, J.F.; Fons, C.; Raynaud, E.; Fédou, C.; Orsetti, A. Influence of circulating lactate on blood rheology during exercise in professional football players. Rev. Port Hemorheol. 1991, 5, 219–229. [Google Scholar]
- Connes, P.; Caillaud, C.; Bouix, D.; Kippelen, P.; Mercier, J.; Varray, A.; Préfaut, C.; Brun, J.F. Red cell rigidity paradoxically decreases during maximal exercise in endurance athletes unless they are prone to exercise-induced hypoxaemia. J. Mal. Vasc. 2000, 25 (Suppl. B), 165. [Google Scholar]
- Peyreigne, C.; Bouix, D.; Aïssa Benhaddad, A.; Raynaud, E.; Perez-Martin, A.; Mercier, J.; Brun, J.F. Hemorheologic effects of a short-term ketogenetic diet. Clin. Hemorheol. Microcirc. 1999, 21, 147–153. [Google Scholar] [PubMed]
- McMahon, T.J. Red Blood Cell Deformability, Vasoactive Mediators, and Adhesion. Front. Physiol. 2019, 10, 1417. [Google Scholar] [CrossRef]
- Simmonds, M.J.; Detterich, J.A.; Connes, P. Nitric oxide, vasodilation and the red blood cell. Biorheology 2014, 51, 121–134. [Google Scholar] [CrossRef]
- Chen, K.; Popel, A.S. Nitric oxide production pathways in erythrocytes and plasma. Biorheology 2009, 46, 107–119. [Google Scholar] [CrossRef]
- Ulker, P.; Sati, L.; Celik-Ozenci, C.; Meiselman, H.J.; Baskurt, O.K. Mechanical stimulation of nitric oxide synthesizing mechanisms in erythrocytes. Biorheology 2009, 46, 121–132. [Google Scholar] [CrossRef]
- Baskurt, O.K.; Uyuklu, M.; Meiselman, H.J. Protection of erythrocytes from sub-hemolytic mechanical damage by nitric oxide mediated inhibition of potassium leakage. Biorheology 2004, 41, 79–89. [Google Scholar] [PubMed]
- Bor-Kucukatay, M.; Wenby, R.B.; Meiselman, H.J.; Baskurt, O.K. Effects of nitric oxide on red blood cell deformability. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H1577–H1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korbut, R.; Gryglewski, R.J. Nitric oxide from polymorphonuclear leukocytes modulates red blood cell deformability in vitro. Eur. J. Pharmacol. 1993, 234, 17–22. [Google Scholar] [CrossRef]
- Mesquita, R.; Picarra, B.; Saldanha, C.; Martins, E.; Silva, J. Nitric oxide effects on human erythrocytes structural and functional properties--an in vitro study. Clin. Hemorheol. Microcirc. 2002, 27, 137–147. [Google Scholar]
- Bor-Kucukatay, M.; Meiselman, H.J.; Başkurt, O.K. Modulation of density-fractionated RBC deformability by nitric oxide. Clin. Hemorheol. Microcirc. 2005, 33, 363–367. [Google Scholar]
- Rey, J.; Buffet, P.A.; Ciceron, L.; Milon, G.; Mercereau-Puijalon, O.; Safeukui, I. Reduced erythrocyte deformability associated with hypoargininemia during Plasmodium falciparum malaria. Sci. Rep. 2014, 4, 3767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grau, M.; Mozar, A.; Charlot, K.; Lamarre, Y.; Weyel, L.; Suhr, F.; Collins, B.; Jumet, S.; Hardy-Dessources, M.D.; Romana, M.; et al. High red blood cell nitric oxide synthase activation is not associated with improved vascular function and red blood cell deformability in sickle cell anaemia. Br. J. Haematol. 2015, 168, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Mozar, A.; Connes, P.; Collins, B.; Hardy-Dessources, M.D.; Romana, M.; Lemonne, N.; Bloch, W.; Grau, M. Red blood cell nitric oxide synthase modulates red blood cell deformability in sickle cell anemia. Clin. Hemorheol. Microcirc. 2016, 64, 47–53. [Google Scholar] [CrossRef]
- Bor-Küçükatay, M.; Yalçin, O.; Gökalp, O.; Kipmen-Korgun, D.; Yesilkaya, A.; Baykal, A.; Ispir, M.; Senturk, U.K.; Kaputlu, I.; Başkurt, O.K. Red blood cell rheological alterations in hypertension induced by chronic inhibition of nitric oxide synthesis in rats. Clin. Hemorheol. Microcirc. 2000, 22, 267–275. [Google Scholar]
- Tai, Y.H.; Chu, Y.H.; Wu, H.L.; Lin, S.M.; Tsou, M.Y.; Huang, C.H.; Chang, H.H.; Lu, C.C. High-dose nitroglycerin administered during rewarming preserves erythrocyte deformability in cardiac surgery with cardiopulmonary bypass. Microcirculation 2020, 27, e12608. [Google Scholar] [CrossRef]
- Shibuya, N.; Kimura, H. Production of hydrogen sulfide from d-cysteine and its therapeutic potential. Front. Endocrinol. 2013, 4, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muravyov, A.V.; Tikhomirova, I.A.; Avdonin, P.V.; Bulaeva, S.V.; Malisheva, J.V. Comparative efficiency of three gasotransmitters (nitric oxide, hydrogen sulfide and carbon monoxide): Analysis on the model of red blood cell microrheological responses. J. Cell. Biotechnol. 2021, 7, 1–9. [Google Scholar] [CrossRef]
- Wong, W.W.; Jack, R.M.; Smith, J.A.; Kennedy, C.A.; Fearon, D.T. Rapid purification of the human C3b/C4b receptor (CR1) by monoclonal antibody affinity chromatography. J. Immunol. Methods 1985, 82, 303–313. [Google Scholar] [CrossRef]
- Sung, L.A.; Kabat, E.A.; Chien, S. Interaction of lectins with membrane receptors on erythrocyte surfaces. J. Cell Biol. 1985, 101, 646–651. [Google Scholar] [CrossRef]
- Bryszewska, M.; Leyko, W. Effect of insulin on human erythrocyte membrane fluidity in diabetes mellitus. Diabetologia 1983, 24, 311–313. [Google Scholar] [CrossRef] [Green Version]
- Dutta-Roy, A.K.; Ray, T.K.; Sinha, A.K. Control of erythrocyte membrane microviscosity by insulin. Biochim. Biophys. Acta 1985, 816, 187–190. [Google Scholar] [CrossRef]
- Rahmani-Jourdheuil, D.; Mourayre, Y.; Vague, P.; Boyer, J.; Juhan-Vague, I. In vivo insulin effect on ATPase activities in erythrocyte membrane from insulin-dependent diabetics. Diabetes 1987, 36, 991–995. [Google Scholar] [CrossRef]
- Linde, T.; Sandhagen, B.; Berne, C.; Lind, L.; Reneland, R.; Hanni, A.; Lithell, H. Erythrocyte deformability is related to fasting insulin and declines during euglycaemic hyperinsulinaemia in hypertensive patients. J. Hum. Hypertens. 1999, 13, 285–286. [Google Scholar] [CrossRef] [Green Version]
- Aursnes, I.; Dahl-Jørgensen, K.; Hanssen, K.F. ATP-concentrations in erythrocytes influenced by insulin levels in plasma. Clin. Hemorheol. Microcirc. 1986, 12, 429–433. [Google Scholar] [CrossRef]
- Forst, T.; De La Tour, D.D.; Kunt, T.; Pfutzner, A.; Goitom, K.; Pohlmann, T.; Schneider, S.; Johansson, B.L.; Wahren, J.; Lobig, M.; et al. Effects of proinsulin C-peptide on nitric oxide, microvascular blood flow and erythrocyte Na+,K+-ATPase activity in diabetes mellitus type I. Clin. Sci. 2000, 98, 283–290. [Google Scholar] [CrossRef]
- Catanese, V.M.; Grigorescu, F.; King, G.L.; Kahn, C.R. The human erythrocyte insulin-like growth factor I receptor: Characterization and demonstration of ligand-stimulated autophosphorylation. J. Clin. Endocrinol. Metab. 1986, 62, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Monnier, J.F.; Benhaddad, A.A.; Micallef, J.P.; Mercier, J.; Brun, J.F. Relationships between blood viscosity and insulin-like growth factor I status in athletes. Clin. Hemorheol. Microcirc. 2000, 22, 277–286. [Google Scholar] [PubMed]
- Valensi, P.; Gaudey, F.; Attali, J.R. La déformabilité érythrocytaire est réduite par le glucagon. Diabetes Metab. 1986, 12, 281. [Google Scholar]
- Komatsu, R.; Tsushima, N.; Matsuyama, T. Effects of glucagon administration on microcirculation and blood rheology. Clin. Hemorheol. Microcirc. 1997, 17, 271–277. [Google Scholar]
- Schnyder, L.; Walter, R.; Rohrer, A.; Contesse, J.; Reinhart, W.H. No influence of C-peptide, insulin, and glucagon on blood viscosity in vitro in healthy humans and patients with diabetes mellitus. Clin. Hemorheol. Microcirc. 2001, 24, 65–74. [Google Scholar] [PubMed]
- Angel, R.C.; Botta, J.A.; Farias, R.N. High affinity L-triiodothyronine binding to right-side-out and inside-out vesicles from rat and human erythrocyte membrane. J. Biol. Chem. 1989, 264, 19143–19146. [Google Scholar] [CrossRef]
- Larsson, H.; Valdemarsson, S.; Hedner, P.; Odeberg, H. Blood viscosity in hyperthyroidism: Significance of erythrocyte changes and hematocrit. Clin. Hemorheol. Microcirc. 1988, 8, 257–265. [Google Scholar] [CrossRef]
- Zhang, Y.; Chua, S., Jr. Leptin Function and Regulation. Compr. Physiol. 2017, 8, 351–369. [Google Scholar] [CrossRef]
- Tsuda, K.; Kimura, K.; Nishio, I. Leptin improves membrane fluidity of erythrocytes in humans via a nitric oxide-dependent mechanism: An electron paramagnetic resonance investigation. Biochem. Biophys. Res. Commun. 2002, 297, 672–681. [Google Scholar] [CrossRef]
- Brun, J.F.; Perez-Martin, A.; Raynaud, E.; Mercier, J. Correlation between plasma viscosity, erythrocyte disaggregability and leptin. Biorheology 1999, 36, 156–157. [Google Scholar]
- Brun, J.F.; Varlet-Marie, E.; Vachoud, L.; Marion, B.; Roques, C.; Raynaud de Mauverger, E.; Mercier, J. Is leptin a regulator of erythrocyte rheology? Clin. Hemorheol. Microcirc. 2021, in press. [Google Scholar]
- Faulkner, J.L.; Belin de Chantemèle, E.J. Leptin and Aldosterone. Vitam. Horm. 2019, 109, 265–284. [Google Scholar] [CrossRef]
- Faulkner, J.L.; Bruder-Nascimento, T.; Belin de Chantemèle, E.J. The regulation of aldosterone secretion by leptin: Implications in obesity-related cardiovascular disease. Curr. Opin. Nephrol. Hypertens. 2018, 27, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Reinhart, W.H. Molecular biology and self-regulatory mechanisms of blood viscosity: A review. Biorheology 2001, 38, 203–212. [Google Scholar]
- Brun, J.F. Hormones, metabolism and body composition as major determinants of blood rheology: Potential pathophysiological meaning. Clin. Hemorheol. Microcirc. 2002, 26, 63–79. [Google Scholar]
- Muravyov, A.V.; Cheporov, S.V.; Kislov, N.V.; Bulaeva, S.V.; Maimistova, A.A. Comparative efficiency and hemorheological consequences of hemotransfusion and epoetin therapy in anemic cancer patients. Clin. Hemorheol. Microcirc. 2010, 44, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Vota, D.M.; Maltaneri, R.E.; Wenker, S.D.; Nesse, A.B.; Vittori, D.C. Differential erythropoietin action upon cells induced to eryptosis by different agents. Cell Biochem. Biophys. 2013, 65, 145–157. [Google Scholar] [CrossRef]
- Zyznar, E.S.; Pietri, A.O.; Harris, V.; Unger, R.H. Evidence for the hormonal status of somatostatin in man. Diabetes 1981, 30, 883–886. [Google Scholar] [CrossRef] [Green Version]
- Savi, L.; Cardillo, C.; Bombardieri, G. Somatostatin and peripheral blood flow in man. Angiology 1985, 36, 511–515. [Google Scholar] [CrossRef]
- Fuse, I.; Ito, S.; Takagi, A.; Shibata, A. Different effects of three kinds of somatostatin (15–28, 1–14, 1–28) on rabbit’s platelet aggregation. Life Sci. 1985, 36, 2047–2052. [Google Scholar] [CrossRef]
- Brun, J.F.; Rauturier, M.; Ghanem, Y.; Orsetti, A. In vitro effects of somatostatin on red cell filterability measured by three methods. J. Mal. Vasc. 1991, 16, 49–52. [Google Scholar]
- Berker, M.; Dikmenoglu, N.; Bozkurt, G.; Ergönül, Z.; Özgen, T. Hemorheology, melatonin and pinealectomy. What’s the relationship? An experimental study. Clin. Hemorheol. Microcirc. 2004, 30, 47–52. [Google Scholar]
- Vazan, R.; Plauterova, K.; Porubska, G.; Radosinska, J. Changes in erythrocyte deformability during day and possible role of melatonin. Endocr. Regul. 2018, 52, 17–20. [Google Scholar] [CrossRef] [Green Version]
- Freyburger, G.; Larrue, J.; Boisseau, M.R. Effect of two arachidonic acid metabolites of white blood cells on red blood cell filterability studied by cell transit-time analysis. Clin. Hemorheol. Microcirc. 1987, 7, 536. [Google Scholar]
- Mary, A.; Modat, G.; Gal, L.; Bonne, C. Leukotriene B4 decreases RBC deformability as assessed by increased filtration index. Clin. Hemorheol. Microcirc. 1989, 9, 209–217. [Google Scholar] [CrossRef]
- Elgatit, A.M.; Rashid, M.A.; Belboul, A.M.; Ramirez, J.J.; Roberts, D.G.; Olsson, G.W. Effects of aprostadil (PGE1) on red and white cell deformability during cardiopulmonary bypass. In Proceedings of the VIIth European Conference on Clinical Haemorheology, Southampton, UK, 16–19 July 1991; Abstract book. p. 81. [Google Scholar]
- Langer, R.; Rossmanith, K.; Henrich, H.A. Hemorheological actions of the prostaglandins D2, E1, E2, F1a, F2a and Iloprost. Clin. Hemorheol. Microcirc. 1995, 15, 829–839. [Google Scholar] [CrossRef]
- Brun, J.F.; Bouchahda, C.; Aissa-Benhaddad, A.; Sagnes, C.; Granat, M.C.; Bor Kucukatay, M.; Baskurt, O.; Mercier, J. Hemorheological aspects of leuko-platelet activation in atheromatous diseases: Clinical applications. J. Mal. Vasc. 2000, 25, 349–355. [Google Scholar]
- Gonçalves, I.; Saldanha, C.; Martins, E.; Silva, J. Beta-estradiol effect on erythrocyte aggregation—A controlled in vitro study. Clin. Hemorheol. Microcirc. 2001, 25, 127–134. [Google Scholar]
- Grau, M.; Cremer, J.M.; Bloch, W. Comparisons of blood parameters, red blood cell deformability and circulating nitric oxide between males and females considering hormonal contraception: A longitudinal gender study. Front. Physiol. 2018, 9, 1835. [Google Scholar] [CrossRef]
- Swanepoel, A.C.; Emmerson, O.; Pretorius, E. Effect of Progesterone and Synthetic Progestins on Whole Blood Clot Formation and Erythrocyte Structure. Microsc. Microanal. 2017, 23, 607–617. [Google Scholar] [CrossRef]
- Swanepoel, A.C.; Pretorius, E. Erythrocyte-platelet interaction in uncomplicated pregnancy. Microsc. Microanal. 2014, 20, 1848–1860. [Google Scholar] [CrossRef] [Green Version]
- Kon, K.; Maeda, N.; Shiga, T. Functional impairments of human red cells, induced by dehydroepiandrosterone sulfate. Pflugers Arch. 1982, 394, 279–286. [Google Scholar] [CrossRef]
- Kartal, H.; Comu, F.M.; Kucuk, A.; Polat, Y.; Dursun, A.D.; Arslan, M. Effect of apelin-13 on erythrocyte deformability during ischaemia-reperfusion injury of heart in diabetic rats. Bratisl Lek Listy 2017, 118, 133–136. [Google Scholar] [CrossRef]
- Sundquist, J.; Blas, S.D.; Hogan, J.E.; Davis, F.B.; Davis, P.J. The alpha 1-adrenergic receptor in human erythrocyte membranes mediates interaction in vitro of epinephrine and thyroid hormone at the membrane Ca(2+)-ATPase. Cell. Signal. 1992, 4, 795–799. [Google Scholar] [CrossRef]
- Borgese, F.; Garcia-Romeu, F.; Motais, R. Control of cell volume and ion transport by beta-adrenergic catecholamines in erythrocytes of rainbow trout, Salmo gairdneri. J. Physiol. 1987, 382, 123–144. [Google Scholar] [CrossRef] [PubMed]
- Pfafferott, C.; Zaninelli, R.; Bauersachs, R.; Volger, E. In vitro effect of norepinephrine and isoprenaline on erythrocyte filterability under pathologic conditions. Clin. Hemorheol. Microcirc. 1987, 7, 409. [Google Scholar]
- Hilario, S.; Saldanha, C.; Martins-Silva, J. The effect of adrenaline upon human erythrocyte. Sex-related differences? Biorheology 1999, 36, 124. [Google Scholar]
- Chiocchia, G.; Motais, R. Effect of catecholamines on deformability of red cells from trout: Relative roles of cyclic AMP and cell volume. J. Physiol. 1989, 412, 321–332. [Google Scholar] [CrossRef] [Green Version]
- Hilario, S.; Saldanha, C.; Martins-Silva, J. An in vitro study of adrenaline effect on human erythrocyte properties in both genders. Clin. Hemorheol. Microcirc. 2003, 28, 89–98. [Google Scholar] [PubMed]
- Oonishi, T.; Sakashita, K.; Uyesaka, N. Regulation of red blood cell filterability by Ca2+ influx and cAMP-mediated signaling pathways. Am. J. Physiol. 1997, 273, C1828–C1834. [Google Scholar] [CrossRef] [PubMed]
- Tikhomiroiva, I.A.; Muravyov, A.V.; Kruglova, E.V. Hormonal control mechanisms of the rheological properties of erythrocytes under physical exercises and stress. Biorheology 2008, 45, 49. [Google Scholar]
- Muravyov, A.V.; Tikhomirova, I.A.; Maimistova, A.A.; Bulaeva, S.V. Extra- and intracellular signaling pathways under red blood cell aggregation and deformability changes. Clin. Hemorheol. Microcirc. 2009, 43, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Berent, H.; Wocial, B.; Kuczyńska, K.; Kochmański, M.; Ignatowska-Switalska, H.; Januszewicz, A.; Lapiński, M.; Lewandowski, J.; Januszewicz, W. Evaluation of blood rheology indices in patients with pheochromocytoma. Pol. Arch. Med. Wewn. 1996, 95, 190–197. [Google Scholar] [PubMed]
- Mokrushnikov, P.V.; Panin, L.E.; Zaitsev, B.N. The action of stress hormones on the structure and function of erythrocyte membrane. Gen. Physiol. Biophys. 2015, 34, 311–321. [Google Scholar] [CrossRef]
- Windberger, U.; Bartholovitsch, A. Hemorheology in spontaneous animal endocrinopathies. Clin. Hemorheol. Microcirc. 2004, 31, 207–215. [Google Scholar] [PubMed]
- Lowe, H.; Toyang, N.; Steele, B.; Bryant, J.; Ngwa, W. The endocannabinoid system: A potential target for the treatment of various diseases. Int. J. Mol. Sci. 2021, 22, 9472. [Google Scholar] [CrossRef]
- Piazza, P.V.; Cota, D.; Marsicano, G. The CB1 receptor as the cornerstone of exostasis. Neuron 2017, 93, 1252–1274. [Google Scholar] [CrossRef]
- Cavuoto, P.; Wittert, G.A. The role of the endocannabinoid system in the regulation of energy expenditure. Best. Pract. Res. Clin. Endocrinol. Metab. 2009, 23, 79–86. [Google Scholar] [CrossRef]
- Sakashita, K.; Oonishi, T.; Ishioka, N.; Uyesaka, N. Endothelin-1 improves the impaired filterability of red blood cells through the activation of protein kinase C. Jpn. J. Physiol. 1999, 49, 113–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, R.; Mark, M.; Gaudenz, R.; Harris, L.G.; Reinhart, W.H. Influence of nitrovasodilators and endothelin-1 on rheology of human blood in vitro. Br. J. Pharmacol. 1999, 128, 744–750. [Google Scholar] [CrossRef] [Green Version]
- Mark, M.; Walter, R.; Harris, L.G.; Reinhart, W.H. Influence of parathyroid hormone, calcitonin, 1,25(OH)2 cholecalciferol, calcium, and the calcium ionophore A23187 on erythrocyte morphology and blood viscosity. J. Lab. Clin. Med. 2000, 135, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Bester, J.; Pretorius, E. Effects of IL-1β, IL-6 and IL-8 on erythrocytes, platelets and clot viscoelasticity. Sci. Rep. 2016, 6, 32188. [Google Scholar] [CrossRef]
- Olumuyiwa-Akeredolu, O.O.; Pretorius, E. Platelet and red blood cell interactions and their role in rheumatoid arthritis. Rheumatol. Int. 2015, 35, 1955–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, M.J.; Bester, J.; Pretorius, E. Interleukin-12 and its procoagulant effect on erythrocytes, platelets and fibrin(nogen): The lesser-known side of inflammation. Brit. J. Hematol. 2018, 180, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Page, M.J.; Bester, J.; Pretorius, E. The inflammatory effects of TNF-alpha and complement component 3 on coagulation. Sci. Rep. 2018, 8, 1812. [Google Scholar] [CrossRef] [Green Version]
- Dhabhar, F.S. The short-term stress response—Mother nature’s mechanism for enhancing protection and performance under conditions of threat, challenge, and opportunity. Front. Neuroendocrinol. 2018, 49, 175–192. [Google Scholar] [CrossRef] [PubMed]
- Finnerty, F.A.; Buchholz, J.H.; Guillaudeu, R.L. The blood volumes and plasma protein during levarterenol-induced hypertension. J. Clin. Invest. 1958, 37, 425–429. [Google Scholar] [CrossRef] [Green Version]
- Ehrly, A.M.; Landgraf, H.; Hessler, J.; Saeger-Lorenz, K. Influence of videofilm-induced emotional stress on the flow properties of blood. Angiology 1988, 39, 341–344. [Google Scholar] [CrossRef]
- Sakai, A.; Nakano, H.; Ohira, T.; Hosoya, M.; Yasumura, S.; Ohtsuru, A.; Satoh, H.; Kawasaki, Y.; Suzuki, H.; Takahashi, A.; et al. Fukushima Health Management Survey Group. Persistent prevalence of polycythemia among evacuees 4 years after the Great East Japan Earthquake: A follow-up study. Prev. Med. Rep. 2017, 5, 251–256. [Google Scholar] [CrossRef]
- Falvo, M.J.; Chen, Y.; Klein, J.C.; Ndirangu, D.; Condon, M.R. Abnormal rheological properties of red blood cells as a potential marker of Gulf War Illness: A preliminary study. Clin. Hemorheol. Microcirc. 2018, 68, 361–370. [Google Scholar] [CrossRef]
- Qian, W.; Klein-Adams, J.C.; Ndirangu, D.S.; Chen, Y.; Falvo, M.J.; Condon, M.R. Hemorheological responses to an acute bout of maximal exercise in Veterans with Gulf War Illness. Life Sci. 2021, 280, 119714. [Google Scholar] [CrossRef] [PubMed]
- Brun, J.F.; Varlet-Marie, E.; Richou, M.; Mercier, J.; Raynaud de Mauverger, E. Blood rheology as a mirror of endocrine and metabolic homeostasis in health and disease. Clin. Hemorheol. Microcirc. 2018, 69, 239–265. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.K.; Schmidt, B.R.; Wilhelmy, J.; Nguyen, V.; Abugherir, A.; Do, J.K.; Nemat-Gorgani, M.; Davis, R.W.; Ramasubramanian, A.K. Red blood cell deformability is diminished in patients with Chronic Fatigue Syndrome. Clin. Hemorheol. Microcirc. 2019, 71, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Brenu, E.W.; Staines, D.R.; Baskurt, O.K.; Ashton, K.J.; Ramos, S.B.; Christy, R.M.; Marshall-Gradisnikal, S.M. Immune and hemorheological changes in chronic fatigue syndrome. J. Transl. Med. 2010, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, G.; Spence, V.A.; McLaren, M.; Hill, A.; Underwood, C.; Belch, J.J. Oxidative stress levels are raised in chronic fatigue syndrome and are associated with clinical symptoms. Free Radic. Biol. Med. 2005, 39, 584–589. [Google Scholar] [CrossRef]
- Kennedy, G.; Khan, F.; Hill, A.; Underwood, C.; Belch, J.J. Biochemical and vascular aspects of pediatric chronic fatigue syndrome. Arch. Pediatr. Adolesc. Med. 2010, 164, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Choi, S.U.; Youn, J.M.; Cha, S.H.; Shin, H.J.; Ko, E.J.; Lim, C.H. Effects of remote ischemic preconditioning on the deformability and aggregation of red blood cells in a rat endotoxemia model. Clin. Hemorheol. Microcirc. 2021. Online ahead of print. [Google Scholar] [CrossRef]
- Ko, E.; Youn, J.M.; Park, H.S.; Song, M.; Koh, K.H.; Lim, C.H. Early red blood cell abnormalities as a clinical variable in sepsis diagnosis. Clin. Hemorheol. Microcirc. 2018, 70, 355–363. [Google Scholar] [CrossRef]
- Piagnerelli, M.; Boudjeltia, K.Z.; Vanhaeverbeek, M.; Vincent, J.L. Red blood cell rheology in sepsis. Intensive Care Med. 2003, 29, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Donadello, K.; Piagnerelli, M.; Reggiori, G.; Gottin, L.; Scolletta, S.; Occhipinti, G.; Zouaoui Boudjeltia, K.; Vincent, J.L. Reduced red blood cell deformability over time is associated with a poor outcome in septic patients. Microvasc. Res. 2015, 101, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Totsimon, K.; Biro, K.; Szabo, Z.E.; Toth, K.; Kenyeres, P.; Marton, Z. The relationship between hemorheological parameters and mortality in critically ill patients with and without sepsis. Clin. Hemorheol. Microcirc. 2017, 65, 119–129. [Google Scholar] [CrossRef]
- Bateman, R.M.; Jagger, J.E.; Sharpe, M.D.; Ellsworth, M.L.; Mehta, S.; Ellis, C.G. Erythrocyte deformability is a nitric oxide-mediated factor in decreased capillary density during sepsis. Am. J. Physiol. Heart Circ. Physiol. 2001, 280, H2848–H2856. [Google Scholar] [CrossRef]
- Bateman, R.M.; Sharpe, M.D.; Singer, M.; Ellis, C.G. The Effect of Sepsis on the Erythrocyte. Int. J. Mol. Sci. 2017, 18, 1932. [Google Scholar] [CrossRef] [Green Version]
- Poraicu, D.; Mogoseanu, A.; Tomescu, N.; Bota, C.; Menessy, I. Decrease of red blood cell filterability seen in intensive care I. The correlation of low erythrocyte filterability with mortality and its return to normal values in critically ill patients under parenteral nutrition. Resuscitation 1983, 10, 291–303. [Google Scholar] [CrossRef]
- Silva-Herdade, A.S.; Andolina, G.; Faggio, C.; Calado, A.; Saldanha, C. Erythrocyte deformability - A partner of the inflammatory response. Microvasc. Res. 2016, 107, 34–38. [Google Scholar] [CrossRef]
- Korbut, R.; Gryglewski, R.J. The effect of prostacyclin and nitric oxide on deformability of red blood cells in septic shock in rats. J. Physiol. Pharmacol. 1996, 47, 591–599. [Google Scholar]
- Caimi, G.; Montana, M.; Canino, B.; Calandrino, V.; Lo Presti, R.; Hopps, E. Erythrocyte deformability, plasma lipid peroxidation and plasma protein oxidation in a group of OSAS subjects. Clin. Hemorheol. Microcirc. 2016, 64, 7–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canino, B.; Hopps, E.; Calandrino, V.; Montana, M.; Lo Presti, R.; Caimi, G. Nitric oxide metabolites and erythrocyte deformability in a group of subjects with obstructive sleep apnea syndrome. Clin. Hemorheol. Microcirc. 2015, 59, 45–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentim-Coelho, C.; Vaz, F.; Antunes, M.; Neves, S.; Martins, I.L.; Osório, H.; Feliciano, A.; Pinto, P.; Bárbara, C.; Penque, D. Redox-Oligomeric State of Peroxiredoxin-2 and Glyceraldehyde-3-Phosphate Dehydrogenase in Obstructive Sleep Apnea Red Blood Cells under Positive Airway Pressure Therapy. Antioxidants 2020, 9, 1184. [Google Scholar] [CrossRef] [PubMed]
- Buono, M.; Rostomily, K. Acute normobaric hypoxia does not increase blood or plasma viscosity. Clin. Hemorheol. Microcirc. 2021, 78, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Jung, F. COVID-19. Clin. Hemorheol. Microcirc. 2020, 74, 347–348. [Google Scholar] [CrossRef] [Green Version]
- Jung, F.; Krüger-Genge, A.; Franke, R.P.; Hufert, F.; Küpper, J.-H. COVID-19 and the endothelium. Clin. Hemorheol. Microcirc. 2020, 75, 7–11. [Google Scholar] [CrossRef]
- Farber, P.L. Can erythrocytes behavior in microcirculation help the understanding the physiopathology and improve prevention and treatment for covid-19? Clin. Hemorheol. Microcirc. 2021, 78, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A.; et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in Covid-19. N. Engl. J. Med. 2020, 383, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Goshua, G.; Pine, A.B.; Meizlish, M.L.; Chang, C.H.; Zhang, H.; Bahel, P.; Baluha, A.; Bar, N.; Bona, R.D.; Burns, A.J.; et al. Endotheliopathy in COVID-19-associated coagulopathy: Evidence from a single-centre, cross-sectional study. Lancet Haematol. 2020, 7, e575–e582. [Google Scholar] [CrossRef]
- Grobler, C.; Maphumulo, S.C.; Grobbelaar, L.M.; Bredenkamp, J.C.; Laubscher, G.J.; Lourens, P.J.; Steenkamp, J.; Kell, D.B.; Pretorius, E. Covid-19: The Rollercoaster of Fibrin(Ogen), D-Dimer, Von Willebrand Factor, P-Selectin and Their Interactions with Endothelial Cells, Platelets and Erythrocytes. Int. J. Mol. Sci. 2020, 21, 5168. [Google Scholar] [CrossRef]
- Pretorius, E.; Venter, C.; Laubscher, G.J.; Lourens, P.J.; Steenkamp, J.; Kell, D.B. Prevalence of readily detected amyloid blood clots in ‘unclotted’ Type 2 Diabetes Mellitus and COVID-19 plasma: A preliminary report. Cardiovasc. Diabetol. 2020, 19, 193. [Google Scholar] [CrossRef] [PubMed]
- Roberts, I.; Muelas, M.W.; Taylor, J.M.; Davison, A.S.; Xu, Y.; Grixti, J.M.; Gotts, N.; Sorokin, A.; Goodacre, R.; Kell, D.B. Untargeted metabolomics of COVID-19 patient serum reveals potential prognostic markers of both severity and outcome. MedRxiv 2020. [Google Scholar] [CrossRef]
- Venter, C.; Bezuidenhout, J.A.; Laubscher, G.J.; Lourens, P.J.; Steenkamp, J.; Kell, D.B.; Pretorius, E. Erythrocyte, Platelet, Serum Ferritin, and P-Selectin Pathophysiology Implicated in Severe Hypercoagulation and Vascular Complications in COVID-19. Int. J. Mol. Sci. 2020, 21, 8234. [Google Scholar] [CrossRef] [PubMed]
- Bobrova, L.; Kozlovskaya, N.; Korotchaeva, Y.; Bobkova, I.; Kamyshova, E.; Moiseev, S. Microvascular COVID-19 lung vessels obstructive thromboinflammatory syndrome (MicroCLOTS): A new variant of thrombotic microangiopathy? Crit. Care Resusc. 2020, 22, 284. [Google Scholar] [PubMed]
- Ciceri, F.; Beretta, L.; Scandroglio, A.M.; Colombo, S.; Landoni, G.; Ruggeri, A.; Peccatori, J.; D’angelo, A.; De Cobelli, F.; Rovere-Querini, P.; et al. Microvascular COVID-19 lung vessels obstructive thromboinflammatory syndrome (MicroCLOTS): An atypical acute respiratory distress syndrome working hypothesis. Crit. Care Resusc. 2020, 22, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Renzi, S.; Landoni, G.; Zangrillo, A.; Ciceri, F. MicroCLOTS pathophysiology in COVID 19. Korean J. Intern. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Akhter, N.; Ahmad, S.; Alzahrani, F.A.; Dar, S.A.; Wahid, M.; Haque, S.; Bhatia, K.; Sr Almalki, S.; Alharbi, R.A.; Sindi, A.A. Impact of COVID-19 on the cerebrovascular system and the prevention of RBC lysis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 10267–10278. [Google Scholar] [CrossRef]
- Berzuini, A.; Bianco, C.; Paccapelo, C.; Bertolini, F.; Gregato, G.; Cattaneo, A.; Erba, E.; Bandera, A.; Gori, A.; Lamorte, G.; et al. Red cell-bound antibodies and transfusion requirements in hospitalized patients with COVID-19. Blood 2020, 136, 766–768. [Google Scholar] [CrossRef]
- Lam, L.M.; Murphy, S.J.; Kuri-Cervantes, L.; Weisman, A.R.; Ittner, C.A.; Reilly, J.P.; Pampena, M.B.; Betts, M.R.; Wherry, E.J.; Song, W.C.; et al. Erythrocytes Reveal Complement Activation in Patients with COVID-19. medRxiv 2020. [Google Scholar] [CrossRef]
- Pretorius, E.; Vlok, M.; Venter, C.; Bezuidenhout, J.A.; Laubscher, G.J.; Steenkamp, J.; Kell, D.B. Persistent clotting protein pathology in Long COVID/Post-Acute Sequelae of COVID-19 (PASC) is accompanied by increased levels of antiplasmin. Cardiovasc. Diabetol. 2021, 20, 172. [Google Scholar] [CrossRef] [PubMed]
- Grobbelaar, L.M.; Venter, C.; Vlok, M.; Ngoepe, M.; Laubscher, G.J.; Lourens, P.J.; Steenkamp, J.; Kell, D.B.; Pretorius, E. Sars-Cov-2 Spike Protein S1 Induces Fibrin(Ogen) Resistant To Fibrinolysis: Implications For Microclot Formation In Covid-19. Biosci. Rep. 2021, 41, BSR20210611. [Google Scholar] [CrossRef]
- Başkurt, O.K.; Edremitlioğlu, M.; Temiz, A. In Vitro Effects of in Vivo Activated Leukocytes on RBC Filterability and Lipid Peroxidation. Clin. Hemorheol. Microcirc. 1994, 14, 591–596. [Google Scholar] [CrossRef]
- Nash, G.B.; Watts, T.; Thornton, C.; Barigou, M. Red cell aggregation as a factor influencing margination and adhesion of leukocytes and platelets. Clin. Hemorheol. Microcirc. 2008, 39, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Tigno, X.T.; Selaru, I.K.; Angeloni, S.V.; Hansen, B.C. Is microvascular flow rate related to ghrelin, leptin and adiponectin levels? Clin. Hemorheol. Microcirc. 2003, 29, 409–416. [Google Scholar] [PubMed]
- Piecuch, J.; Mertas, A.; Nowowiejska–Wiewiora, A.; Sławomir, G.; Zenon, C.; Wiewiora, M. The relationship between the rheological behavior of RBCs and angiogenesis in the morbidly obese. Clin. Hemorheol. Microcirc. 2019, 71, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Murohara, T.; Asahara, T. Nitric oxide and angiogenesis in cardiovascular disease. Antioxid Redox Signal. 2002, 4, 825–831. [Google Scholar] [CrossRef]
- Kondo, T.; Kobayashi, K.; Murohara, T. Nitric oxide signaling during myocardial angiogenesis. Mol. Cell. Biochem. 2004, 264, 25–34. [Google Scholar] [CrossRef]
- Gaudard, A.; Varlet-Marie, E.; Bressolle, F.; Mercier, J.; Brun, J.F. Nutrition as a determinant of blood rheology and fibrinogen in athletes. Clin. Hemorheol. Microcirc. 2004, 30, 1–8. [Google Scholar]
- Brun, J.F.; Varlet-Marie, E.; Romain, A.J.; Guiraudou, M.; Raynaud De Mauverger, E. Exercise hemorheology: Moving from old simplistic paradigms to a more complex picture. Clin. Hemorheol. Microcirc. 2013, 55, 15–27. [Google Scholar] [CrossRef]
- Brun, J.F.; Varlet-Marie, E.; Chevance, G.; Pollatz, M.; Fedou, C.; Raynaud de Mauverger, E. Versatility of ‘hemorheologic fitness’ according to exercise intensity: Emphasis on the “healthy primitive lifestyle”. Korea-Aust. Rheol. J. 2014, 26, 249–253. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Increases RBC Deformability | Decreases RBC Deformability | Increases Eryptosis (After [24] and [20]) | Decreases Eryptosis (After [24] and [20]) | |
|---|---|---|---|---|
| Biologically active molecules and metabolites | ATP NO H2S Carbon monoxide Zn++ Lactate (in trained athletes) | Ketone bodies Cholesterol Glucose > 200 dg/mL Lactate (in sedentary subjects) | Aluminium Arsenic Cadmium Carbon monoxide Ceramide (acylsphingosine) Chromium Copper Fe2+, Energy depletion Glucose (via glycation) Osmotic shock Zn++ | NO Erythropoietin Catecholamines β and α |
| Hormones and chemical messengers | Acetylcholine Epinephrine Endothelin 1 Apelin Leptin Progesterone Erythropoietin Somatostatin Prostaglandin E1 DHEA | Glucagon Melatonin ADP PGE2 Norepinephrine (?) Leukotriene B4 Thyroxin IGF-I Estradiol | Anandamide Estradiol Leukotriene C(4) Lithium Lysophosphatidic acid Mercury PAF Phosphate Progesterone Prostaglandin E2 Silver ions Sphingosine | Adenosine Chloride Erythropoietin Nitroprusside (NO-donor) Urea |
| Nutritional factors | Tea catechins Vitamin E α-tocopherol, α tocoterol | Carbohydrate intake | Curcurmin Gossypol Oxysterol Phytic acid Retinoic acid Retinoic acid Selenium (sodium selenite) Tannic acid Vitamin K | Caffeine Glutathione Monohydroxyethylrutoside N-acetylcysteine Naringin Vitamin E |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brun, J.-F.; Varlet-Marie, E.; Myzia, J.; Raynaud de Mauverger, E.; Pretorius, E. Metabolic Influences Modulating Erythrocyte Deformability and Eryptosis. Metabolites 2022, 12, 4. https://doi.org/10.3390/metabo12010004
Brun J-F, Varlet-Marie E, Myzia J, Raynaud de Mauverger E, Pretorius E. Metabolic Influences Modulating Erythrocyte Deformability and Eryptosis. Metabolites. 2022; 12(1):4. https://doi.org/10.3390/metabo12010004
Chicago/Turabian StyleBrun, Jean-Frédéric, Emmanuelle Varlet-Marie, Justine Myzia, Eric Raynaud de Mauverger, and Etheresia Pretorius. 2022. "Metabolic Influences Modulating Erythrocyte Deformability and Eryptosis" Metabolites 12, no. 1: 4. https://doi.org/10.3390/metabo12010004