
Agronomic and Metabolomic Side-Effects of a Divergent Selection for Indol-3-Ylmethylglucosinolate Content in Kale (Brassica oleracea var. acephala)

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Results
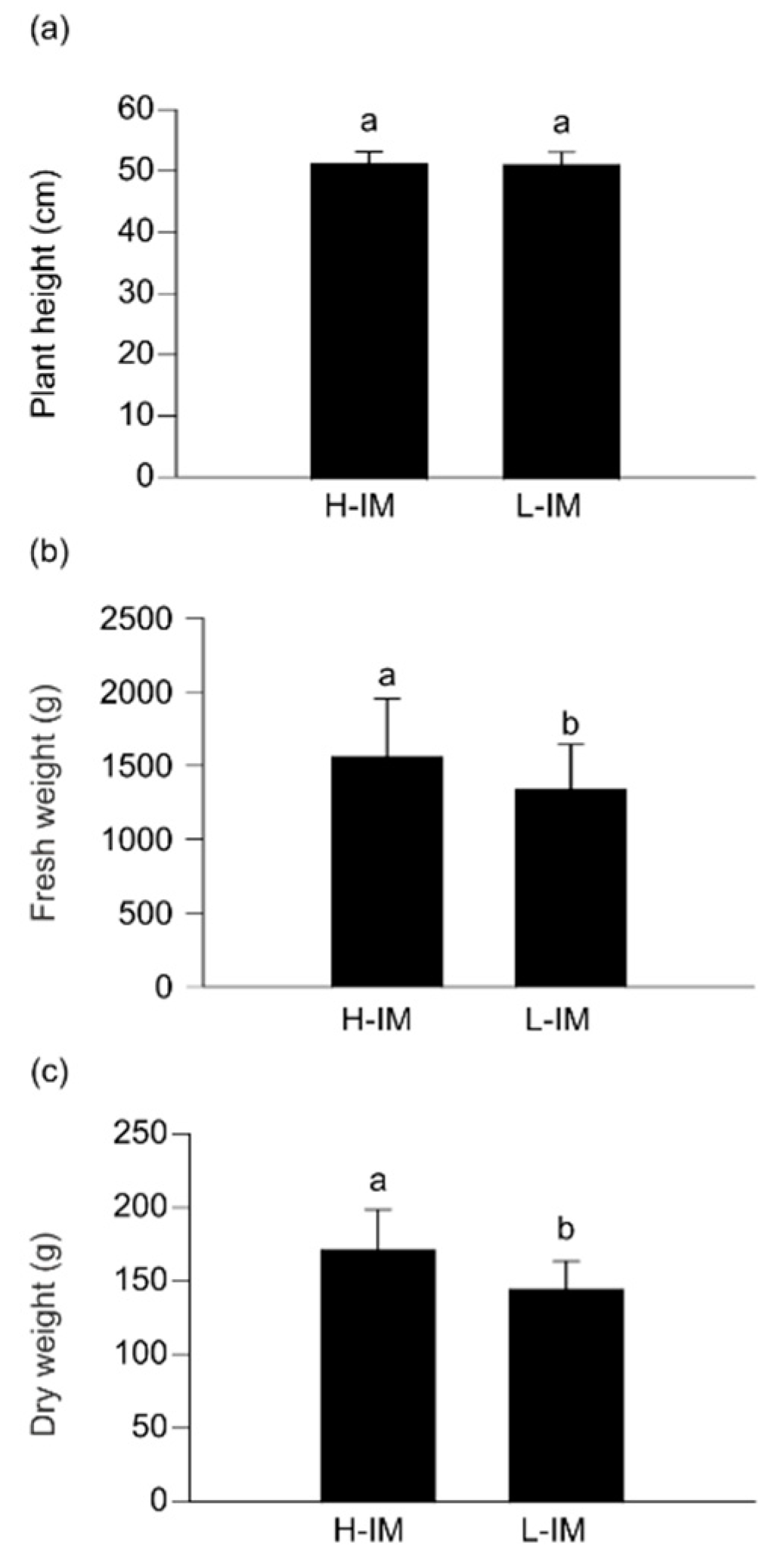
2.1. Differences in Agronomic Parameters
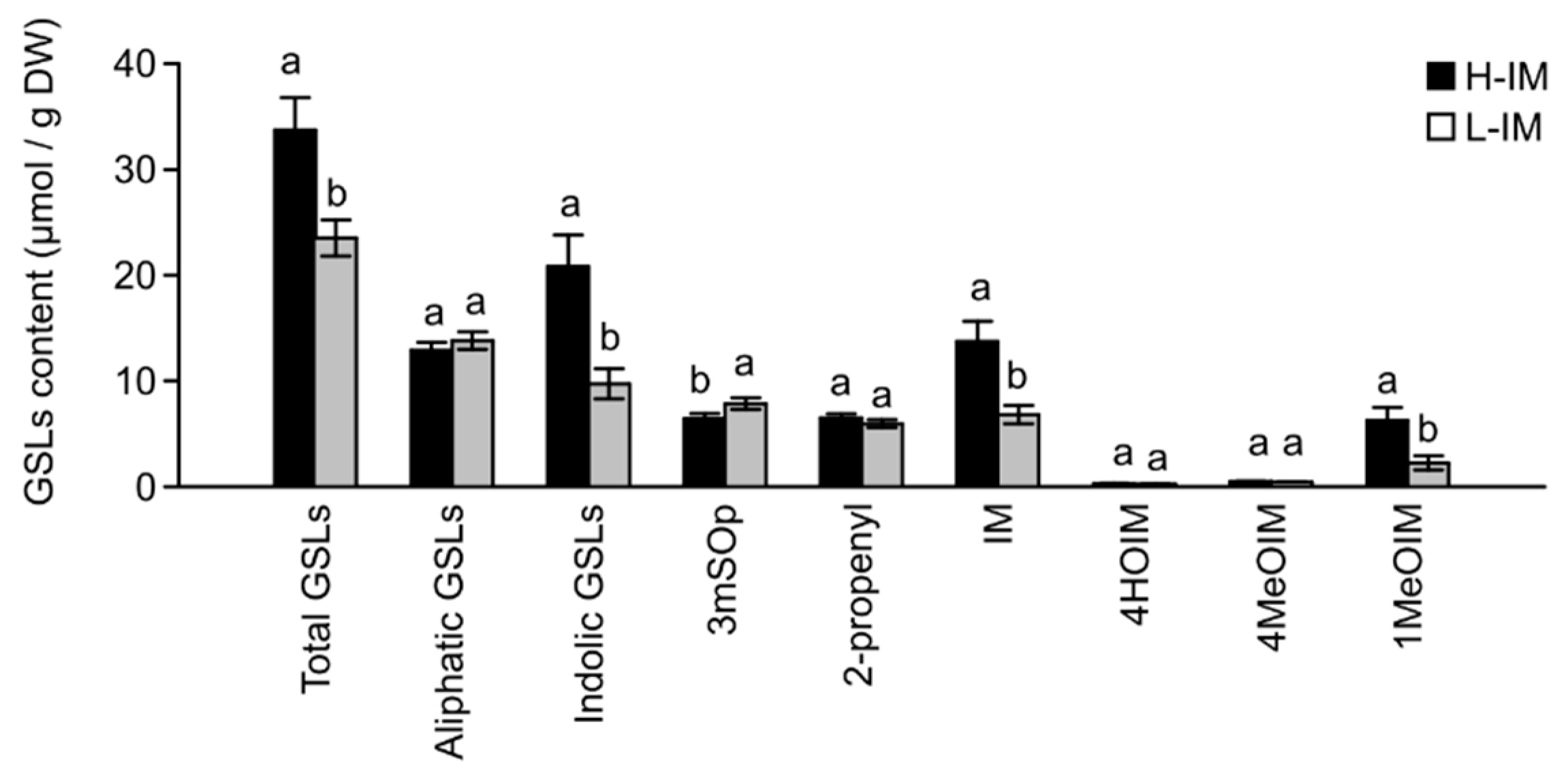
2.2. GSL Profiles
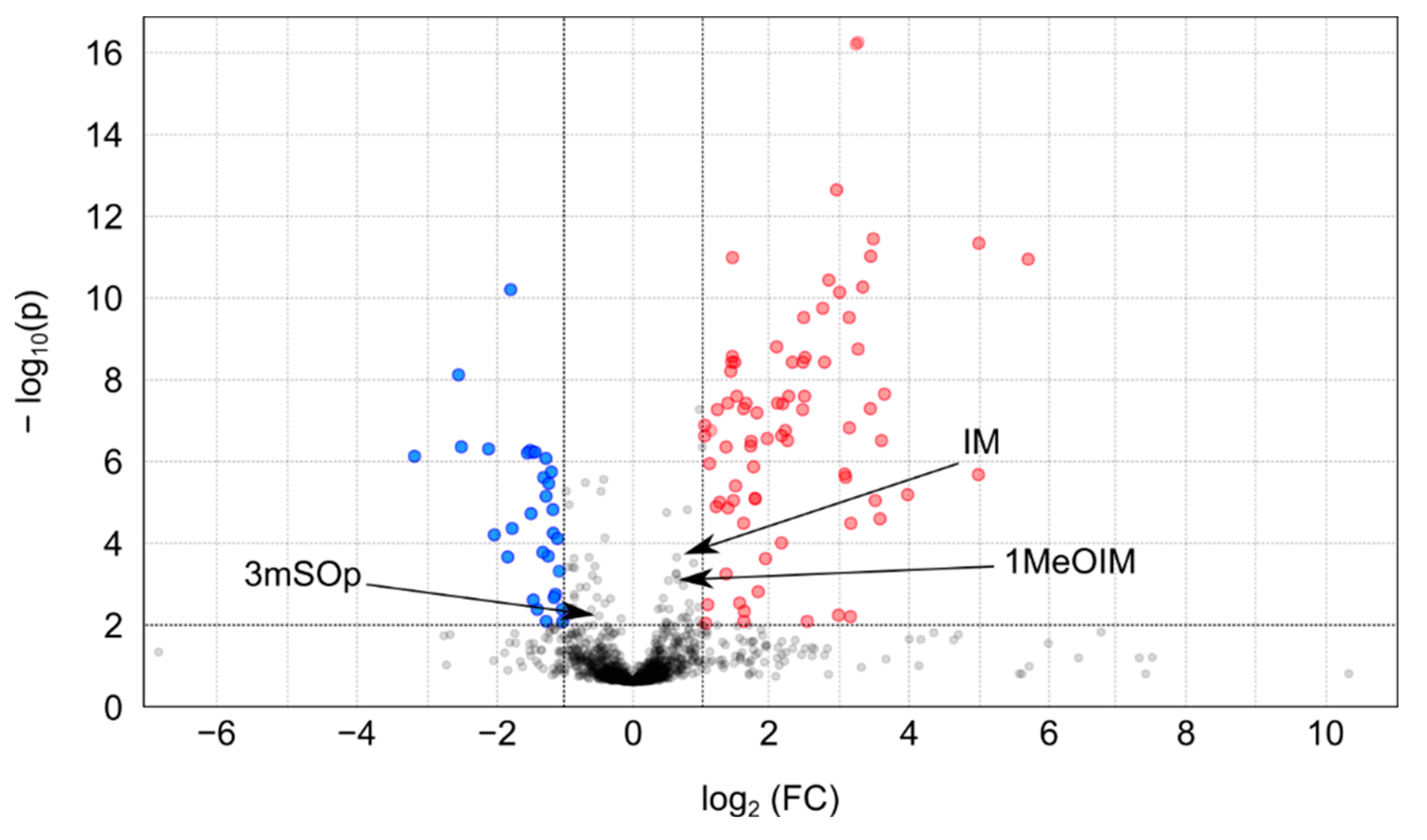
2.3. Metabolomic Profiles
3. Discussion
4. Materials and Methods
4.1. Plant Populations
4.2. Growing Conditions and Locations
4.3. Agronomic Parameters
4.4. Biochemical Analysis
4.4.1. GSLs Analysis
4.4.2. Metabolomic Analysis
4.4.3. Statistical Analysis
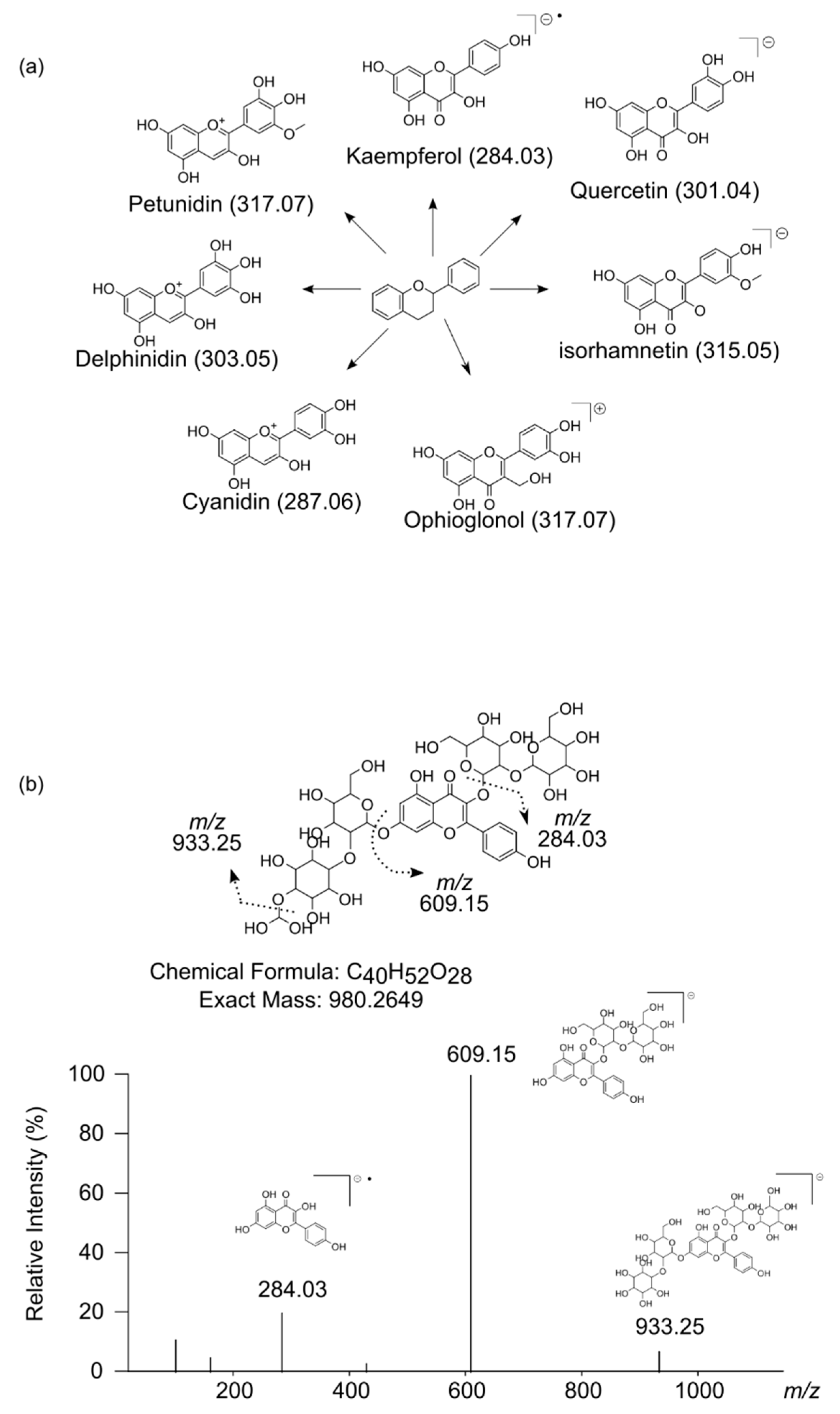
4.4.4. Tentative Metabolite Identification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Francisco, M.; Tortosa, M.; Martínez-Ballesta, M.D.; Velasco, P.; García-Viguera, C.; Moreno, D.A. Nutritional and phytochemical value of Brassica crops from the agri-food perspective. Ann. Appl. Biol. 2017, 170, 273–285. [Google Scholar] [CrossRef]
- Šamec, D.; Urlić, B.; Salopek-Sondi, B. Kale (Brassica oleracea var. acephala) as a superfood: Review of the scientific evidence behind the statement. Crit. Rev. Food Sci. Nutr. 2019, 59, 2411–2422. [Google Scholar]
- Cartea, M.E.; Velasco, P. Glucosinolates in Brassica foods: Bioavailability in food and significance for human health. Phytochem. Rev. 2008, 7, 213–229. [Google Scholar] [CrossRef]
- Poveda, J. Glucosinolates profile of Arabidopsis thaliana modified root colonization of Trichoderma species. Biol. Control 2021, 155, 104522. [Google Scholar] [CrossRef]
- Falk, K.L.; Kästner, J.; Bodenhausen, N.; Schramm, K.; Paetz, C.; Vassão, D.G.; Reichelt, M.; von Knorre, D.; Bergelson, J.; Erb, M.; et al. The role of glucosinolates and the jasmonic acid pathway in resistance of Arabidopsis thaliana against molluscan herbivores. Mol. Ecol. 2014, 23, 1188–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saladino, F.; Bordin, K.; Luciano, F.B.; Franzón, M.F.; Mañes, J.; Meca, G. Antimicrobial Activity of the Glucosinolates BT-Glucosinolates; Mérillon, J.-M., Ramawat, K.G., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 249–274. ISBN 978-3-319-25462-3. [Google Scholar]
- Poveda, J.; Eugui, D.; Velasco, P. Natural control of plant pathogens through glucosinolates: An effective strategy against fungi and oomycetes. Phytochem. Rev. 2020, 19, 1045–1059. [Google Scholar] [CrossRef]
- Jeschke, V.; Kearney, E.E.; Schramm, K.; Kunert, G.; Shekhov, A.; Gershenzon, J.; Vassão, D.G. How Glucosinolates affect generalist lepidopteran larvae: Growth, development and glucosinolate metabolism. Front. Plant Sci. 2017, 8, 1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knauer, A.C.; Schiestl, F.P. The effect of pollinators and herbivores on selection for floral signals: A case study in Brassica rapa. Evol. Ecol. 2017, 31, 285–304. [Google Scholar] [CrossRef]
- Bradburne, R.P.; Mithen, R. Glucosinolate genetics and the attraction of the aphid parasitoid Diaeretiella rapae to Brassica. Proc. Biol. Sci. 2000, 267, 89–95. [Google Scholar] [CrossRef]
- Van Loon, J.J.A.; Schoonhoven, L.M. Specialist deterrent chemoreceptors enable Pieris caterpillars to discriminate between chemically different deterrents. Entomol. Exp. Appl. 1999, 91, 29–35. [Google Scholar] [CrossRef]
- Jensen, L.; Jepsen, H.; Halkier, B.; Kliebenstein, D.; Burow, M. Natural variation in cross-talk between glucosinolates and onset of flowering in Arabidopsis. Front. Plant Sci. 2015, 6, 697. [Google Scholar] [CrossRef] [Green Version]
- Vik, D.; Mitarai, N.; Wulff, N.; Halkier, B.A.; Burow, M. Dynamic modeling of indole glucosinolate hydrolysis and its impact on auxin signaling. Front. Plant Sci. 2018, 9, 550. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.S.; Ye, W.; Hossain, M.A.; Okuma, E.; Uraji, M.; Nakamura, Y.; Mori, I.C.; Murata, Y. Glucosinolate degradation products, isothiocyanates, nitriles, and thiocyanates, induce stomatal closure accompanied by peroxidase-mediated reactive oxygen species production in Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 2013, 77, 977–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Ballesta, M.C.; Muries, B.; Moreno, D.Á.; Dominguez-Perles, R.; García-Viguera, C.; Carvajal, M. Involvement of a glucosinolate (sinigrin) in the regulation of water transport in Brassica oleracea grown under salt stress. Physiol. Plant 2014, 150, 145–160. [Google Scholar] [CrossRef]
- Bonasia, A.; Lazzizera, C.; Elia, A.; Conversa, G. Nutritional, biophysical and physiological characteristics of wild rocket genotypes as affected by soilless cultivation system, salinity level of nutrient solution and growing period. Front. Plant Sci. 2017, 8, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velasco, P.; Cartea, M.E.; González, C.; Vilar, M.; Ordás, A. Factors affecting the glucosinolate content of kale (Brassica oleracea acephala Group). J. Agric. Food Chem. 2007, 55, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Ballesta, M.; Moreno-Fernández, D.; Castejon, D.; Ochando, C.; Morandini, P.; Carvajal, M. The impact of the absence of aliphatic glucosinolates on water transport under salt stress in Arabidopsis thaliana. Front. Plant Sci. 2015, 6, 524. [Google Scholar] [CrossRef] [Green Version]
- Cartea, M.E.; Velasco, P.; Obregón, S.; Padilla, G.; de Haro, A. Seasonal variation in glucosinolate content in Brassica oleracea crops grown in northwestern Spain. Phytochemistry 2008, 69, 403–410. [Google Scholar] [CrossRef]
- Cámara-Martos, F.; Obregón-Cano, S.; Mesa-Plata, O.; Cartea-González, M.E.; de Haro-Bailón, A. Quantification and in vitro bioaccessibility of glucosinolates and trace elements in Brassicaceae leafy vegetables. Food Chem. 2021, 339, 127860. [Google Scholar] [CrossRef]
- Sotelo, T.; Velasco, P.; Soengas, P.; Rodríguez, V.M.; Cartea, M.E. Modification of leaf glucosinolate contents in Brassica oleracea by divergent selection and effect on expression of genes controlling glucosinolate pathway. Front. Plant Sci. 2016, 7, 1012. [Google Scholar] [CrossRef] [Green Version]
- Santolamazza-Carbone, S.; Velasco, P.; Soengas, P.; Cartea, M.E. Bottom-up and top-down herbivore regulation mediated by glucosinolates in Brassica oleracea var. acephala. Oecologia 2014, 174, 893–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santolamazza-Carbone, S.; Sotelo, T.; Velasco, P.; Cartea, M.E. Antibiotic properties of the glucosinolates of Brassica oleracea var. acephala similarly affect generalist and specialist larvae of two lepidopteran pests. J. Pest Sci. 2016, 89, 195–206. [Google Scholar]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules 2010, 16, 251–280. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.K.; Olsen, C.E.; Petersen, M.K. Acylated flavonol glycosides from cabbage leaves. Phytochemistry 1993, 34, 539–544. [Google Scholar] [CrossRef]
- Wu, S.; Tohge, T.; Cuadros-Inostroza, Á.; Tong, H.; Tenenboim, H.; Kooke, R.; Méret, M.; Keurentjes, J.B.; Nikoloski, Z.; Fernie, A.R.; et al. Mapping the Arabidopsis metabolic landscape by untargeted metabolomics at different environmental conditions. Mol. Plant 2018, 11, 118–134. [Google Scholar] [CrossRef] [Green Version]
- Price, K.R.; Casuscelli, F.; Colquhoun, I.J.; Rhodes, M.J.C. Composition and content of flavonol glycosides in broccoli florets (Brassica olearacea) and their fate during cooking. J. Sci. Food Agric. 1998, 77, 468–472. [Google Scholar] [CrossRef]
- Yannai, S. Dictionary of Food Compounds with CD-ROM: Additives, Flavors, and Ingredients, 1st ed.; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Gonzales, G. Mass Spectrometric Characterization of Flavonoids and in vitro Intestinal Transport and Bioactivity Ghent University; Faculty of Bioscience Engineering: Ghent, Belgium, 2016. [Google Scholar]
- Llorach, R.; Gil-Izquierdo, A.; Ferreres, F.; Tomás-Barberán, F.A. HPLC-DAD-MS/MS ESI Characterization of unusual highly glycosylated acylated flavonoids from cauliflower (Brassica oleracea L. var. botrytis) agroindustrial byproducts. J. Agric. Food Chem. 2003, 51, 3895–3899. [Google Scholar] [CrossRef]
- Ormrod, D.P.; Landry, L.G.; Conklin, P.L. Short-term UV-B radiation and ozone exposure effects on aromatic secondary metabolite accumulation and shoot growth of flavonoid-deficient Arabidopsis mutants. Physiol. Plant 1995, 93, 602–610. [Google Scholar] [CrossRef]
- Olsen, H.; Aaby, K.; Borge, G.I.A. Characterization and quantification of flavonoids and hydroxycinnamic acids in curly kale (Brassica oleracea L. convar. acephala var. sabellica) by HPLC-DAD-ESI-MSn. J. Agric. Food Chem. 2009, 57, 2816–2825. [Google Scholar]
- Rosa, A.S.E. Chemical composition. In Biology of Brassica coenospecies; Gomez-Campo, C., Ed.; Elsevier: Amsterdam, The Netherlands, 1999; pp. 315–357. [Google Scholar]
- Martínez-Sánchez, A.; Gil-Izquierdo, A.; Gil, M.I.; Ferreres, F. A Comparative study of flavonoid compounds, vitamin C, and antioxidant properties of baby leaf Brassicaceae species. J. Agric. Food Chem. 2008, 56, 2330–2340. [Google Scholar] [CrossRef] [PubMed]
- Tanchev, S.S.; Timberlake, C.F. The anthocyanins of red cabbage (Brassica oleracea). Phytochemistry 1969, 8, 1825–1827. [Google Scholar] [CrossRef]
- Wiczkowski, W.; Szawara-Nowak, D.; Topolska, J. Red cabbage anthocyanins: Profile, isolation, identification, and antioxidant activity. Food Res. Int. 2013, 51, 303–309. [Google Scholar] [CrossRef]
- Yin, N.-W.; Wang, S.-X.; Jia, L.-D.; Zhu, M.-C.; Yang, J.; Zhou, B.-J.; Yin, J.-M.; Lu, K.; Wang, R.; Li, J.-N.; et al. Identification and characterization of major constituents in different-colored rapeseed petals by UPLC–HESI-MS/MS. J. Agric. Food Chem. 2019, 67, 11053–11065. [Google Scholar] [CrossRef] [PubMed]
- Ferreres, F.; Valentão, P.; Llorach, R.; Pinheiro, C.; Cardoso, L.; Pereira, J.A.; Sousa, C.; Seabra, R.M.; Andrade, P.B. Phenolic compounds in external leaves of tronchuda cabbage (Brassica oleracea L. var. costata DC). J. Agric. Food Chem. 2005, 53, 2901–2907. [Google Scholar] [CrossRef] [PubMed]
- Biondi, F.; Balducci, F.; Capocasa, F.; Visciglio, M.; Mei, E.; Vagnoni, M.; Mezzetti, B.; Mazzoni, L. Environmental conditions and agronomical factors influencing the levels of phytochemicals in Brassica vegetables responsible for nutritional and sensorial properties. Appl. Sci. 2021, 11, 1927. [Google Scholar] [CrossRef]
- Wu, X.; Huang, H.; Childs, H.; Wu, Y.; Yu, L.; Pehrsson, P.R. Glucosinolates in Brassica vegetables: Characterization and factors that influence distribution, content, and intake. Annu. Rev. Food Sci. Technol. 2021, 12, 485–511. [Google Scholar] [CrossRef]
- Chung, I.-M.; Rekha, K.; Rajakumar, G.; Thiruvengadam, M. Production of glucosinolates, phenolic compounds and associated gene expression profiles of hairy root cultures in turnip (Brassica rapa ssp. rapa). 3 Biotech. 2016, 6, 175. [Google Scholar] [CrossRef] [Green Version]
- Galletti, S.; Barillari, J.; Iori, R.; Venturi, G. Glucobrassicin enhancement in woad (Isatis tinctoria) leaves by chemical and physical treatments. J. Sci. Food Agric. 2006, 86, 1833–1838. [Google Scholar] [CrossRef]
- Jo, J.S.; Bhandari, S.R.; Kang, G.H.; Lee, J.G. Comparative analysis of individual glucosinolates, phytochemicals, and antioxidant activities in broccoli breeding lines. Hortic. Environ. Biotechnol. 2016, 57, 392–403. [Google Scholar] [CrossRef]
- Fiorentino, A.; Ricci, A.; D’Abrosca, B.; Golino, A.; Izzo, A.; Pascarella, M.T.; Piccolella, S.; Esposito, A. Kaempferol glycosides from Lobularia maritima and their potential role in plant interactions. Chem. Biodivers. 2009, 6, 204–217. [Google Scholar] [CrossRef]
- Acquaviva, R.; Russo, A.; Galvano, F.; Galvano, G.; Barcellona, M.L.; Li Volti, G.; Vanella, A. Cyanidin and cyanidin 3-O-β-D-glucoside as DNA cleavage protectors and antioxidants. Cell Biol. Toxicol. 2003, 19, 243–252. [Google Scholar] [CrossRef]
- Razavi, S.M.; Zahri, S.; Zarrini, G.; Nazemiyeh, H.; Mohammadi, S. Biological activity of quercetin-3-O-glucoside, a known plant flavonoid. Russ. J. Bioorganic Chem. 2009, 35, 376–378. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, P.S.; Victorelli, F.D.; Fonseca-Santos, B.; Chorilli, M. A review of analytical methods for p-coumaric acid in plant-based products, beverages, and biological matrices. Crit. Rev. Anal. Chem. 2019, 49, 21–31. [Google Scholar] [CrossRef]
- Rodríguez, V.M.; Chételat, A.; Majcherczyk, P.; Farmer, E.E. Chloroplastic phosphoadenosine phosphosulfate metabolism regulates basal levels of the prohormone jasmonic acid in Arabidopsis leaves. Plant Physiol. 2010, 152, 1335–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinrich, M.; Hettenhausen, C.; Lange, T.; Wünsche, H.; Fang, J.; Baldwin, I.T.; Wu, J. High levels of jasmonic acid antagonize the biosynthesis of gibberellins and inhibit the growth of Nicotiana attenuata stems. Plant J. 2013, 73, 591–606. [Google Scholar] [CrossRef] [PubMed]
- Grandmaison, J.; Ibrahim, R.K. Evidence for nuclear protein binding of flavonol sulfate esters in Flaveria chloraefolia. J. Plant Physiol. 1996, 147, 653–660. [Google Scholar] [CrossRef]
- Saslowsky, D.E.; Warek, U.; Winkel, B.S.J. Nuclear localization of flavonoid enzymes in Arabidopsis. J. Biol. Chem. 2005, 280, 23735–23740. [Google Scholar] [CrossRef] [Green Version]
- Böhl, M.; Tietze, S.; Sokoll, A.; Madathil, S.; Pfennig, F.; Apostolakis, J.; Fahmy, K.; Gutzeit, H.O. Flavonoids affect actin functions in cytoplasm and nucleus. Biophys. J. 2007, 93, 2767–2780. [Google Scholar] [CrossRef] [Green Version]
- Besseau, S.; Hoffmann, L.; Geoffroy, P.; Lapierre, C.; Pollet, B.; Legrand, M. Flavonoid accumulation in Arabidopsis; repressed in lignin synthesis affects auxin transport and plant growth. Plant Cell 2007, 19, 148–162. [Google Scholar] [CrossRef] [Green Version]
- Kupidlowska, E.; Kowalec, M.; Sulkowski, G.; Zobel, A.M. The effect of coumarins on root elongation and ultrastructure of meristematic cell protoplast. Ann. Bot. 1994, 73, 525–530. [Google Scholar] [CrossRef]
- Kliebenstein, D.J.; Lambrix, V.M.; Reichelt, M.; Gershenzon, J.; Mitchell-Olds, T. Gene duplication in the diversification of secondary metabolism: Tandem 2-oxoglutarate-dependent dioxygenases control glucosinolate biosynthesis in Arabidopsis. Plant Cell 2001, 13, 681–693. [Google Scholar] [PubMed] [Green Version]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for comprehensive and integrative metabolomics data analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef] [PubMed]
- Steinbrenner, A.D.; Agerbirk, N.; Orians, C.M.; Chew, F.S. Transient abiotic stresses lead to latent defense and reproductive responses over the Brassica rapa life cycle. Chemoecology 2012, 22, 239–250. [Google Scholar] [CrossRef]
- Cavaiuolo, M.; Cocetta, G.; Spadafora, N.D.; Müller, C.T.; Rogers, H.J.; Ferrante, A. Gene expression analysis of rocket salad under pre-harvest and postharvest stresses: A transcriptomic resource for Diplotaxis tenuifolia. PLoS ONE 2017, 12, e0178119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| m/z | Neutral Mass | Ionization | RT (Min) | Log2(FC) | Molecular Formula | Theoretical Mass | Mass Deviation (ppm) | Fragments | Tentative Identification |
|---|---|---|---|---|---|---|---|---|---|
| A. Peaks detected in negative ionization mode | |||||||||
| 337.0937 | 338.1010 | [M−H]− | 10.1 | 2.1 | C16H18O8 | 338.1002 | 2.514 | 119.05, 163.04, 191.06 | coumaroylquinic acid isomer 1 [24] |
| 771.1991 | 772.2064 | [M−H]− | 10.3 | −1.1 | C33H40O21 | 772.2062 | 0.176 | 283.04, 609.17, 255.03, 422.07, 446.10 | kaempferol sophoroside-hexoside [25] |
| 675.1938 | 338.1005 | [2M−H]− | 10.4 | 2.2 | C16H18O8 | 338.1002 | 1.035 | 163.04, 337.09, 191.06 | coumaroylquinic acid isomer 2 [24] |
| 325.0937 | 326.1010 | [M−H]− | 10.8 | 1.8 | C15H18O8 | 326.1007 | 0.889 | 119.05, 163.04 | coumaroylglucoside [26] |
| 489.1252 | 980.2650 | [M−2H]−2 | 11.2 | −1.2 | C40H52O28 | 980.2645 | 0.490 | 609.15, 284.03, 933.25, 101.02 | kaempferol sophoroside-(dihydroxymethoxy) sophoroside 1 |
| 628.1641 | 629.1714 | [M−H]− | 11.9 | −1.0 | 466.11, 284.03, 161.02 | kaempferol hexoside derivative or isomer | |||
| 625.1416 | 626.1488 | [M−H]− | 11.9 | 3.1 | C27H30O17 | 626.1483 | 0.854 | 299.03, 271.04 | quercetin sophoroside [27] |
| 337.0932 | 338.1005 | [M−H]− | 12.0 | 1.5 | C16H18O8 | 338.1002 | 0.917 | 173.05, 119.05, 93.03, 163.04 | coumaroylquinic acid isomer 3 [24] |
| 569.1518 | 1140.3181 | [M−2H]−2 | 12.6 | -1.8 | C50H60O30 | 1140.3170 | 0.991 | 488.12, 407.09, 815.20, 205.05, 284.03 | kaempferol sinapoylsophoroside-gentobioside [28] |
| 635.1728 | 1272.3601 | [M−2H]−2 | 12.7 | −1.3 | C55H68O34 | 1272.3592 | 0.684 | 473.12, 635.17, 947.24, 119.04, 161.05, 263.07, 323.10 | kaempferol feruloylpentaglucoside [29] |
| 609.1463 | 610.1536 | [M−H]− | 12.8 | 2.1 | C27H30O16 | 610.1534 | 0.295 | 477.09, 285.04 | kaempferol dihexoside [30] |
| 554.1465 | 1110.3075 | [M−2H]−2 | 12.8 | −1.3 | C49H58O29 | 1110.3064 | 1.013 | 473.12, 392.09, 785.19, 284.03, 175.04, 609.14 | kaempferol feruloylsophoroside-cellobioside [28] |
| 337.0935 | 338.1008 | [M−H]− | 12.9 | 1.1 | C16H18O8 | 338.1002 | 1.863 | 173.04, 93.03, 119.05, 163.04, 111.04 | coumaroylquinic acid isomer 4 [24] |
| 755.2047 | 756.2119 | [M−H]− | 13.2 | 1.8 | C33H40O20 | 756.2113 | 0.847 | 283.04, 255.04, 609.17, 227.05, 430.10 | kaempferol gentiobioside-rhamnoside isomer 1 [31] |
| 337.0935 | 338.1014 | [M−H]− | 13.9 | 1.5 | C16H18O8 | 338.1002 | 3.668 | 191.05, 119.05, 163.04, 127.04 | coumaroylquinic acid isomer 5 [24] |
| 771.2003 | 772.2076 | [M−H]− | 14.2 | 3.6 | C33H40O21 | 772.2062 | 1.782 | 447.09, 625.14, 301.03 | quercetin-(rhamnosylhexoside)-hexoside or isomer |
| 625.142 | 626.1493 | [M−H]− | 14.3 | −1.2 | C27H30O17 | 626.1483 | 1.526 | 300.03, 179.0, 445.08, 463.09 | quercetin dihexoside [32] |
| 609.1468 | 610.1540 | [M−H]− | 14.3 | 1.6 | C27H30O16 | 610.1534 | 1.048 | 446.08, 283.02, 463.09, 301.03 | quercetin hexoside-rhamnoside [33] |
| 755.2044 | 756.2117 | [M−H]− | 15.0 | 2.9 | C33H40O20 | 756.2113 | 0.517 | 609.15, 431.10, 285.04 | kaempferol gentiobioside-rhamnoside isomer 2 [34] |
| 593.1518 | 594.1584 | [M−H]− | 15.2 | 5.0 | C27H30O15 | 594.1585 | 0.143 | 430.09, 447.09, 285.04 | kaempferol rutinoside or isomer |
| 639.1569 | 640.1642 | [M−H]− | 15.4 | −3.2 | C28H32O17 | 640.1640 | 0.305 | 314.04, 459.09, 609.15 | (iso)rhamnetin-dihexoside |
| 623.1625 | 624.1705 | [M−H]− | 15.5 | 3.6 | C28H32O16 | 624.1690 | 2.339 | 461.11, 477.10, 315.05 | (iso)rhamnetin-rhamnosylhexoside |
| 657.1752 | 1316.3649 | [M−2H]−2 | 17.0 | −1.3 | C60H68O33 | 1316.3643 | 0.437 | 576.14, 284.03, 175.04, 205.05 | kaempferol-(feruloyl)(sinapoyl)-trihexoside-hexoside or isomer 1 |
| 709.1998 | 710.2071 | [M−H]− | 17.1 | 1.6 | C32H38O18 | 710.2058 | 1.816 | 485.13, 161.03, 223.06, 179.04 | kaempferol derivative or isomer |
| 657.1745 | 1316.3641 | [M−2H]−2 | 17.2 | −1.5 | C60H68O33 | 1316.3643 | 0.141 | 576.15, 284.03, 205.05, 175.04, 947.26 | kaempferol-(feruloyl)(sinapoyl)-trihexoside-hexoside or isomer 2 |
| 415.1977 | 416.2050 | [M−H]− | 17.6 | 1.8 | C20H32O9 | 416.2046 | 0.781 | 44.99, 71.02, 113.03, 101.02 | nicotinic acid hexoside derivative |
| 709.4687 | 710.4760 | [M−H]− | 28.7 | −1.8 | |||||
| 683.4655 | 684.4728 | [M−H]− | 29.5 | −1.8 | |||||
| 683.4659 | 684.4732 | [M−H]− | 29.8 | −2.1 | |||||
| 709.4807 | 710.4880 | [M−H]− | 29.8 | −2.0 | |||||
| B. Peaks detected in positive ionization mode | |||||||||
| 110.0702 | 109.0629 | [M+H]+ | 0.8 | 1.1 | 47.77 | ||||
| 190.0502 | 189.0431 | [M+H]+ | 9.7 | −1.5 | C10H7NO3 | 189.0426 | 2.412 | 116.05, 162.05, 89.04, 144.04 | kynurenic acid |
| 361.0893 | 360.0820 | [M+H]+ | 10.1 | 1.1 | 147.05, 167.06, 91.05 | feruloyl derivative | |||
| 147.0433 | 146.0360 | [M+H]+ | 10.1 | 1.4 | C9H6O2 | 146.0368 | 5.478 | 65.04, 91.05 | coumarin isomer 12 |
| 449.1079 | 448.1006 | [M+H]+ | 10.2 | −1.2 | C21H20O11 | 448.1006 | 0.036 | 305.07, 287.05 | cyanidin-hexoside or isomer |
| 339.1074 | 338.1001 | [M+H]+ | 10.4 | 1.2 | C16H18O8 | 338.1002 | 0.177 | 147.04, 119.05, 91.05 | coumaroylquinic acid isomer 2 [24] |
| 361.0894 | 360.0821 | [M+H]+ | 10.4 | 1.1 | 147.04, 167.05, 140.99, 91.05 | feruloyl derivative | |||
| 803.2232 | 802.2160 | [M+H]+ | 11.4 | −2.5 | C34H42O22 | 802.2168 | 1.022 | 317.06, 479.12 | ophioglonol-dihexoside-hexoside or isomer 1 |
| 147.0429 | 146.0356 | [M+H]+ | 12.1 | 1.9 | C9H6O2 | 146.0368 | 8.217 | 65.04, 91.05, 63.03, 55.05 | coumarin isomer 22 |
| 773.2121 | 772.2049 | [M+H]+ | 12.6 | 1.5 | C33H41O21+ | 772.2062 | 1.740 | 287.05, 303.05, 449.10 | cyanidin sophoroside-hexoside [35] |
| 147.0440 | 146.0367 | [M+H]+ | 12.9 | 1.7 | C9H6O2 | 146.0368 | 0.342 | 91.05, 65.04, 53.04 | coumarin isomer 32 |
| 803.2232 | 802.2160 | [M+H]+ | 13.1 | −2.5 | C34H42O22 | 802.2168 | 1.022 | 317.06, 85.03, 145.05, 479.12 | ophioglonol-dihexoside-hexoside or isomer 2 |
| 757.2173 | 756.2100 | [M+H]+ | 13.2 | 2.5 | C33H40O20 | 756.2113 | 1.744 | 287.05, 85.03, 433.11, 145.05 | cyanidin-rhamnosylhexoside-hexoside or isomer 1 |
| 949.2599 | 948.2527 | [M+H]+ | 13.4 | 2.8 | C43H48O24 | 948.2536 | 0.938 | 177.05, 287.05, 339.10, 449.10 | cyanidin-(feruloyldihexoside)-hexoside or isomer |
| 233.1654 | 232.1581 | [M+H]+ | 13.5 | 2.5 | |||||
| 979.2703 | 978.2634 | [M+H]+ | 13.5 | 3.1 | C44H51O25+ | 978.2641 | 0.721 | 287.05, 369.11, 449.11, 611.16 | cyanidin-(sinapoyldihexoside)-hexoside or isomer |
| 919.2493 | 918.2420 | [M+H]+ | 13.7 | 3.1 | C42H46O23 | 918.2430 | 1.040 | 163.04, 287.05, 325.09, 307.08 | cyanidin-(coumaroyldihexoside)-hexoside [36] |
| 147.0393 | 146.0320 | [M+H]+ | 13.9 | 1.3 | 91.05, 65.04,55.06, 53.04 | ||||
| 773.2127 | 772.2054 | [M+H]+ | 14.1 | 3.6 | C33H40O21 | 772.2062 | 1.080 | 303.05, 85.03, 145.05, 287.05, 449.11 | delphinidin-rutinoside-hexoside [36] |
| 963.2752 | 962.2679 | [M+H]+ | 14.2 | 2.5 | C44H51O24+ | 962.2692 | 1.319 | 287.05, 369.12, 433.11, 207.07 | cyanidin-(sinapoylhexoside)-rhamnosylhexoside or isomer |
| 465.1027 | 464.0954 | [M+H]+ | 14.3 | −1.1 | C21H20O12 | 464.0955 | 0.086 | 303.05, 85.03, 127.04 | delphinidin-hexoside isomer [37] |
| 611.1600 | 610.1527 | [M+H]+ | 14.3 | 2.1 | C27H30O16 | 610.1534 | 1.082 | 303.05, 287.05, 85.03, 127.04, 449,11 | cyanidin-dihexoside or isomer 1 |
| 633.2012 | 632.1940 | [M+H]+ | 14.4 | 2.2 | C27H36O17 | 632.1953 | 2.048 | 147.04, 165.05, 127.04, 85.03, 309.10 | feruloyl derivative |
| 933.2648 | 932.2575 | [M+H]+ | 14.5 | 3.0 | C43H48O23 | 932.2586 | 1.260 | 177.05, 287.05, 339.11, 321,10, 127.04, 433.11 | cyanidin-(feruloylhexoside)-rhamnosylhexoside or isomer |
| 903.2545 | 902.2472 | [M+H]+ | 14.7 | 3.2 | C42H47O22+ | 902.2481 | 0.964 | 147.04, 287.05, 309.10, 291.09, 433.11 | cyanidin-(coumaroylhexoside)-rhamnosylhexoside or isomer |
| 757.2172 | 756.2099 | [M+H]+ | 15.0 | 3.2 | C33H40O20 | 756.2113 | 1.889 | 287.05, 85.03, 127.04, 433.11 | cyanidin-rhamnosylhexoside-hexoside or isomer 2 |
| 369.1184 | 368.1111 | [M+H]+ | 15.0 | −1.4 | C17H20O9 | 368.1107 | 0.964 | 175.04, 207.07, 147.04 | feruloyl quinic acid isomer 1 [24] |
| 611.1599 | 610.1526 | [M+H]+ | 15.2 | −1.5 | C27H30O16 | 610.1534 | 1.246 | 287.05, 85.03, 127.04, 97.03, 145.05 | cyanidin-dihexoside or isomer 2 |
| 394.1136 | 393.1063 | [M+H]+ | 15.3 | −1.1 | |||||
| 625.1752 | 624.1679 | [M+H]+ | 15.5 | 4.0 | C28H32O16 | 624.1690 | 1.794 | 317.07, 85.03, 303.05, 127.04, 97.03 | petunidin-rhamnosylhexoside or isomer |
| 500.1758 | 499.1688 | [M+H]+ | 15.8 | 1.6 | C22H29NO12 | 499.1690 | 0.459 | 130.07, 85.03, 160.08, 307.10 | indolylacetyl dihexoside |
| 468.1501 | 467.1428 | [M+H]+ | 15.8 | 1.6 | |||||
| 369.1183 | 368.1110 | [M+H]+ | 16.0 | −1.1 | C17H20O9 | 368.1107 | 0.774 | 175.04, 207.07, 147.04, 119.05 | feruloyl quinic acid isomer 2 [24] |
| 517.1547 | 516.1474 | [M+H]+ | 16.5 | 1.8 | C22H28O14 | 516.1479 | 0.903 | 193.05, 161.02, 85.03, 127.04, 69.03, 97.03 | caffeoyl quinic acid hexoside or isomer 1 |
| 517.1553 | 516.1479 | [M+H]+ | 16.8 | 1.5 | C22H28O14 | 516.1479 | 0.038 | 193.05, 161.02, 85.03, 127.04, 69.03, 97.03 | caffeoyl quinic acid hexoside or isomer 2 |
| 393.1893 | 370.1997 | [M+Na]+ | 17.6 | 2.1 | C19H30O7 | 370.1992 | 1.364 | ||
| 457.2064 | 456.1995 | [M+H]+ | 18.8 | 1.9 | C22H32O10 | 456.1996 | 0.175 | 191.14, 147.11, 121.07, 93.07, 69.07, 209.16 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poveda, J.; Velasco, P.; de Haro, A.; Johansen, T.J.; McAlvay, A.C.; Möllers, C.; Mølmann, J.A.B.; Ordiales, E.; Rodríguez, V.M. Agronomic and Metabolomic Side-Effects of a Divergent Selection for Indol-3-Ylmethylglucosinolate Content in Kale (Brassica oleracea var. acephala). Metabolites 2021, 11, 384. https://doi.org/10.3390/metabo11060384
Poveda J, Velasco P, de Haro A, Johansen TJ, McAlvay AC, Möllers C, Mølmann JAB, Ordiales E, Rodríguez VM. Agronomic and Metabolomic Side-Effects of a Divergent Selection for Indol-3-Ylmethylglucosinolate Content in Kale (Brassica oleracea var. acephala). Metabolites. 2021; 11(6):384. https://doi.org/10.3390/metabo11060384
Chicago/Turabian StylePoveda, Jorge, Pablo Velasco, Antonio de Haro, Tor J. Johansen, Alex C. McAlvay, Christian Möllers, Jørgen A.B. Mølmann, Elena Ordiales, and Víctor M. Rodríguez. 2021. "Agronomic and Metabolomic Side-Effects of a Divergent Selection for Indol-3-Ylmethylglucosinolate Content in Kale (Brassica oleracea var. acephala)" Metabolites 11, no. 6: 384. https://doi.org/10.3390/metabo11060384