
Tree Species and Epiphyte Taxa Determine the “Metabolomic niche” of Canopy Suspended Soils in a Species-Rich Lowland Tropical Rainforest


 ,
,  , , and
, , and
Abstract
:1. Introduction
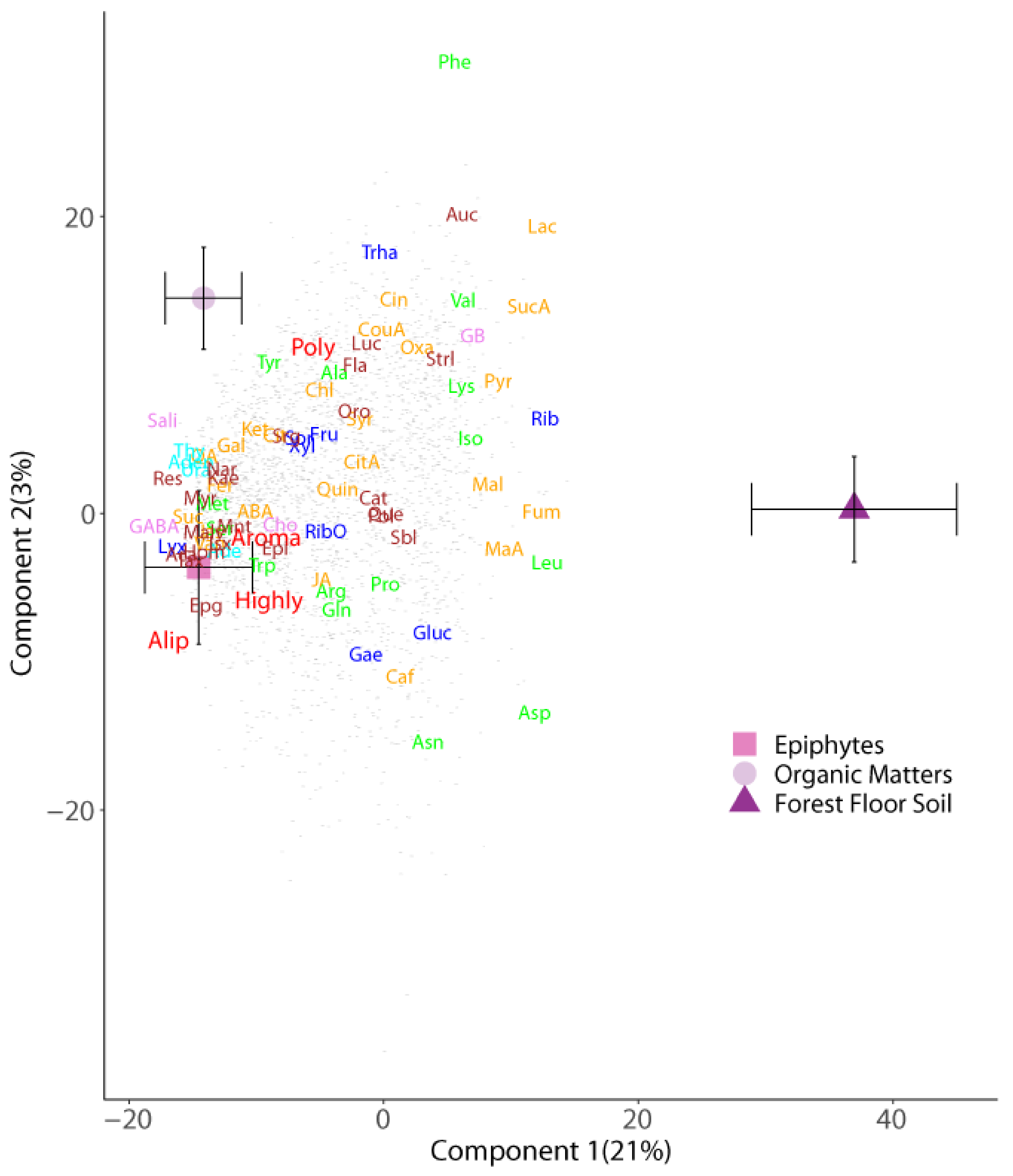
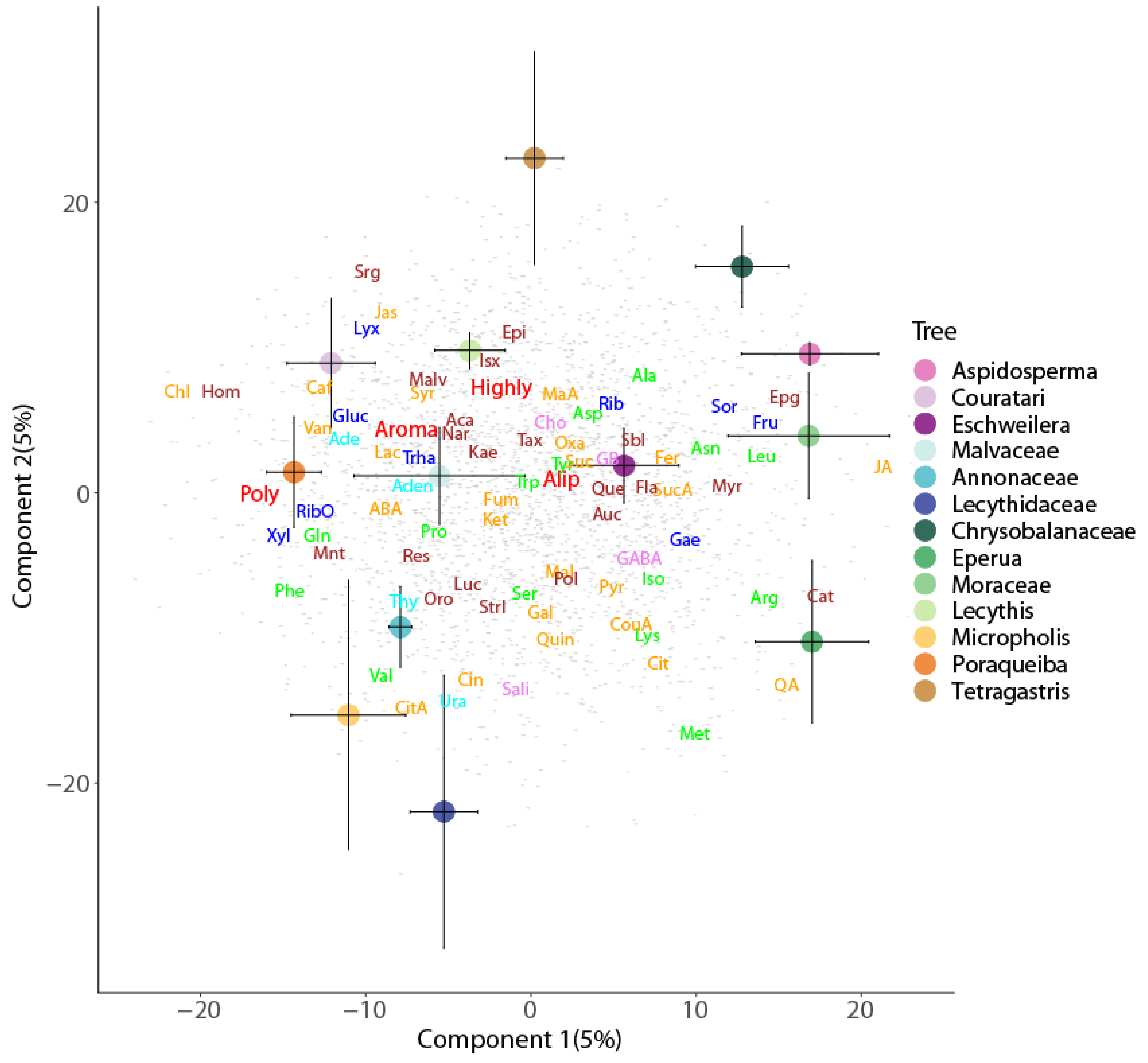
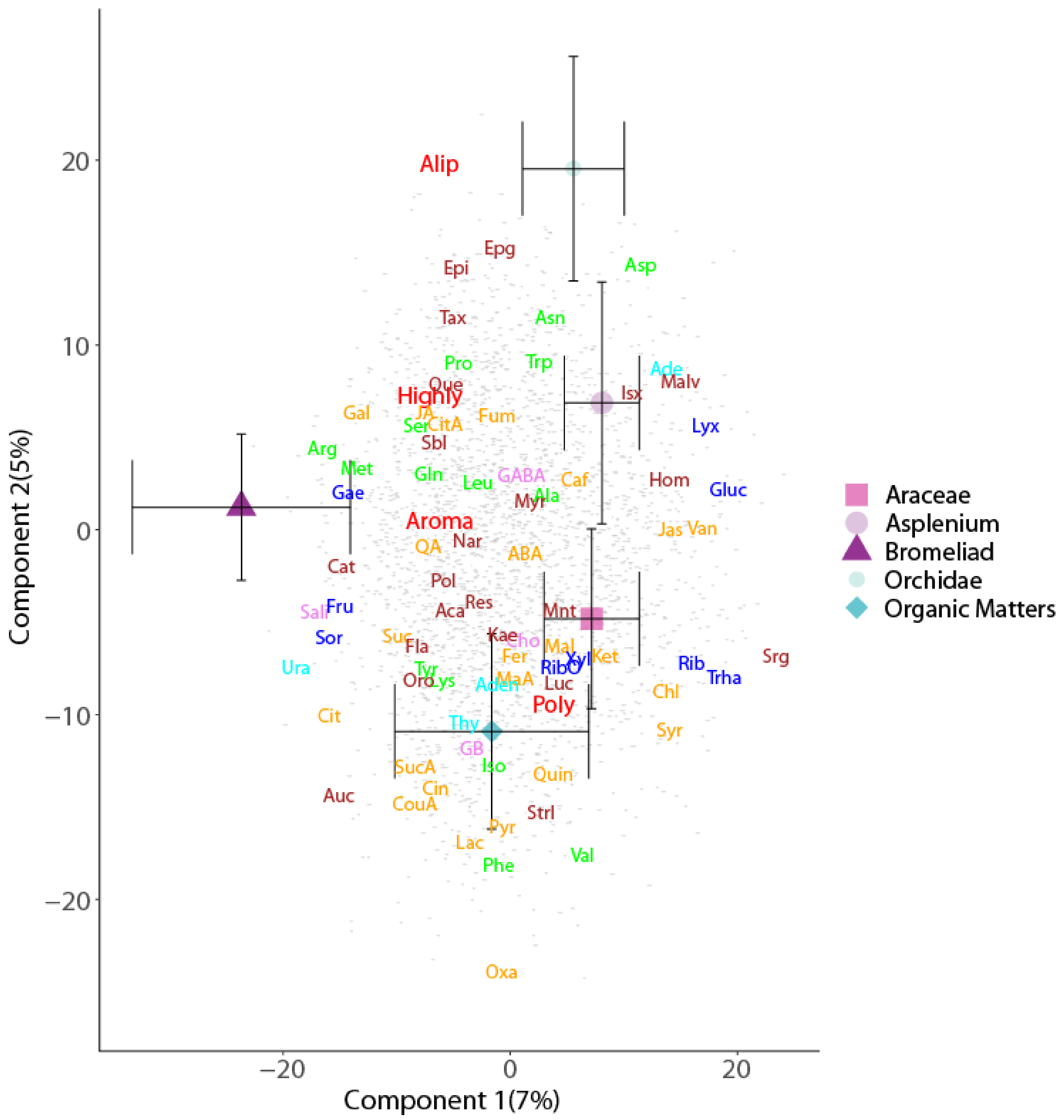
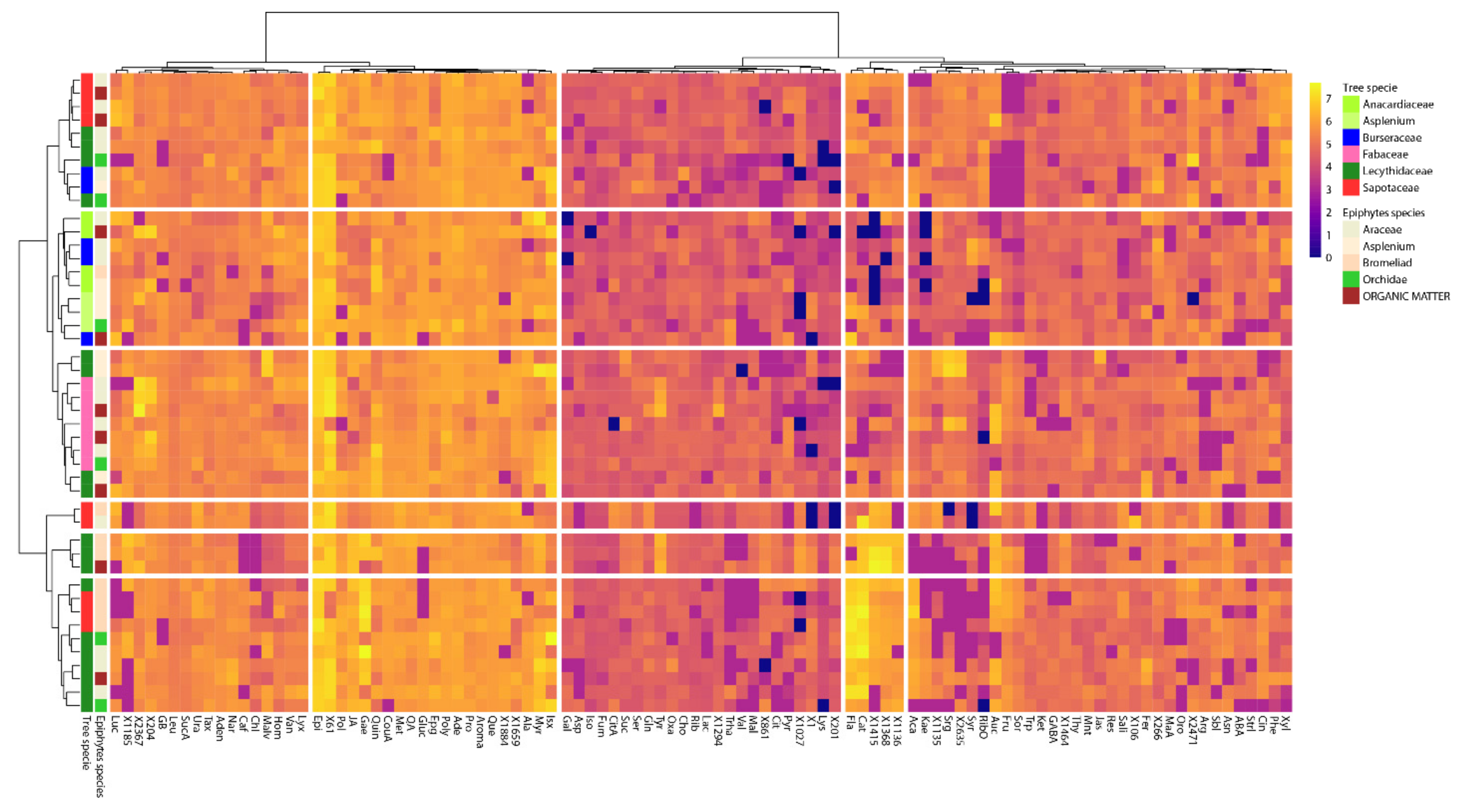
2. Results
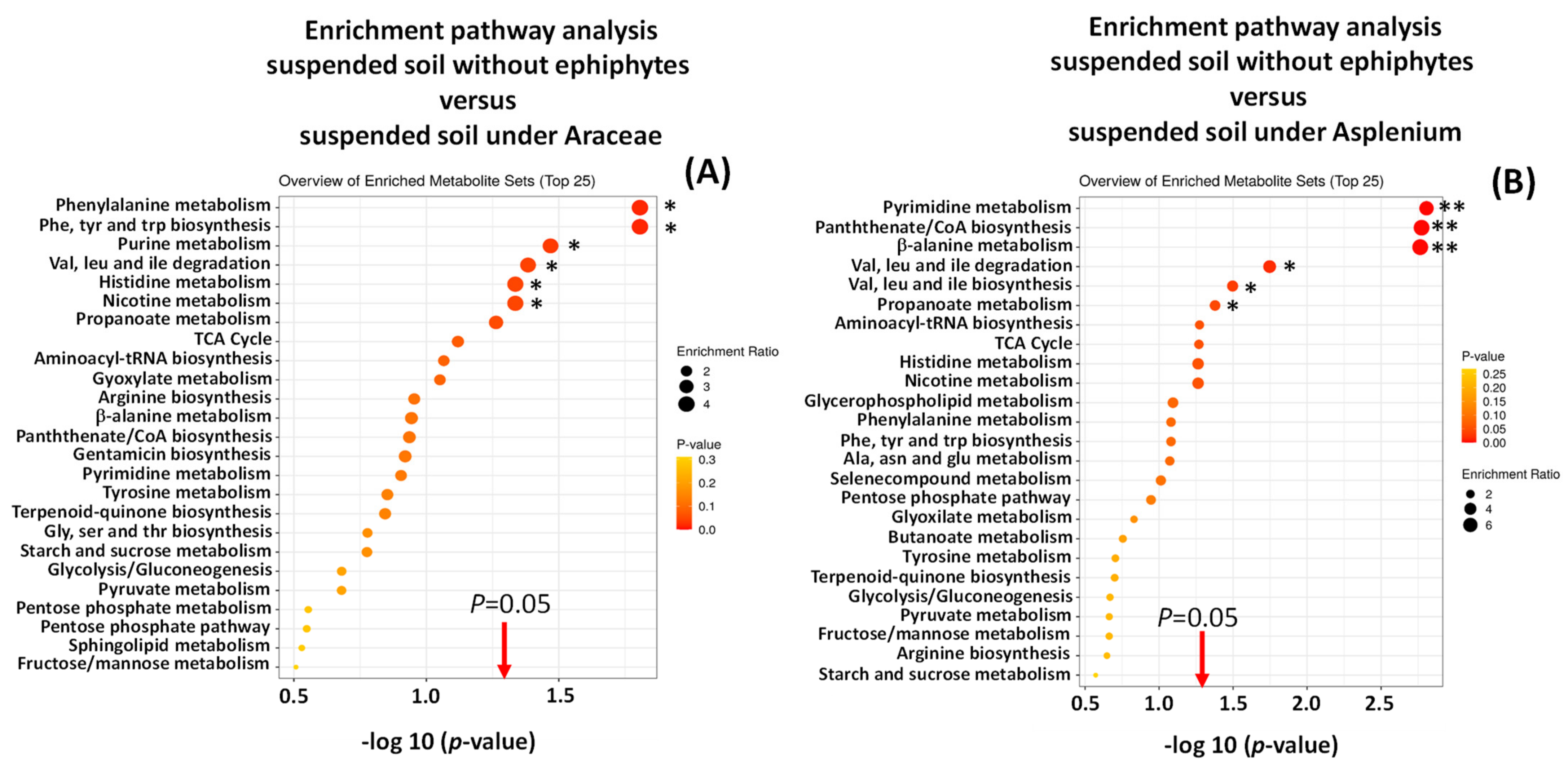
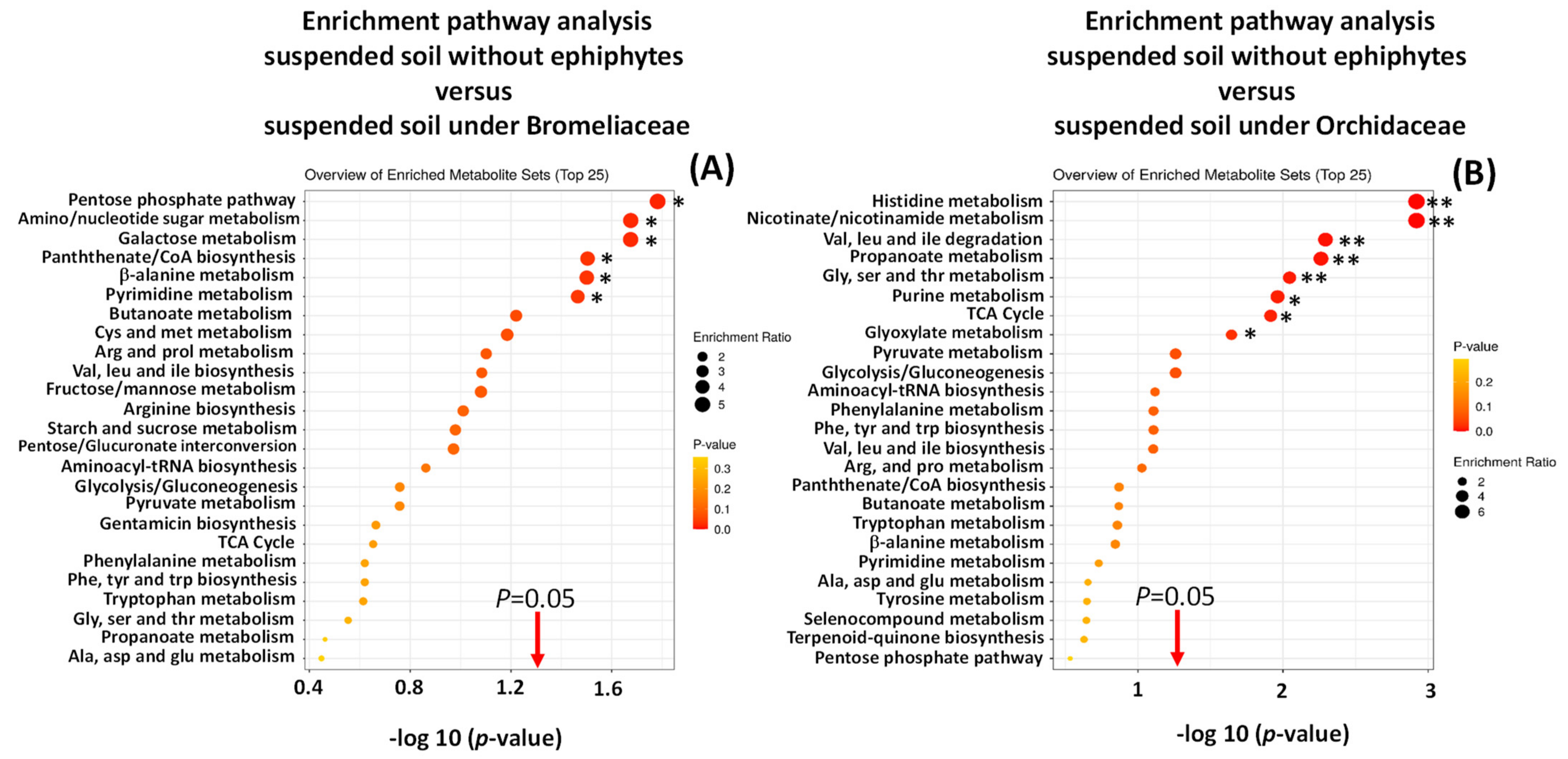
Enrichment of Metabolic Pathways
3. Discussion
3.1. Metabolome of Forest Floor and Canopy Soils
3.2. Metabolic Profile Differences among Canopy Soils
3.3. Global Overview of the High Diversity of Metabolomic Profile in the Studied Soils
4. Materials and Methods
4.1. Study Area
4.2. Soil Sampling and Analysis
4.3. Metabolite Extraction and Analysis
4.4. Metabolite Identification and Quantification
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Connell, J.H. Diversity in Tropical Rain Forests and Coral Reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- John, R.; Dalling, J.W.; Harms, K.E.; Yavitt, J.B.; Stallard, R.F.; Mirabello, M.; Hubbell, S.P.; Valencia, R.; Navarrete, H.; Vallejo, M.; et al. Soil nutrients influence spatial distributions of tropical tree species. Proc. Natl. Acad. Sci. USA 2007, 104, 864–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, J.S. Plant diversity in tropical forests: A review of mechanisms of species coexistence. Oecologia 2002, 130, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nat. Cell Biol. 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Achard, F.; Eva, H.D.; Stibig, H.-J.; Mayaux, P.; Gallego, J.; Richards, T.; Malingreau, J.-P. Determination of Deforestation Rates of the World’s Humid Tropical Forests. Science 2002, 297, 999–1002. [Google Scholar] [CrossRef] [Green Version]
- Volkov, I.; Banavar, J.R.; He, F.; Hubbell, S.P.; Maritan, A. Density dependence explains tree species abundance and diversity in tropical forests. Nat. Cell Biol. 2005, 438, 658–661. [Google Scholar] [CrossRef] [PubMed]
- Aiba, S.-I.; Sawada, Y.; Takyu, M.; Seino, T.; Kitayama, K.; Repin, R. Structure, floristics and diversity of tropical montane rain forests over ultramafic soils on Mount Kinabalu (Borneo) compared with those on non-ultramafic soils. Aust. J. Bot. 2015, 63, 191–203. [Google Scholar] [CrossRef]
- LeBauer, D.S.; Treseder, K.K. Nitrogen Limitation of Net Primary Productivity in Terrestrial Ecosystems Is Globally Distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Fujii, K.; Shibata, M.; Kitajima, K.; Ichie, T.; Kitayama, K.; Turner, B. Plant–soil interactions maintain biodiversity and functions of tropical forest ecosystems. Ecol. Res. 2018, 33, 149–160. [Google Scholar] [CrossRef]
- Xu, W.; Ci, X.; Song, C.; He, T.; Zhang, W.; Li, Q.; Li, J. Soil phosphorus heterogeneity promotes tree species diversity and phylogenetic clustering in a tropical seasonal rainforest. Ecol. Evol. 2016, 6, 8719–8726. [Google Scholar] [CrossRef]
- Pennington, R.T.; Lavin, M.; Oliveira-Filho, A. Woody Plant Diversity, Evolution, and Ecology in the Tropics: Perspectives from Seasonally Dry Tropical Forests. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 437–457. [Google Scholar] [CrossRef] [Green Version]
- Matos, F.A.R.; Magnago, L.; Gastauer, M.; Carreiras, J.M.B.; Simonelli, M.; Meira-Neto, J.A.A.; Edwards, D.P. Effects of landscape configuration and composition on phylogenetic diversity of trees in a highly fragmented tropical forest. J. Ecol. 2016, 105, 265–276. [Google Scholar] [CrossRef]
- Martins, K.; Marques, M.; dos Santos, E.; Marques, R. Effects of soil conditions on the diversity of tropical forests across a successional gradient. For. Ecol. Manag. 2015, 349, 4–11. [Google Scholar] [CrossRef]
- Mandl, N.A.; Kessler, M.; Gradstein, R. Effects of environmental heterogeneity on species diversity and composition of terrestrial bryophyte assemblages in tropical montane forests of southern Ecuador. Plant Ecol. Divers. 2009, 2, 313–321. [Google Scholar] [CrossRef]
- Clark, D.B.; Clark, D.A.; Read, J.M. Edaphic variation and the mesoscale distribution of tree species in a neotropical rain forest. J. Ecol. 1998, 86, 101–112. [Google Scholar] [CrossRef]
- Becerra, J.X. On the factors that promote the diversity of herbivorous insects and plants in tropical forests. Proc. Natl. Acad. Sci. USA 2015, 112, 6098–6103. [Google Scholar] [CrossRef] [Green Version]
- Nadkarni, N.M.; Schafer, D.; Matelson, T.J.; Solano, R. Comparison of srboreal and terrestrial soil characteristics in a lower montane forest, Monteverde, Costa Rica. Pedobiologia 2002, 46, 24–33. [Google Scholar] [CrossRef]
- Lowman, M.D.; Rinker, H.B. Forest Canopies, 2nd ed.; Academic Press: Cambridge, MA, USA, 2004; p. 544. [Google Scholar]
- Ozanne, C.M.P.; Anhuf, D.; Boulter, S.L.; Keller, M.; Kitching, R.L.; Körner, C.; Meinzer, F.C.; Mitchell, A.W.; Nakashizuka, T.; Silva Dias, P.L.; et al. Biodiversity meets the almosphere: A global view of forest canopies. Science 2003, 301, 183–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Floren, A.; von Rintelen, T.; Hebert, P.D.N.; de Araujo, B.C.; Schmidt, S.; Balke, M.; Narakusumo, R.P.; Peggie, D.; Ubaidillah, R.; von Rintelen, K.; et al. Integrative ecological and molecular analysis indicate high diversity and strict elevational separation of canopy beetles in tropical mountain forests. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Paoletti, M.G.; Taylor, R.A.J.; Stinner, B.R.; Stinner, D.H.; Benzing, D.H. Diversity of soil fauna in the canopy and forest floor of a Venezuela cloud forest. J. Trop. Ecol. 1991, 7, 373–383. [Google Scholar] [CrossRef]
- Beaulieu, F.; Walter, D.E.; Proctor, H.C.; Kitching, R.L. The Canopy starts at 0.5 cm: Predatory mites (Acari: Mesostigmata) differ between rain forest floor soil and suspended soil at any height. Biotropica 2010, 42, 704–709. [Google Scholar] [CrossRef]
- Cardelús, C.L.; Mack, M.C.; Woods, C.; Demarco, J.; Treseder, K. The influence of tree species on canopy soil nutrient status in a tropical lowland wet forest in Costa Rica. Plant Soil 2008, 318, 47–61. [Google Scholar] [CrossRef]
- Lesica, P.; Antibus, R.K. Canopy Soils and epiphyte richness. Res. Explor. 1991, 7, 156. [Google Scholar]
- Looby, C.I.; Hollenbeck, E.C.; Treseder, K.K. Fungi in the Canopy: How Soil Fungi and Extracellular Enzymes Differ Between Canopy and Ground Soils. Ecosystems 2019, 23, 768–782. [Google Scholar] [CrossRef] [Green Version]
- Victoriano-Romero, E.; García-Franco, J.G.; Mehltreter, K.; Valencia-Díaz, S.; Toledo-Hernandez, A.; Flores-Palacios, A. Ephiphyte associations and canopy soil volumen: Nutrient capital and factors influencing soil retention in the canopy. Plant Biol. 2019, 22, 541–552. [Google Scholar] [CrossRef]
- Janzen, D.H. Herbivores and the Number of Tree Species in Tropical Forests. Am. Nat. 1970, 104, 501–528. [Google Scholar] [CrossRef]
- Leroy, C.; Carrias, J.-F.; Céréghino, R.; Corbara, B. The contribution of microorganisms and metazoans to mineral nutrition in bromeliads. J. Plant Ecol. 2016, 9, 241–255. [Google Scholar] [CrossRef]
- Lyons, B.; Nadkarni, N.M.; North, M.P. Spatial distribution and succession of epiphytes on Tsuga heterophylla (western hemlock) in an old-growth Douglas-fir forest. Can. J. Bot. 2000, 78, 957–968. [Google Scholar] [CrossRef]
- Neider, J.; Prosperí, J.; Michaloud, G. Epiphytes and their contribution to canopy diversity. Plant Ecol. 2001, 153, 51–63. [Google Scholar] [CrossRef]
- Zotz, G. The systematic distribution of vascular epiphytes—A critical update. Bot. J. Linn. Soc. 2013, 171, 453–481. [Google Scholar] [CrossRef] [Green Version]
- Gotsch, S.G.; Nadkarni, N.; Darby, A.; Glunk, A.; Dix, M.; Davidson, K.; Dawson, T.E. Life in the treetops: Ecophysiological strategies of canopy epiphytes in a tropical montane cloud forest. Ecol. Monogr. 2015, 85, 393–412. [Google Scholar] [CrossRef] [Green Version]
- Benzing, D.H. Bromeliaceae: Profile of an Adaptive Radiation; Cambridge University Press: Cambridge, MA, USA, 2000. [Google Scholar] [CrossRef]
- Ngai, J.T.; Srivastava, D.S. Predators accelerate nutrient cycling in a bromeliad ecosystem. Science 2006, 314, 963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wardle, D.; Yeates, G.; Barker, G.; Bonner, K. The influence of plant litter diversity on decomposer abundance and diversity. Soil Biol. Biochem. 2006, 38, 1052–1062. [Google Scholar] [CrossRef]
- Pittl, E.; Innerebner, G.; Wanek, W.; Insam, H. Microbial communities of arboreal and ground soils in the Esquinas rainforest, Costa Rica. Plant Soil 2009, 329, 65–74. [Google Scholar] [CrossRef]
- Donald, J.; Bonnett, S.; Cutler, M.; Majalap, N.; Maxfield, P.; Ellwood, M.D.F. Physical Conditions Regulate the Fungal to Bacterial Ratios of a Tropical Suspended Soil. Forest 2017, 8, 474. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, A.; Kitching, R.L.; Cao, M.; Creedy, T.J.; Fayle, T.; Freiberg, M.; Hewitt, C.N.; Itioka, T.; Koh, L.P.; Ma, K.; et al. Forests and Their Canopies: Achievements and Horizons in Canopy Science. Trends Ecol. Evol. 2017, 32, 438–451. [Google Scholar] [CrossRef] [Green Version]
- Sardans, J.; Peñuelas, J.; Rivas-Ubach, A. Ecological metabolomics: Overview of current developments and future challenges. Chemoecology 2011, 21, 191–225. [Google Scholar] [CrossRef]
- Chen, M.-S. Inducible direct plant defense against insect herbivores: A review. Insect Sci. 2008, 15, 101–114. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics--the link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef]
- Gargallo-Garriga, A.; Wright, S.J.; Sardans, J.; Pérez-Trujillo, M.; Oravec, M.; Večeřová, K.; Urban, O.; Fernández-Martínez, M.; Parella, T.; Peñuelas, J. Long-term fertilization determines different metabolomic profiles and responses in saplings of three rainforest tree species with different adult canopy position. PLoS ONE 2017, 12, e0177030. [Google Scholar] [CrossRef] [Green Version]
- Hewavitharana, S.S.; Klarer, E.; Reed, A.J.; Leisso, R.; Poirier, B.; Honaas, L.; Rudell, D.R.; Mazzola, M. Temporal Dynamics of the Soil Metabolome and Microbiome during Simulated Anaerobic Soil Disinfestation. Front. Microbiol. 2019, 10, 2365. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Li, X.; Yao, S.; Yang, X.; Jiang, X. Correlations between soil metabolomics and bacterial community structures in the pepper rhizosphere under plastic greenhouse cultivation. Sci. Total Environ. 2020, 728, 138439. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J.; Ogaya, R.; Estiarte, M. Nutrient stoichiometric relations and biogeochemical niche in coexisting plant species: Effect of simulated climate change. Polish J. Ecol. 2008, 56, 613–622. [Google Scholar]
- Bundy, J.G.; Davey, M.P.; Viant, M.R. Environmental metabolomics: A critical review and future perspectives. Metabolomics 2009, 5, 3–21. [Google Scholar] [CrossRef]
- Musila, W.; Todt, H.; Uster, D.; Dalitz, H. Is Geodiversity Correlated to Biodiversity? A Case Study of the Relationship between Spatial Heterogeneity of Soil Resources and Tree Diversity in a Western Kenyan Rainforest. In African Biodiversity; Springer: Boston, MA, USA, 2005; pp. 405–414. [Google Scholar]
- Schreeg, L.A.; Mack, M.C.; Turner, B.L. Leaf litter inputs decrease phosphate sorption in a strongly weathered tropical soil over two time scales. Biogeochemistry 2013, 113, 507–524. [Google Scholar] [CrossRef]
- Osborne, B.B.; Nasto, M.K.; Soper, F.M.; Asner, G.P.; Balzotti, C.S.; Cleveland, C.C.; Taylor, P.G.; Townsend, A.R.; Porder, S. Leaf litter inputs reinforce islands on nitrogen fertility in a lowland tropical rainforest. Biogeochemistry 2020, 147, 293–306. [Google Scholar] [CrossRef]
- Matson, A.L.; Corre, M.D.; Burneo, J.I.; Veldkamp, E. Free-living nitrogen fixation responds to elevated nutrient inputs in tropical montane forest floor and canopy soils of southern Ecuador. Biogeochemistry 2015, 122, 281–294. [Google Scholar] [CrossRef]
- Nadkarni, N.M.; Schaefer, D.; Matelson, T.J.; Solano, R. Biomass and nutrient pools of canopy and terrestrial components in a primary and a secondary montane cloud forest, Costa Rica. For. Ecol. Manag. 2004, 198, 223–236. [Google Scholar] [CrossRef]
- Scheffers, B.R.; Phillips, B.L.; Shoo, L.P. Asplenium bird’s nets ferns in rainforest canopies are climate-contingent refuges for frogs. Glob. Ecol. Conserv. 2014, 2, 37–46. [Google Scholar]
- Aguilar-Cruz, Y.; Garcia-Franco, J.G.; Zotz, G. Microsites and early litter decomposition patterns in the soil and forest canopy at tregional scale. Biogeochemistry 2020, 151, 15–30. [Google Scholar] [CrossRef]
- Zaharescu, D.G.; Burghelea, C.I.; Dontsova, K.; Presler, J.K.; Hunt, E.A.; Domanik, K.J.; Amistadi, M.K.; Sandhaus, S.; Munoz, E.N.; Gaddis, E.E.; et al. Ecosystem-bedrock interaction changes nutrient compartmentalization during early oxidative weathering. Sci. Rep. 2019, 9, 15006. [Google Scholar] [CrossRef] [Green Version]
- Enloe, H.A.; Quideau, S.A.; Graham, R.C.; Sillett, S.C.; Oh, S.-W.; Wasylishen, R.E. Soil Organic Matter Processes in Old-Growth Redwood Forest Canopies. Soil Sci. Soc. Am. J. 2010, 74, 161–171. [Google Scholar] [CrossRef]
- Marí, M.L.G.; Toledo, J.J.; Nascimento, H.E.M.; Zartman, C.E. Regional and Fine Scale Variation of Holoepiphyte Community Structure in Central Amazonian White-Sand Forests. Biotropica 2016, 48, 70–80. [Google Scholar] [CrossRef]
- Gotsch, S.G.; Nadkarni, N.; Amici, A. The functional roles of epiphytes and arboreal soils in tropical montane cloud forests. J. Trop. Ecol. 2016, 32, 455–468. [Google Scholar] [CrossRef]
- Benner, J.W. Epiphytes preferentially colonize high-phosphorus host trees in unfertilized Hawaiian montane forests. Bryol. 2011, 114, 335–345. [Google Scholar] [CrossRef]
- Ledo, A.; Burslem, D.F.; Condés, S.; Montes, F. Micro-scale habitat associations of woody plants in a neotropical cloud forest. J. Veg. Sci. 2013, 24, 1086–1097. [Google Scholar] [CrossRef] [Green Version]
- Zotz, G.; Schultz, S. The vascular epiphytes of a lowland forest in Panama—species composition and spatial structure. Plant Ecol. 2007, 195, 131–141. [Google Scholar] [CrossRef]
- Boelter, C.R.; Dambos, C.S.; Nascimiento, H.E.M.; Zartman, C.E. A tangled web in tropical tree-tops: Effects of edaphic variation, neighborhood phorophyte composition and bark characteristics on epiphytes in a Central Amazonian forest. J. Veg. Sci. 2014, 25, 1090–1099. [Google Scholar] [CrossRef]
- Obregon, A.; Gehrig-Downie, C.; Gradstein, S.R.; Rollenbeck, R.; Bendix, J. Canopy level fog occurrence in a tropical lowland forest of French Guaiana as a prerequisite for high epiphyte diversity. Agric. For. Metereol. 2011, 151, 290–300. [Google Scholar] [CrossRef]
- Bohlman, S.A.; Matelson, T.J.; Nadkarni, N.M. Moisture and Temperature Patterns of Canopy Humus and Forest Floor Soil of a Montane Cloud Forest, Costa Rica. Biotropica 1995, 27, 13. [Google Scholar] [CrossRef]
- Benner, J.W.; Vitousek, P.M. Development of a diverse epiphyte community in response to phosphorus fertilization. Ecol. Lett. 2007, 10, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Ruan, H.H.; Zou, X.M.; Myster, R.W. Response of major soil decomposers to landscape in a Puerto Rican rainforest. Soil Sci. 2005, 170, 202–211. [Google Scholar] [CrossRef]
- Sahu, S.C.; Dhal, N.K.; Lal, B.; Mohanty, R.C. Differences in tree species diversity and soil nutrient status in a tropical sacred forest ecosystem on Niyamgiri hill range, Eastern Ghats, India. J. Mt. Sci. 2012, 9, 492–500. [Google Scholar] [CrossRef]
- Prada, C.M.; Morris, A.; Andersen, K.M.; Turner, B.L.; Caballero, P.; Dalling, J.W. Soils and rainfall drive landscape-scale changes in the diversity and functional composition of three communities in premontane tropical forest. J. Veg. Sci. 2017, 278, 859–870. [Google Scholar] [CrossRef]
- Amani, C. Impact of Soil Heterogeneity on Forest Structure and Diversity of Tree Species in the Central Congo Basin. Int. J. Plant Sci. 2018, 179, 198–208. [Google Scholar] [CrossRef]
- Paoli, G.D.; Curran, L.M.; Zak, D.R. Soil nutrients and beta diversity in the Bornean Dipterocarpaceae: Evidence for niche partitioning by tropical rain forest trees. J. Ecol. 2006, 94, 157–170. [Google Scholar] [CrossRef] [Green Version]
- Kuzmeier, S.; Wiedemann, T.; Biber, P.; Schad, P.; Krasilnikov, P.V. Effects of Edaphic factors on the tree stand siversity in a tropical forest of Sierra Madre del Sur, Mexico. Eurasian Soil Sci. 2012, 45, 740–751. [Google Scholar] [CrossRef]
- Ewel, J.J.; Mazzarino, M.J.; Celis, G. Soil Changes in Model Tropical Ecosystems: Effects of Stand Longevity Outweigh Plant Diversity and Tree Species Identity in a Fertile Volcanic Soil. Ecosystems 2014, 17, 820–836. [Google Scholar] [CrossRef]
- Sarvade, S.; Gupta, B.; Singh, M. Composition, diversity and distribution of tree species in response to changing soil properties with increasing distance from water source—A case study of Gobind Sagar Reservoir in India. J. Mt. Sci. 2016, 13, 522–533. [Google Scholar] [CrossRef]
- Gehrig--Downie, C.; Obregón, A.; Bendix, J.; Gradstein, R.S. Epiphyte biomass and canopy microclimate in the tropical lowland cloud forest of French Guaiana. Biotropica 2011, 43, 591–596. [Google Scholar] [CrossRef]
- Woods, C.L.; Cardelús, C.L.; DeWalt, S.J. Microhabitat associations of vascular epiphytes in a wet tropical forest canopy. J. Ecol. 2015, 103, 421–430. [Google Scholar] [CrossRef]
- Eskov, A.; Zverev, A.; Abakumov, E. Microbiomes in Suspended Soils of Vascular Epiphytes Differ from Terrestrial Soil Microbiomes and from Each Other. Microorganisms 2021, 9, 1033. [Google Scholar] [CrossRef] [PubMed]
- Rascher, U.; Freiberg, M.; Lüttge, U. Functional Diversity of Photosynthetic Light Use of 16 Vascular Epiphyte Species Under Fluctuating Irradiance in the Canopy of a Giant Virola michelii (Myristicaceae) Tree in the Tropical Lowland Forest of French Guyana. Front. Plant Sci. 2012, 2, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardow, A.; Gehrig-Downie, C.; Gradstein, R.; Lakatos, M. Functional diversity of epiphytes in two tropical lowland rain-forest, French Guaiana: Using bryophyte life-forms to detect areas of high biodiversity. Biodivers. Conserv. 2012, 21, 3637–3655. [Google Scholar] [CrossRef]
- Ruiz-Cordova, J.P.; Toledo-Hernández, V.H.; Flores-Palacios, A. The effect of substrate abundance in the vertical stratification of bromeliad epiphytes in a tropical dry forest (Mexico). Flora-Morphol. Distrib. Funct. Ecol. Plants 2014, 209, 375–384. [Google Scholar] [CrossRef]
- Ding, Y.; Liu, G.; Zang, R.; Zhang, J.; Lu, X.; Huang, J. Distribution of vascular epiphytes along a tropical elevational gradient: Disentangling abiotic and biotic determinants. Sci. Rep. 2016, 6, 19706. [Google Scholar] [CrossRef] [Green Version]
- Costa, D.S.; Zotz, G.; Hemp, A.; Kleyer, M. Trait patterns of epiphytes compared to other plant life--Forms along a tropical elevation gradient. Funct. Ecol. 2018, 32, 2073–2084. [Google Scholar] [CrossRef]
- Donald, J.; Maxfield, P.; Leroy, C.; Ellwood, M.F. Epiphytic suspended soils from Borneo and Amazonia differ in their microbial community composition. Acta Oecologica 2020, 106, 103586. [Google Scholar] [CrossRef]
- Peña-Claros, M.; Poorter, L.; Alarcón, A.; Blate, G.; Choque, U.; Fredericksen, T.S.; Justiniano, M.J.; Leaño, C.; Licona, J.C.; Pariona, W.; et al. Soil Effects on Forest Structure and Diversity in a Moist and a Dry Tropical Forest. Biotropica 2012, 44, 276–283. [Google Scholar] [CrossRef]
- Chave, J.; Riéra, B.; Dubois, M.-A. Estimation of biomass in a neotropical forest of French Guiana: Spatial and temporal variability. J. Trop. Ecol. 2001, 17, 79–96. [Google Scholar] [CrossRef] [Green Version]
- Gourlet-Fleury, S.; Guehl, J.-M.; Laroussinie, O. Ecology and Management of a Neotropical Rainforest: Lessons Drawn from Paracou, a Long-Term Experimental Research Site in French Guiana; Elseiver: Paris, France, 2004. [Google Scholar]
- Courtois, E.A.; Stahl, C.; Berge, J.V.D.; Bréchet, L.; Van Langenhove, L.; Richter, A.; Urbina, I.; Soong, J.L.; Peñuelas, J.; Janssens, I. Spatial Variation of Soil CO2, CH4 and N2O Fluxes Across Topographical Positions in Tropical Forests of the Guiana Shield. Ecosystems 2018, 21, 1445–1458. [Google Scholar] [CrossRef] [Green Version]
- Poszwa, A.; Ferry, B.; Pollie, B.; Grimaldi, C.; Charles-Dominique, P.; Loubet, M.; Dambrine, E. Variations of plant and soil 87Sr/86Sr along the slope of a tropical inselberg. Ann. For. Sci. 2009, 66, 512. [Google Scholar] [CrossRef]
- Vand der meer, P.J.; Bongers, F. Patterns of treefall and branch fall in a tropical rain forest in French Guiana. J. Ecol. 1996, 84, 19–29. [Google Scholar] [CrossRef]
- Réjou-Méchain, M.; Tymen, B.; Blanc, L.; Fauset, S.; Feldpausch, T.R.; Monteagudo, A.; Phillips, O.; Richard, H.; Chave, J. Using repeated small-footprint LiDAR acquisitions to infer spatial and temporal variations of a high-biomass Neotropical forest. Remote Sens. Environ. 2015, 169, 93–101. [Google Scholar] [CrossRef]
- FAO-ISRIC-ISSS. World Reference Base for Soil Resources; World Soil Resources Report Nr. 84; FAO: Rome, Italy, 1998. [Google Scholar]
- Minh, D.H.T.; Le Toan, T.; Rocca, F.; Tebaldini, S.; Villard, L.; Réjou-Méchain, M.; Phillips, O.L.; Feldpausch, T.R.; Dubois-Fernandez, P.; Scipal, K.; et al. SAR tomography for the retrieval of forest biomass and height: Cross-validation at two tropical forest sites in French Guaiana. Remote Sens. Environ. 2016, 175, 138–147. [Google Scholar] [CrossRef] [Green Version]
- Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Rivas-Ubach, A.; Oravec, M.; Vecerova, K.; Urban, O.; Jentsch, A.; Kreyling, J.; Beierkuhnlein, C.; et al. Opposite metabolic responses of shoots and roots to drought. Sci. Rep. 2015, 4, 6829. [Google Scholar] [CrossRef] [Green Version]
- Gargallo-Garriga, A.; Sardans, J.; Llusià, J.; Peguero, G.; Asensio, D.; Ogaya, R.; Urbina, I.; Van Langenhove, L.; Verryckt, L.T.; Courtois, E.A.; et al. 31P-NMR Metabolomics Revealed Species-Specific Use of Phosphorous in Trees of a French Guiana Rainforest. Molecules 2020, 25, 3960. [Google Scholar] [CrossRef]
- Koch, B.P.; Dittmar, T.; Witt, M.; Kattner, G. Fundamentals of Molecular Formula Assignment to Ultrahigh Resolution Mass Data of Natural Organic Matter. Anal. Chem. 2007, 79, 1758–1763. [Google Scholar] [CrossRef]
- Kellerman, A.M.; Dittmar, T.; Kothawala, D.N.; Tranvik, L.J. Chemodiversity of dissolved organic matter in lakes driven by climate and hydrology. Nat. Commun. 2014, 5, 3804. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Stevens, M.H.H.; Oksanen, M.J. MASS Suggests. The vegan package. Community Ecol. Package 2007, 10, 631–637. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA + for PRI-MER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| df | Sum of Squares | Mean Square | F | R2 | P | |
|---|---|---|---|---|---|---|
| Soil type | 2 | 2.33 | 1.17 | 8.17 | 0.196 | 0.001 |
| Tree species | 12 | 2.61 | 0.22 | 1.52 | 0.219 | 0.001 |
| Soil type × Tree species | 13 | 1.83 | 0.14 | 0.98 | 0.153 | 0.56 |
| Residuals | 36 | 5.14 | 0.14 | 0.43 | 36 | |
| Total | 63 | 11.9 | 1 |
| df | Sum of Squares | Mean Square | F | R2 | P | |
|---|---|---|---|---|---|---|
| Epiphyte | 4 | 3.47 | 0.50 | 4.00 | 0.291 | 0.001 |
| Tree species | 12 | 2.56 | 0.21 | 1.72 | 0.215 | 0.001 |
| Epiphyte × Tree species | 20 | 2.91 | 0.15 | 1.17 | 0.244 | 0.031 |
| Residuals | 24 | 2.98 | 0.12 | 0.25 | ||
| Total | 63 | 11.9 | 1 |
| Tree Species | DBH | Epiphytes Collected | Epi Family | Epi Genus | Species |
|---|---|---|---|---|---|
| Aspidosperma sprucaneaum Benth. Ex Müll.Arg. | 97.1 | 1 | Bromeliaceae | Achmea | A. aquilega |
| Couratari oblongifolia Ducke & R. Knuth | 79.1 | 1 | Araceae | Unknown | Philodendron sp. |
| Eschweilera coriaceae (DC.) S.A.Mori | 53.0 | 0 | Araceae | Unknown | Philodendron sp. |
| Malvaceae Sterculia pruriens | 53.6 | 3 | Aspleniaceae | Asplenium | Asplenium sp. |
| Bromeliaceae | Unknown | M. pleiosticha | |||
| Araceae | Philodendron | Philodendron sp. | |||
| Annonaceae Oxandra asbeckii | 53.2 | 2 | Bromeliaceae | Mezobromelia | M. pleiosticha |
| Orchidaceae | Unknown | Orchidaceae sp. | |||
| Lecythidaceae Gustavia hexapetala | 48.7 | 1 | Bromeliaceae | Achmea | A. aquilega |
| Chrysobalanaceae Licania alba | 54.7 | 3 | Aspleniaceae | Asplenium | Asplenium sp. 1 |
| Araceae | Philodendron | Philodendron sp. | |||
| Bromeliaceae | Achmea | A. aquilega | |||
| Eperua falcata Aubl. | 81.8 | 3 | Aspleniaceae | Asplenium | Asplenium sp. |
| Orchidaceae | Unknown | Orchidaceae sp. 2 | |||
| Araceae | Philodendron | Philodendron sp. 2 | |||
| Moraceae Brosimum guianense | 73.5 | 2 | Aspleniaceae | Asplenium | Asplenium sp. 1 |
| Orchidaceae | Unknown | Orchidaceae sp. | |||
| Lecythis persistens Sagot | 47.4 | 3 | Araceae | Anthurium | Anthurium sp. 2 |
| Bromeliaceae | Unknown | M. pleiosticha | |||
| Orchidaceae | Unknown | Orchidaceae sp. | |||
| Micropholis sp. | 69.6 | 2 | Aspleniaceae | Asplenium | Asplenium sp. |
| Bromeliaceae | Achmea | A. aquilega | |||
| Poraqueiba guianensis Aubl. | 83.4 | 3 | Orchidaceae | Unknown | Orchidaceae sp. |
| Araceae | Philodendron | Philodendron sp. | |||
| Araceae | Philodendron | Philodendron sp. | |||
| Tetragastris sp. | 47.4 | 2 | Araceae | Philodendron | Philodendron sp. |
| Araceae | Philodendron | Philodendron sp. | |||
| Tetragastris altissima (Aubl.) Swart | 95.5 | 3 | Araceae | Anthurium | Anthurium sp. |
| Aspleniaceae | Asplenium | Asplenium sp. 1 | |||
| Bromeliaceae | Unknown | M. pleiosticha |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gargallo-Garriga, A.; Sardans, J.; Alrefaei, A.F.; Klem, K.; Fuchslueger, L.; Ramírez-Rojas, I.; Donald, J.; Leroy, C.; Langenhove, L.V.; Verbruggen, E.; et al. Tree Species and Epiphyte Taxa Determine the “Metabolomic niche” of Canopy Suspended Soils in a Species-Rich Lowland Tropical Rainforest. Metabolites 2021, 11, 718. https://doi.org/10.3390/metabo11110718
Gargallo-Garriga A, Sardans J, Alrefaei AF, Klem K, Fuchslueger L, Ramírez-Rojas I, Donald J, Leroy C, Langenhove LV, Verbruggen E, et al. Tree Species and Epiphyte Taxa Determine the “Metabolomic niche” of Canopy Suspended Soils in a Species-Rich Lowland Tropical Rainforest. Metabolites. 2021; 11(11):718. https://doi.org/10.3390/metabo11110718
Chicago/Turabian StyleGargallo-Garriga, Albert, Jordi Sardans, Abdulwahed Fahad Alrefaei, Karel Klem, Lucia Fuchslueger, Irene Ramírez-Rojas, Julian Donald, Celine Leroy, Leandro Van Langenhove, Erik Verbruggen, and et al. 2021. "Tree Species and Epiphyte Taxa Determine the “Metabolomic niche” of Canopy Suspended Soils in a Species-Rich Lowland Tropical Rainforest" Metabolites 11, no. 11: 718. https://doi.org/10.3390/metabo11110718