MEATabolomics: Muscle and Meat Metabolomics in Domestic Animals



Abstract
:1. Introduction
2. MEATabolomic Methodologies and Approaches
3. Metabolomes Associated with Meat Quality Traits
3.1. Meat Color, WHC, and pH Decline
3.2. Flavor and Palatability
3.3. Chicken Meat Quality Traits
4. Factors Affecting Muscle and Meat Metabolomes
4.1. Animal Species, Breeds, and Genetic Backgrounds
4.2. Animal Feeding
4.3. Muscle Type
4.4. Postmortem Aging and Storage
4.5. Processing
4.6. Spoilage
4.7. Authentication
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rubert, J.; Zachariasova, M.; Hajslova, J. Advances in high-resolution mass spectrometry based on metabolomics studies for food—A review. Food Addit. Contam. Part. A 2015, 32, 1685–1708. [Google Scholar] [CrossRef] [PubMed]
- Consonni, R.; Cagliani, L.R. The potentiality of NMR-based metabolomics in food science and food authentication assessment. Magn. Reson. Chem. 2019, 57, 558–578. [Google Scholar] [CrossRef] [PubMed]
- García-Cañas, V.; Simó, C.; Herrero, M.; Ibáñez, E.; Cifuentes, A. Present and Future Challenges in Food Analysis: Foodomics. Anal. Chem. 2012, 84, 10150–10159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junot, C.; Fenaille, F.; Colsch, B.; Bécher, F. High resolution mass spectrometry based techniques at the crossroads of metabolic pathways. Mass Spectrom. Rev. 2014, 33, 471–500. [Google Scholar] [CrossRef]
- Sugimoto, M.; Kawakami, M.; Robert, M.; Soga, T.; Tomita, M. Bioinformatics Tools for Mass Spectroscopy-Based Metabolomic Data Processing and Analysis. Curr. Bioinform. 2012, 7, 96–108. [Google Scholar] [CrossRef]
- Straadt, I.K.; Aaslyng, M.D.; Bertram, H.C. An NMR-based metabolomics study of pork from different crossbreeds and relation to sensory perception. Meat Sci. 2014, 96, 719–728. [Google Scholar] [CrossRef]
- Lytou, A.E.; Nychas, G.E.; Panagou, E.Z. Effect of pomegranate based marinades on the microbiological, chemical and sensory quality of chicken meat: A metabolomics approach. Int. J. Food Microbiol. 2018, 267, 42–53. [Google Scholar] [CrossRef]
- Lee, S.M.; Kwon, G.Y.; Kim, K.O.; Kim, Y.S. Metabolomic approach for determination of key volatile compounds related to beef flavor in glutathione-Maillard reaction products. Anal. Chim. Acta 2011, 703, 204–211. [Google Scholar] [CrossRef]
- Adamski, J.; Suhre, K. Metabolomics platforms for genome wide association studies--linking the genome to the metabolome. Curr. Opin. Biotechnol. 2013, 24, 39–47. [Google Scholar] [CrossRef]
- Fontanesi, L. Metabolomics and livestock genomics: Insights into a phenotyping frontier and its applications in animal breeding. Anim. Front. 2016, 6, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Berri, C.; Picard, B.; Lebret, B.; Andueza, D.; Lefevre, F.; Le Bihan-Duval, E.; Beauclercq, S.; Chartrin, P.; Vautier, A.; Legrand, I.; et al. Predicting the Quality of Meat: Myth or Reality? Foods 2019, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zampiga, M.; Flees, J.; Meluzzi, A.; Dridi, S.; Sirri, F. Application of omics technologies for a deeper insight into quali-quantitative production traits in broiler chickens: A review. J. Anim. Sci. Biotechnol. 2018, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Picard, B.; Lebret, B.; Cassar-Malek, I.; Liaubet, L.; Berri, C.; Le Bihan-Duval, E.; Hocquette, J.F.; Renand, G. Recent advances in omic technologies for meat quality management. Meat Sci. 2015, 109, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Muroya, S.; Oe, M.; Nakajima, I.; Ojima, K.; Chikuni, K. CE-TOF MS-based metabolomic profiling revealed characteristic metabolic pathways in postmortem porcine fast and slow type muscles. Meat Sci. 2014, 98, 726–735. [Google Scholar] [CrossRef]
- Ma, D.; Kim, Y.H.B.; Cooper, B.; Oh, J.H.; Chun, H.; Choe, J.H.; Schoonmaker, J.P.; Ajuwon, K.; Min, B. Metabolomics Profiling to Determine the Effect of Postmortem Aging on Color and Lipid Oxidative Stabilities of Different Bovine Muscles. J. Agric. Food Chem. 2017, 65, 6708–6716. [Google Scholar] [CrossRef]
- Yu, Q.; Tian, X.; Shao, L.; Li, X.; Dai, R. Targeted metabolomics to reveal muscle-specific energy metabolism between bovine longissimus lumborum and psoas major during early postmortem periods. Meat Sci. 2019, 156, 166–173. [Google Scholar] [CrossRef]
- Muroya, S.; Oe, M.; Ojima, K.; Watanabe, A. Metabolomic approach to key metabolites characterizing postmortem aged loin muscle of Japanese Black (Wagyu) cattle. Asia Australas. J. Anim. Sci. 2019, 32, 1172–1185. [Google Scholar] [CrossRef] [Green Version]
- Kodani, Y.; Miyakawa, T.; Komatsu, T.; Tanokura, M. NMR-based metabolomics for simultaneously evaluating multiple determinants of primary beef quality in Japanese Black cattle. Sci. Rep. 2017, 7, 1297. [Google Scholar] [CrossRef] [Green Version]
- Graham, S.F.; Kennedy, T.; Chevallier, O.; Gordon, A.; Farmer, L.; Elliott, C.; Moss, B. The application of NMR to study changes in polar metabolite concentrations in beef longissimus dorsi stored for different periods post mortem. Metabolomics 2010, 6, 395–404. [Google Scholar] [CrossRef]
- Carrillo, J.A.; He, Y.; Li, Y.; Liu, J.; Erdman, R.A.; Sonstegard, T.S.; Song, J. Integrated metabolomic and transcriptome analyses reveal finishing forage affects metabolic pathways related to beef quality and animal welfare. Sci. Rep. 2016, 6, 25948. [Google Scholar] [CrossRef]
- Yu, K.; Matzapetakis, M.; Valent, D.; Saco, Y.; De Almeida, A.M.; Terre, M.; Bassols, A. Skeletal muscle metabolomics and blood biochemistry analysis reveal metabolic changes associated with dietary amino acid supplementation in dairy calves. Sci. Rep. 2018, 8, 13850. [Google Scholar] [CrossRef] [PubMed]
- De Zawadzki, A.; Arrivetti, L.O.R.; Vidal, M.P.; Catai, J.R.; Nassu, R.T.; Tullio, R.R.; Berndt, A.; Oliveira, C.R.; Ferreira, A.G.; Neves-Junior, L.F.; et al. Mate extract as feed additive for improvement of beef quality. Food Res. Int. 2017, 99, 336–347. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Fu, Y.; Han, X.; Li, X.; Li, C. Metabolomic investigation of porcine muscle and fatty tissue after Clenbuterol treatment using gas chromatography/mass spectrometry. J. Chromatogr. A 2016, 1456, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, G.; Kobayashi, H.; Shibata, M.; Kubota, M.; Kadowaki, M.; Fujimura, S. Regulation of free glutamate content in meat by dietary lysine in broilers. Anim. Sci. J. 2015, 86, 435–442. [Google Scholar] [CrossRef]
- Liu, C.; Pan, D.; Ye, Y.; Cao, J. 1H NMR and multivariate data analysis of the relationship between the age and quality of duck meat. Food Chem. 2013, 141, 1281–1286. [Google Scholar] [CrossRef]
- Guitton, Y.; Dervilly-Pinel, G.; Jandova, R.; Stead, S.; Takats, Z.; Le Bizec, B. Rapid evaporative ionisation mass spectrometry and chemometrics for high-throughput screening of growth promoters in meat producing animals. Food Addit. Contam. Part A 2018, 35, 900–910. [Google Scholar] [CrossRef]
- Sugimoto, M.; Obiya, S.; Kaneko, M.; Enomoto, A.; Honma, M.; Wakayama, M.; Soga, T.; Tomita, M. Metabolomic Profiling as a Possible Reverse Engineering Tool for Estimating Processing Conditions of Dry-Cured Hams. J. Agric. Food Chem. 2017, 65, 402–410. [Google Scholar] [CrossRef]
- Zhang, J.; Ye, Y.; Sun, Y.; Pan, D.; Ou, C.; Dang, Y.; Wang, Y.; Cao, J.; Wang, D. 1H NMR and multivariate data analysis of the differences of metabolites in five types of dry-cured hams. Food Res. Int. 2018, 113, 140–148. [Google Scholar] [CrossRef]
- Lytou, A.E.; Panagou, E.Z.; Nychas, G.E. Effect of different marinating conditions on the evolution of spoilage microbiota and metabolomic profile of chicken breast fillets. Food Microbiol. 2017, 66, 141–149. [Google Scholar] [CrossRef]
- Yang, Y.; Ye, Y.; Pan, D.; Sun, Y.; Wang, Y.; Cao, J. Metabonomics profiling of marinated meat in soy sauce during processing. J. Sci. Food Agric. 2018, 98, 1325–1331. [Google Scholar] [CrossRef]
- Huff-Lonergan, E.; Zhang, W.; Lonergan, S.M. Biochemistry of postmortem muscle—Lessons on mechanisms of meat tenderization. Meat Sci. 2010, 86, 184–195. [Google Scholar] [CrossRef]
- Ramautar, R.; Somsen, G.W.; De Jong, G.J. CE-MS for metabolomics: Developments and applications in the period 2016–2018. Electrophoresis 2019, 40, 165–179. [Google Scholar] [CrossRef] [Green Version]
- Stolz, A.; Jooss, K.; Hocker, O.; Romer, J.; Schlecht, J.; Neususs, C. Recent advances in capillary electrophoresis-mass spectrometry: Instrumentation, methodology and applications. Electrophoresis 2019, 40, 79–112. [Google Scholar] [CrossRef] [Green Version]
- Welzenbach, J.; Neuhoff, C.; Looft, C.; Schellander, K.; Tholen, E.; Grosse-Brinkhaus, C. Different Statistical Approaches to Investigate Porcine Muscle Metabolome Profiles to Highlight New Biomarkers for Pork Quality Assessment. PLoS ONE 2016, 11, e0149758. [Google Scholar] [CrossRef] [PubMed]
- Welzenbach, J.; Neuhoff, C.; Heidt, H.; Cinar, M.U.; Looft, C.; Schellander, K.; Tholen, E.; Grosse-Brinkhaus, C. Integrative Analysis of Metabolomic, Proteomic and Genomic Data to Reveal Functional Pathways and Candidate Genes for Drip Loss in Pigs. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gredell, D.A.; Schroeder, A.R.; Belk, K.E.; Broeckling, C.D.; Heuberger, A.L.; Kim, S.-Y.; King, D.A.; Shackelford, S.D.; Sharp, J.L.; Wheeler, T.L.; et al. Comparison of Machine Learning Algorithms for Predictive Modeling of Beef Attributes Using Rapid Evaporative Ionization Mass Spectrometry (REIMS) Data. Sci. Rep. 2019, 9, 5721. [Google Scholar] [CrossRef] [Green Version]
- Cameron, S.J.S.; Alexander, J.L.; Bolt, F.; Burke, A.; Ashrafian, H.; Teare, J.; Marchesi, J.R.; Kinross, J.; Li, J.V.; Takáts, Z. Evaluation of Direct from Sample Metabolomics of Human Feces Using Rapid Evaporative Ionization Mass Spectrometry. Anal. Chem. 2019, 91, 13448–13457. [Google Scholar] [CrossRef] [PubMed]
- Balog, J.; Sasi-Szabó, L.; Kinross, J.; Lewis, M.R.; Muirhead, L.J.; Veselkov, K.; Mirnezami, R.; Dezső, B.; Damjanovich, L.; Darzi, A.; et al. Intraoperative tissue identification using rapid evaporative ionization mass spectrometry. Sci. Transl. Med. 2013, 5, 194ra193. [Google Scholar] [CrossRef] [PubMed]
- Ritota, M.; Casciani, L.; Failla, S.; Valentini, M. HRMAS-NMR spectroscopy and multivariate analysis meat characterisation. Meat Sci. 2012, 92, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Subbaraj, A.K.; Kim, Y.H.; Fraser, K.; Farouk, M.M. A hydrophilic interaction liquid chromatography-mass spectrometry (HILIC-MS) based metabolomics study on colour stability of ovine meat. Meat Sci. 2016, 117, 163–172. [Google Scholar] [CrossRef]
- Beauclercq, S.; Nadal-Desbarats, L.; Hennequet-Antier, C.; Collin, A.; Tesseraud, S.; Bourin, M.; Le Bihan-Duval, E.; Berri, C. Serum and Muscle Metabolomics for the Prediction of Ultimate pH, a Key Factor for Chicken-Meat Quality. J. Proteom. Res. 2016, 15, 1168–1178. [Google Scholar] [CrossRef] [PubMed]
- Takakura, Y.; Sakamoto, T.; Hirai, S.; Masuzawa, T.; Wakabayashi, H.; Nishimura, T. Characterization of the key aroma compounds in beef extract using aroma extract dilution analysis. Meat Sci. 2014, 97, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, A.; Kamada, G.; Imanari, M.; Shiba, N.; Yonai, M.; Muramoto, T. Effect of aging on volatile compounds in cooked beef. Meat Sci. 2015, 107, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Ueda, S.; Iwamoto, E.; Kato, Y.; Shinohara, M.; Shirai, Y.; Yamanoue, M. Comparative metabolomics of Japanese Black cattle beef and other meats using gas chromatography-mass spectrometry. Biosci. Biotechnol. Biochem. 2018. [Google Scholar] [CrossRef]
- Xiao, Z.; Ge, C.; Zhou, G.; Zhang, W.; Liao, G. 1H NMR-based metabolic characterization of Chinese Wuding chicken meat. Food Chem. 2019, 274, 574–582. [Google Scholar] [CrossRef]
- Sundekilde, U.K.; Rasmussen, M.K.; Young, J.F.; Bertram, H.C. High resolution magic angle spinning NMR spectroscopy reveals that pectoralis muscle dystrophy in chicken is associated with reduced muscle content of anserine and carnosine. Food Chem. 2017, 217, 151–154. [Google Scholar] [CrossRef]
- Abasht, B.; Mutryn, M.F.; Michalek, R.D.; Lee, W.R. Oxidative Stress and Metabolic Perturbations in Wooden Breast Disorder in Chickens. PLoS ONE 2016, 11, e0153750. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yang, Y.; Pan, D.; He, J.; Cao, J.; Wang, H.; Ertbjerg, P. Metabolite profile based on 1H NMR of broiler chicken breasts affected by wooden breast myodegeneration. Food Chem. 2020, 310, 125852. [Google Scholar] [CrossRef]
- Xing, T.; Zhao, X.; Xu, X.; Li, J.; Zhang, L.; Gao, F. Physiochemical properties, protein and metabolite profiles of muscle exudate of chicken meat affected by wooden breast myopathy. Food Chem. 2020, 316, 126271. [Google Scholar] [CrossRef]
- Boerboom, G.; Van Kempen, T.; Navarro-Villa, A.; Perez-Bonilla, A. Unraveling the cause of white striping in broilers using metabolomics. Poult. Sci. 2018, 97, 3977–3986. [Google Scholar] [CrossRef]
- Consolo, N.R.B.; Silva, J.D.; Buarque, V.L.M.; Higuera-Padilla, A.; Barbosa, L.; Zawadzki, A.; Colnago, L.A.; Netto, A.S.; Gerrard, D.E.; Silva, S.L. Selection for Growth and Precocity Alters Muscle Metabolism in Nellore Cattle. Metabolites 2020, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakuma, H.; Saito, K.; Kohira, K.; Ohhashi, F.; Shoji, N.; Uemoto, Y. Estimates of genetic parameters for chemical traits of meat quality in Japanese black cattle. Anim. Sci. J. 2017, 88, 203–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsu, T.; Komatsu, M.; Uemoto, Y. The NT5E gene variant strongly affects the degradation rate of inosine 5′-monophosphate under postmortem conditions in Japanese Black beef. Meat Sci. 2019, 158, 107893. [Google Scholar] [CrossRef] [PubMed]
- Muroya, S.; Oe, M.; Ojima, K. Thiamine accumulation and thiamine triphosphate decline occur in parallel with ATP exhaustion during postmortem aging of pork muscles. Meat Sci. 2018, 137, 228–234. [Google Scholar] [CrossRef]
- Lana, A.; Longo, V.; Dalmasso, A.; D’Alessandro, A.; Bottero, M.T.; Zolla, L. Omics integrating physical techniques: Aged Piedmontese meat analysis. Food Chem. 2015, 172, 731–741. [Google Scholar] [CrossRef]
- Kim, Y.H.B.; Kemp, R.; Samuelsson, L.M. Effects of dry-aging on meat quality attributes and metabolite profiles of beef loins. Meat Sci. 2016, 111, 168–176. [Google Scholar] [CrossRef]
- Warner, R.D.; Jacob, R.H.; Rosenvold, K.; Rochfort, S.; Trenerry, C.; Plozza, T.; McDonagh, M.B. Altered post-mortem metabolism identified in very fast chilled lamb M. longissimus thoracis et lumborum using metabolomic analysis. Meat Sci. 2015, 108, 155–164. [Google Scholar] [CrossRef]
- Mitacek, R.M.; Ke, Y.; Prenni, J.E.; Jadeja, R.; VanOverbeke, D.L.; Mafi, G.G.; Ramanathan, R. Mitochondrial Degeneration, Depletion of NADH, and Oxidative Stress Decrease Color Stability of Wet-Aged Beef Longissimus Steaks. J. Food Sci. 2019, 84, 38–50. [Google Scholar] [CrossRef] [Green Version]
- García-García, A.B.; Lamichhane, S.; Castejón, D.; Cambero, M.I.; Bertram, H.C. 1H HR-MAS NMR-based metabolomics analysis for dry-fermented sausage characterization. Food Chem. 2018, 240, 514–523. [Google Scholar] [CrossRef]
- Jiang, T.; Bratcher, C.L. Differentiation of commercial ground beef products and correlation between metabolites and sensory attributes: A metabolomic approach. Food Res. Int. 2016, 90, 298–306. [Google Scholar] [CrossRef]
- Suzuki, K.; Shioura, H.; Yokota, S.; Katoh, K.; Roh, S.-g.; Iida, F.; Komatsu, T.; Syoji, N.; Sakuma, H.; Yamada, S. Search for an index for the taste of Japanese Black cattle beef by panel testing and chemical composition analysis. Anim. Sci. J. 2017, 88, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Cheung, W.; Winder, C.L.; Goodacre, R. VOC-based metabolic profiling for food spoilage detection with the application to detecting Salmonella typhimurium-contaminated pork. Anal. Bioanal. Chem. 2010, 397, 2439–2449. [Google Scholar] [CrossRef]
- Xu, Y.; Cheung, W.; Winder, C.L.; Dunn, W.B.; Goodacre, R. Metabolic profiling of meat: Assessment of pork hygiene and contamination with Salmonella typhimurium. Analyst 2011, 136, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Argyri, A.A.; Doulgeraki, A.I.; Blana, V.A.; Panagou, E.Z.; Nychas, G.-J.E. Potential of a simple HPLC-based approach for the identification of the spoilage status of minced beef stored at various temperatures and packaging systems. Int. J. Food Microbiol. 2011, 150, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Argyri, A.A.; Mallouchos, A.; Panagou, E.Z.; Nychas, G.J. The dynamics of the HS/SPME-GC/MS as a tool to assess the spoilage of minced beef stored under different packaging and temperature conditions. Int. J. Food Microbiol. 2015, 193, 51–58. [Google Scholar] [CrossRef]
- Jung, Y.; Lee, J.; Kwon, J.; Lee, K.S.; Ryu, D.H.; Hwang, G.S. Discrimination of the geographical origin of beef by 1H NMR-based metabolomics. J. Agric. Food Chem. 2010, 58, 10458–10466. [Google Scholar] [CrossRef]
- Zaima, N.; Goto-Inoue, N.; Hayasaka, T.; Enomoto, H.; Setou, M. Authenticity assessment of beef origin by principal component analysis of matrix-assisted laser desorption/ionization mass spectrometric data. Anal. Bioanal. Chem. 2011, 400, 1865–1871. [Google Scholar] [CrossRef]
- Osorio, M.T.; Moloney, A.P.; Brennan, L.; Monahan, F.J. Authentication of beef production systems using a metabolomic-based approach. Animal 2012, 6, 167–172. [Google Scholar] [CrossRef]
- Trivedi, D.K.; Hollywood, K.A.; Rattray, N.J.; Ward, H.; Greenwood, J.; Ellis, D.I.; Goodacre, R. Meat, the metabolites: An integrated metabolite profiling and lipidomics approach for the detection of the adulteration of beef with pork. Analyst 2016, 141, 2155–2164. [Google Scholar] [CrossRef] [Green Version]
- Pavlidis, D.E.; Mallouchos, A.; Ercolini, D.; Panagou, E.Z.; Nychas, G.E. A volatilomics approach for off-line discrimination of minced beef and pork meat and their admixture using HS-SPME GC/MS in tandem with multivariate data analysis. Meat Sci. 2019, 151, 43–53. [Google Scholar] [CrossRef]
- Sidwick, K.L.; Johnson, A.E.; Adam, C.D.; Pereira, L.; Thompson, D.F. Use of Liquid Chromatography Quadrupole Time-of-Flight Mass Spectrometry and Metabonomic Profiling To Differentiate between Normally Slaughtered and Dead on Arrival Poultry Meat. Anal. Chem. 2017, 89, 12131–12136. [Google Scholar] [CrossRef]
- Cao, M.; Han, Q.; Zhang, J.; Zhang, R.; Wang, J.; Gu, W.; Kang, W.; Lian, K.; Ai, L. An untargeted and pseudotargeted metabolomic combination approach to identify differential markers to distinguish live from dead pork meat by liquid chromatography-mass spectrometry. J. Chromatograph. A 2020, 1610, 460553. [Google Scholar] [CrossRef] [PubMed]
- Zanardi, E.; Caligiani, A.; Padovani, E.; Mariani, M.; Ghidini, S.; Palla, G.; Ianieri, A. Detection of irradiated beef by nuclear magnetic resonance lipid profiling combined with chemometric techniques. Meat Sci. 2013, 93, 171–177. [Google Scholar] [CrossRef]
- Zanardi, E.; Caligiani, A.; Palla, L.; Mariani, M.; Ghidini, S.; Di Ciccio, P.A.; Palla, G.; Ianieri, A. Metabolic profiling by 1H NMR of ground beef irradiated at different irradiation doses. Meat Sci. 2015, 103, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Hinzman, A.A.; Kang, E.L.; Szczesniak, R.D.; Lu, L.J. Computational and statistical analysis of metabolomics data. Metabolomics 2015, 11, 1492–1513. [Google Scholar] [CrossRef]
- Hollung, K.; Timperio, A.M.; Olivan, M.; Kemp, C.; Coto-Montes, A.; Sierra, V.; Zolla, L. Systems biology: A new tool for farm animal science. Curr. Protein Pept. Sci. 2014, 15, 100–117. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, C.E.; Cornforth, D.P.; Whittier, D. Consumer preferences for beef color and packaging did not affect eating satisfaction. Meat Sci. 2001, 57, 359–363. [Google Scholar] [CrossRef]
- Arihara, K.; Yokoyama, I.; Ohata, M. DMHF (2,5-dimethyl-4-hydroxy-3(2H)-furanone), a volatile food component with attractive sensory properties, brings physiological functions through inhalation. Adv. Food Nutr. Res. 2019, 89, 239–258. [Google Scholar] [PubMed]
- Nishimura, T. Mechanism Involved in the Improvement of Meat Taste during Postmortem Aging. Food Sci. Technol. Int. Tokyo 1998, 4, 241–249. [Google Scholar] [CrossRef] [Green Version]
- Mottram, D.S. Flavour formation in meat and meat products: A review. Food Chem. 1998, 62, 415–424. [Google Scholar] [CrossRef]
- Kuttappan, V.A.; Hargis, B.M.; Owens, C.M. White striping and woody breast myopathies in the modern poultry industry: A review. Poult. Sci. 2016, 95, 2724–2733. [Google Scholar] [CrossRef] [PubMed]
- Andersen, H.J.; Oksbjerg, N.; Young, J.F.; Therkildsen, M. Feeding and meat quality—A future approach. Meat Sci. 2005, 70, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Xing, T.; Gao, F.; Tume, R.K.; Zhou, G.; Xu, X. Stress Effects on Meat Quality: A Mechanistic Perspective. Compr. Rev. Food Sci. Food Saf. 2019, 18, 380–401. [Google Scholar] [CrossRef] [Green Version]
- Hwang, Y.-H.; Kim, G.-D.; Jeong, J.-Y.; Hur, S.-J.; Joo, S.-T. The relationship between muscle fiber characteristics and meat quality traits of highly marbled Hanwoo (Korean native cattle) steers. Meat Sci. 2010, 86, 456–461. [Google Scholar] [CrossRef]
- Chang, K.C.; Da Costa, N.; Blackley, R.; Southwood, O.; Evans, G.; Plastow, G.; Wood, J.D.; Richardson, R.I. Relationships of myosin heavy chain fibre types to meat quality traits in traditional and modern pigs. Meat Sci. 2003, 64, 93–103. [Google Scholar] [CrossRef]
- Lee, S.H.; Choe, J.H.; Choi, Y.M.; Jung, K.C.; Rhee, M.S.; Hong, K.C.; Lee, S.K.; Ryu, Y.C.; Kim, B.C. The influence of pork quality traits and muscle fiber characteristics on the eating quality of pork from various breeds. Meat Sci. 2012, 90, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Nam, Y.J.; Choi, Y.M.; Lee, S.H.; Choe, J.H.; Jeong, D.W.; Kim, Y.Y.; Kim, B.C. Sensory evaluations of porcine longissimus dorsi muscle: Relationships with postmortem meat quality traits and muscle fiber characteristics. Meat Sci. 2009, 83, 731–736. [Google Scholar] [CrossRef]
- Lefaucheur, L. A second look into fibre typing--relation to meat quality. Meat Sci. 2010, 84, 257–270. [Google Scholar] [CrossRef]
- Schiaffino, S.; Reggiani, C. Fiber types in mammalian skeletal muscles. Physiol. Rev. 2011, 91, 1447–1531. [Google Scholar] [CrossRef] [Green Version]
- Eggert, J.M.; Depreux, F.F.S.; Schinckel, A.P.; Grant, A.L.; Gerrard, D.E. Myosin heavy chain isoforms account for variation in pork quality. Meat Sci. 2002, 61, 117–126. [Google Scholar] [CrossRef]
- Oe, M.; Ohnishi-Kameyama, M.; Nakajima, I.; Muroya, S.; Shibata, M.; Ojima, K.; Kushibiki, S.; Chikuni, K. Proteome analysis of whole and water-soluble proteins in masseter and semitendinosus muscles of Holstein cows. Anim. Sci. J. 2011, 82, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Hatazawa, Y.; Senoo, N.; Tadaishi, M.; Ogawa, Y.; Ezaki, O.; Kamei, Y.; Miura, S. Metabolomic Analysis of the Skeletal Muscle of Mice Overexpressing PGC-1α. PLoS ONE 2015, 10, e0129084. [Google Scholar] [CrossRef] [PubMed]
- Bourdeau Julien, I.; Sephton, C.F.; Dutchak, P.A. Metabolic Networks Influencing Skeletal Muscle Fiber Composition. Front. Cell Dev. Biol. 2018, 6, 125. [Google Scholar] [CrossRef]
- Muroya, S.; Nakajima, I.; Oe, M.; Chikuni, K. Difference in postmortem degradation pattern among troponin T isoforms expressed in bovine longissimus, diaphragm, and masseter muscles. Meat Sci. 2006, 72, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Muroya, S.; Ertbjerg, P.; Pomponio, L.; Christensen, M. Desmin and troponin T are degraded faster in type IIb muscle fibers than in type I fibers during postmortem aging of porcine muscle. Meat Sci. 2010, 86, 764–769. [Google Scholar] [CrossRef]
- Kitamura, S.-I.; Muroya, S.; Tanabe, S.; Okumura, T.; Chikuni, K.; Nishimura, T. Mechanism of production of troponin T fragments during postmortem aging of porcine muscle. J. Agric. Food Chem. 2005, 53, 4178–4181. [Google Scholar] [CrossRef] [PubMed]
- Muroya, S.; Kitamura, S.-I.; Tanabe, S.; Nishimura, T.; Nakajima, I.; Chikuni, K. N-terminal amino acid sequences of troponin T fragments, including 30 kDa one, produced during postmortem aging of bovine longissimus muscle. Meat Sci. 2004, 67, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Muroya, S.; Ohnishi-Kameyama, M.; Oe, M.; Nakajima, I.; Chikuni, K. Postmortem changes in bovine troponin T isoforms on two-dimensional electrophoretic gel analyzed using mass spectrometry and western blotting: The limited fragmentation into basic polypeptides. Meat Sci. 2007, 75, 506–514. [Google Scholar] [CrossRef]
- Lametsch, R.; Karlsson, A.; Rosenvold, K.; Andersen, H.J.; Roepstorff, P.; Bendixen, E. Postmortem proteome changes of porcine muscle related to tenderness. J. Agric. Food Chem. 2003, 51, 6992–6997. [Google Scholar] [CrossRef]
- Marino, R.; Albenzio, M.; Della Malva, A.; Santillo, A.; Loizzo, P.; Sevi, A. Proteolytic pattern of myofibrillar protein and meat tenderness as affected by breed and aging time. Meat Sci. 2013, 95, 281–287. [Google Scholar] [CrossRef]
- Della Malva, A.; De Palo, P.; Lorenzo, J.M.; Maggiolino, A.; Albenzio, M.; Marino, R. Application of proteomic to investigate the post-mortem tenderization rate of different horse muscles. Meat Sci. 2019, 157, 107885. [Google Scholar] [CrossRef] [PubMed]
- López-Pedrouso, M.; Pérez-Santaescolástica, C.; Franco, D.; Fulladosa, E.; Carballo, J.; Zapata, C.; Lorenzo, J.M. Comparative proteomic profiling of myofibrillar proteins in dry-cured ham with different proteolysis indices and adhesiveness. Food Chem. 2018, 244, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Nychas, G.-J.E.; Skandamis, P.N.; Tassou, C.C.; Koutsoumanis, K.P. Meat spoilage during distribution. Meat Sci. 2008, 78, 77–89. [Google Scholar] [CrossRef] [PubMed]



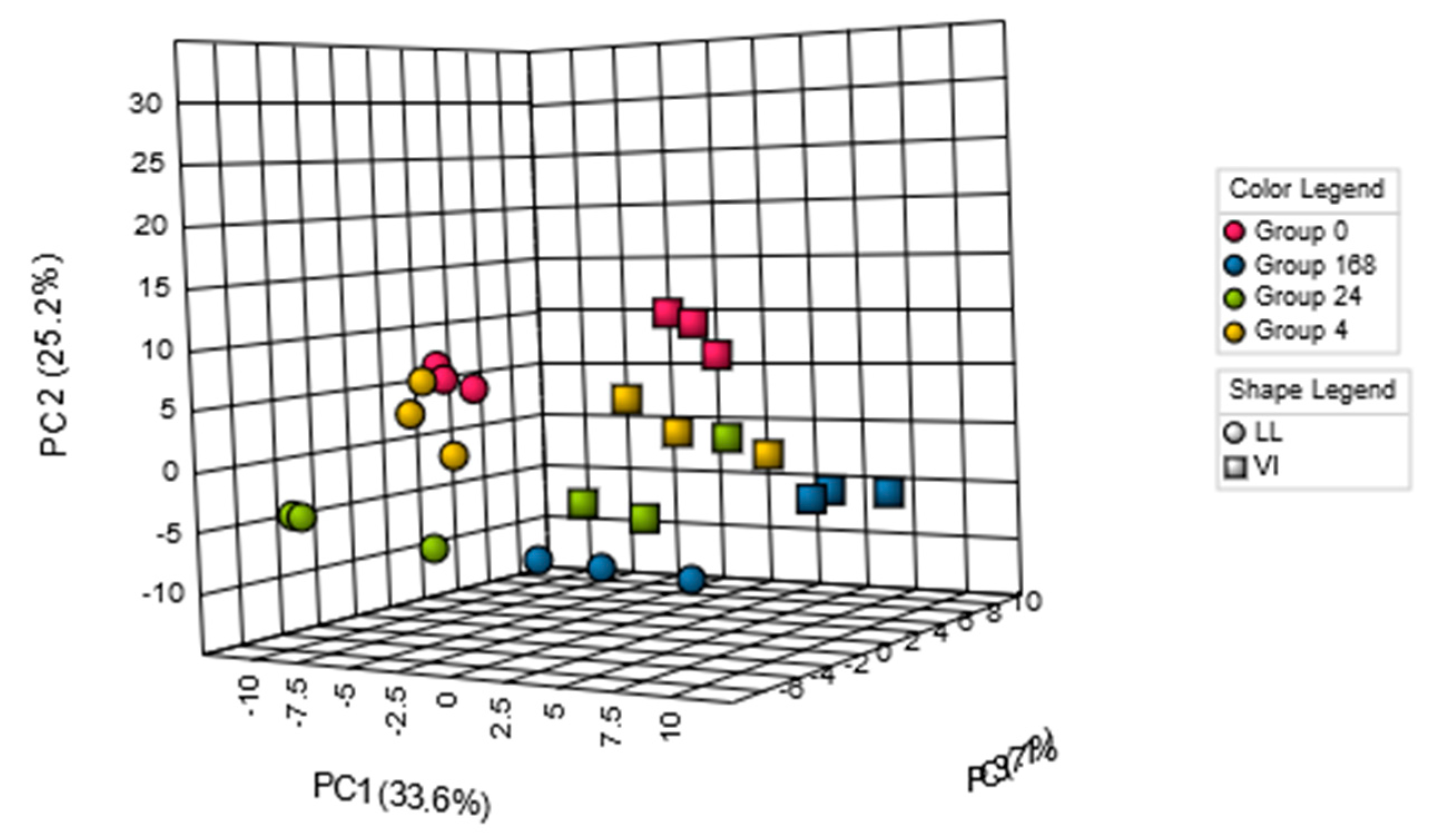{kind=link}
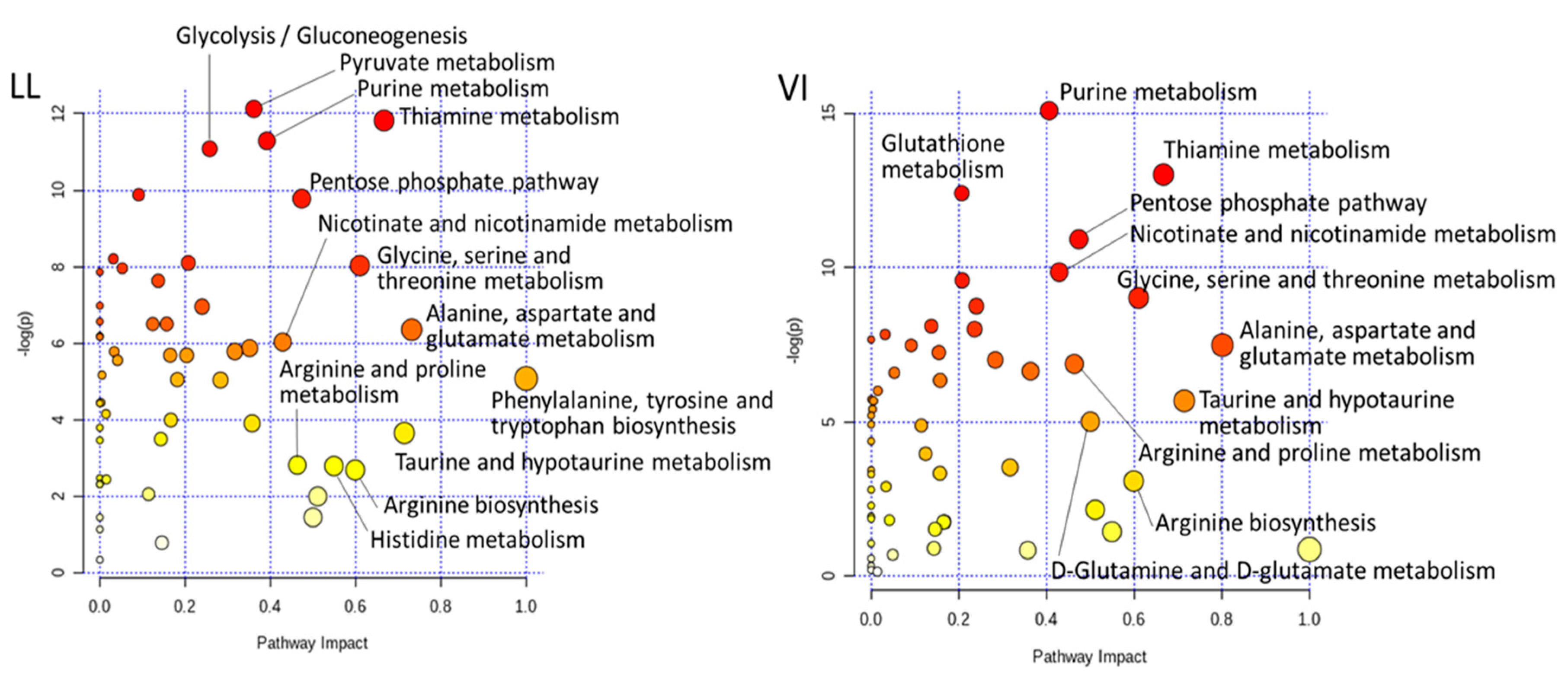{kind=link}
| CE | GC | LC | |
|---|---|---|---|
| Favorable target metabolites | Polar, charged | Mainly volatile | Non-polar, neutral |
| Sample derivatization | Unnecessary | Required for non-volatile compounds | Unnecessary |
| Number of theoretical plate | 105 ~ 106 | 104 ~ 105 | 104 |
| Separation of structural isomer | High | High | Low |
| Running time | >20 min | 20–40 min | 15–40 min |
| Downstream ionization | ESI | EI, CI | ESI, APCI |
| Category of Objective | Species/Meat Type | Factors Analyzed | Methodology | Multivariate Data Analysis | Ref. | Authors |
|---|---|---|---|---|---|---|
| Meat characterization | Cattle | Muscle type | HR–MAS 1H–NMR | PLS–DA, OPLS–DA | [39] | Ritota et al. |
| Lamb | Storage time, display time, packaging conditions | HILIC–MS | PCA | [40] | Subbaraj et al. | |
| Chicken | pHu | 1H–NMR | OPLS–DA, MSEA | [41] | Beauclercq et al. | |
| Beef | Flavor | GC–MS | - | [42] | Takakura et al. | |
| Beef | Flavor, aging period | HS/SPME GC–MS | - | [43] | Watanabe et al. | |
| Beef, pork, chicken | Flavor, species, breeds, tissues | GC–MS | OPLS–DA | [44] | Ueda et al. | |
| Chicken | Age of chicken, muscle type | 1H–NMR | PLS–DA | [45] | Xiao et al. | |
| Meat abnormality | Chicken | Dystrophy of breast | HR–MAS 1H–NMR | PCA, OPLS–DA | [46] | Sundekilde et al. |
| Chicken | Wooden breast | GC–MS, LC–MS/MS | RF | [47] | Abasht et al. | |
| Chicken | Wooden breast | 1H–NMR | OPLS–DA | [48] | Wang et al. | |
| Chicken | Wooden breast | 1H–NMR | OPLS–DA | [49] | Xing et al. | |
| Chicken | White striping | GC–MS, LC–MS | PCA, Pathway | [50] | Boerboom et al. | |
| Genetic background | Pig | Crossbreeds | 1H–NMR | PLS | [6] | Straadt et al. |
| Pig | Drip loss, association with SNP | GC–MS, LC–MS | Pathway, GWAS | [35] | Welzenbach et al. | |
| Cattle | Genetic parameters for growth and precocity | 1H–NMR | PLS–DA | [51] | Consolo et al. | |
| Cattle | Genetic parameters for chemical traits | GC, LC | - | [52] | Sakuma et al. | |
| Cattle | NT5E genotype | GC, LC | - | [53] | Komatsu et al. | |
| Animal feeding | Cattle | Grass-fed/grain-fed | GC–MS, LC–MS/MS | PCA, RF | [20] | Carrillo et al. |
| Cattle | Dietary amino acid supplementation | 1H–NMR | PCA | [21] | Yu et al. | |
| Cattle | Dietary mate extract supplementation | 1H–NMR | PCA | [22] | de Zawadzki et al. | |
| Pig | Clenbuterol supplementation | GC–MS | PCA, PLS–DA, OPLS–DA | [23] | Li et al. | |
| Chicken | Lysine supplementation | CE–MS | - | [24] | Watanabe et al. | |
| Chicken | Age | 1H–NMR | PCA, OPLS–DA | [25] | Liu et al. | |
| Pig | Ractopamine supplementation | REIMS | PCA, LDA, OPLS–DA | [26] | Guitton et al. | |
| Postmortem aging | Pork | Pm. aging period, muscle type | CE–MS | PCA | [14] | Muroya et al. |
| Beef | Pm. aging period, muscle type | LC–MS | PCA | [15] | Ma et al. | |
| Pork | Pm. aging period, muscle type | UPLC–MS/MS | PCA | [16] | Yu et al. | |
| Beef | Pm. aging period | CE–MS | PCA | [17] | Muroya et al. | |
| Beef | Pm. aging period | 1H–NMR | OPLS–DA | [18] | Kodani et al. | |
| Beef | Pm. aging period | 1H–NMR | PCA | [19] | Graham et al. | |
| Pork | Pm. aging period, muscle type (on thiamine) | CE–MS | - | [54] | Muroya et al. | |
| Beef | Pm. aging period | LC–MS | PCA | [55] | Lana et al. | |
| Beef | Pm. period of dry-aging | 1H–NMR | - | [56] | Kim et al. | |
| Lamb | Fast chilling effect | LC–MS, 1H– and 31P–NMR | PCA | [57] | Warner et al. | |
| Beef | Pm. aging period (on oxidative stability) | GC–MS | PCA | [58] | Mitacek et al. | |
| Processing | Pork | Marination time | 1H–NMR | PCA, OPLS–DA | [30] | Yang et al. |
| Pork | Drying/aging period, fermentation of sausage | HR–MAS 1H–NMR | PCA | [59] | García-García et al. | |
| Processing, authentication | Pork | Geographic origin, processing method | CE–MS | PCA | [27] | Sugimoto et al. |
| Pork | Geographic origin, processing method | 1H–NMR | PCA, OPLS–DA | [28] | Zhang et al. | |
| Processing, Spoilage | Chicken | Marinade type, storage time, microbial load, sensory score | GC–MS | PCA, FDA | [7] | Lytou et al. |
| Chicken | Marinade type, marination time and temperature | LC | PCA | [29] | Lytou et al. | |
| Sensory evaluation | Beef | Grinding score, packaging method | LC–MS | PCA, PLS | [60] | Jiang et al. |
| Beef | Commercial brands | GC–MS | – | [61] | Suzuki et al. | |
| Spoilage | Pork | Salmonellae contamination, time of microbial exposure | GC–MS | PCA, etc. | [62,63] | Xu et al. |
| Beef | Packaging, temperature | LC–MS | PCA, FDA, PLS–R | [64] | Argyri et al. | |
| Beef | Packaging, temperature, sensory score, microbial growth | HS/SPME GC–MS | PCA, FDA, PLS–R | [65] | Argyri et al. | |
| Authentication | Beef | Geographic origin | 1H–NMR | PCA, OPLS–DA | [66] | Jung et al. |
| Beef | Geographic origin | IMS | PCA | [67] | Zaima et al. | |
| Beef | Production system | 1H–NMR | PLS–DA | [68] | Osorio et al. | |
| Beef, Pork | Species | GC–MS, UPLC–MS | PCA, PLS–DA, Pathway | [69] | Trivedi, et al. | |
| Beef, Pork | Species | HS/SPME GC–MS | PCA, PLS–DA | [70] | Pavlidis et al. | |
| Chicken | Live/dead on arrival | LC–MS | PCA | [71] | Sidwick et al. | |
| Chicken | Live/dead on arrival | LC–MS | PCA, Pathway | [72] | Cao et al. | |
| Beef | Irradiation doses (on lipids) | 1H–NMR | sLDA, ANN | [73] | Zanardi et al. | |
| Beef | Irradiation doses (on hydrophilic compounds) | 1H–NMR | PCA, CT | [74] | Zanardi et al. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muroya, S.; Ueda, S.; Komatsu, T.; Miyakawa, T.; Ertbjerg, P. MEATabolomics: Muscle and Meat Metabolomics in Domestic Animals. Metabolites 2020, 10, 188. https://doi.org/10.3390/metabo10050188
Muroya S, Ueda S, Komatsu T, Miyakawa T, Ertbjerg P. MEATabolomics: Muscle and Meat Metabolomics in Domestic Animals. Metabolites. 2020; 10(5):188. https://doi.org/10.3390/metabo10050188
Chicago/Turabian StyleMuroya, Susumu, Shuji Ueda, Tomohiko Komatsu, Takuya Miyakawa, and Per Ertbjerg. 2020. "MEATabolomics: Muscle and Meat Metabolomics in Domestic Animals" Metabolites 10, no. 5: 188. https://doi.org/10.3390/metabo10050188