Metabolic Engineering Design Strategies for Increasing Acetyl-CoA Flux


Abstract
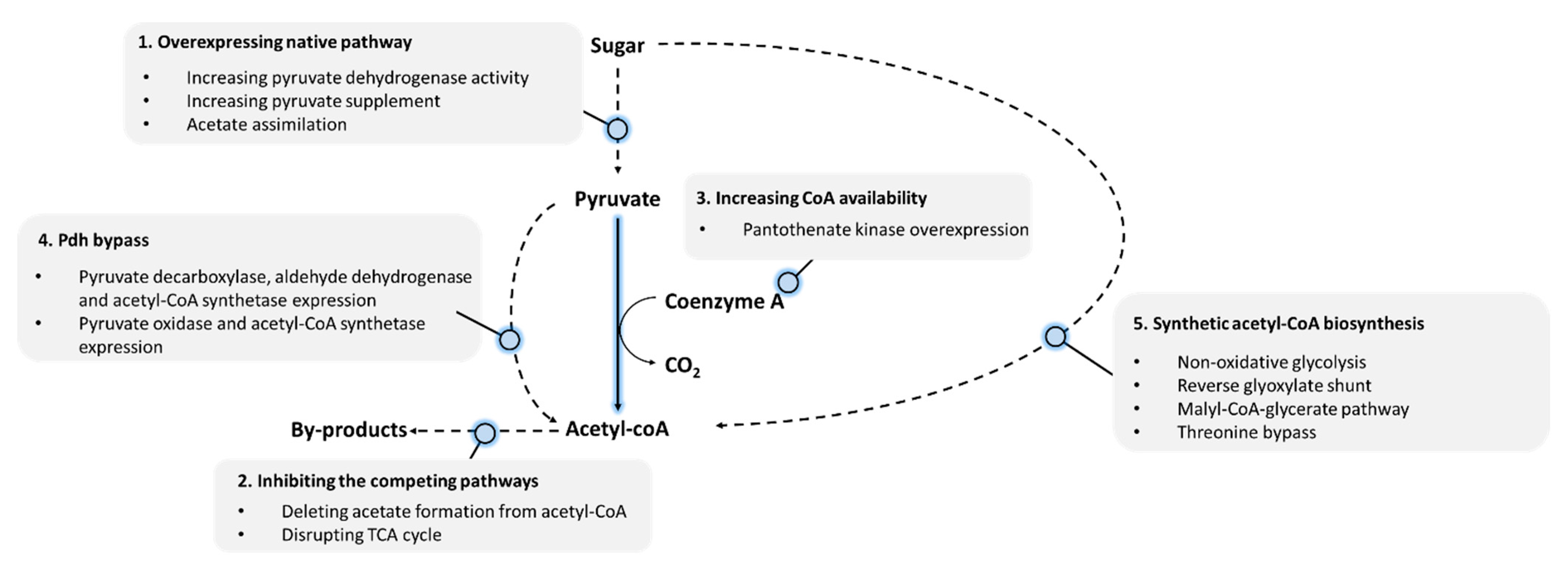
:1. Introduction
2. Overexpressing Native Pathways
2.1. Increasing Pyruvate Dehydrogenase Activity
2.2. Increasing Pyruvate Flux Drives Downstream Acetyl-CoA Formation
2.3. Acetate Assimilation into Acetyl-CoA
3. Inhibition of Competing Pathways
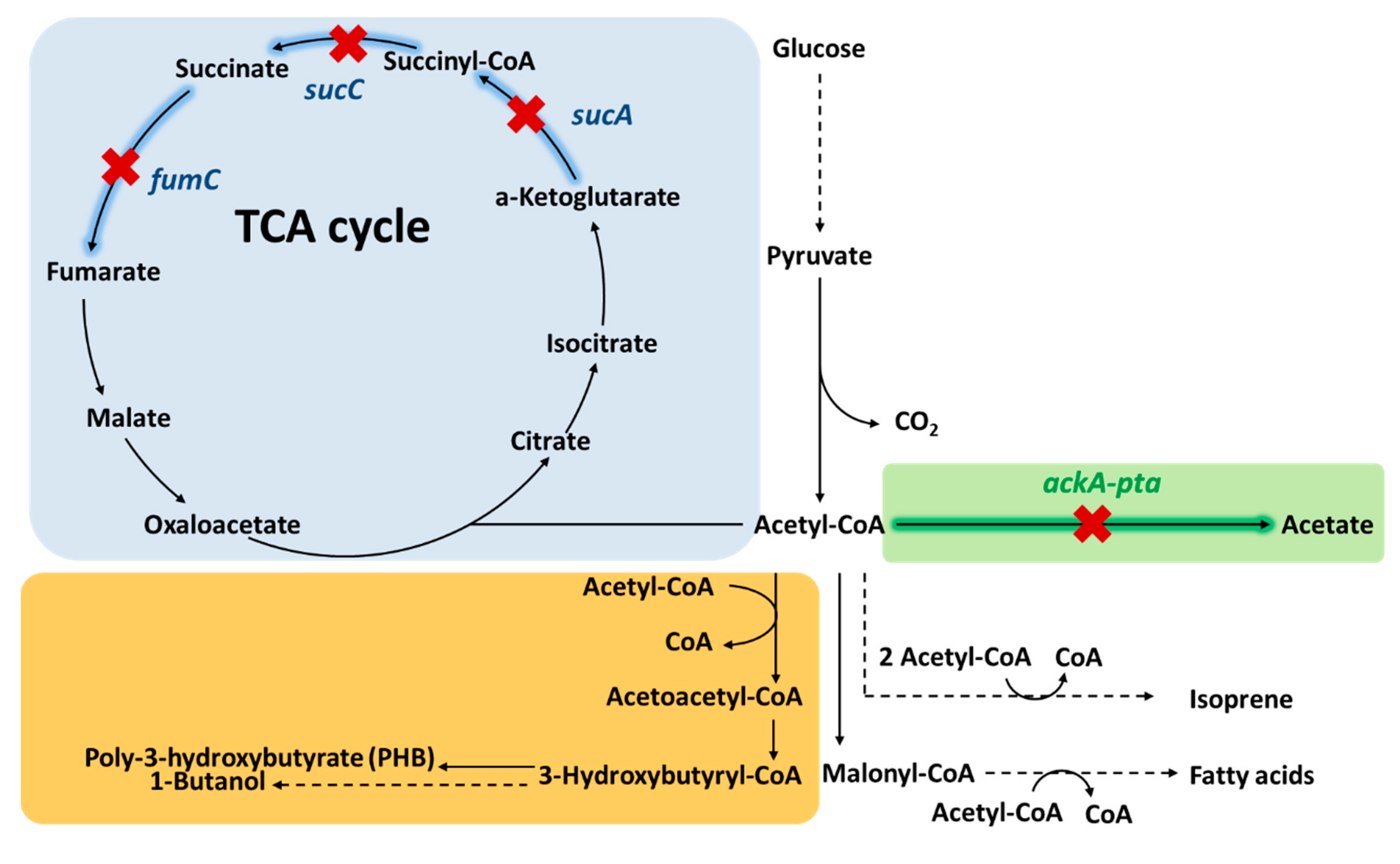
3.1. Deletion of Phosphate Acetyltransferase and Acetyl-CoA Kinase to Reduce Acetate Production
3.2. Disrupting TCA Cycle
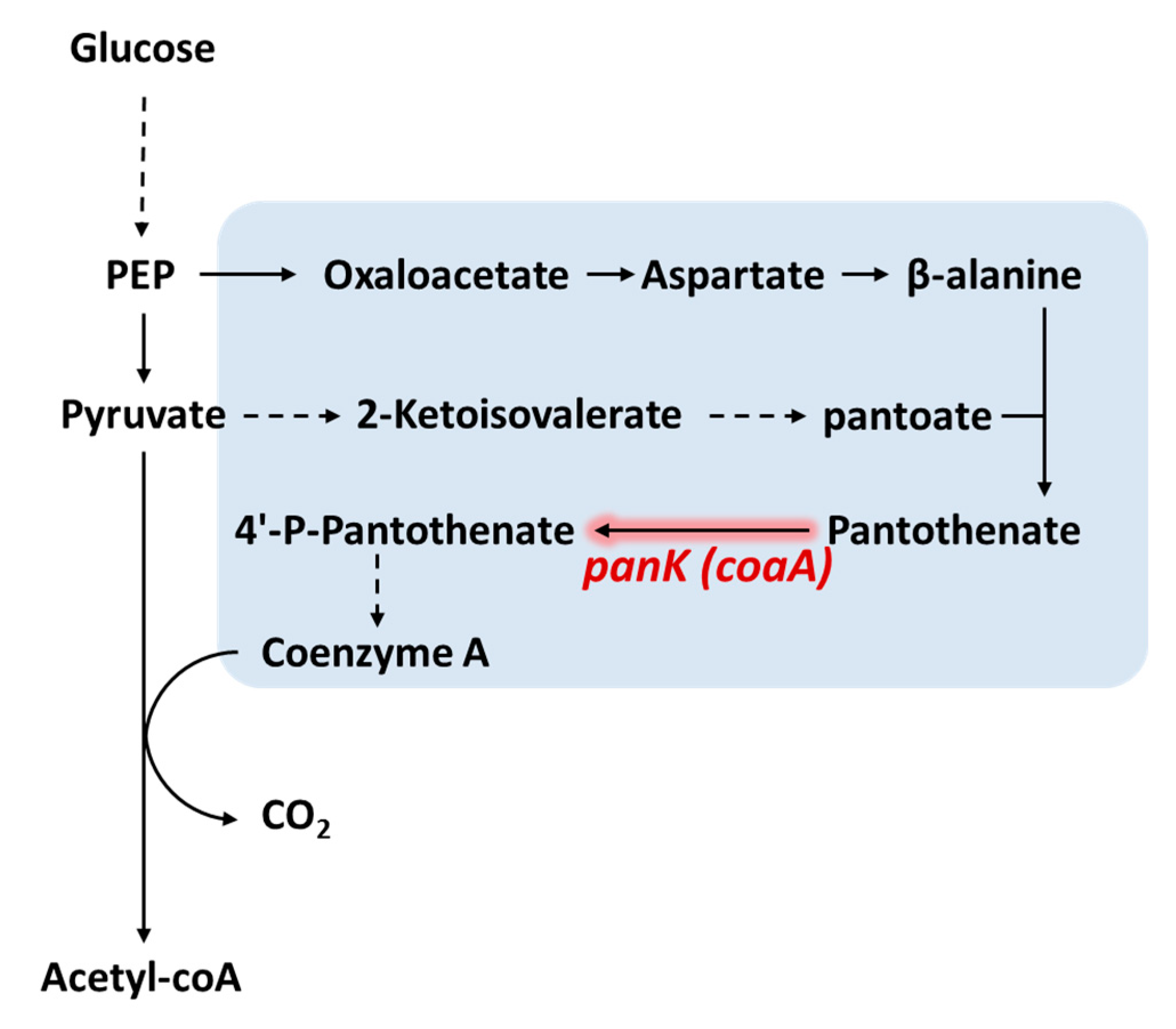
4. Increasing CoA Availability
5. Construction of Pyruvate Dehydrogenase Bypass
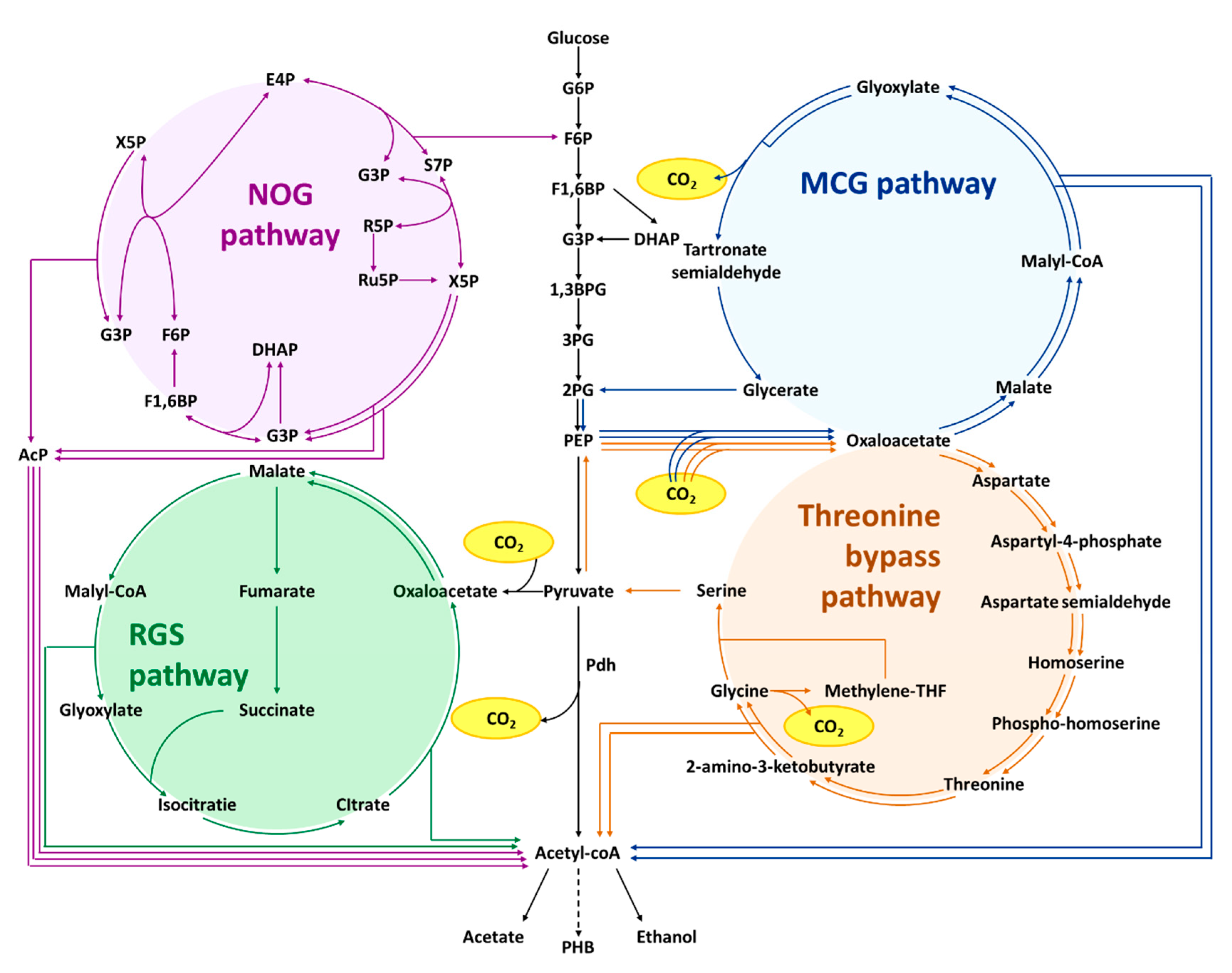
6. Synthetic Acetyl-CoA Biosynthesis
6.1. Non-Oxidative Glycolysis (NOG)
6.2. Reverse Glyoxylate Shunt (rGS)
6.3. Malyl-CoA-Glycerate Pathway (MCG Pathway)
6.4. Threonine Bypass
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- de Kok, A.; Hengeveld, A.F.; Martin, A.; Westphal, A.H. The pyruvate dehydrogenase multi-enzyme complex from Gram-negative bacteria. Biochim. Et Biophys. Acta (Bba)-Protein Struct. Mol. Enzymol. 1998, 1385, 353–366. [Google Scholar] [CrossRef]
- Shen, L.C.; Atkinson, D.E. Regulation of pyruvate dehydrogenase from Escherichia coli interactions of adenylate energy charge and other regulatory parameters. J. Biol. Chem. 1970, 245, 5974–5978. [Google Scholar]
- Dietrich, J.; Henning, U. Regulation of pyruvate dehydrogenase complex synthesis in Escherichia coli K12: Identification of the inducing metabolite. Eur. J. Biochem. 1970, 14, 258–269. [Google Scholar] [CrossRef]
- Kumari, S.; Beatty, C.M.; Browning, D.F.; Busby, S.J.; Simel, E.J.; Hovel-Miner, G.; Wolfe, A.J. Regulation of acetyl coenzyme A synthetase in Escherichia coli. J. Bacteriol. 2000, 182, 4173–4179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennig, J.; Kern, G.; Neef, H.; Spinka, M.; Bisswanger, H.; Hübner, G. Molecular mechanism of regulation of the pyruvate dehydrogenase complex from E. coli. Biochemistry 1997, 36, 15772–15779. [Google Scholar] [CrossRef] [PubMed]
- Broussard, T.C.; Price, A.E.; Laborde, S.M.; Waldrop, G.L. Complex formation and regulation of Escherichia coli acetyl-CoA carboxylase. Biochemistry 2013, 52, 3346–3357. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, S.; Radke, G.A.; Guest, J.R.; Roche, T.E. Lipoyl domain-based mechanism for the integrated feedback control of the pyruvate dehydrogenase complex by enhancement of pyruvate dehydrogenase kinase activity. J. Biol. Chem. 1996, 271, 653–662. [Google Scholar] [CrossRef] [Green Version]
- Sawers, G.; Suppmann, B. Anaerobic induction of pyruvate formate-lyase gene expression is mediated by the ArcA and FNR proteins. J. Bacteriol. 1992, 174, 3474–3478. [Google Scholar] [CrossRef] [Green Version]
- Holms, H. Flux analysis and control of the central metabolic pathways in Escherichia coli. Fems Microbiol. Rev. 1996, 19, 85–116. [Google Scholar] [CrossRef]
- Kitamura, S.; Toya, Y.; Shimizu, H. (13)C-Metabolic Flux Analysis Reveals Effect of Phenol on Central Carbon Metabolism in Escherichia coli. Front. Microbiol. 2019, 10, 1010. [Google Scholar] [CrossRef] [Green Version]
- Takamura, Y.; Nomura, G. Changes in the intracellular concentration of acetyl-CoA and malonyl-CoA in relation to the carbon and energy metabolism of Escherichia coli K12. Microbiology 1988, 134, 2249–2253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chohnan, S.; Izawa, H.; Nishihara, H.; TAKAMURA, Y. Changes in size of intracellular pools of coenzyme A and its thioesters in Escherichia coli K-12 cells to various carbon sources and stresses. Biosci. Biotechnol. Biochem. 1998, 62, 1122–1128. [Google Scholar] [CrossRef] [PubMed]
- Chohnan, S.; Furukawa, H.; Fujio, T.; Nishihara, H.; Takamura, Y. Changes in the size and composition of intracellular pools of nonesterified coenzyme A and coenzyme A thioesters in aerobic and facultatively anaerobic bacteria. Appl. Environ. Microbiol. 1997, 63, 553–560. [Google Scholar] [CrossRef] [Green Version]
- De Graef, M.R.; Alexeeva, S.; Snoep, J.L.; de Mattos, M.J.T. The steady-state internal redox state (NADH/NAD) reflects the external redox state and is correlated with catabolic adaptation in Escherichia coli. J. Bacteriol. 1999, 181, 2351–2357. [Google Scholar] [CrossRef] [Green Version]
- Hasona, A.; Kim, Y.; Healy, F.; Ingram, L.; Shanmugam, K. Pyruvate formate lyase and acetate kinase are essential for anaerobic growth of Escherichia coli on xylose. J. Bacteriol. 2004, 186, 7593–7600. [Google Scholar] [CrossRef] [Green Version]
- Dittrich, C.R.; Vadali, R.V.; Bennett, G.N.; San, K.Y. Redistribution of metabolic fluxes in the central aerobic metabolic pathway of E. coli mutant strains with deletion of the ackA-pta and poxB pathways for the synthesis of isoamyl acetate. Biotechnol. Prog. 2005, 21, 627–631. [Google Scholar] [CrossRef]
- Xu, P.; Ranganathan, S.; Fowler, Z.L.; Maranas, C.D.; Koffas, M.A. Genome-scale metabolic network modeling results in minimal interventions that cooperatively force carbon flux towards malonyl-CoA. Metab. Eng. 2011, 13, 578–587. [Google Scholar] [CrossRef]
- Xu, P.; Gu, Q.; Wang, W.; Wong, L.; Bower, A.G.; Collins, C.H.; Koffas, M.A. Modular optimization of multi-gene pathways for fatty acids production in E. coli. Nat. Commun. 2013, 4, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Hirokawa, Y.; Kubo, T.; Soma, Y.; Saruta, F.; Hanai, T. Enhancement of acetyl-CoA flux for photosynthetic chemical production by pyruvate dehydrogenase complex overexpression in Synechococcus elongatus PCC 7942. Metab. Eng. 2020, 57, 23–30. [Google Scholar] [CrossRef]
- Hansen, H.; Henning, U. Regulation of pyruvate dehydrogenase activity in Escherichia coli K12. Biochim. Et Biophys. Acta 1966, 122, 355–358. [Google Scholar] [CrossRef]
- Kim, Y.; Ingram, L.; Shanmugam, K. Dihydrolipoamide dehydrogenase mutation alters the NADH sensitivity of pyruvate dehydrogenase complex of Escherichia coli K-12. J. Bacteriol. 2008, 190, 3851–3858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Ou, M.S.; Kim, Y.; Ingram, L.; Shanmugam, K. Metabolic flux control at the pyruvate node in an anaerobic Escherichia coli strain with an active pyruvate dehydrogenase. Appl. Environ. Microbiol. 2010, 76, 2107–2114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saini, M.; Li, S.-Y.; Wang, Z.W.; Chiang, C.-J.; Chao, Y.-P. Systematic engineering of the central metabolism in Escherichia coli for effective production of n-butanol. Biotechnol. Biofuels 2016, 9, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yim, H.; Haselbeck, R.; Niu, W.; Pujol-Baxley, C.; Burgard, A.; Boldt, J.; Khandurina, J.; Trawick, J.D.; Osterhout, R.E.; Stephen, R. Metabolic engineering of Escherichia coli for direct production of 1,4-butanediol. Nat. Chem. Biol. 2011, 7, 445. [Google Scholar] [CrossRef]
- Ranganathan, S.; Suthers, P.F.; Maranas, C.D. OptForce: An Optimization Procedure for Identifying All Genetic Manipulations Leading to Targeted Overproductions. PLoS Comput Biol 2010, 6, e1000744. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, Z.; Liu, Q.; Li, Y.; Wang, Z.; Ma, H.; Chen, T.; Zhao, X. Engineering of Serine-Deamination pathway, Entner-Doudoroff pathway and pyruvate dehydrogenase complex to improve poly (3-hydroxybutyrate) production in Escherichia coli. Microb. Cell Factories 2014, 13, 172. [Google Scholar] [CrossRef] [Green Version]
- Dittrich, C.R.; Bennett, G.N.; San, K.Y. Characterization of the acetate-producing pathways in Escherichia coli. Biotechnol. Prog. 2005, 21, 1062–1067. [Google Scholar] [CrossRef]
- Fraenkel, D.; Neidhardt, F. Escherichia coli and Salmonella: Cellular and molecular biology; ASM Press: Washington DC, USA, 1996. [Google Scholar]
- Lin, H.; Castro, N.M.; Bennett, G.N.; San, K.-Y. Acetyl-CoA synthetase overexpression in Escherichia coli demonstrates more efficient acetate assimilation and lower acetate accumulation: A potential tool in metabolic engineering. Appl. Microbiol. Biotechnol. 2006, 71, 870–874. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, X.; Xia, X.; Dong, M. A systematic optimization of medium chain fatty acid biosynthesis via the reverse beta-oxidation cycle in Escherichia coli. Metab. Eng. 2017, 41, 115–124. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, W.; Chen, H.; Liu, B.; Lin, B.; Tao, Y. Metabolic engineering for efficient supply of acetyl-CoA from different carbon sources in Escherichia coli. Microb. Cell Factories 2019, 18, 130. [Google Scholar] [CrossRef]
- Chen, J.; Li, W.; Zhang, Z.-Z.; Tan, T.-W.; Li, Z.-J. Metabolic engineering of Escherichia coli for the synthesis of polyhydroxyalkanoates using acetate as a main carbon source. Microb. Cell Factories 2018, 17, 102. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Cha, S.; Kang, C.W.; Lee, G.M.; Lim, H.G.; Jung, G.Y. Efficient conversion of acetate to 3-hydroxypropionic acid by engineered Escherichia coli. Catalysts 2018, 8, 525. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.M.; Na, D.; Lee, S.Y. Design and use of synthetic regulatory small RNAs to control gene expression in Escherichia coli. Nat. Protoc. 2013, 8, 1694–1707. [Google Scholar] [CrossRef]
- Larson, M.H.; Gilbert, L.A.; Wang, X.W.; Lim, W.A.; Weissman, J.S.; Qi, L.S. CRISPR interference (CRISPRi) for sequence-specific control of gene expression. Nat. Protoc. 2013, 8, 2180–2196. [Google Scholar] [CrossRef] [Green Version]
- Vadali, R.; Horton, C.; Rudolph, F.; Bennett, G.; San, K.-Y. Production of isoamyl acetate in ackA-pta and/or ldh mutants of Escherichia coli with overexpression of yeast ATF2. Appl. Microbiol. Biotechnol. 2004, 63, 698–704. [Google Scholar] [CrossRef]
- van Wegen, R.J.; Lee, S.Y.; Middelberg, A.P. Metabolic and kinetic analysis of poly (3-hydroxybutyrate) production by recombinant Escherichia coli. Biotechnol. Bioeng. 2001, 74, 70–81. [Google Scholar] [CrossRef]
- Ku, J.T.; Lan, E.I. A balanced ATP driving force module for enhancing photosynthetic biosynthesis of 3-hydroxybutyrate from CO2. Metab. Eng. 2018, 46, 35–42. [Google Scholar] [CrossRef]
- Centeno-Leija, S.; Huerta-Beristain, G.; Giles-Gómez, M.; Bolivar, F.; Gosset, G.; Martinez, A. Improving poly-3-hydroxybutyrate production in Escherichia coli by combining the increase in the NADPH pool and acetyl-CoA availability. Antonie Van Leeuwenhoek 2014, 105, 687–696. [Google Scholar] [CrossRef]
- Shen, C.R.; Lan, E.I.; Dekishima, Y.; Baez, A.; Cho, K.M.; Liao, J.C. Driving forces enable high-titer anaerobic 1-butanol synthesis in Escherichia coli. Appl. Environ. Microbiol. 2011, 77, 2905–2915. [Google Scholar] [CrossRef] [Green Version]
- Ohtake, T.; Pontrelli, S.; Laviña, W.A.; Liao, J.C.; Putri, S.P.; Fukusaki, E. Metabolomics-driven approach to solving a CoA imbalance for improved 1-butanol production in Escherichia coli. Metab. Eng. 2017, 41, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Lin, F.; Chen, Y.; Levine, R.; Lee, K.; Yuan, Y.; Lin, X.N. Improving fatty acid availability for bio-hydrocarbon production in Escherichia coli by metabolic engineering. PLoS ONE 2013, 8, e78595. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-H.; Wang, C.; Jang, H.-J.; Cha, M.-S.; Park, J.-E.; Jo, S.-Y.; Choi, E.-S.; Kim, S.-W. Isoprene production by Escherichia coli through the exogenous mevalonate pathway with reduced formation of fermentation byproducts. Microb. Cell Factories 2016, 15, 214. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.H.; Dhamankar, H.; Tseng, H.-C.; Sheppard, M.J.; Reisch, C.R.; Prather, K.L. A platform pathway for production of 3-hydroxyacids provides a biosynthetic route to 3-hydroxy-γ-butyrolactone. Nat. Commun. 2013, 4, 1414. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Gonzalez, R.; Bobik, T.A. Coproduction of acetaldehyde and hydrogen during glucose fermentation by Escherichia coli. Appl. Environ. Microbiol. 2011, 77, 6441–6450. [Google Scholar] [CrossRef] [Green Version]
- Vuoristo, K.S.; Mars, A.E.; Sangra, J.V.; Springer, J.; Eggink, G.; Sanders, J.P.; Weusthuis, R.A. Metabolic engineering of itaconate production in Escherichia coli. Appl. Microbiol. Biotechnol. 2015, 99, 221–228. [Google Scholar] [CrossRef]
- Miyake, M.; Schnackenberg, J.; Kurane, R.; Asada, Y. Phosphotransacetylase as a key factor in biological production of polyhydroxybutyrate. Appl. Biochem. Biotechnol. 2000, 84, 1039–1044. [Google Scholar] [CrossRef]
- Jian, J.; Zhang, S.-Q.; Shi, Z.-Y.; Wang, W.; Chen, G.-Q.; Wu, Q. Production of polyhydroxyalkanoates by Escherichia coli mutants with defected mixed acid fermentation pathways. Appl. Microbiol. Biotechnol. 2010, 87, 2247–2256. [Google Scholar] [CrossRef]
- Ku, J.T.; Simanjuntak, W.; Lan, E.I. Renewable synthesis of n-butyraldehyde from glucose by engineered Escherichia coli. Biotechnol. Biofuels 2017, 10, 291. [Google Scholar] [CrossRef]
- Srirangan, K.; Liu, X.; Akawi, L.; Bruder, M.; Moo-Young, M.; Chou, C.P. Engineering Escherichia coli for microbial production of butanone. Appl. Environ. Microbiol. 2016, 82, 2574–2584. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Niyompanich, S.; Tai, Y.-S.; Wang, J.; Bai, W.; Mahida, P.; Gao, T.; Zhang, K. Engineering of a highly efficient Escherichia coli strain for mevalonate fermentation through chromosomal integration. Appl. Environ. Microbiol. 2016, 82, 7176–7184. [Google Scholar] [CrossRef] [Green Version]
- Lan, E.I.; Wei, C.T. Metabolic engineering of cyanobacteria for the photosynthetic production of succinate. Metab. Eng. 2016, 38, 483–493. [Google Scholar] [CrossRef]
- Parimi, N.S.; Durie, I.A.; Wu, X.; Niyas, A.M.; Eiteman, M.A. Eliminating acetate formation improves citramalate production by metabolically engineered Escherichia coli. Microb. Cell Factories 2017, 16, 114. [Google Scholar] [CrossRef]
- Lan, E.I.; Ro, S.Y.; Liao, J.C. Oxygen-tolerant coenzyme A-acylating aldehyde dehydrogenase facilitates efficient photosynthetic n-butanol biosynthesis in cyanobacteria. Energy Environ. Sci. 2013, 6, 2672–2681. [Google Scholar] [CrossRef]
- Vallari, D.; Jackowski, S. Biosynthesis and degradation both contribute to the regulation of coenzyme A content in Escherichia coli. J. Bacteriol. 1988, 170, 3961–3966. [Google Scholar] [CrossRef] [Green Version]
- Vadali, R.V.; Bennett, G.N.; San, K.-Y. Cofactor engineering of intracellular CoA/acetyl-CoA and its effect on metabolic flux redistribution in Escherichia coli. Metab. Eng. 2003, 6, 133–139. [Google Scholar] [CrossRef]
- Jackowski, S.; Rock, C. Regulation of coenzyme A biosynthesis. J. Bacteriol. 1981, 148, 926–932. [Google Scholar] [CrossRef] [Green Version]
- Rock, C.O.; Calder, R.B.; Karim, M.A.; Jackowski, S. Pantothenate kinase regulation of the intracellular concentration of coenzyme A. J. Biol. Chem. 2000, 275, 1377–1383. [Google Scholar] [CrossRef] [Green Version]
- Song, W.-J.; Jackowski, S. Kinetics and regulation of pantothenate kinase from Escherichia coli. J. Biol. Chem. 1994, 269, 27051–27058. [Google Scholar]
- Vallari, D.; Jackowski, S.; Rock, C. Regulation of pantothenate kinase by coenzyme A and its thioesters. J. Biol. Chem. 1987, 262, 2468–2471. [Google Scholar]
- Vadali, R.V.; Bennett, G.N.; San, K.-Y. Applicability of CoA/acetyl-CoA manipulation system to enhance isoamyl acetate production in Escherichia coli. Metab. Eng. 2004, 6, 294–299. [Google Scholar] [CrossRef]
- Lin, H.; Vadali, R.V.; Bennett, G.N.; San, K.Y. Increasing the acetyl-CoA pool in the presence of overexpressed phosphoenolpyruvate carboxylase or pyruvate carboxylase enhances succinate production in Escherichia coli. Biotechnol. Prog. 2004, 20, 1599–1604. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.-Y.; Masuda, T.; Kawamura, T.; Hata, S.; Izui, K. Cloning, expression, and characterization of a root-form phosphoenolpyruvate carboxylase from Zea mays: Comparison with the C4-form enzyme. Plant Cell Physiol. 1998, 39, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-m.; Omiya, T.; Hata, S.; Izui, K. Molecular characterization of a phosphoenolpyruvate carboxylase from a thermophilic cyanobacterium, Synechococcus vulcanus with unusual allosteric properties. Plant Cell Physiol. 2002, 43, 159–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogata, Y.; Chohnan, S. Prokaryotic type III pantothenate kinase enhances coenzyme A biosynthesis in Escherichia coli. J. Gen. Appl. Microbiol. 2015, 61, 266–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiba, Y.; Paradise, E.M.; Kirby, J.; Ro, D.-K.; Keasling, J.D. Engineering of the pyruvate dehydrogenase bypass in Saccharomyces cerevisiae for high-level production of isoprenoids. Metab. Eng. 2007, 9, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhang, B.; Jiang, R. Improving acetyl-CoA biosynthesis in Saccharomyces cerevisiae via the overexpression of pantothenate kinase and PDH bypass. Biotechnol. Biofuels 2017, 10, 41. [Google Scholar] [CrossRef] [Green Version]
- Soma, Y.; Yamaji, T.; Matsuda, F.; Hanai, T. Synthetic metabolic bypass for a metabolic toggle switch enhances acetyl-CoA supply for isopropanol production by Escherichia coli. J. Biosci. Bioeng. 2017, 123, 625–633. [Google Scholar] [CrossRef]
- Lai, M.J.; Lan, E.I. Photoautotrophic synthesis of butyrate by metabolically engineered cyanobacteria. Biotechnol. Bioeng. 2019, 116, 893–903. [Google Scholar] [CrossRef]
- Lan, E.I.; Liao, J.C. ATP drives direct photosynthetic production of 1-butanol in cyanobacteria. Proc. Natl. Acad. Sci. USA 2012, 109, 6018–6023. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Son, J.; Sim, S.J.; Woo, H.M. Metabolic rewiring of synthetic pyruvate dehydrogenase bypasses for acetone production in cyanobacteria. Plant Biotechnol. J. 2020. [Google Scholar] [CrossRef]
- Bogorad, I.W.; Lin, T.-S.; Liao, J.C. Synthetic non-oxidative glycolysis enables complete carbon conservation. Nature 2013, 502, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Henard, C.A.; Freed, E.F.; Guarnieri, M.T. Phosphoketolase pathway engineering for carbon-efficient biocatalysis. Curr. Opin. Biotechnol. 2015, 36, 183–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, P.P.; Jaeger, A.J.; Wu, T.-Y.; Xu, S.C.; Lee, A.S.; Gao, F.; Chen, P.-W.; Liao, J.C. Construction and evolution of an Escherichia coli strain relying on nonoxidative glycolysis for sugar catabolism. Proc. Natl. Acad. Sci. USA 2018, 115, 3538–3546. [Google Scholar] [CrossRef] [Green Version]
- Mainguet, S.E.; Gronenberg, L.S.; Wong, S.S.; Liao, J.C. A reverse glyoxylate shunt to build a non-native route from C4 to C2 in Escherichia coli. Metab. Eng. 2013, 19, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Li, X.; Duchoud, F.; Chuang, D.S.; Liao, J.C. Augmenting the Calvin–Benson–Bassham cycle by a synthetic malyl-CoA-glycerate carbon fixation pathway. Nat. Commun. 2018, 9, 2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.; Zhang, Y.; Yuan, Q.; Liu, Q.; Li, Y.; Wang, Z.; Ma, H.; Chen, T.; Zhao, X. Metabolic engineering of Escherichia coli for poly (3-hydroxybutyrate) production via threonine bypass. Microb. Cell Factories 2015, 14, 185. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Ma, W.; Fang, Y.; Yang, J.; Zhan, J.; Chen, S.; Wang, X. Increasing l-threonine production in Escherichia coli by overexpressing the gene cluster phaCAB. J. Ind. Microbiol. Biotechnol. 2019, 46, 1557–1568. [Google Scholar] [CrossRef]
- Rodriguez, S.; Denby, C.M.; Van Vu, T.; Baidoo, E.E.; Wang, G.; Keasling, J.D. ATP citrate lyase mediated cytosolic acetyl-CoA biosynthesis increases mevalonate production in Saccharomyces cerevisiae. Microb. Cell Factories 2016, 15, 48. [Google Scholar] [CrossRef] [Green Version]
- Kozak, B.U.; van Rossum, H.M.; Luttik, M.A.; Akeroyd, M.; Benjamin, K.R.; Wu, L.; de Vries, S.; Daran, J.-M.; Pronk, J.T.; van Maris, A.J. Engineering acetyl coenzyme A supply: Functional expression of a bacterial pyruvate dehydrogenase complex in the cytosol of Saccharomyces cerevisiae. MBio 2014, 5, e01696-01614. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Approach | Strategy | Genetic Manipulations | Rational | Example |
|---|---|---|---|---|
| Overexpressing native pathways | Increase pyruvate dehydrogenase activity | Overexpressing aceEF, lpd | Overexpression of pyruvate dehydrogenase to increase activity | [16,17] |
| Lpd * (E354K) | Introducing the mutation in E3 subunit of Pdh increased anaerobic activity due to insensitive to NADH | [22,23] | ||
| Increase pyruvate supplement | △serC::Ptrc-162::pgk, serABC; | Entner–Doudoroff pathway converts glucose-6-phopshate to pyruvate using the genes, zwf, edd, and eda | [26] | |
| PsdaA::Ptrc; Pedd-eda::PJ23119; Pzwf::Ptrc-162; | Pgk helps to provide 3PG from 1,3BPG. Then, the serine-deamination pathway converts 3PG into pyruvate using the genes serABC and sdaA. | [26] | ||
| Assimilate acetate | Overexpressing acs or pta-ack | Activating acetate into acetyl-CoA, allowing the assimilation of naturally secreted or supplemented acetate | [31,32,33] | |
| Inhibiting the competing pathway | Deleting acetate formation from acetyl-CoA | △pta | Deletion of phospho acetyltransferase blocks the consumption of acetyl-CoA into acetate | [40,42,43] |
| Disrupting TCA cycle | △sucA | Deletion of α-ketoglutarate dehydrogenase (sucA) disrupts the TCA cycle but maintains glutamate synthesis | [51] | |
| △scuC △fumC | Deletion of succinyl-CoA synthetase (sucC) and fumarase (fumC) disrupt the TCA cycle | [17] | ||
| △gltA | Deletion of citrate synthase to block the first step of the TCA cycle | [53] | ||
| Enhancing CoA availability | Enhance CoA synthesis | Overexpressing panK | Pantothenate kinase serves as the limiting step of CoA synthesis | [56,61] |
| Pdh bypass | Pdh bypass | Overexpressing PDC, ACDH, and ACS | PDC, ACDH, and ACS allows the conversion of pyruvate to acetyl-CoA via acetaldehyde and acetate as intermediate | [66] |
| Overexpressing poxB and acs | poxB and acs allow the conversion of pyruvate to acetyl-CoA via acetate as intermediate | [68] | ||
| Synthetic acetyl-CoA biosynthesis | Non-oxidative glycolysis | ΔadhE, ΔldhA, ΔfrdBC, ΔpflB, PLlacO1::fbp, fxpk | Expression of fructose 1,6-bisphosphatase and phosphoketolase, allowing the conversion of glucose-phosphate to stoichiometry amount acetyl-phosphate or acetyl-CoA | [72] |
| Non-oxidative glycolysis | Evolved strain with 50 genomic mutation and PLlacO1::xpkBA *, glfZM, glk, tkt2MB, talKP, glpX PLlacO1::tkt2MB, tkt1MB | Combining evolution, the NOG pathway supports E. coli growth and nearly complete carbon conservation from glucose to acetyl-phosphate or acetyl-CoA | [74] | |
| Reverse glyoxylate shunt | ΔgltA Δmdh Δppc ΔcitE Δmqo ΔaceB Δicd PLlacO1::DctA, PLlacO1::AceA; PLlacO1:AcnA; PLlacO1::AclB, AclA; PLlacO1::SucCD-2, Mcl1 | rGS provides a net conversion of C4 compound such as malate into two acetyl-CoA | [75] | |
| Malyl-CoA-glycerate cycle | ∆aceB ∆glcB ∆frdB ∆ldhA ∆pstG PLlacO1::mtkB, mtkA(M.c), mcl; PLlacO1::gcl, hyi PLlacO1::garK, mdh PLlacO1::ppc | PEP carboxylase, malate dehydrogenase, malate thiokinase, and malyl-CoA lyase together with glyoxylate assimilation pathway allows two acetyl-CoA from PEP | [76] | |
| Threonine bypass | Pkbl-tdh::Ptrc, PsdaA::Ptrc, thrA::thrA * (C1034T), PthrABC::PTrc-162, Pppc::Ptac, PglyA::PTrc-162 | Expression of threonine synthesis pathway and threonine degradation converts PEP to acetyl-CoA and glycine, which is next converted to pyruvate. Together, generating an extra acetyl-CoA. | [77] |
| Acetyl-CoA Derived Biochemical | Acetyl-CoA Increasing Strategy Applied | Fold Increased | Titer Achieved | Reference |
|---|---|---|---|---|
| Isoamyl acetate | Increasing pyruvate dehydrogenase activity; gene deletion to reduce acetate formation | 1.5 | 0.023 g/L | [16] |
| Gene deletion to reduce acetate formation | 4.8 | 0.23 g/L | [36] | |
| Gene deletion to reduce acetate formation; increasing CoA availability | 2.3 | 0.144 g/L | [61] | |
| Isopropanol | Pyruvate dehydrogenase bypass | 4.4 | 3.8 g/L | [68] |
| 1,4-Butanediol | Increasing pyruvate dehydrogenase activity | 2.0 | 7.5 g/L | [24] |
| 1-Butanol | Increasing pyruvate dehydrogenase activity | 1.6 | 4.3 g/L | [23] |
| Gene deletion to reduce acetate formation | 10 | 30 g/L | [40] | |
| Poly-3-hydroxybutyrate | Increasing pyruvate flux; increasing pyruvate dehydrogenase activity | 2.7 | 5.5 g/L | [26] |
| Threonine bypass | 2.9 | 5.97 g/L | [77] | |
| N-Acetylglutamate | Gene deletion to reduce acetate formation; acetate assimilation | 2.0 | 17.8 g/L | [31] |
| 3-Hydroxypropionate | Acetate assimilation | 2.5 | 0.25 g/L | [33] |
| Citramalate | Gene deletion to reduce acetate formation; gene deletion to disrupt TCA cycle | 1.2 | 19.8 g/L | [53] |
| Succinate | Increasing CoA availability | 1.3 | 2.7 g/L | [62] |
| Fatty acid | Increasing pyruvate flux; increasing pyruvate dehydrogenase activity; gene deletion to disrupt TCA cycle | 5.6 | 0.47 g/L | [17] |
| Acetate assimilation | 2.2 | 0.68 g/L | [30] | |
| Mevalonate | Pyruvate dehydrogenase bypass | 1.4 | 0.37 g/L | [66] |
| Isoprene | Gene deletion to reduce acetate formation | 2.0 | 1.8 g/L | [43] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ku, J.T.; Chen, A.Y.; Lan, E.I. Metabolic Engineering Design Strategies for Increasing Acetyl-CoA Flux. Metabolites 2020, 10, 166. https://doi.org/10.3390/metabo10040166
Ku JT, Chen AY, Lan EI. Metabolic Engineering Design Strategies for Increasing Acetyl-CoA Flux. Metabolites. 2020; 10(4):166. https://doi.org/10.3390/metabo10040166
Chicago/Turabian StyleKu, Jason T., Arvin Y. Chen, and Ethan I. Lan. 2020. "Metabolic Engineering Design Strategies for Increasing Acetyl-CoA Flux" Metabolites 10, no. 4: 166. https://doi.org/10.3390/metabo10040166