The Urinary Metabolome of Healthy Newborns

 , , , , ,
, , , , , 
Abstract
:
1. Introduction
2. Results
2.1. Concentration Data of 136 Metabolites Measured by Targeted Metabolomics
2.2. Gender-Sex Differences Associated Metabolites
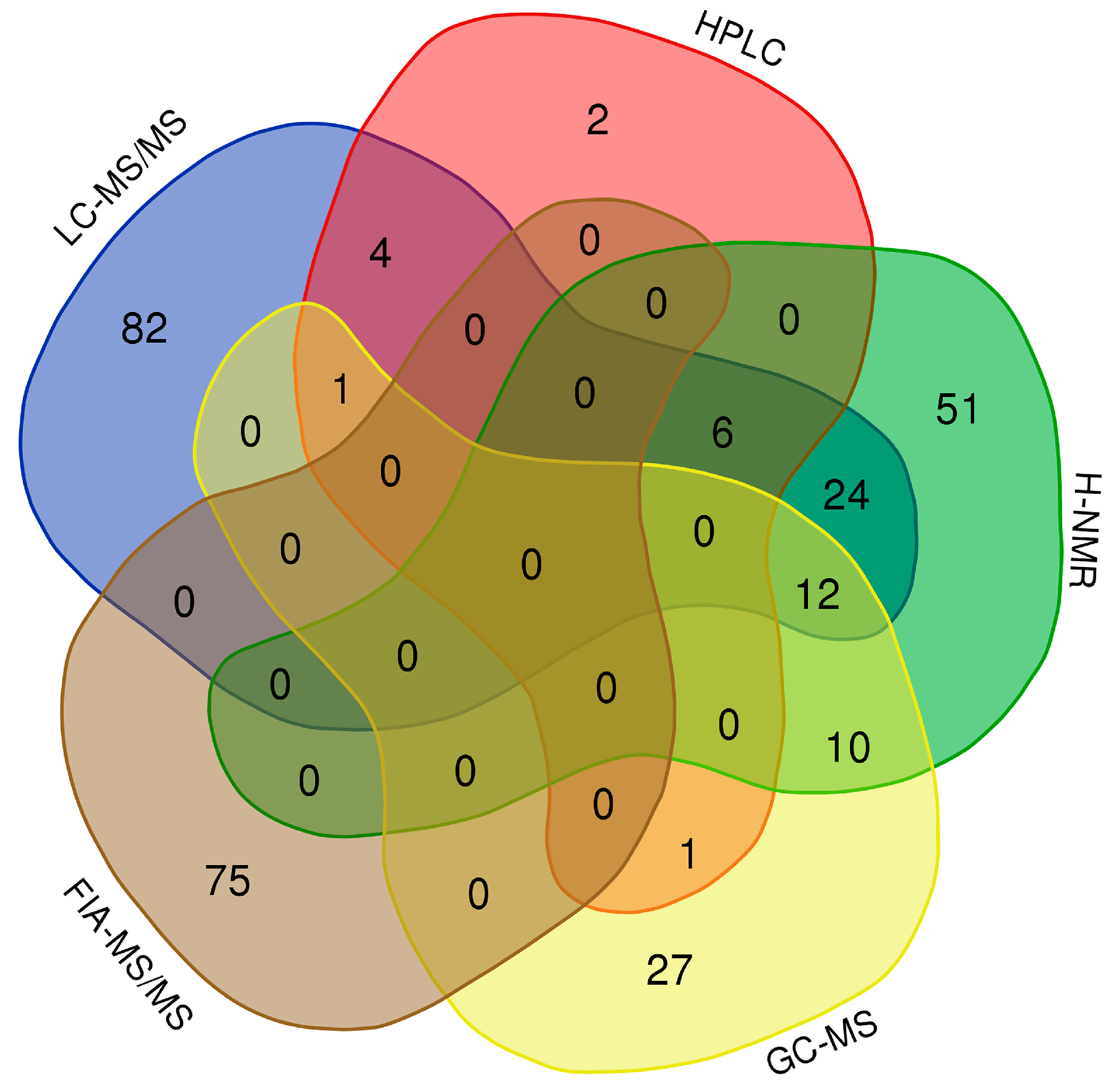
2.3. Impact of Resolution Mode on the Urinary Metabolome of Healthy Newborns
2.4. Literature Review of the Urinary Metabolome of Healthy Newborns
3. Discussion
3.1. Comparison of Experimental Values with Reported Reference Values
3.2. Gender-Sex Differences Associated Metabolites
3.3. Impact of Birth Resolution on the Urinary Metabolome of Healthy Newborns
3.4. Urinary Metabolites Associated with IEMs
3.5. The Importance of Age-Specific Intervals
4. Materials and Methods
4.1. Sample Collection and Research Ethics Approvals
4.2. Chemicals and Internal Standards (ISTD)
4.3. Metabolite Measurement
4.3.1. Sample Preparation
4.3.2. LC-MS/MS Method
4.3.3. FIA-MS/MS Method
4.3.4. Quantification
4.4. Statistical Analysis
4.5. Literature Review and Data Entry
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- United Nations Inter-agency Group for Child Mortality Estimation (UN IGME). Levels & Trends in Child Mortality: Report 2019; Estimates developed by the United Nations Inter-agency Group for Child Mortality Estimation; United Nations Children’s Fund: New York, NY, USA, 2019. [Google Scholar]
- Lehtonen, L.; Gimeno, A.; Parra-Llorca, A.; Vento, M. Early neonatal death: A challenge worldwide. Semin. Fetal Neonatal Med. 2017, 22, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Gemme, G.; Serra, G.; Rovetta, G.D. Causes of neonatal death in premature infants. Analysis of 1053 autopsies. Minerva Pediatr. 1973, 25, 273–279. [Google Scholar] [PubMed]
- Guthrie, R.; Susi, A. A Simple Phenylalanine Method for Detecting Phenylketonuria in Large Populations of Newborn Infants. Pediatrics 1963, 32, 338–343. [Google Scholar]
- Mussap, M.; Zaffanello, M.; Fanos, V. Metabolomics: A challenge for detecting and monitoring inborn errors of metabolism. Ann. Transl. Med. 2018, 6, 338. [Google Scholar] [CrossRef]
- Forfar, J. Normal and abnormal calcium, phosphorus and magnesium metabolism in the perinatal period. Clin. Endocrinol. Metab. 1976, 5, 123–148. [Google Scholar] [CrossRef]
- Hajjar, F.M. Neutrophils in the newborn: Normal characteristics and quantitative disorders. Semin. Perinatol. 1990, 14, 374–383. [Google Scholar]
- Knobel, R.B.; Smith, J.M. Laboratory Blood Tests Useful in Monitoring Renal Function in Neonates. Neonatal Netw. 2013, 33, 35–40. [Google Scholar] [CrossRef]
- Rennie, J.M.; Roberton, N.R.C. Textbook of Neonatology, 3rd ed.; Churchill Livingstone: Edinburgh, UK, 1999. [Google Scholar]
- Srinivasan, G.; Pildes, R.; Cattamanchi, G.; Voora, S.; Lilien, L. Plasma glucose values in normal neonates: A new look. J. Pediatr. 1986, 109, 114–117. [Google Scholar] [CrossRef]
- Wishart, D.S.; Tzur, D.; Knox, C.; Eisner, R.; Guo, A.C.; Young, N.; Cheng, D.; Jewell, K.; Arndt, D.; Sawhney, S.; et al. HMDB: The Human Metabolome Database. Nucleic Acids Res. 2007, 35, D521–D526. [Google Scholar] [CrossRef]
- Bouatra, S.; Aziat, F.; Mandal, R.; Guo, A.C.; Wilson, M.R.; Knox, C.; Bjorndahl, T.C.; Krishnamurthy, R.; Saleem, F.; Liu, P.; et al. The Human Urine Metabolome. PLoS ONE 2013, 8, e73076. [Google Scholar] [CrossRef] [Green Version]
- Psychogios, N.; Hau, D.D.; Peng, J.; Guo, A.C.; Mandal, R.; Bouatra, S.; Sinelnikov, I.; Krishnamurthy, R.; Eisner, R.; Gautam, B.; et al. The Human Serum Metabolome. PLoS ONE 2011, 6, e16957. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S.; Lewis, M.J.; Morrissey, J.A.; Flegel, M.D.; Jeroncic, K.; Xiong, Y.; Cheng, D.; Eisner, R.; Gautam, B.; Tzur, D.; et al. The human cerebrospinal fluid metabolome. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2008, 871, 164–173. [Google Scholar] [CrossRef]
- Dame, Z.; Aziat, F.; Mandal, R.; Krishnamurthy, R.; Bouatra, S.; Borzouie, S.; Guo, A.C.; Sajed, T.; Deng, L.; Lin, H.; et al. The human saliva metabolome. Metabolomics 2015, 11, 1864–1883. [Google Scholar] [CrossRef]
- Karu, N.; Deng, L.; Slae, M.; Guo, A.C.; Sajed, T.; Huynh, H.; Wine, E.; Wishart, D.S. A review on human fecal metabolomics: Methods, applications and the human fecal metabolome database. Anal. Chim. Acta 2018, 1030, 1–24. [Google Scholar] [CrossRef]
- Sinues, P.M.-L.; Tarokh, L.; Li, X.; Kohler, M.; Brown, S.A.; Zenobi, R.; Dallmann, R. Circadian Variation of the Human Metabolome Captured by Real-Time Breath Analysis. PLoS ONE 2014, 9, e114422. [Google Scholar] [CrossRef]
- Villaseñor, A.; Garcia-Perez, I.; Garcia, A.; Posma, J.M.; López, M.F.; Nicholas, A.J.; Modi, N.; Holmes, E.; Barbas, C. Breast Milk Metabolome Characterization in a Single-Phase Extraction, Multiplatform Analytical Approach. Anal. Chem. 2014, 86, 8245–8252. [Google Scholar] [CrossRef]
- Su, L.; Mao, J.; Hao, M.; Lu, T.; Mao, C.; Ji, D.; Tong, H.; Fei, C. Integrated Plasma and Bile Metabolomics Based on an UHPLC-Q/TOF-MS and Network Pharmacology Approach to Explore the Potential Mechanism of Schisandra chinensis-Protection from Acute Alcoholic Liver Injury. Front. Pharmacol. 2020, 10, 10. [Google Scholar] [CrossRef]
- Sulek, K.; Han, T.-L.; Villas-Boas, S.; Wishart, D.S.; E Soh, S.; Kwek, K.; Gluckman, P.D.; Chong, Y.-S.; Kenny, L.; Baker, P. Hair Metabolomics: Identification of Fetal Compromise Provides Proof of Concept for Biomarker Discovery. Theranostics 2014, 4, 953–959. [Google Scholar] [CrossRef]
- Dunn, W.; Lin, W.; Broadhurst, D.; Begley, P.; Brown, M.; Zelená, E.; Vaughan, A.A.; Halsall, A.; Harding, N.; Knowles, J.; et al. Molecular phenotyping of a UK population: Defining the human serum metabolome. Metabolomics 2014, 11, 9–26. [Google Scholar] [CrossRef] [Green Version]
- Rist, M.; Roth, A.; Frommherz, L.; Weinert, C.; Krüger, R.; Merz, B.; Bunzel, D.; Mack, C.; Egert, B.; Bub, A.; et al. Metabolite patterns predicting sex and age in participants of the Karlsruhe Metabolomics and Nutrition (KarMeN) study. PLoS ONE 2017, 12, e0183228. [Google Scholar] [CrossRef]
- Trabado, S.; Al-Salameh, A.; Croixmarie, V.; Masson, P.; Corruble, E.; Feve, B.; Colle, R.; Ripoll, L.; Walther, B.; Boursier-Neyret, C.; et al. The human plasma-metabolome: Reference values in 800 French healthy volunteers; impact of cholesterol, gender and age. PLoS ONE 2017, 12, e0173615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Embade, N.; Cannet, C.; Diercks, T.; Gil-Redondo, R.; Bruzzone, C.; Ansó, S.; Echevarría, L.R.; Ayucar, M.M.M.; Collazos, L.; Lodoso, B.; et al. NMR-based newborn urine screening for optimized detection of inherited errors of metabolism. Sci. Rep. 2019, 9, 13067–13069. [Google Scholar] [CrossRef] [PubMed]
- Van Gennip, A.H.; Abeling, N.G.G.M.; Stroomer, A.E.M.; Overmars, H.; Bakker, H.D. The detection of molybdenum cofactor deficiency: Clinical symptomatology and urinary metabolite profile. J. Inherit. Metab. Dis. 1994, 17, 142–145. [Google Scholar] [CrossRef]
- Zaima, K. Taurine Concentration in the Perinatal Period. Pediatr. Int. 1984, 26, 169–177. [Google Scholar] [CrossRef]
- Aerts, L.; Van Assche, F.A. Taurine and taurine-deficiency in the perinatal period. J. Périnat. Med. 2002, 30, 281–286. [Google Scholar] [CrossRef]
- Huynh, J.; Xiong, G.; Bentley-Lewis, R. A systematic review of metabolite profiling in gestational diabetes mellitus. Diabetologia 2014, 57, 2453–2464. [Google Scholar] [CrossRef] [Green Version]
- Amino Acids, Quantitative, Random, Urine. Available online: https://neurology.testcatalog.org/show/AAPD (accessed on 2 March 2020).
- Scalabre, A.; Jobard, E.; Demède, D.; Gaillard, S.; Pontoizeau, C.; Mouriquand, P.; Elena-Herrmann, B.; Mure, P.-Y. Evolution of Newborns’ Urinary Metabolomic Profiles According to Age and Growth. J. Proteome Res. 2017, 16, 3732–3740. [Google Scholar] [CrossRef]
- Shelley, H.J. Carbohydrate Reserves in the Newborn Infant. BMJ 1964, 1, 273–275. [Google Scholar] [CrossRef] [Green Version]
- Guneral, F.; Bachmann, C. Age-related reference values for urinary organic acids in a healthy Turkish pediatric population. Clin. Chem. 1994, 40, 862–868. [Google Scholar] [CrossRef]
- Shaw, W. Increased urinary excretion of a 3-(3-hydroxyphenyl)-3-hydroxypropionic acid (HPHPA), an abnormal phenylalanine metabolite ofClostridiaspp. in the gastrointestinal tract, in urine samples from patients with autism and schizophrenia. Nutr. Neurosci. 2010, 13, 135–143. [Google Scholar] [CrossRef]
- Gregersen, N.; Ingerslev, J.; Rasmussen, K. Low Molecular Weight Organic Acids in the Urine of the Newborn. Acta Paediatr. 1977, 66, 85–89. [Google Scholar] [CrossRef]
- Challis, J.; Newnham, J.P.; Petraglia, F.; Yeganegi, M.; Bocking, A. Fetal sex and preterm birth. Placenta 2013, 34, 95–99. [Google Scholar] [CrossRef]
- Caterino, M.; Ruoppolo, M.; Villani, G.R.D.; Marchese, E.; Costanzo, M.; Sotgiu, G.; Dore, S.; Franconi, F.; Campesi, I. Influence of Sex on Urinary Organic Acids: A Cross-Sectional Study in Children. Int. J. Mol. Sci. 2020, 21, 582. [Google Scholar] [CrossRef] [Green Version]
- Diaz, S.O.; Pinto, J.; Barros, A.; Morais, E.; Duarte, D.; Negrão, F.; Pita, C.; Almeida, M.D.C.; Carreira, I.M.; Spraul, M.; et al. Newborn Urinary Metabolic Signatures of Prematurity and Other Disorders: A Case Control Study. J. Proteome Res. 2015, 15, 311–325. [Google Scholar] [CrossRef] [Green Version]
- Pierre Martin, F.; Rezzi, S.; Lussu, M.; Pintus, R.; Pattumelli, M.G.; Noto, A.; Dessì, A.; Da Silva, L.; Collino, S.; Ciccarelli, S.; et al. Urinary metabolomics in term newborns delivered spontaneously or with cesarean section: Preliminary data. J. Pediatr. Neonatal Individ. Med. 2018, 7, 1–9. [Google Scholar] [CrossRef]
- Sachan, D.S.; Hoppel, C.L. Carnitine biosynthesis. Hydroxylation of N6-trimethyl-lysine to 3-hydroxy-N6-trimethyl-lysine. Biochem. J. 1980, 188, 529–534. [Google Scholar] [CrossRef] [Green Version]
- Shenai, J.P.; Borum, P.R. Tissue Carnitine Reserves of Newborn Infants. Pediatr. Res. 1984, 18, 679–681. [Google Scholar] [CrossRef] [Green Version]
- Pitt, J.J.; Eggington, M.; Kahler, S.G. Comprehensive Screening of Urine Samples for Inborn Errors of Metabolism by Electrospray Tandem Mass Spectrometry. Clin. Chem. 2002, 48, 1970–1980. [Google Scholar] [CrossRef]
- Hampe, M.H.; Panaskar, S.N.; Yadav, A.A.; Ingale, P.W. Gas chromatography/mass spectrometry-based urine metabolome study in children for inborn errors of metabolism: An Indian experience. Clin. Biochem. 2017, 50, 121–126. [Google Scholar] [CrossRef]
- Kennedy, A.D.; Miller, M.J.; Beebe, K.; Wulff, J.E.; Evans, A.M.; Miller, L.A.; Sutton, V.R.; Sun, Q.; Elsea, S.H. Metabolomic Profiling of Human Urine as a Screen for Multiple Inborn Errors of Metabolism. Genet. Test. Mol. Biomark. 2016, 20, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Almannai, M.; Alfadhel, M.; El-Hattab, A.W. Carnitine Inborn Errors of Metabolism. Molecules 2019, 24, 3251. [Google Scholar] [CrossRef] [Green Version]
- Lamari, F.; Mochel, F.; Sedel, F.; Saudubray, J.M. Disorders of phospholipids, sphingolipids and fatty acids biosynthesis: Toward a new category of inherited metabolic diseases. J. Inherit. Metab. Dis. 2012, 36, 411–425. [Google Scholar] [CrossRef]
- Lamari, F.; Mochel, F.; Saudubray, J.M. An overview of inborn errors of complex lipid biosynthesis and remodelling. J. Inherit. Metab. Dis. 2014, 38, 3–18. [Google Scholar] [CrossRef]
- Jazwinski, S.M.; Kim, S. Examination of the Dimensions of Biological Age. Front. Genet. 2019, 10, 263. [Google Scholar] [CrossRef]
- Chiu, C.-Y.; Yeh, K.-W.; Lin, G.; Chiang, M.-H.; Yang, S.-C.; Chao, W.-J.; Yao, T.-C.; Tsai, M.-H.; Hua, M.-C.; Liao, S.-L.; et al. Metabolomics Reveals Dynamic Metabolic Changes Associated with Age in Early Childhood. PLoS ONE 2016, 11, e0149823. [Google Scholar] [CrossRef] [Green Version]
- Jove, M.; Maté, I.; Naudí, A.; Mota-Martorell, N.; Portero-Otin, M.; De La Fuente, M.; Pamplona, R. Human Aging Is a Metabolome-related Matter of Gender. J. Gerontol. Ser. A Boil. Sci. Med. Sci. 2015, 71, 578–585. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S. Emerging Insights into the Metabolic Alterations in Aging Using Metabolomics. Metabolites 2019, 9, 301. [Google Scholar] [CrossRef] [Green Version]
- Saito, K.; Maekawa, K.; Kinchen, J.M.; Tanaka, R.; Kumagai, Y.; Saito, Y. Gender- and Age-Associated Differences in Serum Metabolite Profiles among Japanese Populations. Boil. Pharm. Bull. 2016, 39, 1179–1186. [Google Scholar] [CrossRef] [Green Version]
- Chaleckis, R.; Murakami, I.; Takada, J.; Kondoh, H.; Yanagida, M. Individual variability in human blood metabolites identifies age-related differences. Proc. Natl. Acad. Sci. USA 2016, 113, 4252–4259. [Google Scholar] [CrossRef] [Green Version]
- Thévenot, E.A.; Roux, A.; Xu, Y.; Ezan, E.; Junot, C. Analysis of the Human Adult Urinary Metabolome Variations with Age, Body Mass Index, and Gender by Implementing a Comprehensive Workflow for Univariate and OPLS Statistical Analyses. J. Proteome Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef]
- Slupsky, C.M.; Rankin, K.N.; Wagner, J.R.; Fu, H.; Chang, D.; Weljie, A.M.; Saude, E.J.; Lix, B.; Adamko, D.; Shah, S.; et al. Investigations of the Effects of Gender, Diurnal Variation, and Age in Human Urinary Metabolomic Profiles. Anal. Chem. 2007, 79, 6995–7004. [Google Scholar] [CrossRef]
- Psihogios, N.G.; Gazi, I.F.; Elisaf, M.; Seferiadis, K.I.; Bairaktari, E.T. Gender-related and age-related urinalysis of healthy subjects by NMR-based metabonomics. NMR Biomed. 2008, 21, 195–207. [Google Scholar] [CrossRef]
- Gu, H.; Pan, Z.; Xi, B.; Hainline, B.E.; Shanaiah, N.; Asiago, V.; Gowda, G.A.N.; Raftery, D. 1H NMR metabolomics study of age profiling in children. NMR Biomed. 2009, 22, 826–833. [Google Scholar] [CrossRef] [Green Version]
- Lau, C.-H.E.; Siskos, A.P.; Maitre, L.; Robinson, O.; Athersuch, T.J.; Want, E.; Urquiza, J.; Casas, M.; Vafeiadi, M.; Roumeliotaki, T.; et al. Determinants of the urinary and serum metabolome in children from six European populations. BMC Med. 2018, 16, 202. [Google Scholar] [CrossRef] [Green Version]
- Matos, V.; Drukker, A.; Guignard, J.-P. Spot urine samples for evaluating solute excretion in the first week of life. Arch. Dis. Child. Fetal Neonatal Ed. 1999, 80, F240–F242. [Google Scholar] [CrossRef] [Green Version]
- Timmerman, K.L.; Volpi, E. Amino acid metabolism and regulatory effects in aging. Curr. Opin. Clin. Nutr. Metab. Care 2008, 11, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Wu, G. Functional Amino Acids in Growth, Reproduction, and Health12. Adv. Nutr. 2010, 1, 31–37. [Google Scholar] [CrossRef]
- Hipkiss, A.R.; Cartwright, S.P.; Bromley, C.; Gross, S.; Bill, R.M. Carnosine: Can understanding its actions on energy metabolism and protein homeostasis inform its therapeutic potential? Chem. Central J. 2013, 7, 38. [Google Scholar] [CrossRef] [Green Version]
- Rutledge, R.B.; Smittenaar, P.; Zeidman, P.; Brown, H.R.; Adams, R.A.; Lindenberger, U.; Dayan, P.; Dolan, R.J. Risk Taking for Potential Reward Decreases across the Lifespan. Curr. Boil. 2016, 26, 1634–1639. [Google Scholar] [CrossRef] [Green Version]
- Rudman, D.; Kutner, M.H.; Chawla, R.K.; Goldsmith, M.A.; Blackston, R.D.; Bain, R. Serum and Urine Polyamines in Normal and in Short Children. J. Clin. Investig. 1979, 64, 1661–1668. [Google Scholar] [CrossRef]
- Petitclerc, C.; Solberg, H. Approved recommendation (1987) on the theory of reference values. Part 2. Selection of individuals for the production of reference values. Clin. Chim. Acta 1987, 170, S1–S11. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Newborns (N = 48) | |
| Sex | |
| Female | 17 (35%) |
| Male | 31 (65%) |
| Gestational age (weeks) | 38.2 ± 1.5 |
| Ave. Apgar at 1 min | 8 (100%) |
| Ave. Apgar at 5 min | 9 (100%) |
| Ave. Silverman–Anderson | 0 (100%) |
| Weight (g) | 2979 ± 472 |
| Weight (g) males | 2992 ± 86.59 |
| Weight (g) females | 2882 ± 112.4 |
| Mothers (N = 48) | |
| BMI (pre-gestational) | 27.1 ± 4.9 |
| Age (years) | 29 ± 7 |
| Resolution | |
| Vaginal delivery | 22 (45.8%) |
| Caesarean section | 26 (54.2%) |
| Euglycemic | 22 (45.8%) |
| Gestational Diabetes Mellitus (GDM) | 26 (54.2%) |
| Metabolite | HMDB ID | Mean ± SD (μM) | Mean ± SD (μM/mM Creatinine) | 2.5–97.5% Percentile (μM/mM Creatinine) |
|---|---|---|---|---|
| Histamine | HMDB0000870 | 0.08 ± 0.03 | 0.02 ± 0.01 | 0.01–0.04 |
| Putrescine | HMDB0001414 | 1.09 ± 1.54 | 0.31 ± 0.63 | 0.03–3.42 |
| Methionine sulfoxide | HMDB0002005 | 7.02 ± 4.70 | 1.60 ± 0.73 | 0.60–3.60 |
| N2-Acetylornithine | HMDB0003357 | 3.28 ± 4.00 | 0.67 ± 0.75 | 0.12–3.50 |
| Serotonin | HMDB0000259 | 0.98 ± 0.67 | 0.20 ± 0.06 | 0.10–0.35 |
| DOPA | HMDB0000181 | 0.20 ± 0.10 | 0.05 ± 0.03 | 0.01–0.15 |
| Asymmetric dimethylarginine | HMDB0001539 | 10.6 ± 6.40 | 2.31 ± 0.83 | 0.98–4.90 |
| Symmetric dimethylarginine | HMDB0003334 | 45.7 ± 30.8 | 9.80 ± 3.00 | 5.33–18.1 |
| Spermidine | HMDB0001257 | 0.25 ± 0.19 | 0.06 ± 0.05 | 0.02–0.30 |
| Spermine | HMDB0001256 | 0.23 ± 0.28 | 0.06 ± 0.11 | 0.01–0.62 |
| Diacetylspermine | HMDB0002172 | 4.64 ± 3.52 | 1.03 ± 0.59 | 0.37–3.11 |
| Trimethylamine N-oxide | HMDB0000925 | 59.4 ± 54.0 | 12.2 ± 10.3 | 0.30–43.1 |
| p-Hydroxyhippuric acid | HMDB0013678 | 37.1 ± 24.8 | 8.03 ± 3.48 | 4.20–19.6 |
| LysoPC a C16:1 | HMDB0010383 | 0.013 ± 0.011 | 0.003 ± 0.003 | 0.0001–0.0150 |
| LysoPC a C16:0 | HMDB0010382 | 0.19 ± 0.20 | 0.043 ± 0.036 | 0.007–0.169 |
| LysoPC a C17:0 | HMDB0012108 | 0.018 ± 0.013 | 0.005 ± 0.001 | 0.0008–0.0582 |
| LysoPC a C18:2 | HMDB0010386 | 0.03 ± 0.04 | 0.007 ± 0.007 | 0.001–0.030 |
| LysoPC a C18:0 | HMDB0010384 | 0.06 ± 0.06 | 0.015 ± 0.012 | 0.0008–0.0652 |
| LysoPC a C20:4 | HMDB0010395 | 0.02 ± 0.04 | 0.005 ± 0.006 | 0.0–0.03 |
| PC ae C36:0 | HMDB0013406 | 0.02 ± 0.03 | 0.006 ± 0.013 | 0.0–0.08 |
| PC aa C36:6 | HMDB0008206 | 0.007 ± 0.006 | 0.002 ± 0.003 | 0.0–0.02 |
| PC aa C36:0 | HMDB0007886 | 0.05 ± 0.06 | 0.013 ± 0.023 | 0.001–0.130 |
| PC aa C38:6 | HMDB0008116 | 0.08 ± 0.12 | 0.017 ± 0.020 | 0.002–0.089 |
| PC aa C38:0 | HMDB0007893 | 0.08 ± 0.04 | 0.022 ± 0.021 | 0.005–0.124 |
| PC ae C40:6 | HMDB0013422 | 0.02 ± 0.02 | 0.005 ± 0.004 | 0.001–0.021 |
| PC aa C40:6 | HMDB0008057 | 0.04 ± 0.05 | 0.010 ± 0.013 | 0.0–0.07 |
| SM(OH) C14:1 | HMDB0013462 | 10.1 ± 0.09 | 0.02 ± 0.04 | 0.00–0.25 |
| SM C16:1 | HMDB0013464 | 0.10 ± 0.11 | 0.02 ± 0.02 | 0.0–0.11 |
| SM C16:0 | HMDB0010168 | 2.02 ± 2.28 | 0.50 ± 0.92 | 0.07–5.31 |
| SM(OH) C16:1 | HMDB0013463 | 0.06 ± 0.08 | 0.02 ± 0.04 | 0.0–0.24 |
| SM C18:1 | HMDB0012101 | 0.08 ± 0.10 | 0.02 ± 0.02 | 0.0–0.06 |
| SM C18:0 | HMDB0012087 | 0.31 ± 0.37 | 0.07 ± 0.79 | 0.01–0.43 |
| SM C20:2 | HMDB0013465 | 0.005 ± 0.005 | 0.001 ± 0.001 | 0.0–0.004 |
| SM(OH) C22:2 | HMDB0013467 | 0.03 ± 0.04 | 0.01 ± 0.01 | 0.0–0.05 |
| SM(OH) C22:1 | HMDB0013466 | 0.16 ± 0.15 | 0.04 ± 0.03 | 0.0–0.15 |
| SM(OH) C24:1 | HMDB0013469 | 0.04 ± 0.05 | 0.01 ± 0.02 | 0.0–0.14 |
| Carnitine (C0) | HMDB0000062 | 8.96 ± 7.07 | 2.01 ± 1.13 | 0.79–5.67 |
| l-Acetylcarnitine (C2) | HMDB0000201 | 4.49 ± 4.32 | 0.89 ± 0.38 | 0.38–1.86 |
| Propenoylcarnitine (C3:1) | HMDB0013124 | 0.08 ± 0.08 | 0.02 ± 0.01 | 0.0–0.06 |
| Propionylcarnitine (C3) | HMDB0000824 | 0.14 ± 0.09 | 0.03 ± 0.02 | 0.01–0.07 |
| Butenylcarnitine (C4:1) | HMDB0013126 | 0.09 ± 0.04 | 0.02 ± 0.01 | 0.01–0.04 |
| Butyrylcarnitine (C4) | HMDB0002013 | 0.53 ± 0.39 | 0.12 ± 0.07 | 0.05–0.33 |
| Hydroxypropionyl carnitine (C3OH) | HMDB0013125 | 0.11 ± 0.05 | 0.03 ± 0.01 | 0.01–0.06 |
| Tiglylcarnitine (C5:1) | HMDB0002366 | 0.34 ± 0.23 | 0.07 ± 0.03 | 0.03–0.16 |
| Hydroxybutyryl carnitine (C4OH) | HMDB0002095 | 0.18 ± 0.10 | 0.04 ± 0.02 | 0.01–0.08 |
| Hexenoylcarnitine (C6:1) | HMDB0013161 | 0.06 ± 0.03 | 0.020 ± 0.004 | 0.01–0.02 |
| Hexanoylcarnitine (C6) | HMDB0000756 | 0.12 ± 0.07 | 0.03 ± 0.08 | 0.02–0.05 |
| Hydroxyvalerylcarnitine (C5OH) | HMDB0013132 | 0.36 ± 0.20 | 0.08 ± 0.05 | 0.04–0.29 |
| Glutaconylcarnitine (C5:1DC) | HMDB0013129 | 0.07 ± 0.04 | 0.02 ± 0.01 | 0.0–0.03 |
| Glutarylcarnitine (C5DC) | HMDB0013130 | 0.20 ± 0.11 | 0.05 ± 0.02 | 0.02–0.09 |
| Octanoylcarnitine (C8) | HMDB0000791 | 0.24 ± 0.19 | 0.05 ± 0.05 | 0.02–0.31 |
| Methylglutarylcarnitine (C5MDC) | HMDB0000552 | 0.27 ± 0.16 | 0.06 ± 0.02 | 0.03–0.13 |
| Nonaylcarnitine (C9) | HMDB0013288 | 0.46 ± 0.38 | 0.09 ± 0.04 | 0.03–0.18 |
| Pimelylcarnitine (C7DC) | HMDB0013328 | 0.21 ± 0.14 | 0.05 ± 0.02 | 0.02–0.09 |
| Decenoylcarnitine (C10:1) | HMDB0013205 | 0.34 ± 0.12 | 0.09 ± 0.04 | 0.03–0.20 |
| Decanoylcarnitine (C10) | HMDB0000651 | 0.29 ± 0.18 | 0.06 ± 0.02 | 0.04–0.13 |
| Dodecenoylcarnitine (C12:1) | HMDB0013326 | 0.24 ± 0.10 | 0.06 ± 0.02 | 0.02–0.14 |
| Dodecanoylcarnitine (C12) | HMDB0002250 | 0.35 ± 0.27 | 0.08 ± 0.04 | 0.03–0.25 |
| Tetradecadienyl carnitine (C14:2) | HMDB0013331 | 0.05 ± 0.03 | 0.010 ± 0.003 | 0.01–0.02 |
| Tetradecenoylcarnitine (C14:1) | HMDB0013329 | 0.06 ± 0.03 | 0.01 ± 0.02 | 0.0–0.1 |
| Tetradecanoylcarnitine (C14) | HMDB0005066 | 0.11 ± 0.10 | 0.02 ± 0.02 | 0.01–0.10 |
| Hydroxytetradecadienylcarnitine (C14:2OH) | HMDB0013332 | 0.03 ± 0.02 | 0.007 ± 0.002 | 0.003–0.014 |
| Hydroxytetradecenoyl carnitine (C14:1OH) | HMDB0013330 | 0.04 ± 0.02 | 0.008 ± 0.002 | 0.004–0.015 |
| Hexadecadienyl carnitine (C16:2) | HMDB0013334 | 0.02 ± 0.01 | 0.004 ± 0.002 | 0.0–0.01 |
| Hexadecanoylcarnitine (C16) | HMDB0000222 | 0.05 ± 0.03 | 0.01 ± 0.01 | 0.01–0.06 |
| Octadecadienylcarnitine (C18:2) | HMDB0006469 | 0.010 ± 0.003 | 0.003 ± 0.001 | 0.0–0.01 |
| Metabolite | HMDB ID | Mean ± SD (μM) | Mean ± SD (μM/mM Creatinine) | 2.5–97.5% Percentile (μM/mM Creatinine) |
|---|---|---|---|---|
| LysoPC a C14:0 | HMDB0010379 | 0.02 ± 0.02 | 0.006 ± 0.003 | 0.002–0.012 |
| LysoPC a C18:1 | HMDB0002815 | 0.07 ± 0.09 | 0.02 ± 0.03 | 0.0–0.20 |
| LysoPC a C20:3 | HMDB0010394 | 0.014 ± 0.002 | 0.004 ± 0.003 | 0.0–0.01 |
| LysoPC a C24:0 | HMDB0010405 | 0.07 ± 0.02 | 0.02 ± 0.01 | 0.0–0.05 |
| LysoPC a C26:1 | HMDB0029220 | 0.01 ± 0.01 | 0.002 ± 0.002 | 0.0–0.01 |
| LysoPC a C26:0 | HMDB0029205 | 0.01 ± 0.01 | 0.004 ± 0.005 | 0.0–0.03 |
| LysoPC a C28:1 | HMDB0029221 | 0.01 ± 0.01 | 0.002 ± 0.002 | 0.0–0.01 |
| LysoPC a C28:0 | HMDB0029206 | 0.04 ± 0.01 | 0.011 ± 0.007 | 0.0–0.04 |
| PC aa C32:2 | HMDB0007874 | 0.03 ± 0.03 | 0.01 ± 0.01 | 0.0–0.03 |
| PC aa C40:2 | HMDB0008276 | 0.01 ± 0.01 | 0.003 ± 0.008 | 0.0–0.05 |
| PC aa C40:1 | HMDB0007993 | 0.03 ± 0.04 | 0.01 ± 0.02 | 0.0–0.13 |
| Decadienylcarnitine (C10:2) | HMDB0013325 | 0.14 ± 0.07 | 0.03 ± 0.01 | 0.01–0.07 |
| Dodecanedioylcarnitine (C12DC) | HMDB0013327 | 0.27 ± 0.29 | 0.05 ± 0.05 | 0.01–0.29 |
| Hexadecenoylcarnitine (C16:1) | HMDB0006317 | 0.05 ± 0.02 | 0.01 ± 0.01 | 0.0–0.03 |
| Hydroxyhexadecadienylcarnitine (C16:2OH) | HMDB0013335 | 0.02 ± 0.01 | 0.004 ± 0.001 | 0.0–0.01 |
| Hydroxyhexadecenoyl carnitine (C16:1OH) | HMDB0013333 | 0.04 ± 0.02 | 0.010 ± 0.004 | 0.0–0.02 |
| Hydroxyhexadecanoylcarnitine (C16OH) | HMDB0061642 | 0.04 ± 0.02 | 0.008 ± 0.003 | 0.0–0.02 |
| Octadecenoylcarnitine (C18:1) | HMDB0006464 | 0.02 ± 0.01 | 0.005 ± 0.004 | 0.0–0.03 |
| Octadecanoylcarnitine (C18) | HMDB0000848 | 0.03 ± 0.03 | 0.01 ± 0.01 | 0.0–0.07 |
| Hydroxyoctadecenoylcarnitine (C18:1OH) | HMDB0013339 | 0.02 ± 0.01 | 0.004 ± 0.002 | 0.0–0.01 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Hernández, Y.; Oropeza-Valdez, J.J.; Blanco-Sandate, J.O.; Herrera-Van Oostdam, A.S.; Zheng, J.; Chi Guo, A.; Lima-Rogel, V.; Rajabzadeh, R.; Salgado-Bustamante, M.; Adrian-Lopez, J.; et al. The Urinary Metabolome of Healthy Newborns. Metabolites 2020, 10, 165. https://doi.org/10.3390/metabo10040165
López-Hernández Y, Oropeza-Valdez JJ, Blanco-Sandate JO, Herrera-Van Oostdam AS, Zheng J, Chi Guo A, Lima-Rogel V, Rajabzadeh R, Salgado-Bustamante M, Adrian-Lopez J, et al. The Urinary Metabolome of Healthy Newborns. Metabolites. 2020; 10(4):165. https://doi.org/10.3390/metabo10040165
Chicago/Turabian StyleLópez-Hernández, Yamilé, Juan José Oropeza-Valdez, Jorge O. Blanco-Sandate, Ana Sofia Herrera-Van Oostdam, Jiamin Zheng, An Chi Guo, Victoria Lima-Rogel, Rahmatollah Rajabzadeh, Mariana Salgado-Bustamante, Jesus Adrian-Lopez, and et al. 2020. "The Urinary Metabolome of Healthy Newborns" Metabolites 10, no. 4: 165. https://doi.org/10.3390/metabo10040165