Photostimulation of Extravasation of Beta-Amyloid through the Model of Blood-Brain Barrier

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subject
2.2. The Injected Model of AD
2.3. The Protocol of PS Course In Vivo and In Vitro Experiments
2.4. Memory and Neurological Tests
2.5. The BBB Model In Vitro
2.6. Measurement of BBB Permeability
2.7. Analysis of Level of Aβ in the Tested Tissues
2.8. Measure the Termal Impact of PS
2.9. Statistical Analysis
3. Results
3.1. PS-Therapeutic Effects on the Memory and Neurological Deficit in Mice with AD
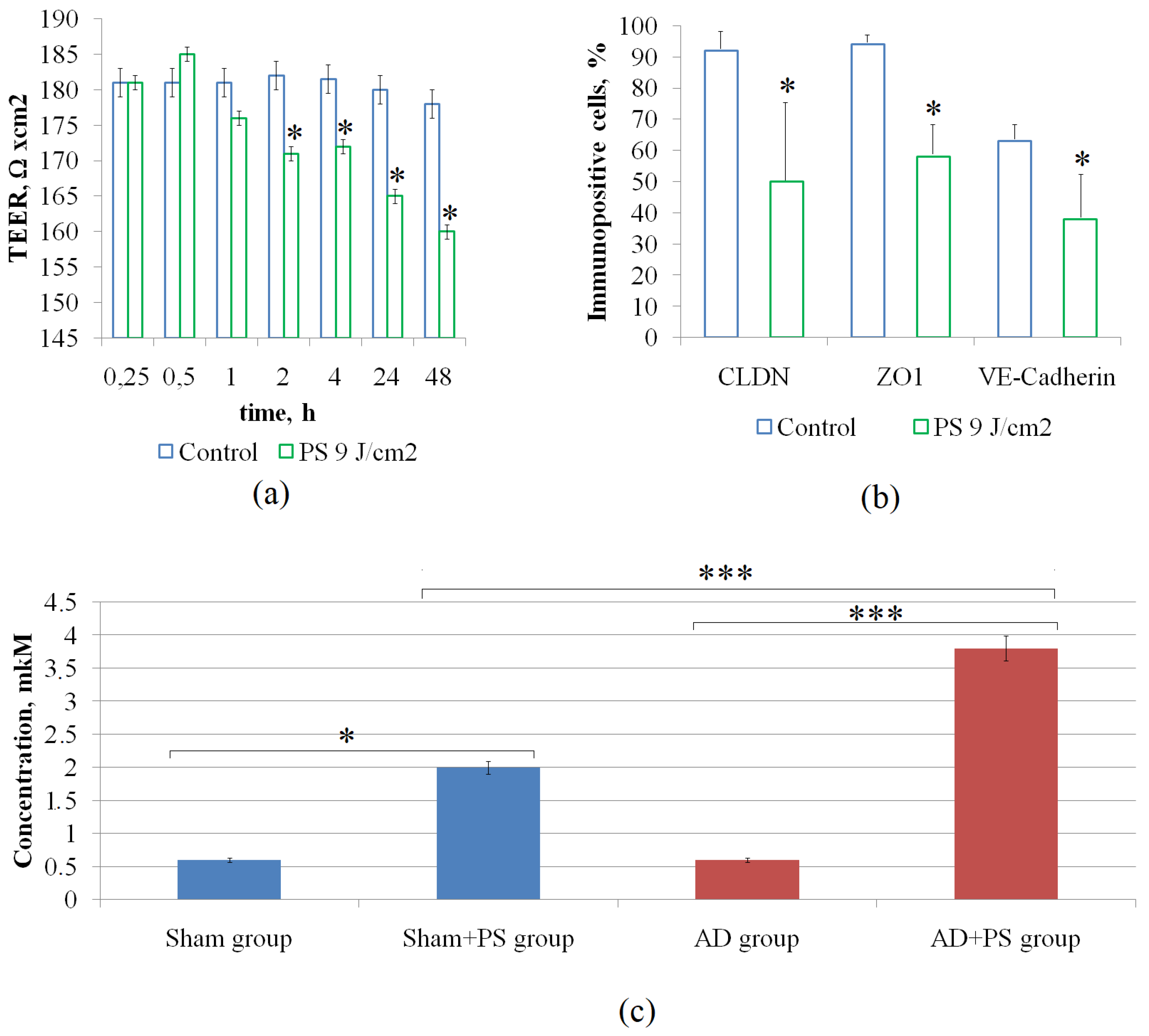
3.2. PS-Course Causes Clearance of Aβ from the Brain in Mice with AD
3.3. PS-Mediated Increase in the BBB Permeability
3.4. The Termal Impact of PS
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Karran, E.; De Strooper, B. The amyloid cascade hypothesis: Are we poised for success or failure? J. Neurochem. 2016, 139, 237–252. [Google Scholar] [CrossRef]
- Sevigny, J.; Chiao, P.; Bussière, T.; Weinreb, P.; Williams, L.; Maier, M.; Dunstan, R.; Salloway, S.; Chen, T.; Ling, Y.; et al. The antibody aducanumab reduces Aβ plaques in Alzheimer’s disease. Nature 2016, 537, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Zinchenko, E.M.; Navolokin, N.A.; Shirokov, A.A.; Khlebcov, B.N.; Dubrovsky, A.I.; Saranceva, E.I.; Abdurashitov, A.S.; Khorovodov, A.P.; Terskov, A.V.; Mamedova, A.T.; et al. Pilot study of transcranial photobiomodulation of lymphatic clearance of beta-amyloid from the mouse brain: Breakthrough strategies for nonpharmacologic therapy of Alzheimer’s disease. Biomed. Opt. Express 2019, 10, 4003–4017. [Google Scholar] [CrossRef] [PubMed]
- Lane, N. Cell biology: Power games. Nature 2006, 443, 901–903. [Google Scholar] [CrossRef] [PubMed]
- Grillo, S.L.; Duggett, N.; Ennaceur, A.; Chazot, P. Non-invasive infra-red therapy (1072 nm) reduces β-amyloid protein levels in the brain of an Alzheimer’s disease mouse model, TASTPM. J. Photochem. Photobiol. B 2013, 123, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Wang, R.; Dong, Y.; Tucker, D.; Zhao, N.; Ahmed, E.; Zhu, L.; Liu, T.C.-Y.; Cohen, R.M.; Zhang, Q. Low-level laser therapy for beta amyloid toxicity in rat hippocampus. Neurobiol. Aging 2017, 49, 165–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Taboada, L.; Yu, J.; El-Amouri, S.; Gattoni-Celli, S.; Richieri, S.; McCarthy, T.; Streeter, J.; Kindy, M. Transcranial laser therapy attenuates amyloid-beta peptide neuropathology in amyloid-beta protein precursor transgenic mice. J. Alzheimer Dis. 2011, 23, 521–535. [Google Scholar] [CrossRef] [PubMed]
- Purushothuman, S.; Johstone, D.M.; Nandasena, C.; Van Eersel, J.; Ittner, L.M.; Stone, J. Near infrared light mitigates cerebellar pathology in transgenic mouse models of dementia. Neurosci. Lett. 2015, 591, 155–159. [Google Scholar] [CrossRef]
- Gonzalez-Lima, F.; Barksdale, B.R.; Rojas, J.C. Mitochondrial respiration as a target for neuroprotection and cognitive enhancement. Biochem. Pharmacol. 2014, 88, 584–593. [Google Scholar] [CrossRef]
- Grimm, A.; Friedland, K.; Eckert, A. Mitochondrial dysfunction: The missing link between aging and sporadic Alzheimer’s disease. Biogerontology 2016, 17, 281–296. [Google Scholar] [CrossRef]
- Da Mesquita, S.; Louveau, A.; Vaccari, A.; Smirnov, I.; Cornelison, R.C.; Kingsmore, K.M.; Contarino, C.; Onengut-Gumuscu, S.; Farber, E.; Raper, D.; et al. Functional aspects of meningeal lymphatics in ageing and Alzheimer’s disease. Nature 2018, 560, 185–191. [Google Scholar] [CrossRef]
- Semyachkina-Glushkovskaya, O.V.; Abdurashitov, A.S.; Dubrovsky, A.I.; Klimova, M.M.; Agranovich, I.M.; Terskov, A.V.; Shirokov, A.A.; Vinnik, V.V.; Kuznecova, A.S.; Lezhnev, N.D.; et al. Photobiomodulation of lymphatic drainage and clearance: Perspective strategy for augmentation of meningeal lymphatic functions. Biomed. Opt. Express 2020, 11, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Semyachkina-Glushkovskaya, O.V.; Abdurashitov, A.S.; Klimova, M.M.; Dubrovsky, A.I.; Shirokov, A.A.; Fomin, A.S.; Terskov, A.V.; Agranovich, I.M.; Mamedova, A.T.; Khorovodov, A.P.; et al. Photobiostimulation of cerebral and peripheral lymphatic functions. Transl. Biophotonics 2020, 2, e201900036. [Google Scholar] [CrossRef]
- Sokolovski, S.G.; Zolotovskaya, S.A.; Goltsov, A.; Pourreyron, C.; South, A.P.; Rafailov, E.U. Infrared laser pulsetriggers increased singlet oxygen production in tumourcells. Sci. Rep. 2013, 3, 3484. [Google Scholar] [CrossRef] [PubMed]
- Semyachkina-Glushkovskaya, O.V.; Chehonin, V.P.; Borisova, E.G.; Fedosov, I.V.; Namykin, A.A.; Abdurashitov, A.S.; Shirokov, A.A.; Khlebtsov, B.N.; Lyubun, E.V.; Navolokin, N.A.; et al. Photodynamic opening of the blood-brain barrier and pathways of brain clearing pathways. J. Biophotonics 2018, 11, e201700287. [Google Scholar] [CrossRef]
- Semyachkina-Glushkovskaya, O.V.; Abdurashitov, A.S.; Dubrovsky, A.I.; Bragin, D.E.; Bragina, O.A.; Shushunova, N.A.; Maslyakova, G.N.; Navolokin, N.A.; Bucharskaya, A.B.; Tuchin, V.V.; et al. Application of optical coherent tomography for in vivo monitoring of the meningeal lymphatic vessels during opening of blood-brain barrier: Mechanisms of brain clearing. J. Biomed. Opt. 2017, 22, 121719. [Google Scholar] [CrossRef] [Green Version]
- Semyachkina-Glushkovskaya, O.V.; Postnov, D.E.; Kurths, J. Blood–Brain Barrier, Lymphatic Clearance, and Recovery: Ariadne’s Thread in Labyrinths of Hypotheses. Int. J. Mol. Sci. 2018, 19, 3818. [Google Scholar] [CrossRef] [Green Version]
- Jordão, J.F.; Thévenot, E.; Markham-Coultes, K.; Scarcelli, T.; Weng, Y.-Q.; Xhima, K.; O’Reilly, M.; Huang, Y.; McLaurin, J.; Hynynen, K.; et al. Amyloid-β plaque reduction, endogenous antibody delivery and glial activation by brain-targeted, transcranial focused ultrasound. Exp. Neurol. 2013, 248, 16–29. [Google Scholar] [CrossRef] [Green Version]
- Leinenga, G.; Götz, J. Scanning ultrasound removes amyloid-β and restores memory in an Alzheimer’s disease mouse model. Sci. Transl. Med. 2015, 7, 278ra33. [Google Scholar] [CrossRef] [Green Version]
- Burgess, A.; Dubey, S.; Yeung, S.; Hough, O.; Eterman, N.; Aubert, I.; Hynynen, K. Alzheimer disease in a mouse model: MR imaging-guided focused ultrasound targeted to the hippocampus opens the blood-brain barrier and improves pathologic abnormalities and behavior. Radiology 2014, 273, 736–745. [Google Scholar] [CrossRef] [Green Version]
- Lipsman, N.; Meng, Y.; Bethune, A.J.; Huang, Y.; Lam, B.; Masellis, M.; Herrmann, N.; Heyn, C.; Aubert, I.; Boutet, A.; et al. Blood–brain barrier opening in Alzheimer’s disease using MR-guided focused ultrasound. Nat. Commun. 2018, 9, 2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Facchinetti, R.; Bronzuoli, M.R.; Scuderi, C. An Animal Model of Alzheimer Disease Based on the Intrahippocampal Injection of Amyloid β-Peptide (1–42). Methods Mol. Biol. 2018, 1727, 343–352. [Google Scholar] [CrossRef]
- Ahn, J.H.; Cho, H.; Kim, J.H.; Kim, S.H.; Ham, J.S.; Park, I.; Suh, S.H.; Hong, S.P.; Song, J.H.; Hong, Y.K.; et al. Meningeal lymphatic vessels at the skull base drain cerebrospinal fluid. Nature 2019, 572, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Bevins, R.A.; Besheer, J. Object recognition in rats and mice: A one-trial nonmatching-to-sample learning task to study ‘recognition memory’. Nat. Protoc. 2006, 1, 1306–1311. [Google Scholar] [CrossRef] [PubMed]
- Garberg, P.; Ball, M.; Borg, N.; Cecchelli, R.; Fenart, L.; Hurst, R.D.; Lindmark, T.; Mabondzo, A.; Nilsson, J.E.; Raub, T.J.; et al. In Vitro Models for the Blood-Brain Barrier. Toxicol. Vitr. 2005, 19, 299–334. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, I.; Fazakas, C.; Krizbai, I.A. In Vitro Models of the Blood-Brain Barrier. Acta Neurobiol. Exp. Wars 2011, 71, 113–128. [Google Scholar]
- Liu, Y.; Xue, Q.; Tang, Q.; Hou, M.; Qi, H.; Chen, G.; Chen, W.; Zhang, J.; Chen, Y.; Xu, X.-Y. A simple method for isolating and culturing the rat brain microvascular endothelial cells. Microvasc. Res. 2013, 90, 199–205. [Google Scholar] [CrossRef]
- Khilazheva, E.D.; Boytsova, E.B.; Pozhilenkova, E.A.; Solonchuk, Y.R.; Salmina, A.B. Obtaining a three-cell model of a neurovascular unit in vitro. Cell Tissue Biol. 2015, 9, 447–451. [Google Scholar] [CrossRef]
- Louveau, A.; Smirnov, I.; Keyes, T.J.; Eccles, J.D.; Rouhani, S.J.; Peske, J.D.; Derecki, N.C.; Castle, D.; Mandell, J.W.; Lee, K.S.; et al. Structural and functional features of central nervous system lymphatic vessels. Nature 2015, 523, 337–341. [Google Scholar] [CrossRef]
- Aspelund, A.; Antila, S.; Proulx, S.T.; Karlsen, T.V.; Karaman, S.; Detmar, M.; Wiig, H.; Alitalo, K. A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J. Exp. Med. 2015, 212, 991–999. [Google Scholar] [CrossRef]
- De Boer, A.G.; Gaillard, P. The blood-brain barrier and drug transport to the brain. STP Pharma Sci. 2002, 12, 229–234. [Google Scholar]
- Greene, C.; Campbell, M. Tight junction modulation of the blood brain barrier: CNS delivery of small molecules. Tissue Barriers 2016, 4, e1138017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Z.; Qiao, P.-F.; Wan, C.-Q.; Cai, M.; Zhou, N.-K.; Li, Q. Role of Blood-Brain Barrier in Alzheimer’s Disease. J. Alzheimer Dis. 2018, 63, 1223–1234. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, H.M.; Dionne, V.E. Temperature dependence of ion permeation at the endplate channel. J. Gen. Physiol. 1983, 81, 687–703. [Google Scholar] [CrossRef] [Green Version]
- Moser, E.; Mathiesen, I.; Andersen, P. Association between brain temperature and dentate field potentials in exploring and swimming rats. Science 1993, 259, 1324–1326. [Google Scholar] [CrossRef]
- Faucher, P.; Mons, N.; Micheau, J.; Louis, C.; Beracochea, D.J. Hippocampal Injections of Oligomeric Amyloid β-peptide (1–42) Induce Selective Working Memory Deficits and Long-lasting Alterations of ERK Signaling Pathway. Front. Aging Neurosci. 2016, 7, 245. [Google Scholar] [CrossRef]
- Terry, R.D.; Masliah, E.; Salmon, D.P.; Butters, N.; DeTeresa, R.; Hill, R.; Hansen, L.A.; Katzman, R. Physical basis of cognitive alterations in Alzheimer’s disease: Synapse loss is the major correlate of cognitive impairment. Ann. Neurol. 1991, 30, 572–580. [Google Scholar] [CrossRef]
- Andreasen, N.; Blennow, K. CSF biomarkers for mild cognitive impairment and early Alzheimer’s disease. Clin. Neurol. Neurosurg. 2005, 107, 165–173. [Google Scholar] [CrossRef]
- Nordberg, A. Amyloid imaging in Alzheimer’s disease. Neuropsychologia 2008, 46, 1636–1641. [Google Scholar] [CrossRef]
- Villemagne, V.L.; Pike, K.E.; Darby, D.; Maruff, P.; Savage, G.; Ng, S.; Ackermann, U.; Cowie, T.F.; Currie, J.; Chan, S.G.; et al. Abeta deposits in older non-demented individuals with cognitive decline are indicative of preclinical Alzheimer’s disease. Neuropsychologia 2008, 46, 1688–1697. [Google Scholar] [CrossRef]
- Cleary, J.P.; Walsh, D.M.; Hofmeister, J.J.; Shankar, G.M.; Kuskowski, M.A.; Selkoe, D.J.; Ashe, K.H. Natural oligomers of the amyloid-beta protein specifically disrupt cognitive function. Nat. Neurosci. 2005, 8, 79–84. [Google Scholar] [CrossRef]
- Haass, C.; Selkoe, D. Soluble protein oligomers in neurodegeneration: Lessons from the Alzheimer’s amyloid beta-peptide. Nat. Rev. Mol. Cell Biol. 2007, 8, 101–112. [Google Scholar] [CrossRef]
- Benilova, I.; Karran, E.; De Strooper, B. The toxic Abeta oligomer and Alzheimer’s disease: An emperor in need of clothes. Nat. Neurosci. 2012, 15, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Mucke, L.; Selkoe, D. Neurotoxicity of amyloid beta-protein: Synaptic and network dysfunction. Cold Spring Harb. Perspect. Med. 2012, 27, a006338. [Google Scholar] [CrossRef] [Green Version]
- Chambon, C.; Wegener, N.; Gravius, A.; Danysz, W. Behavioural and cellular effects of exogenous amyloid-beta peptides in rodents. Behav. Brain Res. 2011, 225, 623–641. [Google Scholar] [CrossRef] [PubMed]
- Murakami, H.; Takanaga, H.; Matsuo, H.; Ohtani, H.; Sawada, Y. Comparison of Blood-Brain Barrier Permeability in Mice and Rats Using in Situ Brain Perfusion Technique. Am. J. Physiol. Heart Circ. Physiol. 2000, 279, H1022–H1028. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zinchenko, E.; Klimova, M.; Mamedova, A.; Agranovich, I.; Blokhina, I.; Antonova, T.; Terskov, A.; Shirokov, A.; Navolokin, N.; Morgun, A.; et al. Photostimulation of Extravasation of Beta-Amyloid through the Model of Blood-Brain Barrier. Electronics 2020, 9, 1056. https://doi.org/10.3390/electronics9061056
Zinchenko E, Klimova M, Mamedova A, Agranovich I, Blokhina I, Antonova T, Terskov A, Shirokov A, Navolokin N, Morgun A, et al. Photostimulation of Extravasation of Beta-Amyloid through the Model of Blood-Brain Barrier. Electronics. 2020; 9(6):1056. https://doi.org/10.3390/electronics9061056
Chicago/Turabian StyleZinchenko, Ekaterina, Maria Klimova, Aysel Mamedova, Ilana Agranovich, Inna Blokhina, Tatiana Antonova, Andrey Terskov, Alexander Shirokov, Nikita Navolokin, Andrey Morgun, and et al. 2020. "Photostimulation of Extravasation of Beta-Amyloid through the Model of Blood-Brain Barrier" Electronics 9, no. 6: 1056. https://doi.org/10.3390/electronics9061056