Preliminary Assessment of Microbial Community Structure of Wind-Tidal Flats in the Laguna Madre, Texas, USA

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
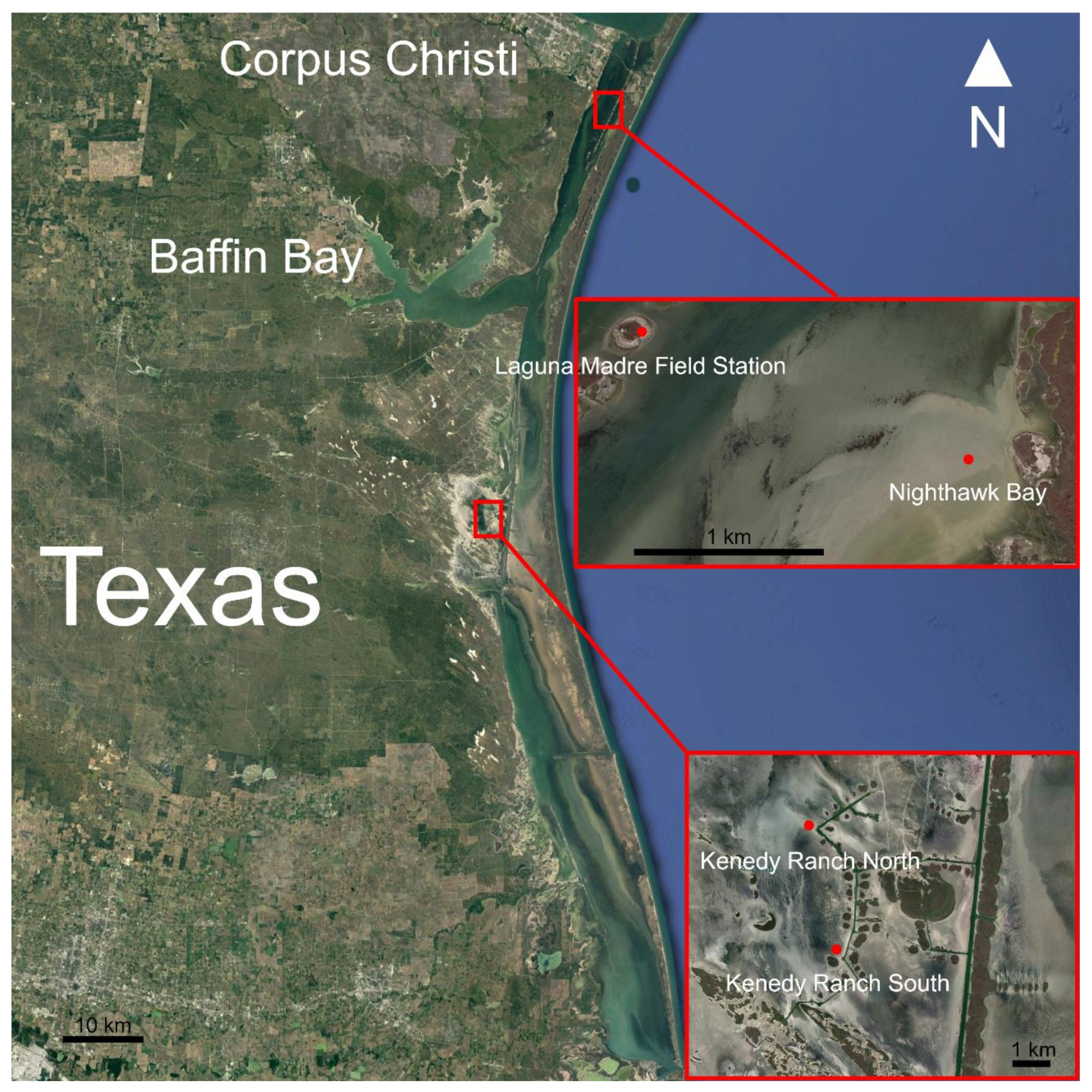
2.1. Study Site
2.2. Microscopy
2.3. Molecular Methods and Sequencing
2.4. Mat Cyanotoxin Analysis
3. Results
3.1. Microscopic Based Taxonomic Structure of the Mat
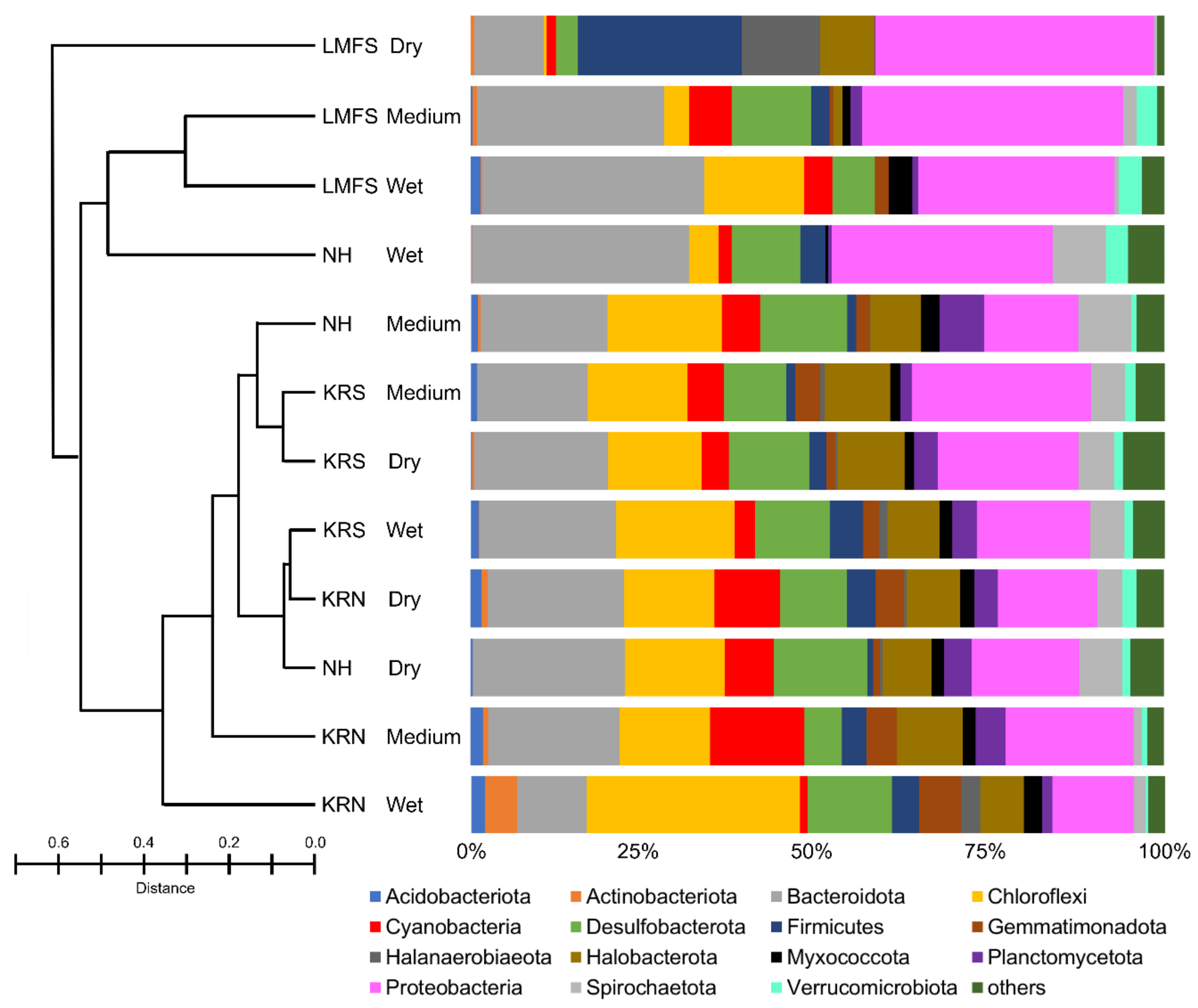
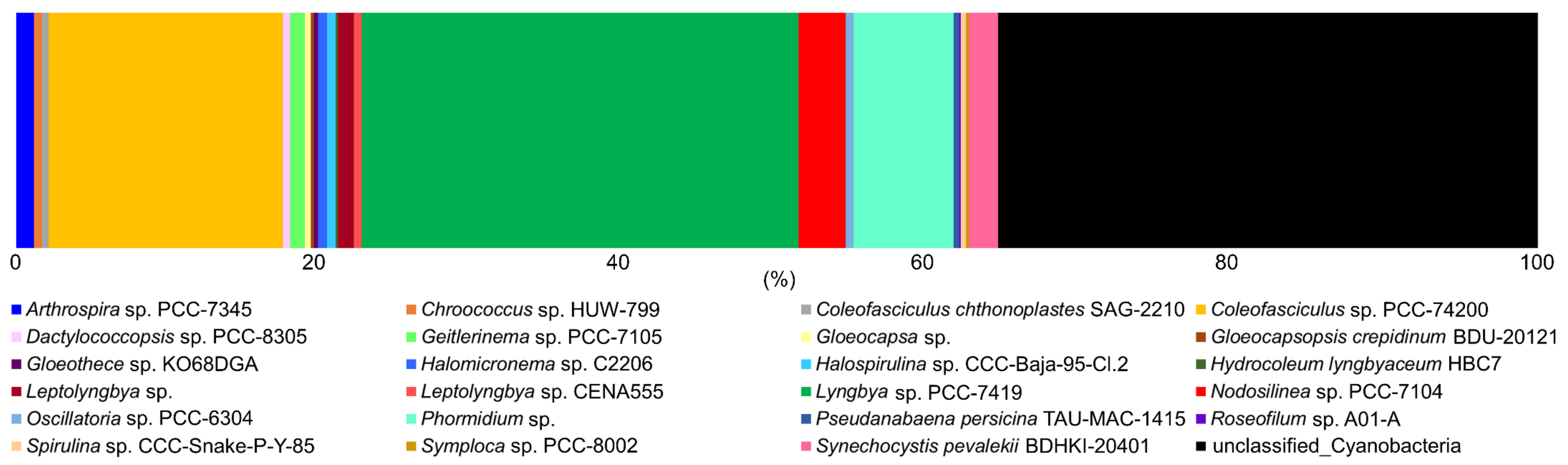
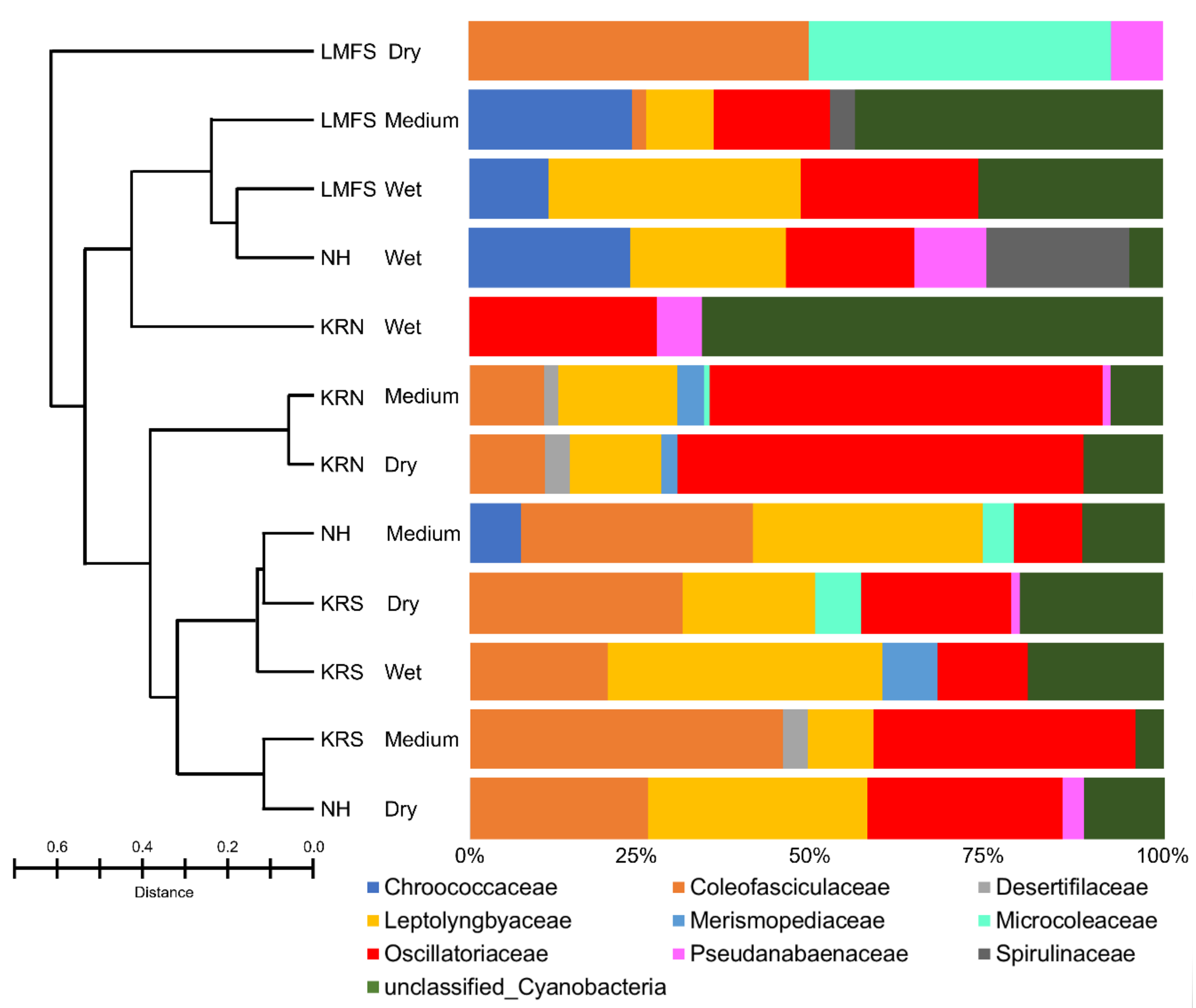
3.2. Molecular and Phylogenetic Analysis
3.3. Cyanobacterial Bioactive Metabolite Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Belnap, J. The world at your feet: Desert biological soil crusts. Front. Ecol. Environ. 2003, 1, 181–189. [Google Scholar] [CrossRef]
- Williams, A.; Kane, D.A.; Ewing, P.M.; Atwood, L.W.; Jilling, A.; Li, M.; Lou, Y.; Davis, A.S.; Grandy, A.S.; Huerd, S.C.; et al. Soil functional zone management: A vehicle for enhancing production and soil ecosystem services in row-crop agroecosystems. Front. Plant Sci. 2016, 7, 65. [Google Scholar] [CrossRef] [PubMed]
- Dudgeon, D. The functional significance of selection of particles by aquatic animals during building behavior. In The Biology of Particles in Aquatic Systems, 2nd ed.; Wooten, R.S., Ed.; Lewis: Boca Raton, FL, USA, 1994; pp. 289–312. [Google Scholar]
- Allison, D.G.; Sutherland, L.W. Role of exopolysaccharides in adhesion of freshwater bacteria. J. Gen. Microbiol. 1987, 133, 1319–1327. [Google Scholar] [CrossRef] [Green Version]
- Evans, R.D.; Johansen, J.R. Microbiotic crusts and ecosystem processes. Crit. Rev. Plant Sci. 1999, 18, 183–225. [Google Scholar] [CrossRef]
- Osorio-Santos, K.; Pietrasiak, N.; Bohunická, M.; Miscoe, L.H.; Kováčik, L.; Martin, M.P.; Johansen, J.R. Seven new species of Oculatella (Pseudanabaenales, Cyanobacteria): Taxonomically recognizing cryptic diversification. Eur. J. Phycol. 2014, 49, 450–470. [Google Scholar] [CrossRef] [Green Version]
- Castenholz, R.W. Composition of hot spring microbial mats: A summary. In Microbial Mats Stromatolites; Cohen, Y., Castenholz, R.W., Halvorson, H.O., Alan, R., Eds.; Liss Publ.: New York, NY, USA, 1984; pp. 101–119. [Google Scholar]
- Oremland, R.S.; Des Marms, D.J. Distribution, abundance and carbon isotopic composition of gaseous hydrocarbons in Big Soda Lake, Nevada an alkaline, meromictic lake. Geochim. Cosmochim. Ac. 1983, 47, 2107–2114. [Google Scholar] [CrossRef]
- Gerasimenko, L.M.; Mityushina, L.L.; Namsaraev, B.B. Microcoleus mats from alkaliphilic and halophilic communities. Microbiology 2002, 72, 71–79. [Google Scholar] [CrossRef]
- Tunnell, J.W.J.; Judd, F.W. The Laguna Madre of Texas and Tamaulipas; Texas A&M University Press: College Station, TX, USA, 2002; pp. 1–346. [Google Scholar]
- Gallardo, J.C.; Macias, V.; Velarde, E. Birds (Vertebrate: Aves) of the Gulf of Mexico. In Gulf of Mexico Origin, Water, and Biota. Biodiversity; Felder, D.L., Camp, D.K., Eds.; Texas A&M University: College Station, TX, USA, 2009; pp. 1321–1342. [Google Scholar]
- Amdurer, M.; Land, L.S. Geochemistry, hydrology, and mineralogy of the sand bulge area, Laguna Madre Flats, south Texas. J. Sediment. Petrol. 1982, 52, 703–716. [Google Scholar]
- Fisk, H.N. Padre Island and the Laguna Madre flats, coastal South Texas. Louisiana State University, 2nd Coastal Geography Conference 1959, 6–9, 103–151. [Google Scholar]
- Sorensen, L.O.; Conover, J.T. Algal mat communities of Lyngbya confervoides (Agardh) Gomont. Publ. Inst. Mar. Sci. Univ. Texas 1962, 8, 61–74. [Google Scholar]
- Pulich, W.; Rabalais, S. Primary production potential of blue-green algal mats on southern Texas tidal flats. Southwest. Nat. 1986, 31, 39–47. [Google Scholar] [CrossRef]
- Withers, K. Wind-tidal flats. In The Laguna Madre of Texas and Tamaulipas; Tunnell, J.W., Jr., Judd, F.W., Eds.; Texas A&M Press: College Station, TX, USA, 2002; pp. 114–126. [Google Scholar]
- Kirkwood, A.E.; Buchheim, J.A.; Buchheim, M.A.; Henley, W.J. Cyanobacterial diversity and halotolerance in a variable hypersaline environment. Microb. Ecol. 2008, 55, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Bernt Sørensen, K.; Canfield, D.E.; Oren, A. Salinity responses of benthic microbial communities in a solar saltern (Eilat, Israel). Appl. Environ. Microbiol. 2004, 70, 1608–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Yang, H.; Zhao, M.; Zhanga, X. Spatial distribution patterns of benthic microbial communities along the Pearl Estuary, China. Syst. Appl. Microbiol. 2014, 37, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Urakawa, H.; Yoshida, T.; Nishimura, M.; Ohwada, K. Characterization of depth-related population variation in microbial communities of a coastal marine sediment using 16S rDNA-based approaches and quinone profiling. Environ. Microbiol. 2000, 2, 542–554. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, B.B.; Revsbech, N.P.; Cohen, Y. Photosynthesis and structure of benthic microbial mats: microelectrode and SEM studies of four cyanobacterial communities. Limnol. Oceanogr. 1983, 28, 1075–1093. [Google Scholar] [CrossRef]
- John, J.; Hay, M.; Paton, J. Cyanobacteria in benthic microbial communities in coastal salt lakes in Western Australia. Algol. Stud. 2009, 130, 125–135. [Google Scholar] [CrossRef]
- Vogt, J.C.; Albach, D.C.; Palinska, K.A. Cyanobacteria of the Wadden Sea: Seasonality and sediment influence on community composition. Hydrobiologia 2018, 811, 103–117. [Google Scholar] [CrossRef]
- Echnique-Subiabre, I.; Villeneuve, A.; Golubic, S.; Turquet, J.; Humbert, J.-F.; Gugger, M. Influence of local and global environmental parameters on the composition of cyanobacterial mats in a tropical lagoon. Microb. Ecol. 2015, 69, 234–244. [Google Scholar] [CrossRef] [Green Version]
- Ramos, V.M.C.; Castelo-Branco, R.; Leão, P.N.; Martins, J.; Carvalhal-Gomes, S.; Sobrinho da Silva, F.; Mendonça Filho, J.G.; Vasconcelos, V.M. Cyanobacterial diversity in microbial mats from the hypersaline lagoon system of Araruama, Brazil: An in-depth polyphasic study. Front. Microbiol. 2017, 8, 1233. [Google Scholar] [CrossRef] [Green Version]
- Bauer, K.; Diez, B.; Seppala, S.; Borg, A.J.; Bergman, B. Variability in diazotrophy and cyanobacterial diversity in a tropical intertidal lagoon. FEMS Microbiol. Ecol. 2008, 63, 205–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omoregie, E.; Crumbliss, L.L.; Bebout, B.M.; Zehr, J.P. Determination of nitrogen-fixing phylotypes in Lyngbya sp. and Microcoleus chthonoplastes cyanobacterial mats from Geurro Negro, Baja California, Mexico. Appl. Environ. Microb. 2004, 70, 2119–2128. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.K.; Caporaso, J.G.; Walker, J.J.; Spear, J.R.; Gold, N.J.; Robertson, C.E.; Hugenholtz, P.; Goodrich, J.; McDonald, D.; Knights, D.; et al. Phylogenetic stratigraphy in the Guerrero Negro hypersaline microbial mat. ISME J. 2013, 7, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Bolhuis, H.; Fillinger, L.; Stal, L.S. Coastal microbial mat diversity along a natural salinity gradient. PLoS ONE 2013, 8, e63166. [Google Scholar] [CrossRef] [Green Version]
- Jungblut, A.D.; Hawes, I.; Mountfort, D.; Hitzfeld, B.; Dietrich, D.R.; Burns, B.P.; Neilan, B.A. Diversity within cyanobacterial mat communities in variable salinity meltwater ponds of McMurdo Ice Shelf, Antarctica. Environ. Microbiol. 2005, 7, 519–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archer, S.D.J.; McDonald, I.R.; Herbold, C.W.; Lee, C.K.; Cary, C.S. Benthic microbial communities of coastal terrestrial and ice shelf Antarctic meltwater ponds. Front. Microbiol. 2015, 6, 485. [Google Scholar] [CrossRef] [Green Version]
- Zoppini, A.; Amalfitano, S.; Fazi, S.; Puddu, A. Dynamics of a benthic microbial community in a riverine environment subject to hydrological fluctuations (Mulargia River, Italy). Hydrobiologia 2010, 215, 37–51. [Google Scholar] [CrossRef]
- Fourçans, A.; García de Oteyza, T.; Wieland, A.; Solé, A.; Diestra, E.; van Bleijswijk, J.; Grimalt, J.O.; Kühl, M.; Esteve, I.; Muyzer, G.; et al. Characterization of functional bacterial groups in a hypersaline microbial mat community (Salins-de-Giraud, Camargue, France). FEMS Microbiol. Ecol. 2004, 51, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Yannarell, A.; Steppe, T.F.; Paerl, H.W. Genetic variance in the composition of two functional group (diazotrophs and cyanobacteria) from a hypersaline microbial mat. Appl. Environ. Microbiol. 2006, 72, 1207–1217. [Google Scholar] [CrossRef] [Green Version]
- Komárek, J. Subwasserflora von Mitteleuropa, Bd. 19/1: Cyanprokaryota. Teil/Part 1: Chrococcales; Gustav Fisher: Jena, Germany, 1999. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Subwasserflora von Mitteleuropa, Bd. 19/2: Cyanprokaryota. Tiel/ Part 2: Oscillatoriales; Springer: Heidelburg, Germany, 2007. [Google Scholar]
- Hindak, F. Colour Atlas of Cyanophytes; VEDA Publ.: Bratislavia, Slovak, 2008. [Google Scholar]
- Komárek, J. Subwasserflora von Mitteleuropa, Bd. 19/3: Cyanprokaryota. Teil/Part 3: Heterocystous genera; Springer: Heidelburg, Germany, 2013. [Google Scholar]
- Komárek, J.; Kaštovský, J.; Mares, J.; Joansen, J.R. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera), using a polyphasic approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Guillard, R.R.L.; Ryther, J.H. Studies of marine planktonic diatoms. I. Cyclotella nana Hustedt and Detonula confervacea Cleve. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Walters, W.; Hyde, E.R.; Berg-Lyons, D.; Ackermann, G.; Humphrey, A.; Gilbert, J.A.; Jannson, J.K.; Caporaso, J.G.; Fuhrman, J.A.; Apprill, A.; et al. Improved bacterial 16S rRNA gene (V4 and V4-5) and fungal internal transcribed spacer marker gene primers for microbial community surveys. mSystems 2016, 1, e9–e15. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Huttley, G.A.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, I.S.; Zimba, P.V. Cyanobacterial bioactive metabolites—A review of their chemistry and biology. Harmful Algae 2019, 86, 139–209. [Google Scholar]
- Zimba, P.V.; Huang, I.H.; Foley, J.E.; Linton, E.W. Identification of a new to science cyanobacterium, Toxifilum mysidocida gen. nov. and sp. nov. (Cyanobacteria, cyanophyceae). J. Phycol. 2017, 53, 188–197. [Google Scholar] [CrossRef]
- Siegesmund, M.A.; Johansen, J.R.; Karsten, U.; Friedl, T. Coleofasciculus gen. nov. (cyanobacteria): Morphological and molecular criteria for revision of the genus Microcoleus Gomont. J. Phycol. 2008, 44, 1572–1585. [Google Scholar] [CrossRef] [PubMed]
- Murray, N.J.; Phinn, S.R.; DeWitt, M.; Ferrari, R.; Johnston, R.; Lyons, M.R.; Clinton, C.; Thau, D.; Fuller, R.A. The global distribution and trajectory of tidal flats. Nature 2019, 565, 222–225. [Google Scholar] [CrossRef]
- Preisner, E.C.; Fichot, F.B.; Norman, R.S. Microbial mat composition and functional sensitivity to environmental disturbnace. Front. Microbiol. 2016, 7, 1632. [Google Scholar] [CrossRef] [Green Version]
- Blanco, Y.; Rivas, L.A.; Gonzalez-Toril, E.; Ruiz-Bermejo, M.; Morenzo-Paz, M.; Parro, V.; Palacin, A.; Aquilera, A.; Puente-Sanchez, F. Environmental parameters, and not phylogeny, determine the composition of extracellular polymeric substances in microbial mats from extreme environments. Sci. Total Environ. 2019, 650, 384–393. [Google Scholar] [CrossRef]
- Callejas, C.; Azziz, G.; Souza, E.M.; Gill, P.R.; Batista, S. Prokaroytic diversity in four microbial mats of the Fildes Peninsula, King George Island, maritime Antarctica. Polar Biol. 2018, 41, 935–943. [Google Scholar] [CrossRef]
- Engene, N.; Tronholm, A.; Paul, V.J. Uncovering cryptic diversity of Lyngbya: The new tropical marine cyanobacterial genus Dapis (Oscillatoriales). J. Phycol. 2018, 54, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Rudnick, P.; Meletzus, D.; Green, A.; He, L.; Kennedy, C. Regulation of nitrogen fixation by ammonium in diazotrophic species of proteobacteria. Soil Biol. Biochem. 1997, 5-6, 831–841. [Google Scholar] [CrossRef]
- Perlova, O.; Ureta, A.; Meletzus, D.; Nordlund, S. Sensing of n-status in Gluconacetobacter diazotrophicus: Biochemistry and genetics of nitrogen fixation and assimilation. Symbiosis 2003, 35, 73–84. [Google Scholar]
- Campbell, B.J.; Summers Engel, A.; Porter, M.L.; Takai, K. The versatile ε-proteobacteria: Key players in sulphidic habitats. Nat. Rev. Microbiol. 2006, 4, 458–468. [Google Scholar] [CrossRef]
- Gregersen, L.H.; Bryant, D.A.; Frigaard, N. Mechanisms and evolution of oxidative sulfur metabolism in green sulfur bacteria. Front. Microbiol. 2011, 2, 116. [Google Scholar] [CrossRef] [Green Version]
- Inoue, J.; Oshima, K.; Suda, W.; Sakamoto, M.; Iino, T.; Noda, S.; Hongoh, Y.; Hattori, M.; Ohkuma, M. Distribution and evolution of nitrogen fixation genes in the phylum Bacteroidetes. Microbes Environ. 2015, 30, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Sekiguchi, Y.; Yamada, T.; Hanada, S.; Ohashi, A.; Harada, H.; Kamagata, Y. Anaerolinea thermophila gen. nov., sp. nov. and Caldilinea aerophila gen. nov., sp. nov., novel filamentous thermophiles that represent a previously uncultured lineage of the domain Bacteria at the subphylum level. Int. J. Syst. Evol. Microbiol. 2003, 53, 1843–1851. [Google Scholar] [CrossRef]
- Yamada, T.; Imachi, H.; Ohashi, A.; Harada, H.; Hanada, S.; Kamagata, Y.; Sekiguchi, Y. Bellilinea caldifistulae gen. nov., sp. nov. and Longilinea arvoryzae gen. nov., sp. nov., strictly anaerobic, filamentous bacteria of the phylum Chloroflexi isolated from methanogenic propionate-degrading consortia. Int. J. Syst. Evol. Microbiol. 2007, 57, 2299–2306. [Google Scholar] [CrossRef] [Green Version]
- Sorokin, D.Y.; Toshchakov, S.V.; Kolganova, T.V.; Kublanov, I.V. Halo (natrono) archaea isolated from hypersaline lakes utilize cellulose and chitin as growth substrates. Front. Microbiol. 2015, 6, 942. [Google Scholar] [CrossRef] [Green Version]
- Quadri, I.; Hassani, I.I.; l’Haridon, S.; Chalopin, M.; Hacène, H.; Jebbar, M. Characterization and antimicrobial potential of extremely halophilic archaea isolated from hypersaline environments of the Algerian Sahara. Microbiol. Res. 2016, 186–187, 119–131. [Google Scholar] [CrossRef]
- Rao, S.; Chan, O.W.; Lacap-Bugler, D.C.; Pointing, S.B. Radiation-tolerant bacteria isolated from high altitude soil in Tibet. Indian J. Microbiol. 2016, 56, 508–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaoz, U.; Couradeau, E.; da Rocha, U.M.; Lim, H.; Northen, T.; Garcia-Pichel, F.; Brodie, E.L. Large blooms of Bacillales (Firmicutes) underlie the response to wetting of cyanobacterial biocrusts at various stages of maturity. mBio 2018, 9, e01366-e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jetten, M.S.M.; Sliekers, O.; Kuypers, M.; Dalsgaard, T.; van Niftrik, L.; Cirpus, I.; van de Pas-Schoonen, K.; Lavik, G.; Thamdrup, B.; Le Paslier, D.; et al. Anaerobic ammonium oxidation by marine and freshwater planctomycete-like bacteria. Appl. Microbiol. Biotechnol. 2003, 63, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Kaštovský, J.; Berrendero Gómez, E.; Hladil, J.; Johansen, J.R. Cyanocohniella calida gen. et sp. nov. (Cyanobacteria: Aphanizomenonaceae) a new cyanobacterium from the thermal springs from Karlovy Vary, Czech Republic. Phytotaxa 2014, 18, 3. [Google Scholar] [CrossRef]
- Caroppo, C.; Turicchia, S.; Margheri, M.C. Phytoplankton assemblages in coastal waters of the northern Ionian Sea (eastern Mediterranean), with special reference to cyanobacteria. J. Mar. Biolog. Assoc. U.K. 2006, 86, 927–937. [Google Scholar] [CrossRef]
- Engene, N.; Rottacker, E.C.; Kastovsky, J.; Byrum, T.; Choi, H.; Ellisman, M.H.; Komárek, J.; Gerwick, W.H. Moorea producens gen. nov., sp. nov. and Moorea bouillonii comb. nov., tropical marine cyanobacteria rich in bioactive secondary metabolites. Int. J. Syst. Evol. Microbiol. 2012, 62, 1171–1178. [Google Scholar] [CrossRef]
- Zimba, P.V.; Shalygin, S.; Huang, I.S.; Momčilović, M.; Abdulla, H. A new boring toxin producer-Perforafilum tunnelli gen. & sp. nov. (Oscillatoriales, Cyanobacteria) isolated from Laguna Madre sand-tidal flats, Texas, USA. Phycologia 2020. submitted for publication. [Google Scholar]
- Quiblier, C.; Wood, S.; Echenique-Subiabre, I.; Heath, M.; Villeneuve, A.; Humbert, J.-F. A review of current knowledge on toxic benthic freshwater cyanobacteria-ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar]
- Tidgewell, J.; Clark, B.R.; Gerwick, W.H. 2.06-The Natural Products Chemistry of Cyanobacteria. In Comprehensive Natural Products; Liu, H.-W., Mander, L., Eds.; Elsevier: Oxford, UK, 2010. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dominant Taxa | |||||||
|---|---|---|---|---|---|---|---|
| Environments | Salinity | Temp (°C) | Location | Analyses | Bacteria | Cyanobacteria | Reference |
| Brine mats | 159–310 | n.d. | Great Salt Plains, USA | Cloning, LM | n.d. | Geitlerinema, Komvophoron | [17] |
| 215 | 23 | Eilat, Israel | LM, isolation | Desulfocella, Halochromatium, Methanohalobium | Halothece, Halospirulina, Oscillatoria | [18] | |
| Estuarine mats | 0–33.6 | 26.5–30.1 | Pearl Estuary, China | Sanger | Bacteroidetes, Proteobacteria | Found, but not specified | [19] |
| 29–31.2 | 25 | Tokyo Bay, Japan | Cloning | Proteobacteria | n.d. | [20] | |
| Hypersaline lake mats | 150 | 45 | Solar Lake, Egypt | LM, SEM | n.d. | Microcoleus (syn. Coleofasciculus) | [21] |
| 69–108 | 17.3–25.5 | Lake Clifton, Garden Lake, Government House Lake, Serpentine Lake, Herschell Lake, Australia | LM | n.d. | Aphanothece, Oscillatoria, Microcoleus, Spirulina, Schizothrix, Gloeocapsa | [22] | |
| Lagoonal mats | 27–36 | 0.1–22.5 | Hooksiel, Germany | NGS, SEM, LM | n.d. | Coleofasciculus, Hydrocoleum, Lyngbya | [23] |
| 34.2–35.1 | 28.3–30.6 | La Réunion lagoon, France | Cloning, LM | n.d. | Anabaena, Hydrocoleum, Leptolyngbya, Lyngbya, Symploca | [24] | |
| 35–60 | 24–34 | Araruama Lagoon, Brazil | Sanger, NGS, LM | n.d. | Geitlerinema, Halomicronema, Microcoleus, Oxynema | [25] | |
| 14–32 | 30–37 | Paje lagoon, Tanzania | Cloning, LM | Nitrogen-fixing bacteria detected | Lyngbya, Microcoleus, Oscillatoria, Spirulina | [26] | |
| 78–90 | n.d. | Guerrero Negro, Mexico | Cloning | Azotobacter, Desulfovibrio | Anabaena, Halothece, Myxosarcina, Phormidium, Plectonema, Synechocystis | [27] | |
| 90 | n.d. | Guerrero Negro, Mexico | Sanger | Bacteroidetes, Chloroflexi, Planctomycetes, Proteobacteria | Lyngbya | [28] | |
| 225–280 (pore water) | n.d. | Schiermonnikoog Island, Netherlands | Sanger | n.d. | Coleofasciculus, Lyngbya | [29] | |
| 0–150 | 10–32 | Laguna Madre, USA | NGS, LM, SEM | Bacteroidetes, Chloroflexi, Desulfobacteria, Firmicutes, Halobacterota, Proteobacteria | Coleofasciculus, Lyngbya | This Study | |
| Meltwater ponds | 0.158–52.9 (mS/cm; cond.) | n.d. | McMurdo Ice Shelf, Bratina Island | Cloning, LM | n.d. | Oscillatoria | [30] |
| 0.6–40.8 (mS/cm; cond.) | n.d. | Bratina Island and the Miers Valley | Sanger | Actinobacteria, Bacteroidetes, Chloroflexi, Proteobacteria | Anabaena, Microcoleus, Phormidesmis | [31] | |
| Rivermouth mats | 0 | 9–29 | Mulargia River, Italy | FISH | Proteobacteria | n.d. | [32] |
| Salt ponds | 75–103 | 20–23 | Salins-de-Giraud, France | CLSM, DGGE | n.d. | Microcoleus (syn. Coleofasciculus), Halomicronema | [33] |
| 87–114 | 28–40 | San Salvador Island, Bahamas | Sanger | n.d. | Leptolyngbya, Phormidium | [34] | |
| Order | Family | Genus/Species | Wet | Med | Dry | Genomic |
|---|---|---|---|---|---|---|
| Chroococcales | Aphanothecaceae | Aphanothece utahensis Tilden | R | |||
| Chroococcaceae | Chroococcus cf. pulcherrimus Welsh | |||||
| Chroococcus obliteratus P. G. Richter | R | |||||
| Chroococcus sp. | R | R | R | R | ||
| Cyanosarcina sp. | ||||||
| Dactylococcopsis sp. | R | |||||
| Gloeocapsa sp. | R | R | ||||
| Gloeocapsopsis sp. | R | R | R | |||
| Gloeothece sp. | R | |||||
| Cyanothrichaceae | Johannesbaptistia pellucida (Dickie) W. R. Taylor et Drouet | R | R | R | ||
| Johannesbaptistia sp. | R | R | R | |||
| Chroococcidiopsidales | Chroococcidiopsidaceae | Chroococcidiopsis sp. | ||||
| Nostocales | Aphanizomenonaceae | Cyanocohniella sp. | R | R | ||
| Oscillatoriales | Coleofasciculaceae | Anagnostidinema sp. | R | R | ||
| Coleofasciculus chthonoplastes (Thuret ex Gomont) M. Siegesmund et al. | R | R | R | R | ||
| Coleofasciculus sp. | R | R | R | A | ||
| Roseofilum sp. | R | |||||
| Gomontiellaceae | Komvophoron sp. | R | ||||
| Microcoleaceae | Arthrospira sp. | C | ||||
| Hydrocoleum lyngbyaceum | R | |||||
| Kamptonema sp. | R | R | ||||
| Porphyrosiphon sp. | R | R | ||||
| Symploca sp. | R | |||||
| Symplocastrum sp. | R | R | R | |||
| Oscillatoriaceae | Lyngbya sp. | C | A | A | A | |
| New genus - akin to Lyngbya aestuarii Liebman ex Gomont | R | C | A | |||
| Oscillatoria nigro-viridis Thwaites ex Gomont | R | |||||
| Oscillatoria sp. | C | R | ||||
| Phormidium sp. | R | C | ||||
| Pleurocapsales | Hyellaceae | New genus - akin to Pleurocapsa minuta Geitler | R | |||
| Xenococcaceae | Xenococcus sp. | R | ||||
| Spirulinales | Spirulinaceae | Halospirulina sp. | R | |||
| Spirulina sp. | C | R | ||||
| Spirulina tenerrima Kützing ex Gomont | C | C | C | |||
| Synechococcales | Leptolyngbyaceae | Halomicronema sp. | R | R | ||
| Leibleinia nordgaardii (Wille) Anagnostidis et Komárek | R | |||||
| Leibleinia sp. | R | R | R | |||
| Leibleinia subtilis (Holden) Anagnostidis et Komárek | C | |||||
| Leptolyngbya sp. | C | |||||
| Nodosilinea sp. | C | |||||
| Stenomitos sp. | R | |||||
| Merismopediaceae | Aphanocapsa sp. | C | R | |||
| Eucapsis prescottii (Drouet et Daily) Komárek et Hindák | R | R | ||||
| Synechocystis sp. | R | C | ||||
| Pseudanabaenaceae | Geitlerinema sp. | C | C | |||
| Jaaginema sp. | R | |||||
| Pseudanabaena sp. | C | C | C | R | ||
| Schizotrichaceae | Schizothrix helva Frémy | R | R | |||
| Schizothrix sp. | R | R | R | |||
| Synechococcaceae | Bacularia sp. | C | ||||
| Synechococcus sp. | C | R | ||||
| Trichocoleusaceae | Trichocoleus sp. | R | ||||
| Trichocoleus tenerrimus (Gomont) Anagnostidis | R | |||||
| Oscillatoriales | Laspinemaceae fam. nov. | Perforafilum tunnelli (new genus - akin to Oxynema sp.) | R |
| Literature Origins | ||||||||
|---|---|---|---|---|---|---|---|---|
| Main Group | Subgroup | Compound | Bioactivity | Occr | Cy | Family | Genus | In This Study |
| Aeruginosin | AER 103-A | Protease inhibitors | 1 | Y | Microcystaceae | Microcystis | Order found | |
| AER 98-A | 2 | |||||||
| AER 98-B | 1 | |||||||
| AER 98-C | 2 | |||||||
| AER DA 642-A | 1 | |||||||
| AER DA 642-B | 1 | |||||||
| AER DA-688 | 3 | |||||||
| AER GE-686 | 1 | |||||||
| AER KB-676 | 1 | |||||||
| Spumigin | AER-SPM B-1 | 1 | Y | Nostocaceae | Nodularia | Morph found | ||
| AER-SPM B-2* | 2 | |||||||
| AER-SPM E* | 1 | |||||||
| AER-SPM G | 1 | |||||||
| AER-SPM H | 1 | |||||||
| Anabaenopeptin | ABPN B | Protease/protein phosphate inhibitors | 5 | Y | Pseudanabaenaceae | Toxifilum | Family found | |
| Feritonic acid | ABPN-FTA B | 1 | Y | Microcystaceae | Microcystis | Order found | ||
| Konbamide | ABPN-KBA | 1 | N | Theonellidae | Theonella | N/A | ||
| Paltolide | ABPN-PAL A | 2 | N | Theonellidae | Theonella | N/A | ||
| Anatoxin | ANA-a | Neurotoxicity | 4 | Y | Microcoleaceae | Arthrospira | Genus found | |
| 11-carboxy-ANA | 1 | |||||||
| Antillatoxin | ATX B | Cytotoxicity | 1 | Y | Oscillatoriaceae | Moorea | Family found | |
| Apratoxin | APX F | Cytotoxicity | 1 | Y | Oscillatoriaceae | Moorea | Family found | |
| APX G | 1 | |||||||
| Coibacin | CBC C | Cytotoxicity, anti-parasite, neurotoxicity | 4 | Y | Oscillatoriaceae | Oscillatoria | Genus found | |
| CBC D | 1 | |||||||
| Cyanopeptolin | Aeruginopeptin | CPT-APP 228-A | Protease/protein phosphate inhibitors, cytotoxicity, anti-crustacean | 1 | Y | Microcystaceae | Microcystis | Order found |
| CPT-APP 917-S-C | 1 | |||||||
| Micropeptin | CPT-MPP MM-836 | 1 | Y | Microcystaceae | Microcystis | Order found | ||
| Oscillapeptin | CPT-OPP A | 2 | Y | Microcoleaceae | Planktothrix | Family found | ||
| Scyptolin | CPT-SPT A | 1 | Y | Scytonemataceae | Scytonema | Morph found | ||
| Tasipeptin | CPT-TSP B | 1 | Y | Microcoleaceae | Symploca | Genus found | ||
| Largamide | Dolastatin | LRA-DST D | Neurotoxicity, cytotoxicity | 2 | N | Aplysiidae | Dolabella | N/A |
| Linear-Dolastatin | l-DST 15 | Protease inhibitors, cytotoxicity, anti-bacteria, anti-fungi, anti-parasite | 1 | Y | Microcoleaceae | Symploca | Genus found | |
| l-DST 18 | 1 | |||||||
| Lyngbyabellin | LBB F | Cytotoxicity, anti-fungi | 1 | Y | Oscillatoriaceae | Moorea | Family found | |
| LBB G | 1 | |||||||
| Lyngbyaloside | Phorbaside | LBL-PBA A* | Cytotoxicity | 2 | Y | Oscillatoriaceae | Moorea | Family found |
| Malyngamide | MGA 4 | Cytotoxicity, anti-fungi, anti-parasite | 2 | Y | Oscillatoriaceae | Moorea | Family found | |
| Microcolin | Majusculamide | MCL-MSA D | Cytotoxicity | 1 | Y | Oscillatoriaceae | Moorea | Family found |
| Microcyclamide | MCA 7806-B | Cytotoxicity, anti-parasite | 7 | Y | Microcystaceae | Microcystis | Order found | |
| Aerucyclamide | MCA-ACA C | 2 | Y | Microcystaceae | Microcystis | Order found | ||
| Balgacyclamide | MCA-BCA B* | 1 | Y | Microcystaceae | Microcystis | Order found | ||
| MCA-BCA C | 5 | |||||||
| Bistratamide | MCA-BTA I | 5 | N | Didemnidae | Lissoclinum | N/A | ||
| Venturamide | MCA-tetraOH-VTA A | 1 | Y | Oscillatoriaceae | Oscillatoria | Genus found | ||
| Microcystin | [(6Z)-Adda5]MCYST-RR | Hepatotoxicity | 1 | Y | Microcystaceae | Microcystis | Order found | |
| [ADMAdda5]MCYST-HilR | 4 | |||||||
| [ADMAdda5]MCYST-LHar | 4 | |||||||
| [ADMAdda5]MCYST-LR | 4 | |||||||
| [D-Asp3, ADMAdda5]MCYST-LHar | 3 | |||||||
| [D-Asp3, Dha7]MCYST-E(OMe)E(OMe) | 7 | |||||||
| MCYST-OiaA | 1 | |||||||
| MCYST-OiaAba | 3 | |||||||
| MCYST-OiaR | 1 | |||||||
| MCYST-RF | 1 | |||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, I.-S.; Pinnell, L.J.; Turner, J.W.; Abdulla, H.; Boyd, L.; Linton, E.W.; Zimba, P.V. Preliminary Assessment of Microbial Community Structure of Wind-Tidal Flats in the Laguna Madre, Texas, USA. Biology 2020, 9, 183. https://doi.org/10.3390/biology9080183
Huang I-S, Pinnell LJ, Turner JW, Abdulla H, Boyd L, Linton EW, Zimba PV. Preliminary Assessment of Microbial Community Structure of Wind-Tidal Flats in the Laguna Madre, Texas, USA. Biology. 2020; 9(8):183. https://doi.org/10.3390/biology9080183
Chicago/Turabian StyleHuang, I.-Shuo, Lee J. Pinnell, Jeffrey W. Turner, Hussain Abdulla, Lauren Boyd, Eric W. Linton, and Paul V. Zimba. 2020. "Preliminary Assessment of Microbial Community Structure of Wind-Tidal Flats in the Laguna Madre, Texas, USA" Biology 9, no. 8: 183. https://doi.org/10.3390/biology9080183