Phytoremediation of Cadmium: Physiological, Biochemical, and Molecular Mechanisms

 ,
,  ,
,  ,
, 

Abstract
:1. Introduction
2. Plant Responses to Cadmium Toxicity
3. Phytoremediation Processes and Their Salient Features
3.1. Phytoextraction
3.2. Phytostabilization
3.3. Phytofiltration
3.4. Phtytostimulation
4. Effect of Phytoremediation on Cd Removal from Soils
5. Role of Transporters in Cadmium Accumulation on Hyperaccumulator Plants
5.1. Long-Distance Cd Transport
5.2. Vacuolar Storage and Sequestration
5.3. Mechanism of Cd Crossing the Plasma Membrane of Root
6. Antioxidant Defense: A Key Mechanism of Cadmium Tolerance and Phytoremediation
7. Chelate-Assisted Cadmium Phytoremediation
8. Phytochelatins and Metallothionein for Cadmium Phytoremediation
9. Omics Approaches for Cadmium Phytoremediation
9.1. Genomics
9.2. Transcriptomics
9.3. Proteomics
9.4. Metabolomics
10. Genetic Engineering for Cadmium Phytoremediation
11. Employing Microbes for Cadmium Phytoremediation
11.1. Role of Arbuscular Mycorrhizal Fungi
11.2. Role of Plant Growth-Promoting Bacteria (PGRB) in the Phytoremediation of Cd
12. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dhaliwal, S.S.; Singh, J.; Taneja, P.K.; Mandal, A. Remediation techniques for removal of heavy metals from the soil contaminated through different sources: A review. Environ. Sci. Pollut. Res. 2020, 27, 1319–1333. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Dong, M.; Mao, P.; Zhuang, P.; Paz-Ferreiro, J.; Li, Y.; Li, Y.; Hu, X.; Netherway, P.; Li, Z. Evaluation of phytoremediation potential of five Cd (hyper) accumulators in two Cd contaminated soils. Sci. Total Environ. 2020, 721, 137581. [Google Scholar] [CrossRef] [PubMed]
- Baliardini, C.; Meyer, C.-L.; Salis, P.; Saumitou-Laprade, P.; Verbruggen, N. Cation EXCHANGER1 cosegregates with cadmium tolerance in the metal hyperaccumulator Arabidopsis halleri and plays a role in limiting oxidative stress in Arabidopsis spp. Plant Physiol. 2015, 169, 549–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges, K.L.R.; Salvato, F.; Alcântara, B.K.; Nalin, R.S.; Piotto, F.Â.; Azevedo, R.A. Temporal dynamic responses of roots in contrasting tomato genotypes to cadmium tolerance. Ecotoxicology 2018, 27, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Fattahi, B.; Arzani, K.; Souri, M.K.; Barzegar, M. Effects of cadmium and lead on seed germination, morphological traits, and essential oil composition of sweet basil (Ocimum basilicum L.). Ind. Crops Prod. 2019, 138, 111584. [Google Scholar] [CrossRef]
- Shah, V.; Daverey, A. Phytoremediation: A multidisciplinary approach to clean up heavy metal contaminated soil. Environ. Technol. Innov. 2020, 18, 100774. [Google Scholar] [CrossRef]
- Shanying, H.; Xiaoe, Y.; Zhenli, H.; Baligar, V.C. Morphological and physiological responses of plants to cadmium toxicity: A review. Pedosphere 2017, 27, 421–438. [Google Scholar]
- Navarro-León, E.; Oviedo-Silva, J.; Ruiz, J.M.; Blasco, B. Possible role of HMA4a TILLING mutants of Brassica rapa in cadmium phytoremediation programs. Ecotoxicol. Environ. Saf. 2019, 180, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.A.; Castagna, A.; Ranieri, A.; di Toppi, L.S. Cadmium tolerance in Brassica juncea roots and shoots is affected by antioxidant status and phytochelatin biosynthesis. Plant Physiol. Biochem. 2012, 57, 15–22. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Pan, L.-B.; Ma, J.; Wang, X.-L.; Hou, H. Heavy metals in soils from a typical county in Shanxi Province, China: Levels, sources and spatial distribution. Chemosphere 2016, 148, 248–254. [Google Scholar] [CrossRef]
- Khan, S.; Rehman, S.; Khan, A.Z.; Khan, M.A.; Shah, M.T. Soil and vegetables enrichment with heavy metals from geological sources in Gilgit, northern Pakistan. Ecotoxicol. Environ. Saf. 2010, 73, 1820–1827. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xiao, T.; Ning, Z.; Li, H.; Tang, J.; Zhou, G. High cadmium concentration in soil in the Three Gorges region: Geogenic source and potential bioavailability. Appl. Geochem. 2013, 37, 149–156. [Google Scholar] [CrossRef]
- Khan, S.; Munir, S.; Sajjad, M.; Li, G. Urban park soil contamination by potentially harmful elements and human health risk in Peshawar City, Khyber Pakhtunkhwa, Pakistan. J. Geochem. Explor. 2016, 165, 102–110. [Google Scholar] [CrossRef]
- Nawab, J.; Khan, S.; Aamir, M.; Shamshad, I.; Qamar, Z.; Din, I.; Huang, Q. Organic amendments impact the availability of heavy metal (loid) s in mine-impacted soil and their phytoremediation by Penisitum americanum and Sorghum bicolor. Environ. Sci. Pollut. Res. 2016, 23, 2381–2390. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Sharma, A. Cadmium toxicity: Effects on human reproduction and fertility. Rev. Environ. Health 2019, 34, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Genchi, G.; Sinicropi, M.S.; Lauria, G.; Carocci, A.; Catalano, A. The effects of cadmium toxicity. Int. J. Environ. Res. Public Health 2020, 17, 3782. [Google Scholar] [CrossRef] [PubMed]
- Huff, J.; Lunn, R.M.; Waalkes, M.P.; Tomatis, L.; Infante, P.F. Cadmium-induced cancers in animals and in humans. Int. J. Occup. Environ. Health 2007, 13, 202–212. [Google Scholar] [CrossRef] [Green Version]
- Djordjevic, V.R.; Wallace, D.R.; Schweitzer, A.; Boricic, N.; Knezevic, D.; Matic, S.; Grubor, N.; Kerkez, M.; Radenkovic, D.; Bulat, Z.; et al. Environmental cadmium exposure and pancreatic cancer: Evidence from case control, animal and in vitro studies. Environ. Int. 2019, 128, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Li, J.; Xie, H.; Yu, C. Review on remediation technologies of soil contaminated by heavy metals. Procedia Environ. Sci. 2012, 16, 722–729. [Google Scholar] [CrossRef] [Green Version]
- Paz-Ferreiro, J.; Gascó, G.; Méndez, A.; Reichman, S.M. Soil Pollution and Remediation. Int. J. Environ. Res. Public Health 2018, 15, 1657. [Google Scholar] [CrossRef] [Green Version]
- Maity, J.P.; Huang, Y.M.; Fan, C.-W.; Chen, C.-C.; Li, C.-Y.; Hsu, C.-M.; Chang, Y.-F.; Wu, C.-I.; Chen, C.-Y.; Jean, J.-S. Evaluation of remediation process with soapberry derived saponin for removal of heavy metals from contaminated soils in Hai-Pu, Taiwan. J. Environ. Sci. 2013, 25, 1180–1185. [Google Scholar] [CrossRef]
- Juwarkar, A.A.; Nair, A.; Dubey, K.V.; Singh, S.; Devotta, S. Biosurfactant technology for remediation of cadmium and lead contaminated soils. Chemosphere 2007, 68, 1996–2002. [Google Scholar] [CrossRef]
- Guo, J.; Zhou, Y. Transformation of heavy metals and dewaterability of waste activated sludge during the conditioning by Fe2+-activated peroxymonosulfate oxidation combined with rice straw biochar as skeleton builder. Chemosphere 2020, 238, 124628. [Google Scholar] [CrossRef]
- Ren, W.-X.; Li, P.-J.; Geng, Y.; Li, X.-J. Biological leaching of heavy metals from a contaminated soil by Aspergillus niger. J. Hazard. Mater. 2009, 167, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Kapahi, M.; Sachdeva, S. Mycoremediation potential of Pleurotus species for heavy metals: A review. Bioresour. Bioprocessing 2017, 4, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fazli, M.M.; Soleimani, N.; Mehrasbi, M.; Darabian, S.; Mohammadi, J.; Ramazani, A. Highly cadmium tolerant fungi: Their tolerance and removal potential. J. Environ. Health Sci. Eng. 2015, 13, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagot, D.; Lebeau, T.; Jezequel, K. Microorganisms for remediation of cadmium-contaminated soils. Environ. Chem. Lett. 2006, 4, 207–211. [Google Scholar] [CrossRef]
- Tang, J.; He, J.; Liu, T.; Xin, X.; Hu, H. Removal of heavy metal from sludge by the combined application of a biodegradable biosurfactant and complexing agent in enhanced electrokinetic treatment. Chemosphere 2017, 189, 599–608. [Google Scholar] [CrossRef]
- Virkutyte, J.; Sillanpää, M.; Latostenmaa, P. Electrokinetic soil remediation—Critical overview. Sci. Total Environ. 2002, 289, 97–121. [Google Scholar] [CrossRef]
- Shen, Z.; Chen, X.; Jia, J.; Qu, L.; Wang, W. Comparison of electrokinetic soil remediation methods using one fixed anode and approaching anodes. Environ. Pollut. 2007, 150, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, Z.; He, X.; Liu, Y. Optimization analysis and mechanism exploration on the removal of cadmium from contaminated soil by electrokinetic remediation. Sep. Purif. Technol. 2020. [Google Scholar] [CrossRef]
- Azizollahi, Z.; Ghaderian, S.M.; Ghotbi-Ravandi, A.A. Cadmium accumulation and its effects on physiological and biochemical characters of summer savory (Satureja hortensis L.). Int. J. Phytoremediat. 2019, 21, 1241–1253. [Google Scholar] [CrossRef]
- Yu, S.; Sheng, L.; Zhang, C.; Deng, H. Physiological response of Arundo donax to cadmium stress by Fourier transform infrared spectroscopy. Spectrochim. Acta A 2018, 198, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.S.; Yang, Y.; Gu, T.; Wu, Z.; Zhang, Z. The Arabidopsis defensin gene AtPDF2.5 mediates cadmium tolerance and accumulation. Plant Cell Environ. 2019, 42, 2681–2695. [Google Scholar] [CrossRef]
- Gajewska, E.; SkŁodowska, M. Differential effect of equal copper, cadmium and nickel concentration on biochemical reactions in wheat seedlings. Ecotoxicol. Environ. Saf. 2010, 73, 996–1003. [Google Scholar] [CrossRef]
- Popova, L.P.; Maslenkova, L.T.; Yordanova, R.Y.; Ivanova, A.P.; Krantev, A.P.; Szalai, G.; Janda, T. Exogenous treatment with salicylic acid attenuates cadmium toxicity in pea seedlings. Plant Physiol. Biochem. 2009, 47, 224–231. [Google Scholar] [CrossRef]
- Yamada, M.; Malambane, G.; Yamada, S.; Suharsono, S.; Tsujimoto, H.; Moseki, B.; Akashi, K. Differential physiological responses and tolerance to potentially toxic elements in biodiesel tree Jatropha curcas. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Meena, M.; Aamir, M.; Kumar, V.; Swapnil, P.; Upadhyay, R. Evaluation of morpho-physiological growth parameters of tomato in response to Cd induced toxicity and characterization of metal sensitive NRAMP3 transporter protein. Environ. Exp. Bot. 2018, 148, 144–167. [Google Scholar] [CrossRef]
- Huang, L.; Li, W.C.; Tam, N.F.Y.; Ye, Z. Effects of root morphology and anatomy on cadmium uptake and translocation in rice (Oryza sativa L.). J. Environ. Sci. 2019, 75, 296–306. [Google Scholar] [CrossRef]
- Chun, C.-P.; Zhou, W.; Ling, L.-L.; Cao, L.; Fu, X.-Z.; Peng, L.-Z.; Li, Z.-G. Uptake of cadmium (Cd) by selected citrus rootstock cultivars. Sci. Hortic. 2020, 263, 109061. [Google Scholar] [CrossRef]
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Iannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Environ. Exp. Bot. 2012, 83, 33–46. [Google Scholar] [CrossRef]
- Zheng, R.-L.; Li, H.-F.; Jiang, R.-F.; Zhang, F.-S. Cadmium accumulation in the edible parts of different cultivars of radish, Raphanus sativus L.; and carrot, Daucus carota var. sativa, grown in a Cd-contaminated soil. Bull. Environ. Cont. Toxicol. 2008, 81, 75–79. [Google Scholar] [CrossRef]
- Ulusu, Y.; Öztürk, L.; Elmastaş, M. Antioxidant capacity and cadmium accumulation in parsley seedlings exposed to cadmium stress. Russ. J. Plant Physiol. 2017, 64, 883–888. [Google Scholar] [CrossRef]
- Li, L.-Z.; Tu, C.; Peijnenburg, W.J.; Luo, Y.-M. Characteristics of cadmium uptake and membrane transport in roots of intact wheat (Triticum aestivum L.) seedlings. Environ. Pollut. 2017, 221, 351–358. [Google Scholar] [CrossRef]
- Rahman, M.F.; Ghosal, A.; Alam, M.F.; Kabir, A.H. Remediation of cadmium toxicity in field peas (Pisum sativum L.) through exogenous silicon. Ecotoxicol. Environ. Saf. 2017, 135, 165–172. [Google Scholar] [CrossRef]
- Saleem, M.H.; Fahad, S.; Khan, S.U.; Ahmar, S.; Khan, M.H.U.; Rehman, M.; Maqbool, Z.; Liu, L. Morpho-physiological traits, gaseous exchange attributes, and phytoremediation potential of jute (Corchorus capsularis L.) grown in different concentrations of copper-contaminated soil. Ecotoxicol. Environ. Saf. 2020, 189, 109915. [Google Scholar] [CrossRef]
- Zhang, X.; Li, M.; Yang, H.; Li, X.; Cui, Z. Physiological responses of Suaeda glauca and Arabidopsis thaliana in phytoremediation of heavy metals. J. Environ. Manag. 2018, 223, 132–139. [Google Scholar] [CrossRef]
- Kaya, C.; Okant, M.; Ugurlar, F.; Alyemeni, M.N.; Ashraf, M.; Ahmad, P. Melatonin-mediated nitric oxide improves tolerance to cadmium toxicity by reducing oxidative stress in wheat plants. Chemosphere 2019, 225, 627–638. [Google Scholar] [CrossRef]
- Lian, J.; Zhao, L.; Wu, J.; Xiong, H.; Bao, Y.; Zeb, A.; Tang, J.; Liu, W. Foliar spray of TiO2 nanoparticles prevails over root application in reducing Cd accumulation and mitigating Cd-induced phytotoxicity in maize (Zea mays L.). Chemosphere 2020, 239, 124794. [Google Scholar] [CrossRef]
- Carvalho, M.E.; Piotto, F.A.; Gaziola, S.A.; Jacomino, A.P.; Jozefczak, M.; Cuypers, A.; Azevedo, R.A. New insights about cadmium impacts on tomato: Plant acclimation, nutritional changes, fruit quality and yield. Food Energy Secur. 2018, 7, e00131. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Luo, Q.; Zhao, Y.; Long, Y.; Liu, S.; Pan, Y. Physiological and biochemical mechanisms preventing Cd toxicity in the new hyperaccumulator Abelmoschus manihot. J. Plant Growth Regul. 2018, 37, 709–718. [Google Scholar] [CrossRef]
- Ahmed, H.; Häder, D.-P. Rapid ecotoxicological bioassay of nickel and cadmium using motility and photosynthetic parameters of Euglena gracilis. Environ. Exp. Bot. 2010, 69, 68–75. [Google Scholar] [CrossRef]
- Liu, M.; Bi, J.; Liu, X.; Kang, J.; Korpelainen, H.; Niinemets, Ü.; Li, C. Microstructural and physiological responses to cadmium stress under different nitrogen levels in Populus cathayana females and males. Tree Physiol. 2020, 40, 30–45. [Google Scholar] [CrossRef] [PubMed]
- Alyemeni, M.N.; Ahanger, M.A.; Wijaya, L.; Alam, P.; Bhardwaj, R.; Ahmad, P. Selenium mitigates cadmium-induced oxidative stress in tomato (Solanum lycopersicum L.) plants by modulating chlorophyll fluorescence, osmolyte accumulation, and antioxidant system. Protoplasma 2018, 255, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Wang, X.; Shang, L.; Zhou, Z.; Wang, R. Cadmium accumulation and its effects on nutrient uptake and photosynthetic performance in cucumber (Cucumis sativus L.). Philipp. Agric. Sci. 2017, 100, 263–270. [Google Scholar]
- Rossi, L.; Bagheri, M.; Zhang, W.; Chen, Z.; Burken, J.G.; Ma, X. Using artificial neural network to investigate physiological changes and cerium oxide nanoparticles and cadmium uptake by Brassica napus plants. Environ. Pollut. 2019, 246, 381–389. [Google Scholar] [CrossRef]
- Iannone, M.F.; Rosales, E.P.; Groppa, M.D.; Benavides, M.P. Reactive oxygen species formation and cell death in catalase-deficient tobacco leaf disks exposed to cadmium. Protoplasma 2010, 245, 15–27. [Google Scholar] [CrossRef]
- Bertin, G.; Averbeck, D. Cadmium: Cellular effects, modifications of biomolecules, modulation of DNA repair and genotoxic consequences (a review). Biochimie 2006, 88, 1549–1559. [Google Scholar] [CrossRef]
- Gjorgieva, D.; Kadifkova-Panovska, T.; Mitrev, S.; Kovacevik, B.; Kostadinovska, E.; Bačeva, K.; Stafilov, T. Assessment of the genotoxicity of heavy metals in Phaseolus vulgaris L. as a model plant system by Random Amplified Polymorphic DNA (RAPD) analysis. J. Environ. Sci. Health 2012, 47, 366–373. [Google Scholar] [CrossRef]
- Surgun-Acar, Y. Determination of heavy metal-induced DNA damage in Pisum Sativum, L. at the molecular and population level. J. Anim. Plant Sci. 2018, 28, 1825–1834. [Google Scholar]
- Ahmad, P.; Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alam, P. Exogenous application of nitric oxide modulates osmolyte metabolism, antioxidants, enzymes of ascorbate-glutathione cycle and promotes growth under cadmium stress in tomato. Protoplasma 2018, 255, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Chen, W.; Liu, F.; Wan, Y. Physiological responses of peanut seedlings to exposure to low or high cadmium concentration and the alleviating effect of exogenous nitric oxide to high cadmium concentration stress. Plant Biosyst. 2020, 154, 405–412. [Google Scholar] [CrossRef]
- Çatav, Ş.S.; Genç, T.O.; Oktay, M.K.; Küçükakyüz, K. Cadmium toxicity in wheat: Impacts on element contents, antioxidant enzyme activities, oxidative stress, and genotoxicity. Bull. Environ. Cont. Toxicol. 2020, 104, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Barman, F.; Majumdar, S.; Arzoo, S.H.; Kundu, R. Genotypic variation among 20 rice cultivars/landraces in response to cadmium stress grown locally in West Bengal, India. Plant Physiol. Biochem. 2020, 148, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Irfan, M.; Ahmad, A.; Hayat, S. Effect of cadmium on the growth and antioxidant enzymes in two varieties of Brassica juncea. Saudi J. Biol. Sci. 2014, 21, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Abdula, S.E.; Jang, D.W.; Park, S.-H.; Yoon, U.-H.; Jung, Y.J.; Kang, K.K.; Nou, I.S.; Cho, Y.-G. Overexpression of the glutamine synthetase gene modulates oxidative stress response in rice after exposure to cadmium stress. Plant Cell Rep. 2013, 32, 1521–1529. [Google Scholar] [CrossRef]
- Viehweger, K. How plants cope with heavy metals. Bot. Stud. 2014, 55, 35. [Google Scholar] [CrossRef] [Green Version]
- Ahmadpour, P.; Soleimani, M. Cadmium accumulation and translocation in Jatropha curcas grown in contaminated soils. JWSS-Isfahan Uni. Technol. 2015, 19, 179–190. [Google Scholar] [CrossRef]
- Suman, J.; Uhlik, O.; Viktorova, J.; Macek, T. Phytoextraction of heavy metals: A promising tool for clean-up of polluted environment? Front. Plant Sci. 2018, 9, 1476. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Liu, J.; Long, Y.; Chen, Z.; Sunahara, G.I.; Jiang, P.; You, S.; Lin, H.; Xiao, H. Phytoextraction of cadmium-contaminated soils: Comparison of plant species and low molecular weight organic acids. Int. J. Phytoremediat. 2020, 22, 383–391. [Google Scholar] [CrossRef]
- El-Mahrouk, E.-S.M.; Eisa, E.A.-H.; Hegazi, M.A.; Abdel-Gayed, M.E.-S.; Dewir, Y.H.; El-Mahrouk, M.E.; Naidoo, Y. Phytoremediation of cadmium-, copper-, and lead-contaminated soil by Salix mucronata (Synonym Salix safsaf). HortScience 2019, 54, 1249–1257. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.; Fernandes, A.; Junior, M.S.; Santos, C.; Lobato, A. Tolerance mechanisms in Cassia alata exposed to cadmium toxicity–potential use for phytoremediation. Photosynthetica 2018, 56, 495–504. [Google Scholar] [CrossRef]
- Ali, S.Y.; Banerjee, S.N.; Chaudhury, S. Phytoextraction of cadmium and lead by three vegetable-crop plants. Plant Sci. Today 2016, 3, 298–303. [Google Scholar] [CrossRef]
- Liu, L.; Li, Y.; Tang, J.; Hu, L.; Chen, X. Plant coexistence can enhance phytoextraction of cadmium by tobacco (Nicotiana tabacum L.) in contaminated soil. J. Environ. Sci. 2011, 23, 453–460. [Google Scholar] [CrossRef]
- Fan, K.-C.; Hsi, H.-C.; Chen, C.-W.; Lee, H.-L.; Hseu, Z.-Y. Cadmium accumulation and tolerance of mahogany (Swietenia macrophylla) seedlings for phytoextraction applications. J. Environ. Manag. 2011, 92, 2818–2822. [Google Scholar] [CrossRef] [PubMed]
- Farid, M.; Ali, S.; Shakoor, M.B.; Bharwana, S.A.; Rizvi, H.; Ehsan, S.; Tauqeer, H.M.; Iftikhar, U.; Hannan, F. EDTA assisted phytoremediation of cadmium, lead and zinc. Int. J. Agron. Plant Prod. 2013, 4, 2833–2846. [Google Scholar]
- Hassan, M.; Dagari, M.; Babayo, A. Effect of citric acid on cadmium ion uptake and stress response of hydroponically grown jute mallow (Corchorus olitorius). J. Environ. Anal. Toxicol. 2016, 6. [Google Scholar] [CrossRef]
- Anderson, C.; Brooks, R.; Stewart, R.; Simcock, R.; Robinson, B. The phytoremediation and phytomining of heavy metals. In Proceedings of the Pacrim International Congress on Earth Science, Exploration and Mining Around Pacific Rim, Bali, Indonesia, 10–13 October 1999; pp. 127–135. [Google Scholar]
- Ranieri, E.; Moustakas, K.; Barbafieri, M.; Ranieri, A.C.; Herrera-Melián, J.A.; Petrella, A.; Tommasi, F. Phytoextraction technologies for mercury-and chromium-contaminated soil: A review. J. Chem. Technol. Biotechnol. 2020, 95, 317–327. [Google Scholar] [CrossRef]
- Ernst, W.H. Phytoextraction of mine wastes—Options and impossibilities. Geochemistry 2005, 65, 29–42. [Google Scholar] [CrossRef]
- Robinson, B.; Anderson, C.; Dickinson, N. Phytoextraction: Where’s the action? J. Geochem. Explor. 2015, 151, 34–40. [Google Scholar] [CrossRef]
- Cheraghi, M.; Lorestani, B.; Khorasani, N.; Yousefi, N.; Karami, M. Findings on the phytoextraction and phytostabilization of soils contaminated with heavy metals. Biol. Trace Elem. Res. 2011, 144, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Dalvi, A.A.; Bhalerao, S.A. Response of plants towards heavy metal toxicity: An overview of avoidance, tolerance and uptake mechanism. Ann. Plant Sci. 2013, 2, 362–368. [Google Scholar]
- Andrade Júnior, W.V.; de Oliveira Neto, C.F.; Santos Filho, B.G.d.; do Amarante, C.B.; Cruz, E.D.; Okumura, R.S.; Barbosa, A.V.C.; de Sousa, D.J.P.; Teixeira, J.S.S.; Botelho, A.D.S. Effect of cadmium on young plants of Virola surinamensis. AoB Plants 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Zgorelec, Z.; Bilandzija, N.; Knez, K.; Galic, M.; Zuzul, S. cadmium and Mercury phytostabilization from soil using Miscanthus× giganteus. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Boros-Lajszner, E.; Wyszkowska, J.; Kucharski, J. Application of white mustard and oats in the phytostabilisation of soil contaminated with cadmium with the addition of cellulose and urea. J. Soil Sediment. 2020, 20, 931–942. [Google Scholar] [CrossRef] [Green Version]
- Lan, M.-M.; Liu, C.; Liu, S.-J.; Qiu, R.-L.; Tang, Y.-T. Phytostabilization of Cd and Pb in Highly polluted Farmland soils using ramie and amendments. Int. J. Environ. Res. Public Health 2020, 17, 1661. [Google Scholar] [CrossRef] [Green Version]
- Sheoran, V.; Sheoran, A.; Poonia, P. Phytostabilization of metalliferous mine waste. J. Indus. Pollut. Control 2013, 29, 183–192. [Google Scholar]
- Wong, M.H. Ecological restoration of mine degraded soils, with emphasis on metal contaminated soils. Chemosphere 2003, 50, 775–780. [Google Scholar] [CrossRef]
- Ghosh, M.; Singh, S. A review on phytoremediation of heavy metals and utilization of it’s by products. Asian. J. Energy Environ. 2005, 6, 18. [Google Scholar]
- Sarma, H. Metal hyperaccumulation in plants: A review focusing on phytoremediation technology. J. Environ. Sci. Technol. 2011, 4, 118–138. [Google Scholar] [CrossRef] [Green Version]
- Galal, T.M.; Eid, E.M.; Dakhil, M.A.; Hassan, L.M. Bioaccumulation and rhizofiltration potential of Pistia stratiotes L. for mitigating water pollution in the Egyptian wetlands. Int. J. Phytoremediat. 2018, 20, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Abhilash, P.; Pandey, V.C.; Srivastava, P.; Rakesh, P.; Chandran, S.; Singh, N.; Thomas, A. Phytofiltration of cadmium from water by Limnocharis flava (L.) Buchenau grown in free-floating culture system. J. Hazard. Mater. 2009, 170, 791–797. [Google Scholar] [CrossRef]
- Islam, M.S.; Saito, T.; Kurasaki, M. Phytofiltration of arsenic and cadmium by using an aquatic plant, Micranthemum umbrosum: Phytotoxicity, uptake kinetics, and mechanism. Ecotoxicol. Environ. Saf. 2015, 112, 193–200. [Google Scholar] [CrossRef]
- Dürešová, Z.; Šuňovská, A.; Horník, M.; Pipíška, M.; Gubišová, M.; Gubiš, J.; Hostin, S. Rhizofiltration potential of Arundo donax for cadmium and zinc removal from contaminated wastewater. Chem. Pap. 2014, 68, 1452–1462. [Google Scholar] [CrossRef]
- Olguín, E.J.; Sánchez-Galván, G. Heavy metal removal in phytofiltration and phycoremediation: The need to differentiate between bioadsorption and bioaccumulation. New Biotech. 2012, 30, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Wang, H.; Lu, H.; Jiang, S.; Dai, M.; Liu, J.; Yan, C. Rhizodegradation potential and tolerance of Avicennia marina (Forsk.) Vierh in phenanthrene and pyrene contaminated sediments. Mar. Pollut. Bull. 2016, 110, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Yanai, J.; Zhao, F.-J.; McGrath, S.P.; Kosaki, T. Effect of soil characteristics on Cd uptake by the hyperaccumulator Thlaspi caerulescens. Environ. Pollut. 2006, 139, 167–175. [Google Scholar] [CrossRef]
- Sharma, H.D.; Reddy, K.R. Geoenvironmental Engineering: Site Remediation, Waste Containment, and Emerging Waste Management Technologies; John Wiley and Sons: Hoboken, NJ, USA, 2004. [Google Scholar]
- Chen, M.; Xu, P.; Zeng, G.; Yang, C.; Huang, D.; Zhang, J. Bioremediation of soils contaminated with polycyclic aromatic hydrocarbons, petroleum, pesticides, chlorophenols and heavy metals by composting: Applications, microbes and future research needs. Biotechnol. Adv. 2015, 33, 745–755. [Google Scholar] [CrossRef]
- Li, X.; Zhang, X.; Wang, X.; Yang, X.; Cui, Z. Bioaugmentation-assisted phytoremediation of lead and salinity co-contaminated soil by Suaeda salsa and Trichoderma asperellum. Chemosphere 2019, 224, 716–725. [Google Scholar] [CrossRef]
- Braud, A.M.; Gaudin, P.; Hazotte, A.; Le Guern, C.; Lebeau, T. Chelate-assisted phytoextraction of lead using Fagopyrum esculentum: Laboratory vs. field experiments. Int. J. Phytoremediat. 2019, 21, 1072–1079. [Google Scholar] [CrossRef]
- Bani, A.; Echevarria, G. Can organic amendments replace chemical fertilizers in nickel agromining cropping systems in Albania? Int. J. Phytoremediat. 2019, 21, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Bingöl, N.A.; Özmal, F.; Akın, B. Phytoremediation and Biosorption Potential of Lythrum salicaria L. for Nickel Removal from Aqueous Solutions. Pol. J. Environ. Stud. 2017, 26. [Google Scholar] [CrossRef]
- da Conceição Gomes, M.A.; Hauser-Davis, R.A.; de Souza, A.N.; Vitória, A.P. Metal phytoremediation: General strategies, genetically modified plants and applications in metal nanoparticle contamination. Ecotoxicol. Environ. Saf. 2016, 134, 133–147. [Google Scholar] [CrossRef]
- Nasr, M. Phytomanagement in Egypt: A Sustainable Approach for Clean Environment Coupled with Meeting Future Energy Demand. In Waste Management in MENA Regions; Springer: Berlin/Heidelberg, Germany, 2020; pp. 93–109. [Google Scholar]
- Mesjasz-Przybyłowicz, J.; Nakonieczny, M.; Migula, P.; Augustyniak, M.; Tarnawska, M.; Reimold, U.; Koeberl, C.; Przybyłowicz, W.; Głowacka, E. Uptake of cadmium, lead nickel and zinc from soil and water solutions by the nickel hyperaccumulator Berkheya coddii. Acta Biol. Cracoviensia Ser. Bot. 2004, 46, 75–85. [Google Scholar]
- Ojuederie, O.B.; Babalola, O.O. Microbial and plant-assisted bioremediation of heavy metal polluted environments: A review. Int. J. Environ. Res. Public Health 2017, 14, 1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, P.; Ye, Z.; Lan, C.; Xie, Z.; Shu, W. Chemically assisted phytoextraction of heavy metal contaminated soils using three plant species. Plant Soil 2005, 276, 153–162. [Google Scholar] [CrossRef]
- Mahajan, P.; Kaushal, J. Role of phytoremediation in reducing cadmium toxicity in soil and water. J. Toxicol. 2018, 2018, 4864365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Toppi, L.S.; Gabbrielli, R. Response to cadmium in higher plants. Environ. Exp. Bot. 1999, 41, 105–130. [Google Scholar] [CrossRef]
- Yang, M.; Lin, X.; Yang, X. Impact of Cd on growth and nutrient accumulation of different plant species. Chin. J. Appl. Ecol. 1998, 9, 89–94. [Google Scholar]
- Khaokaew, S.; Landrot, G. A field-scale study of cadmium phytoremediation in a contaminated agricultural soil at Mae Sot District, Tak Province, Thailand: (1) Determination of Cd-hyperaccumulating plants. Chemosphere 2015, 138, 883–887. [Google Scholar] [CrossRef] [PubMed]
- Lux, A.; Martinka, M.; Vaculík, M.; White, P.J. Root responses to cadmium in the rhizosphere: A review. J. Exp. Bot. 2011, 62, 21–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loix, C.; Huybrechts, M.; Vangronsveld, J.; Gielen, M.; Keunen, E.; Cuypers, A. Reciprocal interactions between cadmium-induced cell wall responses and oxidative stress in plants. Front. Plant Sci. 2017, 8, 1867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, J.M.; Lee, D.A.; Schroeder, J.I. Long-distance root-to-shoot transport of phytochelatins and cadmium in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 10118–10123. [Google Scholar] [CrossRef] [Green Version]
- Verret, F.; Gravot, A.; Auroy, P.; Leonhardt, N.; David, P.; Nussaume, L.; Vavasseur, A.; Richaud, P. Overexpression of AtHMA4 enhances root-to-shoot translocation of zinc and cadmium and plant metal tolerance. FEBS Lett. 2004, 576, 306–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Ogo, Y.; Senoura, T.; Nishizawa, N.K.; Nakanishi, H. The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice. Plant Cell Environ. 2012, 35, 1948–1957. [Google Scholar] [CrossRef]
- Uraguchi, S.; Kamiya, T.; Sakamoto, T.; Kasai, K.; Sato, Y.; Nagamura, Y.; Yoshida, A.; Kyozuka, J.; Ishikawa, S.; Fujiwara, T. Low-affinity cation transporter (OsLCT1) regulates cadmium transport into rice grains. Proc. Natl. Acad. Sci. USA 2011, 108, 20959–20964. [Google Scholar] [CrossRef] [Green Version]
- Lux, A.; Vaculík, M.; Martinka, M.; Lišková, D.; Kulkarni, M.G.; Stirk, W.A.; Van Staden, J. Cadmium induces hypodermal periderm formation in the roots of the monocotyledonous medicinal plant Merwilla plumbea. Ann. Bot. 2011, 107, 285–292. [Google Scholar] [CrossRef]
- Sasaki, A.; Yamaji, N.; Yokosho, K.; Ma, J.F. Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice. Plant Cell 2012, 24, 2155–2167. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, R.; Ishimaru, Y.; Senoura, T.; Shimo, H.; Ishikawa, S.; Arao, T.; Nakanishi, H.; Nishizawa, N.K. The OsNRAMP1 iron transporter is involved in Cd accumulation in rice. J. Exp. Bot. 2011, 62, 4843–4850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milner, M.J.; Seamon, J.; Craft, E.; Kochian, L.V. Transport properties of members of the ZIP family in plants and their role in Zn and Mn homeostasis. J. Exp. Bot. 2013, 64, 369–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murata, Y.; Ma, J.F.; Yamaji, N.; Ueno, D.; Nomoto, K.; Iwashita, T. A specific transporter for iron (III)–phytosiderophore in barley roots. Plant J. 2006, 46, 563–572. [Google Scholar] [CrossRef]
- Ishimaru, Y.; Takahashi, R.; Bashir, K.; Shimo, H.; Senoura, T.; Sugimoto, K.; Ono, K.; Yano, M.; Ishikawa, S.; Arao, T. Characterizing the role of rice NRAMP5 in manganese, iron and cadmium transport. Sci. Rep. 2012, 2, 286. [Google Scholar] [CrossRef] [PubMed]
- Nevo, Y.; Nelson, N. The NRAMP family of metal-ion transporters. Biochim. Biophys. Acta Mol. Cell Res. 2006, 1763, 609–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshihara, T.; Hodoshima, H.; Miyano, Y.; Shoji, K.; Shimada, H.; Goto, F. Cadmium inducible Fe deficiency responses observed from macro and molecular views in tobacco plants. Plant Cell Rep. 2006, 25, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Yamaji, N.; Yamane, M.; Kashino-Fujii, M.; Sato, K.; Ma, J.F. The HvNramp5 transporter mediates uptake of cadmium and manganese, but not iron. Plant Physiol. 2016, 172, 1899–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Liu, X.; Peijnenburg, W.J.; Zhao, J.; Chen, X.; Yu, J.; Wu, H. Pathways of cadmium fluxes in the root of the halophyte Suaeda salsa. Ecotoxicol. Environ. Saf. 2012, 75, 1–7. [Google Scholar] [CrossRef]
- Kim, D.Y.; Bovet, L.; Maeshima, M.; Martinoia, E.; Lee, Y. The ABC transporter AtPDR8 is a cadmium extrusion pump conferring heavy metal resistance. Plant J. 2007, 50, 207–218. [Google Scholar] [CrossRef]
- Clemens, S.; Aarts, M.G.; Thomine, S.; Verbruggen, N. Plant science: The key to preventing slow cadmium poisoning. Trend. Plant Sci. 2013, 18, 92–99. [Google Scholar] [CrossRef]
- Migocka, M.; Kosieradzka, A.; Papierniak, A.; Maciaszczyk-Dziubinska, E.; Posyniak, E.; Garbiec, A.; Filleur, S. Two metal-tolerance proteins, MTP1 and MTP4, are involved in Zn homeostasis and Cd sequestration in cucumber cells. J. Exp. Bot. 2015, 66, 1001–1015. [Google Scholar] [CrossRef] [Green Version]
- Migocka, M.; Papierniak, A.; Kosieradzka, A.; Posyniak, E.; Maciaszczyk-Dziubinska, E.; Biskup, R.; Garbiec, A.; Marchewka, T. Cucumber metal tolerance protein Cs MTP 9 is a plasma membrane H+-coupled antiporter involved in the Mn2+ and Cd2+ efflux from root cells. Plant J. 2015, 84, 1045–1058. [Google Scholar] [CrossRef] [Green Version]
- Lekeux, G.; Crowet, J.-M.; Nouet, C.; Joris, M.; Jadoul, A.; Bosman, B.; Carnol, M.; Motte, P.; Lins, L.; Galleni, M. Homology modeling and in vivo functional characterization of the zinc permeation pathway in a heavy metal P-type ATPase. J. Exp. Bot. 2019, 70, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, N.; Sasaki, A.; Xia, J.X.; Yokosho, K.; Ma, J.F. A node-based switch for preferential distribution of manganese in rice. Nat. Commun. 2013, 4, 1–11. [Google Scholar] [CrossRef]
- Satoh-Nagasawa, N.; Mori, M.; Nakazawa, N.; Kawamoto, T.; Nagato, Y.; Sakurai, K.; Takahashi, H.; Watanabe, A.; Akagi, H. Mutations in rice (Oryza sativa) heavy metal ATPase 2 (OsHMA2) restrict the translocation of zinc and cadmium. Plant Cell Physiol. 2012, 53, 213–224. [Google Scholar] [CrossRef]
- Yamaji, N.; Xia, J.; Mitani-Ueno, N.; Yokosho, K.; Ma, J.F. Preferential delivery of zinc to developing tissues in rice is mediated by P-type heavy metal ATPase OsHMA2. Plant Physiol. 2013, 162, 927–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza-Cózatl, D.G.; Jobe, T.O.; Hauser, F.; Schroeder, J.I. Long-distance transport, vacuolar sequestration, tolerance, and transcriptional responses induced by cadmium and arsenic. Curr. Opin. Plant Biol. 2011, 14, 554–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, M.; Ishikawa, S.; Inagaki, K.; Chiba, K.; Hayashi, H.; Yanagisawa, S.; Yoneyama, T. Possible chemical forms of cadmium and varietal differences in cadmium concentrations in the phloem sap of rice plants (Oryza sativa L.). Soil Sci. Plant Nutr. 2010, 56, 839–847. [Google Scholar] [CrossRef] [Green Version]
- Harris, W.R.; Sammons, R.D.; Grabiak, R.C. A speciation model of essential trace metal ions in phloem. J. Inorganic Biochem. 2012, 116, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Uraguchi, S.; Kamiya, T.; Clemens, S.; Fujiwara, T. Characterization of OsLCT1, a cadmium transporter from indica rice (Oryza sativa). Physiol. Plant. 2014, 151, 339–347. [Google Scholar] [CrossRef]
- Park, J.; Song, W.Y.; Ko, D.; Eom, Y.; Hansen, T.H.; Schiller, M.; Lee, T.G.; Martinoia, E.; Lee, Y. The phytochelatin transporters AtABCC1 and AtABCC2 mediate tolerance to cadmium and mercury. Plant J. 2012, 69, 278–288. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, J.; Lu, L.; Zhu, Z.; Yang, X. Functional analysis of CAX2-like transporters isolated from two ecotypes of Sedum alfredii. Biol. Plant. 2016, 60, 37–47. [Google Scholar] [CrossRef]
- Brunetti, P.; Zanella, L.; De Paolis, A.; Di Litta, D.; Cecchetti, V.; Falasca, G.; Barbieri, M.; Altamura, M.M.; Costantino, P.; Cardarelli, M. Cadmium-inducible expression of the ABC-type transporter AtABCC3 increases phytochelatin-mediated cadmium tolerance in Arabidopsis. J. Exp. Bot. 2015, 66, 3815–3829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanquar, V.; Lelièvre, F.; Bolte, S.; Hamès, C.; Alcon, C.; Neumann, D.; Vansuyt, G.; Curie, C.; Schröder, A.; Krämer, U. Mobilization of vacuolar iron by AtNRAMP3 and AtNRAMP4 is essential for seed germination on low iron. EMBO J. 2005, 24, 4041–4051. [Google Scholar] [CrossRef]
- Korenkov, V.; King, B.; Hirschi, K.; Wagner, G.J. Root-selective expression of AtCAX4 and AtCAX2 results in reduced lamina cadmium in field-grown Nicotiana tabacum L. Plant Biotechnol. J. 2009, 7, 219–226. [Google Scholar] [CrossRef]
- Ueno, D.; Yamaji, N.; Kono, I.; Huang, C.F.; Ando, T.; Yano, M.; Ma, J.F. Gene limiting cadmium accumulation in rice. Proc. Natl. Acad. Sci. USA 2010, 107, 16500–16505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyadate, H.; Adachi, S.; Hiraizumi, A.; Tezuka, K.; Nakazawa, N.; Kawamoto, T.; Katou, K.; Kodama, I.; Sakurai, K.; Takahashi, H. OsHMA3, a P1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles. New Phytol. 2011, 189, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Ueno, D.; Milner, M.J.; Yamaji, N.; Yokosho, K.; Koyama, E.; Clemencia Zambrano, M.; Kaskie, M.; Ebbs, S.; Kochian, L.V.; Ma, J.F. Elevated expression of TcHMA3 plays a key role in the extreme Cd tolerance in a Cd-hyperaccumulating ecotype of Thlaspi caerulescens. Plant J. 2011, 66, 852–862. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, M.; Shohag, M.J.I.; Tian, S.; Song, H.; Feng, Y.; Yang, X. Enhanced expression of SaHMA3 plays critical roles in Cd hyperaccumulation and hypertolerance in Cd hyperaccumulator Sedum alfredii Hance. Planta 2016, 243, 577–589. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, H.; Wu, L.; Liu, A.; Zhao, F.J.; Xu, W. Heavy metal ATPase 3 (HMA3) confers cadmium hypertolerance on the cadmium/zinc hyperaccumulator Sedum plumbizincicola. New Phytol. 2017, 215, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Krämer, U.; Talke, I.N.; Hanikenne, M. Transition metal transport. FEBS Let. 2007, 581, 2263–2272. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, N.; Mori, S.; Baba, K.; Kaburagi-Yada, S.; Arao, T.; Kitajima, N.; Hokura, A.; Terada, Y. Cadmium distribution in the root tissues of solanaceous plants with contrasting root-to-shoot Cd translocation efficiencies. Environ. Exp. Bot. 2011, 71, 198–206. [Google Scholar] [CrossRef]
- Seregin, I.; Ivanov, V. Histochemical investigation of cadmium and lead distribution in plants. Russ. J. Plant Physiol. 1997, 44, 791–796. [Google Scholar]
- Sidhu, G.P.S.; Bali, A.S.; Bhardwaj, R. Use of fungi in mitigating cadmium toxicity in plants. In Cadmium Toxicity and Tolerance in Plants; Hasanuzzaman, M., Prasad, M.N.V., Fujita, M., Eds.; Academic Press: Cambridge, UK, 2019; pp. 397–426. [Google Scholar]
- Tan, L.; Qu, M.; Zhu, Y.; Peng, C.; Wang, J.; Gao, D.; Chen, C. Zinc TRANSPORTER5 and zinc TRANSPORTER9 function synergistically in zinc/cadmium uptake. Plant Physiol. 2020, 183, 1235–1249. [Google Scholar] [CrossRef]
- Yuan, L.; Yang, S.; Liu, B.; Zhang, M.; Wu, K. Molecular characterization of a rice metal tolerance protein, OsMTP1. Plant Cell Rep. 2012, 31, 67–79. [Google Scholar] [CrossRef]
- Song, Y.; Hudek, L.; Freestone, D.; Puhui, J.; Michalczyk, A.; Senlin, Z.; Ackland, M. Comparative analyses of cadmium and zinc uptake correlated with changes in natural resistance-associated macrophage protein (NRAMP) expression in Solanum nigrum L. and Brassica rapa. Environ. Chem. 2014, 11, 653–660. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Kim, Y.-Y.; Lee, Y.; An, G. Rice P1B-type heavy-metal ATPase, OsHMA9, is a metal efflux protein. Plant Physiol. 2007, 145, 831–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bækgaard, L.; Mikkelsen, M.D.; Sørensen, D.M.; Hegelund, J.N.; Persson, D.P.; Mills, R.F.; Yang, Z.; Husted, S.; Andersen, J.P.; Buch-Pedersen, M. A combined zinc/cadmium sensor and zinc/cadmium export regulator in a heavy metal pump. J. Biol. Chem. 2010, 285, 31243–31252. [Google Scholar] [CrossRef] [Green Version]
- Moons, A. Ospdr9, which encodes a PDR-type ABC transporter, is induced by heavy metals, hypoxic stress and redox perturbations in rice roots. FEBS Lett. 2003, 553, 370–376. [Google Scholar] [CrossRef] [Green Version]
- Hirschi, K.D.; Korenkov, V.D.; Wilganowski, N.L.; Wagner, G. Expression of Arabidopsis CAX2 in tobacco. Altered metal accumulation and increased manganese tolerance. Plant Physiol. 2000, 124, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Shigaki, T.; Pittman, J.K.; Hirschi, K.D. Manganese Specificity Determinants in the Arabidopsis Metal/H+ Antiporter CAX2. J. Biol. Chem. 2003, 278, 6610–6617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, S.; Tan, J.; Zhang, Y.; Liang, S.; Xiang, S.; Wang, H.; Chai, T. Isolation and characterization of a novel cadmium-regulated yellow stripe-like transporter (SnYSL3) in Solanum nigrum. Plant Cell Rep. 2017, 36, 281–296. [Google Scholar] [CrossRef]
- Koike, S.; Inoue, H.; Mizuno, D.; Takahashi, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. OsYSL2 is a rice metal-nicotianamine transporter that is regulated by iron and expressed in the phloem. Plant J. 2004, 39, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, Y.; Zheng, L.; Li, Y.; Zhou, X.; Li, J.; Gu, D.; Xu, E.; Lu, Y.; Chen, X. The intracellular transporter AtNRAMP6 is involved in Fe homeostasis in Arabidopsis. Front. Plant Sci. 2019, 10, 1124. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Kim, S.A.; Lee, J.; Guerinot, M.L.; An, G. Zinc deficiency-inducible OsZIP8 encodes a plasma membrane-localized zinc transporter in rice. Mol. Cell. 2010, 29, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Jiang, J.; Li, S.; Li, M.; Tan, Y.; Song, S.; Shu, Q.; Huang, J. Glutamate alleviates cadmium toxicity in rice via suppressing cadmium uptake and translocation. J. Hazard. Mater. 2020, 384, 121319. [Google Scholar] [CrossRef]
- Song, W.-Y.; Park, J.; Mendoza-Cózatl, D.G.; Suter-Grotemeyer, M.; Shim, D.; Hörtensteiner, S.; Geisler, M.; Weder, B.; Rea, P.A.; Rentsch, D. Arsenic tolerance in Arabidopsis is mediated by two ABCC-type phytochelatin transporters. Proc. Natl. Acad. Sci. USA 2010, 107, 21187–21192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mary, V.; Ramos, M.S.; Gillet, C.; Socha, A.L.; Giraudat, J.; Agorio, A.; Merlot, S.; Clairet, C.; Kim, S.A.; Punshon, T. Bypassing iron storage in endodermal vacuoles rescues the iron mobilization defect in the natural resistance associated-macrophage protein3natural resistance associated-macrophage protein4 double mutant. Plant Physiol. 2015, 169, 748–759. [Google Scholar] [CrossRef] [Green Version]
- Hassan, Z.; Aarts, M.G. Opportunities and feasibilities for biotechnological improvement of Zn, Cd or Ni tolerance and accumulation in plants. Environ. Exp. Bot. 2011, 72, 53–63. [Google Scholar] [CrossRef]
- Kawachi, M.; Kobae, Y.; Kogawa, S.; Mimura, T.; Krämer, U.; Maeshima, M. Amino acid screening based on structural modeling identifies critical residues for the function, ion selectivity and structure of Arabidopsis MTP1. FEBS J. 2012, 279, 2339–2356. [Google Scholar] [CrossRef]
- Deng, F.; Yamaji, N.; Xia, J.; Ma, J.F. A member of the heavy metal P-type ATPase OsHMA5 is involved in xylem loading of copper in rice. Plant Physiol. 2013, 163, 1353–1362. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Fujii, Y.; Yamaji, N.; Masuda, S.; Takemoto, Y.; Kamiya, T.; Yusuyin, Y.; Iwasaki, K.; Kato, S.-I.; Maeshima, M. Mn tolerance in rice is mediated by MTP8. 1, a member of the cation diffusion facilitator family. J. Exp. Bot. 2013, 64, 4375–4387. [Google Scholar] [CrossRef] [Green Version]
- Bahmani, R.; Kim, D.; Na, J.; Hwang, S. Expression of the tobacco non-symbiotic class 1 hemoglobin gene Hb1 reduces cadmium levels by modulating Cd transporter expression through decreasing nitric oxide and ROS level in Arabidopsis. Front. Plant Sci. 2019, 10, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.; Yang, M.; Li, Y.; Tian, J.; Zhang, Y.; Liang, L.; Liu, Z.; Chen, K.; Li, Y.; Lv, K. Comprehensive analysis of variation of cadmium accumulation in rice and detection of a new weak allele of OsHMA3. J. Exp. Bot. 2019, 70, 6389–6400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Yu, K.-F.; Poysa, V.; Shi, C.; Zhou, Y.-H. A single point mutation in GmHMA3 affects cadimum (Cd) translocation and accumulation in soybean seeds. Mol. Plant 2012, 5, 1154–1156. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, D.N.; Carvalho, M.E.A.; Piotto, F.A.; Batagin-Piotto, K.D.; Nogueira, M.L.; Gaziola, S.A.; Azevedo, R.A. Antioxidant defense response in plants to cadmium stress. In Cadmium Tolerance in Plants: Agronomic, Molecular, Signaling, and Omic Approaches; Hasanuzzaman, M., Prasad, M.N.V., Nahar, L., Eds.; Academic Press: New York, NY, USA, 2019; pp. 423–461. [Google Scholar]
- Guo, J.; Qin, S.; Rengel, Z.; Gao, W.; Nie, Z.; Liu, H.; Li, C.; Zhao, P. Cadmium stress increases antioxidant enzyme activities and decreases endogenous hormone concentrations more in Cd-tolerant than Cd-sensitive wheat varieties. Ecotoxicol. Environ. Saf. 2019, 172, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.; Nahakpam, S. Heat exposure alters the expression of SOD, POD, APX and CAT isozymes and mitigates low cadmium toxicity in seedlings of sensitive and tolerant rice cultivars. Plant Physiol. Biochem. 2012, 57, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Forde, B.G. Glutamate signalling in roots. J. Exp. Bot. 2014, 65, 779–787. [Google Scholar] [CrossRef] [Green Version]
- Nie, J.; Liu, Y.; Zeng, G.; Zheng, B.; Tan, X.; Liu, H.; Xie, J.; Gan, C.; Liu, W. Cadmium accumulation and tolerance of Macleaya cordata: A newly potential plant for sustainable phytoremediation in Cd-contaminated soil. Environ. Sci. Pollut. Res. 2016, 23, 10189–10199. [Google Scholar] [CrossRef]
- Arshad, M.; Ali, S.; Noman, A.; Ali, Q.; Rizwan, M.; Farid, M.; Irshad, M.K. Phosphorus amendment decreased cadmium (Cd) uptake and ameliorates chlorophyll contents, gas exchange attributes, antioxidants, and mineral nutrients in wheat (Triticum aestivum L.) under Cd stress. Arch. Agron. Soil Sci. 2016, 62, 533–546. [Google Scholar] [CrossRef]
- Pan, C.; Lu, H.; Yu, J.; Liu, J.; Liu, Y.; Yan, C. Identification of Cadmium-responsive Kandelia obovata SOD family genes and response to Cd toxicity. Environ. Exp. Bot. 2019, 162, 230–238. [Google Scholar] [CrossRef]
- Jung, H.-i.; Lee, B.-R.; Chae, M.-J.; Kong, M.-S.; Lee, C.-H.; Kang, S.-S.; Kim, Y.-H. Sulfur alleviates cadmium toxicity in rice (Oryza sativa L.) seedlings by altering antioxidant levels. J. Crop Sci. Biotechnol. 2017, 20, 213–220. [Google Scholar] [CrossRef]
- Ci, D.; Jiang, D.; Dai, T.; Jing, Q.; Cao, W. Effects of cadmium on plant growth and physiological traits in contrast wheat recombinant inbred lines differing in cadmium tolerance. Chemosphere 2009, 77, 1620–1625. [Google Scholar] [CrossRef] [PubMed]
- Alyemeni, M.N.; Ahanger, M.A.; Wijaya, L.; Alam, P.; Ahmad, P. Contrasting tolerance among soybean genotypes subjected to different levels of cadmium stress. Pak. J. Bot. 2017, 49, 903–911. [Google Scholar]
- Gratao, P.L.; Monteiro, C.C.; Tezotto, T.; Carvalho, R.F.; Alves, L.R.; Peters, L.P.; Azevedo, R.A. Cadmium stress antioxidant responses and root-to-shoot communication in grafted tomato plants. Biometals 2015, 28, 803–816. [Google Scholar] [CrossRef]
- Nouairi, I.; Ammar, W.B.; Youssef, N.B.; Miled, D.D.B.; Ghorbal, M.H.; Zarrouk, M. Antioxidant defense system in leaves of Indian mustard (Brassica juncea) and rape (Brassica napus) under cadmium stress. Acta Physiol. Plant. 2009, 31, 237–247. [Google Scholar] [CrossRef]
- Iqbal, N.; Masood, A.; Nazar, R.; Syeed, S.; Khan, N.A. Photosynthesis, growth and antioxidant metabolism in mustard (Brassica juncea L.) cultivars differing in cadmium tolerance. Agric. Sci. China 2010, 9, 519–527. [Google Scholar] [CrossRef]
- Luo, H.; Li, H.; Zhang, X.; Fu, J. Antioxidant responses and gene expression in perennial ryegrass (Lolium perenne L.) under cadmium stress. Ecotoxicology 2011, 20, 770–778. [Google Scholar] [CrossRef]
- Gill, S.S.; Khan, N.A.; Tuteja, N. Differential cadmium stress tolerance in five Indian mustard (Brassica juncea L.) cultivars: An evaluation of the role of antioxidant machinery. Plant Signal. Behave. 2011, 6, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Zhao, X.; Sun, X.; Tan, Q.; Tang, Y.; Nie, Z.; Qu, C.; Chen, Z.; Hu, C. Antioxidant enzyme systems and the ascorbate–glutathione cycle as contributing factors to cadmium accumulation and tolerance in two oilseed rape cultivars (Brassica napus L.) under moderate cadmium stress. Chemosphere 2015, 138, 526–536. [Google Scholar] [CrossRef]
- Mesnoua, M.; Mateos-Naranjo, E.; Barcia-Piedras, J.M.; Pérez-Romero, J.A.; Lotmani, B.; Redondo-Gómez, S. Physiological and biochemical mechanisms preventing Cd-toxicity in the hyperaccumulator Atriplex halimus L. Plant Physiol. Biochem. 2016, 106, 30–38. [Google Scholar] [CrossRef]
- Asopa, P.P.; Bhatt, R.; Sihag, S.; Kothari, S.; Kachhwaha, S. Effect of cadmium on physiological parameters of cereal and millet plants—A comparative study. Int. J. Phytoremediat. 2017, 19, 225–230. [Google Scholar] [CrossRef]
- Ashrafzadeh, S.; Leung, D.W. Novel potato plants with enhanced cadmium resistance and antioxidative defence generated after in vitro cell line selection. PLoS ONE 2017, 12, e0185621. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Huang, Y.; Wu, X.; Liu, Z.; Zou, J.; Chen, Y.; Su, N.; Cui, J. Increased antioxidative capacity and decreased cadmium uptake contribute to hemin-induced alleviation of cadmium toxicity in Chinese cabbage seedlings. Ecotoxicol. Environ. Saf. 2019, 177, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Sozoniuk, M.; Nowak, M.; Dudziak, K.; Bulak, P.; Leśniowska-Nowak, J.; Kowalczyk, K. Antioxidative system response of pedunculate oak (Quercus robur L.) seedlings to Cd exposure. Physiol. Mol. Biol. Plants 2019, 25, 1377–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Lu, L.; Tian, S.; Li, S.; Liu, X.; Gao, X.; Zhou, W.; Lin, X. Cadmium-induced nitric oxide burst enhances Cd tolerance at early stage in roots of a hyperaccumulator Sedum alfredii partially by altering glutathione metabolism. Sci. Total Environ. 2019, 650, 2761–2770. [Google Scholar] [CrossRef] [PubMed]
- Khalid, S.; Shahid, M.; Niazi, N.K.; Murtaza, B.; Bibi, I.; Dumat, C. A comparison of technologies for remediation of heavy metal contaminated soils. J. Geochem. Exp. 2017, 182, 247–268. [Google Scholar] [CrossRef] [Green Version]
- Evangelou, M.W.; Ebel, M.; Schaeffer, A. Chelate assisted phytoextraction of heavy metals from soil. Effect, mechanism, toxicity, and fate of chelating agents. Chemosphere 2007, 68, 989–1003. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, A.; Carmona, F.F.; Bhargava, M.; Srivastava, S. Approaches for enhanced phytoextraction of heavy metals. J. Environ. Manag. 2012, 105, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Luo, D.; Yao, G.; Huang, X.; Wei, L.; Liu, Y.; Wu, Q.; Mai, X.; Liu, G.; Xiao, T. Comparative Activation Process of Pb, Cd and Tl Using Chelating Agents from Contaminated Red Soils. Int. J. Environ. Res. Public Health 2020, 17, 497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, M.; Uddin, M.; Ara-Sharmeen, I.; Alharby, F.H.; Alzahrani, Y.; Hakeem, K.R.; Zhang, L. Assisting phytoremediation of heavy metals using chemical amendments. Plants 2019, 8, 295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, C.; Shen, Z.; Li, X. Enhanced phytoextraction of Cu, Pb, Zn and Cd with EDTA and EDDS. Chemosphere 2005, 59, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Engelen, D.L.; Sharpe-Pedler, R.C.; Moorhead, K.K. Effect of chelating agents and solubility of cadmium complexes on uptake from soil by Brassica juncea. Chemosphere 2007, 68, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Evangelou, M.W.; Bauer, U.; Ebel, M.; Schaeffer, A. The influence of EDDS and EDTA on the uptake of heavy metals of Cd and Cu from soil with tobacco Nicotiana tabacum. Chemosphere 2007, 68, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.H.; Ali, S.; Rehman, M.; Hasanuzzaman, M.; Rizwan, M.; Irshad, S.; Shafiq, F.; Iqbal, M.; Alharbi, B.M.; Alnusaire, T.S. Jute: A potential candidate for phytoremediation of metals—A review. Plants 2020, 9, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.; Wang, X.; Guo, Z.; Xiao, X.; Peng, C.; Yang, J.; Zhou, C.; Zeng, P. Chelator-assisted phytoextraction of arsenic, cadmium and lead by Pteris vittata L. and soil microbial community structure response. Int. J. Phytoremed. 2019, 21, 1032–1040. [Google Scholar] [CrossRef]
- Turgut, C.; Pepe, M.K.; Cutright, T.J. The effect of EDTA and citric acid on phytoremediation of Cd, Cr, and Ni from soil using Helianthus annuus. Environ. Pollut. 2004, 131, 147–154. [Google Scholar] [CrossRef]
- Bian, X.; Cui, J.; Tang, B.; Yang, L. Chelant-induced phytoextraction of heavy metals from contaminated soils: A review. Pol. J. Environ. Stud. 2018, 27, 2417–2424. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.W.; Wong, W.W.; Wei, Z.; Jagadeesan, H. Alkaline biosolids and EDTA for phytoremediation of an acidic loamy soil spiked with cadmium. Sci. Total Environ. 2004, 324, 235–246. [Google Scholar] [CrossRef]
- Tahmasbian, I.; Sinegani, A.S. Chelate-assisted phytoextraction of cadmium from a mine soil by negatively charged sunflower. Int. J. Environ. Sci. Technol. 2014, 11, 695–702. [Google Scholar] [CrossRef] [Green Version]
- Redjala, T.; Sterckeman, T.; Morel, J.L. Cadmium uptake by roots: Contribution of apoplast and of high-and low-affinity membrane transport systems. Environ. Exp. Bot. 2009, 67, 235–242. [Google Scholar] [CrossRef]
- Tack, F.M.; Meers, E. Assisted phytoextraction: Helping plants to help us. Elements 2010, 6, 383–388. [Google Scholar] [CrossRef]
- Nowack, B.; Schulin, R.; Robinson, B.H. Critical assessment of chelant-enhanced metal phytoextraction. Environ. Sci. Technol. 2006, 40, 5225–5232. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, N.; Hermans, C.; Schat, H. Molecular mechanisms of metal hyperaccumulation in plants. New Phytol. 2009, 181, 759–776. [Google Scholar] [CrossRef] [PubMed]
- Anjum, N.A.; Hasanuzzaman, M.; Hossain, M.A.; Thangavel, P.; Roychoudhury, A.; Gill, S.S.; Rodrigo, M.A.M.; Adam, V.; Fujita, M.; Kizek, R. Jacks of metal/metalloid chelation trade in plants—An overview. Front. Plant Sci. 2015, 6, 192. [Google Scholar] [CrossRef] [Green Version]
- Sytar, O.; Kumar, A.; Latowski, D.; Kuczynska, P.; Strzałka, K.; Prasad, M. Heavy metal-induced oxidative damage, defense reactions, and detoxification mechanisms in plants. Acta Physiol. Plant. 2013, 35, 985–999. [Google Scholar] [CrossRef]
- Filiz, E.; Saracoglu, I.A.; Ozyigit, I.I.; Yalcin, B. Comparative analyses of phytochelatin synthase (PCS) genes in higher plants. Biotechnol. Biotechnol. Equip. 2019, 33, 178–194. [Google Scholar] [CrossRef] [Green Version]
- Vatamaniuk, O.K.; Mari, S.; Lu, Y.-P.; Rea, P.A. Mechanism of heavy metal ion activation of phytochelatin (PC) synthase blocked thiols are sufficient for PC synthase-catalyzed transpeptidation of glutathione and related thiol peptides. J. Biol. Chem. 2000, 275, 31451–31459. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.D.; Hwang, S. Tobacco phytochelatin synthase (NtPCS1) plays important roles in cadmium and arsenic tolerance and in early plant development in tobacco. Plant Biotechnol. Rep. 2015, 9, 107–114. [Google Scholar] [CrossRef]
- Franchi, N.; Piccinni, E.; Ferro, D.; Basso, G.; Spolaore, B.; Santovito, G.; Ballarin, L. Characterization and transcription studies of a phytochelatin synthase gene from the solitary tunicate Ciona intestinalis exposed to cadmium. Aquatic Toxicol. 2014, 152, 47–56. [Google Scholar] [CrossRef]
- Fu, S.; Lu, Y.; Zhang, X.; Yang, G.; Chao, D.; Wang, Z.; Shi, M.; Chen, J.; Chao, D.-Y.; Li, R. The ABC transporter ABCG36 is required for cadmium tolerance in rice. J. Exp. Bot. 2019, 70, 5909–5918. [Google Scholar] [CrossRef]
- Iqbal, M.; Ahmad, A.; Ansari, M.; Qureshi, M.; Aref, I.M.; Khan, P.R.; Hegazy, S.S.; El-Atta, H.; Husen, A.; Hakeem, K.R. Improving the phytoextraction capacity of plants to scavenge metal (loid)-contaminated sites. Environ. Rev. 2015, 23, 44–65. [Google Scholar] [CrossRef]
- Küpper, H.; Mijovilovich, A.; Meyer-Klaucke, W.; Kroneck, P.M. Tissue-and age-dependent differences in the complexation of cadmium and zinc in the cadmium/zinc hyperaccumulator Thlaspi caerulescens (Ganges ecotype) revealed by X-ray absorption spectroscopy. Plant Physiol. 2004, 134, 748–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, R.-L.; Zhou, Q.-X.; Sun, F.-H.; Jin, C.-X. Antioxidative defense and proline/phytochelatin accumulation in a newly discovered Cd-hyperaccumulator, Solanum nigrum L. Environ. Exp. Bot. 2007, 60, 468–476. [Google Scholar] [CrossRef]
- Xu, D.; Chen, Z.; Sun, K.; Yan, D.; Kang, M.; Zhao, Y. Effect of cadmium on the physiological parameters and the subcellular cadmium localization in the potato (Solanum tuberosum L.). Ecotoxicol. Environ. Saf. 2013, 97, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Yao, P.; Pan, J.; Dai, C.; Cao, H.; Chen, Z.; Zhang, S.; Xu, S.; Shen, W. Transcriptome analysis reveals insight into molecular hydrogen-induced cadmium tolerance in alfalfa: The prominent role of sulfur and (homo) glutathione metabolism. BMC Plant Biol. 2020, 20, 1–19. [Google Scholar] [CrossRef]
- Hossain, M.A.; Piyatida, P.; da Silva, J.A.T.; Fujita, M. Molecular mechanism of heavy metal toxicity and tolerance in plants: Central role of glutathione in detoxification of reactive oxygen species and methylglyoxal and in heavy metal chelation. J. Bot. 2012, 2012. [Google Scholar] [CrossRef]
- Jiang, W.; Wu, Z.; Wang, T.; Mantri, N.; Huang, H.; Li, H.; Tao, Z.; Guo, Q. Physiological and transcriptomic analyses of cadmium stress response in Dendrobium officinale seedling. Plant Physiol. Biochem. 2020, 148, 152–165. [Google Scholar] [CrossRef]
- DalCorso, G.; Farinati, S.; Furini, A. Regulatory networks of cadmium stress in plants. Plant Signal. Behav. 2010, 5, 663–667. [Google Scholar] [CrossRef]
- Mendoza-Cózatl, D.G.; Butko, E.; Springer, F.; Torpey, J.W.; Komives, E.A.; Kehr, J.; Schroeder, J.I. Identification of high levels of phytochelatins, glutathione and cadmium in the phloem sap of Brassica napus. A role for thiol-peptides in the long-distance transport of cadmium and the effect of cadmium on iron translocation. Plant J. 2008, 54, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Freisinger, E. Plant MTs—Long neglected members of the metallothionein superfamily. Dalton Trans. 2008, 47, 6663–6675. [Google Scholar] [CrossRef]
- Ziller, A.; Fraissinet-Tachet, L. Metallothionein diversity and distribution in the tree of life: A multifunctional protein. Metallomics 2018, 10, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Heavy metal stress and some mechanisms of plant defense response. Sci. World J. 2015, 2015. [Google Scholar] [CrossRef]
- Singh, G.; Tripathi, S.; Shanker, K.; Sharma, A. Cadmium-induced conformational changes in type 2 metallothionein of medicinal plant Coptis japonica: Insights from molecular dynamics studies of apo, partially and fully metalated forms. J. Biomol. Struct. Dyn. 2019, 37, 1520–1533. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Zhu, M.; Wang, S.; Ma, G.; Huang, X.; Qiao, C.; Wang, R.; Xu, X.; Liang, Y.; Lu, K. Genome-wide characterization and analysis of metallothionein family genes that function in metal stress tolerance in Brassica napus L. Int. J. Mol. Sci. 2018, 19, 2181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.-J.; Meetam, M.; Goldsbrough, P.B. Examining the specific contributions of individual Arabidopsis metallothioneins to copper distribution and metal tolerance. Plant Physiol. 2008, 146, 1697–1706. [Google Scholar] [CrossRef] [Green Version]
- Li, L.-S.; Meng, Y.-P.; Cao, Q.-F.; Yang, Y.-Z.; Wang, F.; Jia, H.-S.; Wu, S.-B.; Liu, X.-G. Type 1 metallothionein (ZjMT) is responsible for heavy metal tolerance in Ziziphus jujuba. Biochemistry 2016, 81, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Pan, Y.; Zhai, J.; Xiong, Y.; Li, J.; Du, X.; Su, C.; Zhang, X. Cucumber metallothionein-like 2 (CsMTL2) exhibits metal-binding properties. Genes 2016, 7, 106. [Google Scholar] [CrossRef] [Green Version]
- Duan, L.; Yu, J.; Xu, L.; Tian, P.; Hu, X.; Song, X.; Pan, Y. Functional characterization of a type 4 metallothionein gene (CsMT4) in cucumber. Hortic. Plant J. 2019, 5, 120–128. [Google Scholar] [CrossRef]
- Balasundaram, U.; Venkataraman, G.; George, S.; Parida, A. Metallothioneins from a hyperaccumulating plant Prosopis juliflora show difference in heavy metal accumulation in transgenic tobacco. Int. J. Agric. Environ. Biotechnol. 2014, 7, 241–246. [Google Scholar] [CrossRef]
- Gonzalez-Mendoza, D.; Moreno, A.Q.; Zapata-Perez, O. Coordinated responses of phytochelatin synthase and metallothionein genes in black mangrove, Avicennia germinans, exposed to cadmium and copper. Aquatic Toxicol. 2007, 83, 306–314. [Google Scholar] [CrossRef]
- Ruta, L.L.; Lin, Y.-F.; Kissen, R.; Nicolau, I.; Neagoe, A.D.; Ghenea, S.; Bones, A.M.; Farcasanu, I.C. Anchoring plant metallothioneins to the inner face of the plasma membrane of Saccharomyces cerevisiae cells leads to heavy metal accumulation. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Zhu, Y.X.; Du, W.X.; Fang, X.Z.; Zhang, L.L.; Jin, C.W. Knockdown of BTS may provide a new strategy to improve cadmium-phytoremediation efficiency by improving iron status in plants. J. Hazard. Mater. 2020, 384, 121473. [Google Scholar] [CrossRef] [PubMed]
- Clemens, S.; Palmgren, M.G.; Krämer, U. A long way ahead: Understanding and engineering plant metal accumulation. Trend. Plant Sci. 2002, 7, 309–315. [Google Scholar] [CrossRef]
- Zhao, Q.; Wang, H.; Du, Y.; Rogers, H.J.; Wu, Z.; Jia, S.; Yao, X.; Xie, F.; Liu, W. MSH2 and MSH6 in Mismatch Repair System Account for Soybean (Glycine max (L.) Merr.) Tolerance to Cadmium Toxicity by Determining DNA Damage Response. J. Agric. Food Chem. 2020, 68, 1974–1985. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, Y.; Fu, Y.; Xie, H.; Song, S.; Qiu, M.; Wen, J.; Chen, M.; Chen, G.; Tian, Y. Mutation at different sites of metal transporter gene OsNramp5 affects Cd accumulation and related agronomic traits in rice (Oryza sativa L.). Front. Plant Sci. 2019, 10, 1081. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Xiao, X.; Yan, G.; Hu, J.; Cheng, X.; Li, L.; Li, H.; Wu, X. Association mapping of cadmium-tolerant QTLs in Brassica napus L. and insight into their contributions to phytoremediation. Environ. Exp. Bot. 2018, 155, 420–428. [Google Scholar] [CrossRef]
- Chen, L.; Wan, H.; Qian, J.; Guo, J.; Sun, C.; Wen, J.; Yi, B.; Ma, C.; Tu, J.; Song, L. Genome-wide association study of cadmium accumulation at the seedling stage in rapeseed (Brassica napus L.). Front. Plant Sci. 2018, 9, 375. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Chen, X.; Zhao, J.; Wang, S.; Tan, L.; Sun, C.; Liu, F. Identification of QTLs related to cadmium tolerance from wild rice (Oryza nivara) using a high-density genetic map for a set of introgression lines. Euphytica 2019, 215, 205. [Google Scholar] [CrossRef]
- Chao, D.-Y.; Silva, A.; Baxter, I.; Huang, Y.S.; Nordborg, M.; Danku, J.; Lahner, B.; Yakubova, E.; Salt, D.E. Genome-wide association studies identify heavy metal ATPase3 as the primary determinant of natural variation in leaf cadmium in Arabidopsis thaliana. PLoS Genet. 2012, 8, e1002923. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Mao, B.; Li, Y.; Lv, Q.; Zhang, L.; Chen, C.; He, H.; Wang, W.; Zeng, X.; Shao, Y. Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.-q.; Bai, Z.-Y.; Xiao, Y.-F.; Li, Y.; Liu, Q.-L.; Zhang, L.; Pan, Y.-Z.; Jiang, B.-B.; Zhang, F. Transcriptomic analysis of Verbena bonariensis roots in response to cadmium stress. BMC Genom. 2019, 20, 877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalyan, E.; Yüzbaşıoğlu, E.; Keskin, B.C.; Yıldızhan, Y.; Memon, A.; Ünal, M.; Yüksel, B. The identification of genes associated with Pb and Cd response mechanism in Brassica juncea L. by using Arabidopsis expression array. Environ. Exp. Bot. 2017, 139, 105–115. [Google Scholar] [CrossRef]
- Wang, J.; Li, H.; Zou, D.; Zhao, J.; Fan, L.; Wu, T. Transcriptome profile analysis of cadmium tolerance in Chinese flowering cabbage. Hortic. Environ. Biotechnol. 2017, 58, 56–65. [Google Scholar] [CrossRef]
- Yu, R.; Ma, Y.; Li, Y.; Li, X.; Liu, C.; Du, X.; Shi, G. Comparative transcriptome analysis revealed key factors for differential cadmium transport and retention in roots of two contrasting peanut cultivars. BMC Genom. 2018, 19, 938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Bai, Y.; Chao, Y.; Sun, X.; He, C.; Liang, X.; Xie, L.; Han, L. Genome-wide analysis reveals four key transcription factors associated with cadmium stress in creeping bentgrass (Agrostis stolonifera L.). PeerJ 2018, 6, e5191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.; Zhang, Y.; He, Y.; Cao, Q.; Zhang, T.; Lou, L.; Cai, Q. Full-length transcriptome assembly of italian ryegrass root integrated with RNA-Seq to identify genes in response to plant cadmium stress. Int. J. Mol. Sci. 2020, 21, 1067. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhi, J.; Zhang, H.; Li, J.; Zhao, Q.; Xu, J. Transcriptome analysis of Phytolacca americana L. in response to cadmium stress. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Yu, C.; Xia, X.; Li, M.; Li, H.; Wang, Y.; Wang, S.; Wang, C.; Ma, Y.; Zhou, G. Comparative transcriptome analysis of duckweed (Landoltia punctata) in response to cadmium provides insights into molecular mechanisms underlying hyperaccumulation. Chemosphere 2018, 190, 154–165. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, S.; Cheng, Z.; Li, T.; Jia, Y.; Wang, G.; Yang, Z.; Xian, J.; Yang, Y.; Zhou, W. Transcriptome analysis revealed cadmium accumulation mechanisms in hyperaccumulator Siegesbeckia orientalis L. Environ. Sci. Pollut. Res. 2020, 27, 18853–18865. [Google Scholar] [CrossRef]
- Liu, A.; Zhou, Z.; Yi, Y.; Chen, G. Transcriptome analysis reveals the roles of stem nodes in cadmium transport to rice grain. BMC Genom. 2020, 21, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Jien, S.H.; Lin, Y.H. Proteins in xylem exudates from rapeseed plants (Brassica napus L.) play a crucial role in cadmium phytoremediation. Clean 2018, 46, 1700164. [Google Scholar] [CrossRef]
- Durand, T.C.; Sergeant, K.; Planchon, S.; Carpin, S.; Label, P.; Morabito, D.; Hausman, J.F.; Renaut, J. Acute metal stress in Populus tremula× P. alba (717-1B4 genotype): Leaf and cambial proteome changes induced by cadmium2+. Proteomics 2010, 10, 349–368. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.K.; Cho, S.-W.; Kwon, S.J.; Kamal, A.H.M.; Kim, S.-W.; Oh, M.-W.; Lee, M.-S.; Chung, K.-Y.; Xin, Z.; Woo, S.-H. Morpho-physiological and proteome level responses to cadmium stress in sorghum. PLoS ONE 2016, 11, e0150431. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Sheng, H.; Li, X.; Wang, L. iTRAQ-based proteomic analysis reveals the mechanisms of silicon-mediated cadmium tolerance in rice (Oryza sativa) cells. Plant Physiol. Biochem. 2016, 104, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Lan, X.-Y.; Yan, Y.-Y.; Yang, B.; Li, X.-Y.; Xu, F.-L. Differential expression of proteins in the leaves and roots of cadmium-stressed Microsorum pteropus, a novel potential aquatic cadmium hyperaccumulator. Sci. Total Environ. 2018, 642, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, J.M.; Chacón-Madrid, K.; Galazzi, R.M.; Campos, B.K.; Arruda, S.C.; Azevedo, R.A.; Arruda, M.A. Evaluation of silicon influence on the mitigation of cadmium-stress in the development of Arabidopsis thaliana through total metal content, proteomic and enzymatic approaches. J. Trace Elem. Med. Biol. 2017, 44, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M.; Li, S.; Huang, F.; Qiu, J.; Zhang, J.; Sheng, Z.; Tang, S.; Wei, X.; Hu, P. The phosphoproteomic response of rice seedlings to cadmium stress. Int. J. Mol. Sci. 2017, 18, 2055. [Google Scholar] [CrossRef] [Green Version]
- Gong, B.; Nie, W.; Yan, Y.; Gao, Z.; Shi, Q. Unravelling cadmium toxicity and nitric oxide induced tolerance in Cucumis sativus: Insight into regulatory mechanisms using proteomics. J. Hazard. Mater. 2017, 336, 202–213. [Google Scholar] [CrossRef]
- Borges, K.L.R.; Salvato, F.; Loziuk, P.L.; Muddiman, D.C.; Azevedo, R.A. Quantitative proteomic analysis of tomato genotypes with differential cadmium tolerance. Environ. Sci. Pollut. Res. 2019, 26, 26039–26051. [Google Scholar] [CrossRef]
- Pernía, B.; Calabokis, M.; Noris, K.; Bubis, J.; Guerra, M.; Castrillo, M. Effects of cadmium in plants of Sphagneticola trilobata (L.) Pruski. Bioagro 2019, 31, 133–142. [Google Scholar]
- Liu, S.; Li, Y.; Liu, L.; Min, J.; Liu, W.; Li, X.; Pan, X.; Lu, X.; Deng, Q. Comparative proteomics in rice seedlings to characterize the resistance to cadmium stress by high-performance liquid chromatography–tandem mass spectrometry (HPLC-MS/MS) with isobaric tag for relative and absolute quantitation (iTRAQ). Anal. Lett. 2020, 53, 807–820. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, L.; Shen, H.; Wang, J.; Liu, W.; Zhu, X.; Wang, R.; Sun, X.; Liu, L. Metabolomic analysis with GC-MS to reveal potential metabolites and biological pathways involved in Pb and Cd stress response of radish roots. Sci. Rep. 2015, 5, 18296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razzaq, A.; Sadia, B.; Raza, A.; Khalid Hameed, M.; Saleem, F. Metabolomics: A way forward for crop improvement. Metabolites 2019, 9, 303. [Google Scholar] [CrossRef] [Green Version]
- Siripornadulsil, S.; Traina, S.; Verma, D.P.S.; Sayre, R.T. Molecular mechanisms of proline-mediated tolerance to toxic heavy metals in transgenic microalgae. Plant Cell 2002, 14, 2837–2847. [Google Scholar] [CrossRef]
- Collin, V.C.; Eymery, F.; Genty, B.; Rey, P.; Havaux, M. Vitamin E is essential for the tolerance of Arabidopsis thaliana to metal-induced oxidative stress. Plant Cell Environ. 2008, 31, 244–257. [Google Scholar] [CrossRef]
- Xie, M.; Chen, W.; Lai, X.; Dai, H.; Sun, H.; Zhou, X.; Chen, T. Metabolic responses and their correlations with phytochelatins in Amaranthus hypochondriacus under cadmium stress. Environ. Pollut. 2019, 252, 1791–1800. [Google Scholar] [CrossRef]
- Navarro-Reig, M.; Jaumot, J.; Piña, B.; Moyano, E.; Galceran, M.T.; Tauler, R. Metabolomic analysis of the effects of cadmium and copper treatment in Oryza sativa L. using untargeted liquid chromatography coupled to high resolution mass spectrometry and all-ion fragmentation. Metallomics 2017, 9, 660–675. [Google Scholar] [CrossRef]
- Mengdi, X.; Haibo, D.; Jiaxin, L.; Zhe, X.; Yi, C.; Xuan, L.; Haiyan, M.; Hui, S.; Tianqi, A.; Yunzhen, L. Metabolomics reveals the “Invisible” detoxification mechanisms of Amaranthus hypochondriacus at three ages upon exposure to different levels of cadmium. Ecotoxicol. Environ. Saf. 2020, 195, 110520. [Google Scholar] [CrossRef]
- Hediji, H.; Djebali, W.; Cabasson, C.; Maucourt, M.; Baldet, P.; Bertrand, A.; Zoghlami, L.B.; Deborde, C.; Moing, A.; Brouquisse, R. Effects of long-term cadmium exposure on growth and metabolomic profile of tomato plants. Ecotoxicol. Environ. Saf. 2010, 73, 1965–1974. [Google Scholar] [CrossRef] [PubMed]
- Cosio, C.; Renault, D. Effects of cadmium, inorganic mercury and methylmercury on the physiology and metabolomic profiles of shoots of the macrophyte Elodea nuttallii. Environ. Pollut. 2020, 257, 113557. [Google Scholar] [CrossRef]
- Mwamba, T.; Islam, F.; Ali, B.; Lwalaba, J.; Gill, R.; Zhang, F.; Farooq, M.; Ali, S.; Ulhassan, Z.; Huang, Q. Comparative metabolomic responses of low-and high-cadmium accumulating genotypes reveal the cadmium adaptive mechanism in Brassica napus. Chemosphere 2020, 250, 126308. [Google Scholar] [CrossRef]
- He, X.L.; Fan, S.K.; Zhu, J.; Guan, M.Y.; Liu, X.X.; Zhang, Y.S.; Jin, C.W. Iron supply prevents Cd uptake in Arabidopsis by inhibiting IRT1 expression and favoring competition between Fe and Cd uptake. Plant Soil 2017, 416, 453–462. [Google Scholar] [CrossRef]
- Zhang, Z.; Luo, J.-S.; Xiao, Y.; Yao, J.; Wu, Z.; Yang, Y.; Ismail, A.M. Overexpression of a defensin-like gene CAL2 enhances cadmium accumulation in plants. Front. Plant Sci. 2020, 11, 217. [Google Scholar]
- Luo, J.-S.; Gu, T.; Yang, Y.; Zhang, Z. A non-secreted plant defensin AtPDF2. 6 conferred cadmium tolerance via its chelation in Arabidopsis. Plant Mol. Biol. 2019, 100, 561–569. [Google Scholar] [CrossRef]
- Reisinger, S.; Schiavon, M.; Terry, N.; Pilon-Smits, E.A. Heavy metal tolerance and accumulation in Indian mustard (Brassica juncea L.) expressing bacterial γ-glutamylcysteine synthetase or glutathione synthetase. Int. J. Phytoremediat. 2008, 10, 440–454. [Google Scholar] [CrossRef] [PubMed]
- Luo, B.F.; Du, S.T.; Lu, K.X.; Liu, W.J.; Lin, X.Y.; Jin, C.W. Iron uptake system mediates nitrate-facilitated cadmium accumulation in tomato (Solanum lycopersicum) plants. J. Exp. Bot. 2012, 63, 3127–3136. [Google Scholar] [CrossRef]
- Mao, Q.Q.; Guan, M.Y.; Lu, K.X.; Du, S.T.; Fan, S.K.; Ye, Y.-Q.; Lin, X.Y.; Jin, C.W. Inhibition of nitrate transporter 1.1-controlled nitrate uptake reduces cadmium uptake in Arabidopsis. Plant Physiol. 2014, 166, 934–944. [Google Scholar] [CrossRef]
- Siemianowski, O.; Barabasz, A.; Kendziorek, M.; Ruszczyńska, A.; Bulska, E.; Williams, L.E.; Antosiewicz, D.M. HMA4 expression in tobacco reduces Cd accumulation due to the induction of the apoplastic barrier. J. Exp. Bot. 2014, 65, 1125–1139. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Wang, P.; Wang, P.; Yang, M.; Lian, X.; Tang, Z.; Huang, C.F.; Salt, D.E.; Zhao, F.J. A loss-of-function allele of OsHMA3 associated with high cadmium accumulation in shoots and grain of Japonica rice cultivars. Plant Cell Environ. 2016, 39, 1941–1954. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Feng, S.J.; Chen, J.; Zhao, W.T.; Yang, Z.M. A cadmium stress-responsive gene AtFC1 confers plant tolerance to cadmium toxicity. BMC Plant Biol. 2017, 17, 187. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, W.; Chao, Y.; Wang, S.; Tang, Y.-T.; Qiu, R.-L. Metal-tolerant Enterobacter sp. strain EG16 enhanced phytoremediation using Hibiscus cannabinus via siderophore-mediated plant growth promotion under metal contamination. Plant Soil 2017, 413, 203–216. [Google Scholar] [CrossRef]
- Yang, P.; Zhou, X.-F.; Wang, L.-L.; Li, Q.-S.; Zhou, T.; Chen, Y.-K.; Zhao, Z.-Y.; He, B.-Y. Effect of phosphate-solubilizing bacteria on the mobility of insoluble cadmium and metabolic analysis. Int. J. Environ. Res. Public Health 2018, 15, 1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajkumar, M.; Sandhya, S.; Prasad, M.; Freitas, H. Perspectives of plant-associated microbes in heavy metal phytoremediation. Biotechnol. Adv. 2012, 30, 1562–1574. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Mushtaq, H.; Ali, H.; Munis, M.F.H.; Javed, M.T.; Chaudhary, H.J. Diazotrophs-assisted phytoremediation of heavy metals: A novel approach. Environ. Sci. Pollut. Res. 2015, 22, 2505–2514. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, B.; Ohtomo, R. Mycorrhizal effects on growth, P uptake and Cd tolerance of the host plant vary among different AM fungal species. Soil Sci. Plant Nutr. 2015, 61, 359–368. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Wang, L.; Ma, F.; You, Y.; Wang, Y.; Yang, D. Integration of earthworms and arbuscular mycorrhizal fungi into phytoremediation of cadmium-contaminated soil by Solanum nigrum L. J. Hazard. Mater. 2020, 389. [Google Scholar] [CrossRef]
- Gunathilakae, N.; Yapa, N.; Hettiarachchi, R. Effect of arbuscular mycorrhizal fungi on the cadmium phytoremediation potential of Eichhornia crassipes (Mart.) Solms. Groundwater Sustain. Dev. 2018, 7, 477–482. [Google Scholar] [CrossRef]
- Hashem, A.; Abd_Allah, E.; Alqarawi, A.; Egamberdieva, D. Bioremediation of adverse impact of cadmium toxicity on Cassia italica Mill by arbuscular mycorrhizal fungi. Saudi J. Biol. Sci. 2016, 23, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Hashem, A.; Abd_Allah, E.; Alqarawi, A.; Al Huqail, A.A.; Egamberdieva, D.; Wirth, S. Alleviation of cadmium stress in Solanum lycopersicum L. by arbuscular mycorrhizal fungi via induction of acquired systemic tolerance. Saudi J. Biol. Sci. 2016, 23, 272–281. [Google Scholar] [CrossRef] [Green Version]
- Bissonnette, L.; St-Arnaud, M.; Labrecque, M. Phytoextraction of heavy metals by two Salicaceae clones in symbiosis with arbuscular mycorrhizal fungi during the second year of a field trial. Plant Soil 2010, 332, 55–67. [Google Scholar] [CrossRef]
- Gao, Y.; Miao, C.; Mao, L.; Zhou, P.; Jin, Z.; Shi, W. Improvement of phytoextraction and antioxidative defense in Solanum nigrum L. under cadmium stress by application of cadmium-resistant strain and citric acid. J. Hazard. Mater. 2010, 181, 771–777. [Google Scholar] [CrossRef]
- Ling-Zhi, L.; Zong-Qiang, G.; ZHANG, Y.-L.; Pei-Jun, L. Growth, cadmium accumulation and physiology of marigold (Tagetes erecta L.) as affected by arbuscular mycorrhizal fungi. Pedosphere 2011, 21, 319–327. [Google Scholar]
- Bhaduri, A.M.; Fulekar, M. Assessment of arbuscular mycorrhizal fungi on the phytoremediation potential of Ipomoea aquatica on cadmium uptake. 3 Biotech 2012, 2, 193–198. [Google Scholar] [CrossRef] [Green Version]
- Garg, N.; Chandel, S. Role of arbuscular mycorrhizal (AM) fungi on growth, cadmium uptake, osmolyte, and phytochelatin synthesis in Cajanus cajan (L.) Millsp. under NaCl and Cd stresses. J. Plant Growth Regul. 2012, 31, 292–308. [Google Scholar] [CrossRef]
- Hassan, S.E.; Hijri, M.; St-Arnaud, M. Effect of arbuscular mycorrhizal fungi on trace metal uptake by sunflower plants grown on cadmium contaminated soil. New Biotechnol. 2013, 30, 780–787. [Google Scholar] [CrossRef]
- Liu, L.; Gong, Z.; Zhang, Y.; Li, P. Growth, cadmium uptake and accumulation of maize (Zea mays L.) under the effects of arbuscular mycorrhizal fungi. Ecotoxicology 2014, 23, 1979–1986. [Google Scholar] [CrossRef]
- Ali, N.; Masood, S.; Mukhtar, T.; Kamran, M.A.; Rafique, M.; Munis, M.F.H.; Chaudhary, H.J. Differential effects of cadmium and chromium on growth, photosynthetic activity, and metal uptake of Linum usitatissimum in association with Glomus intraradices. Environ. Monit. Assess. 2015, 187, 311. [Google Scholar]
- Jiang, Q.-Y.; Zhuo, F.; Long, S.-H.; Zhao, H.-D.; Yang, D.-J.; Ye, Z.-H.; Li, S.-S.; Jing, Y.-X. Can arbuscular mycorrhizal fungi reduce Cd uptake and alleviate Cd toxicity of Lonicera japonica grown in Cd-added soils? Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Jin, Z.; Deng, S.; Wen, Y.; Jin, Y.; Pan, L.; Zhang, Y.; Black, T.; Jones, K.C.; Zhang, H.; Zhang, D. Application of Simplicillium chinense for Cd and Pb biosorption and enhancing heavy metal phytoremediation of soils. Sci. Total Environ. 2019, 697, 134148. [Google Scholar] [CrossRef]
- Ullah, A.; Heng, S.; Munis, M.F.H.; Fahad, S.; Yang, X. Phytoremediation of heavy metals assisted by plant growth promoting (PGP) bacteria: A review. Environ. Exp. Bot. 2015, 117, 28–40. [Google Scholar] [CrossRef]
- Prapagdee, B.; Khonsue, N. Bacterial-assisted cadmium phytoremediation by Ocimum gratissimum L. in polluted agricultural soil: A field trial experiment. Int. J. Environ. Sci. Technol. 2015, 12, 3843–3852. [Google Scholar] [CrossRef] [Green Version]
- Rojjanateeranaj, P.; Sangthong, C.; Prapagdee, B. Enhanced cadmium phytoremediation of Glycine max L. through bioaugmentation of cadmium-resistant bacteria assisted by biostimulation. Chemosphere 2017, 185, 764–771. [Google Scholar] [CrossRef]
- Guo, J.; Chi, J. Effect of Cd-tolerant plant growth-promoting rhizobium on plant growth and Cd uptake by Lolium multiflorum Lam. and Glycine max (L.) Merr. in Cd-contaminated soil. Plant Soil 2014, 375, 205–214. [Google Scholar] [CrossRef]
- Sriprang, R.; Hayashi, M.; Ono, H.; Takagi, M.; Hirata, K.; Murooka, Y. Enhanced accumulation of Cd2+ by a Mesorhizobium sp. transformed with a gene from Arabidopsis thaliana coding for phytochelatin synthase. Appl. Environ. Microbiol. 2003, 69, 1791–1796. [Google Scholar] [CrossRef] [Green Version]
- Pan, F.; Meng, Q.; Luo, S.; Shen, J.; Chen, B.; Khan, K.Y.; Japenga, J.; Ma, X.; Yang, X.; Feng, Y. Enhanced Cd extraction of oilseed rape (Brassica napus) by plant growth-promoting bacteria isolated from Cd hyperaccumulator Sedum alfredii Hance. Int. J. Phytoremediat. 2017, 19, 281–289. [Google Scholar] [CrossRef]
- Liu, W.; Wang, Q.; Wang, B.; Hou, J.; Luo, Y.; Tang, C.; Franks, A.E. Plant growth-promoting rhizobacteria enhance the growth and Cd uptake of Sedum plumbizincicola in a Cd-contaminated soil. J. Soil Sediments 2015, 15, 1191–1199. [Google Scholar] [CrossRef]
- Wan, Y.; Luo, S.; Chen, J.; Xiao, X.; Chen, L.; Zeng, G.; Liu, C.; He, Y. Effect of endophyte-infection on growth parameters and Cd-induced phytotoxicity of Cd-hyperaccumulator Solanum nigrum L. Chemosphere 2012, 89, 743–750. [Google Scholar] [CrossRef]
- Chen, Z.-j.; Sheng, X.-f.; He, L.-y.; Huang, Z.; Zhang, W.-h. Effects of root inoculation with bacteria on the growth, Cd uptake and bacterial communities associated with rape grown in Cd-contaminated soil. J. Hazard. Mater. 2013, 244, 709–717. [Google Scholar] [CrossRef]
- Chen, L.; Luo, S.; Li, X.; Wan, Y.; Chen, J.; Liu, C. Interaction of Cd-hyperaccumulator Solanum nigrum L. and functional endophyte Pseudomonas sp. Lk9 on soil heavy metals uptake. Soil Biol. Biochem. 2014, 68, 300–308. [Google Scholar] [CrossRef]
- Kamran, M.A.; Syed, J.H.; Eqani, S.A.M.A.S.; Munis, M.F.H.; Chaudhary, H.J. Effect of plant growth-promoting rhizobacteria inoculation on cadmium (Cd) uptake by Eruca sativa. Environ. Sci. Pollut. Res. 2015, 22, 9275–9283. [Google Scholar] [CrossRef]
- Złoch, M.; Kowalkowski, T.; Tyburski, J.; Hrynkiewicz, K. Modeling of phytoextraction efficiency of microbially stimulated Salix dasyclados L. in the soils with different speciation of heavy metals. Int. J. Phytoremediat. 2017, 19, 1150–1164. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Jiao, J.; Fan, X.; Sun, H.; Zhang, Y.; Jiang, J.; Liu, C. Endophytic bacterium Pseudomonas fluorescens RG11 may transform tryptophan to melatonin and promote endogenous melatonin levels in the roots of four grape cultivars. Front. Plant Sci. 2017, 7, 2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nayak, A.; Panda, S.; Basu, A.; Dhal, N. Enhancement of toxic Cr (VI), Fe, and other heavy metals phytoremediation by the synergistic combination of native Bacillus cereus strain and Vetiveria zizanioides L. Int. J. Phytoremediat. 2018, 20, 682–691. [Google Scholar] [CrossRef] [PubMed]







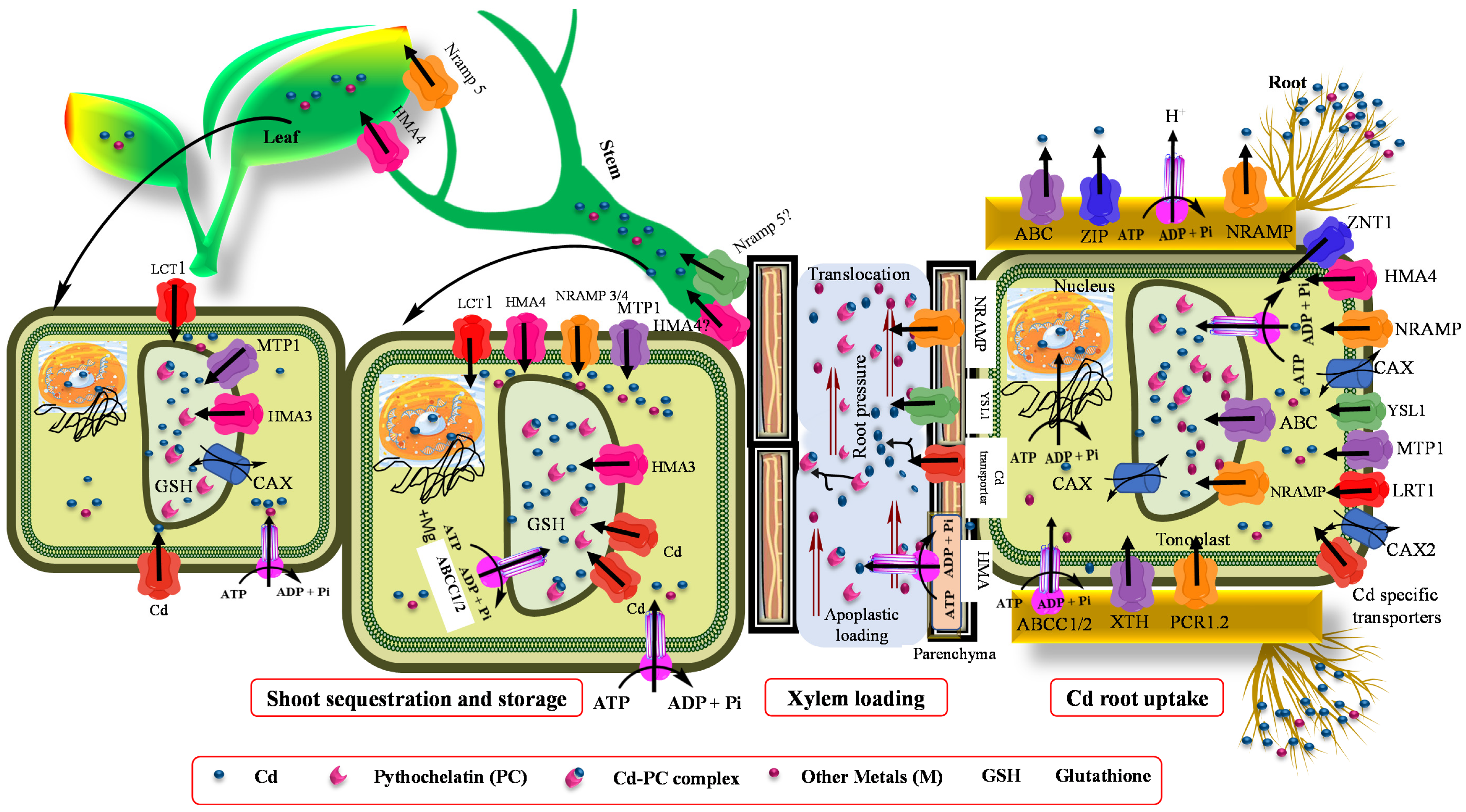{kind=link}
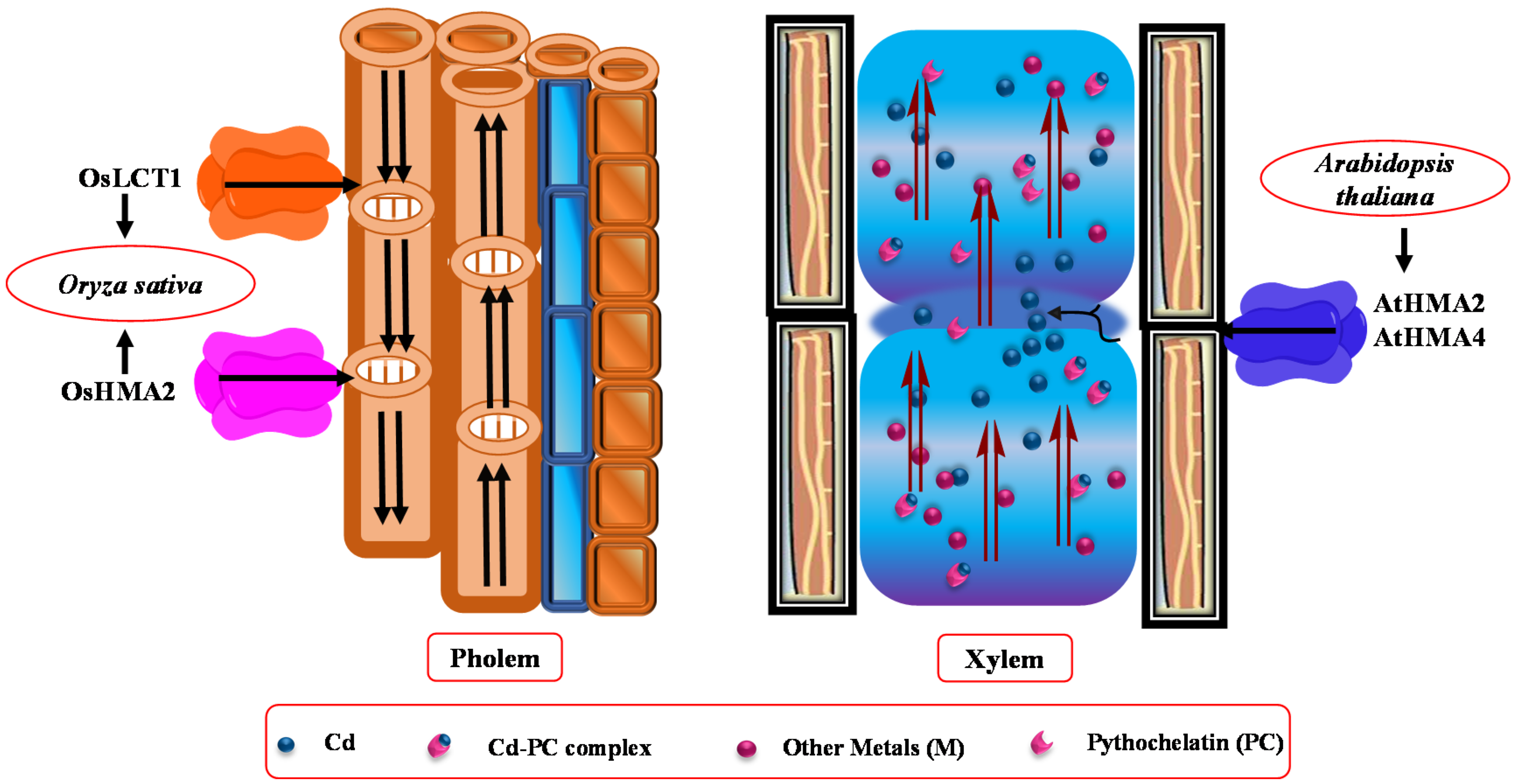{kind=link}
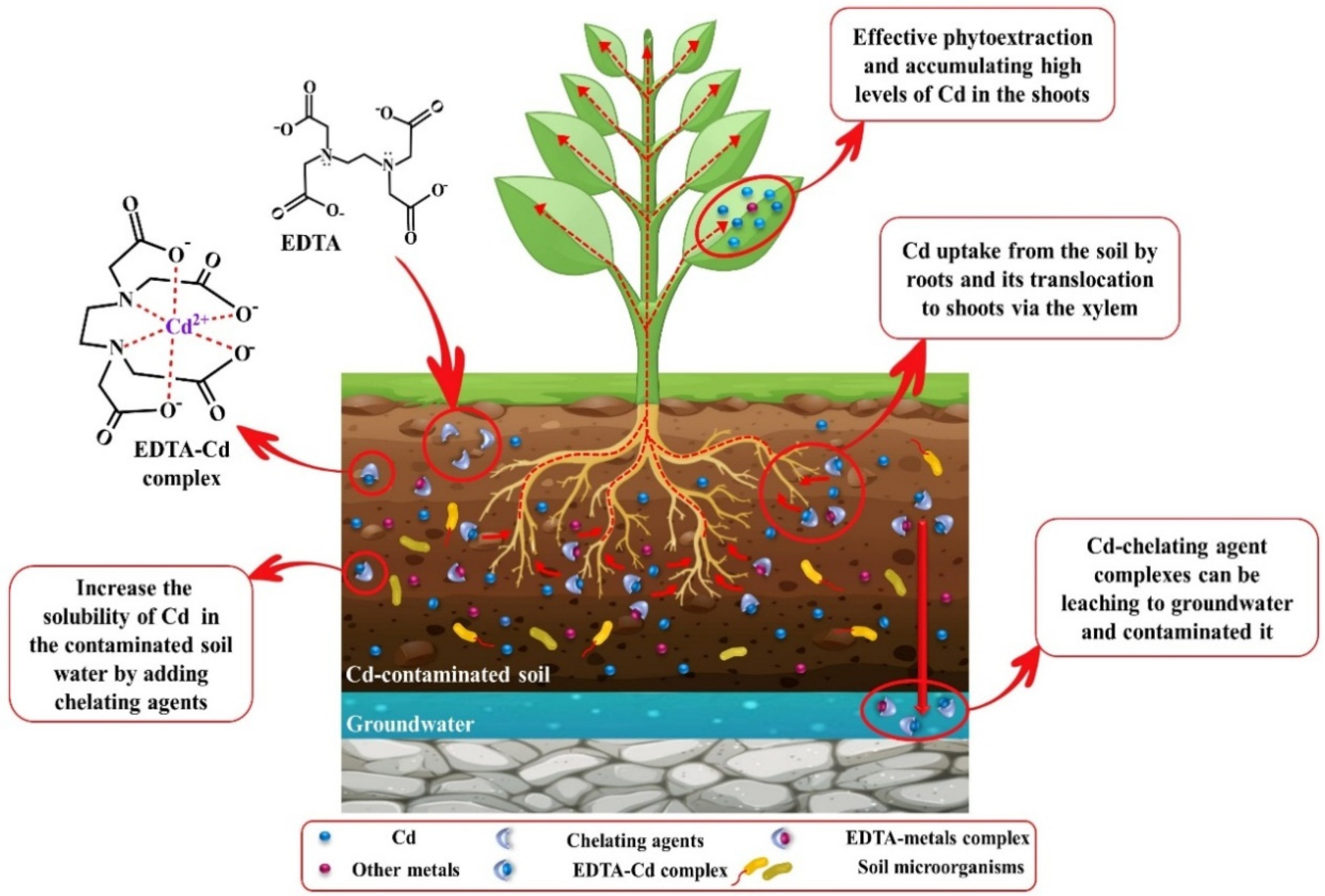{kind=link}
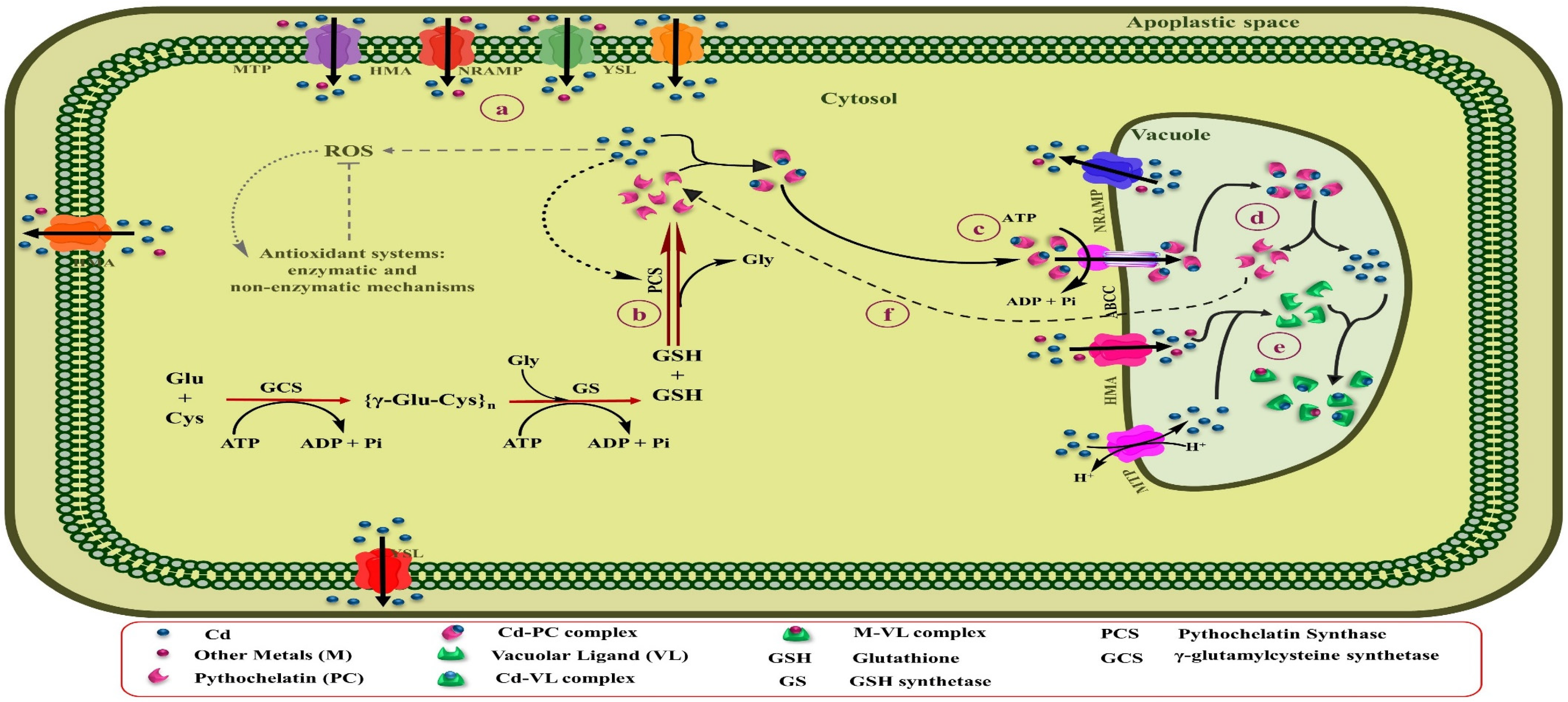{kind=link}
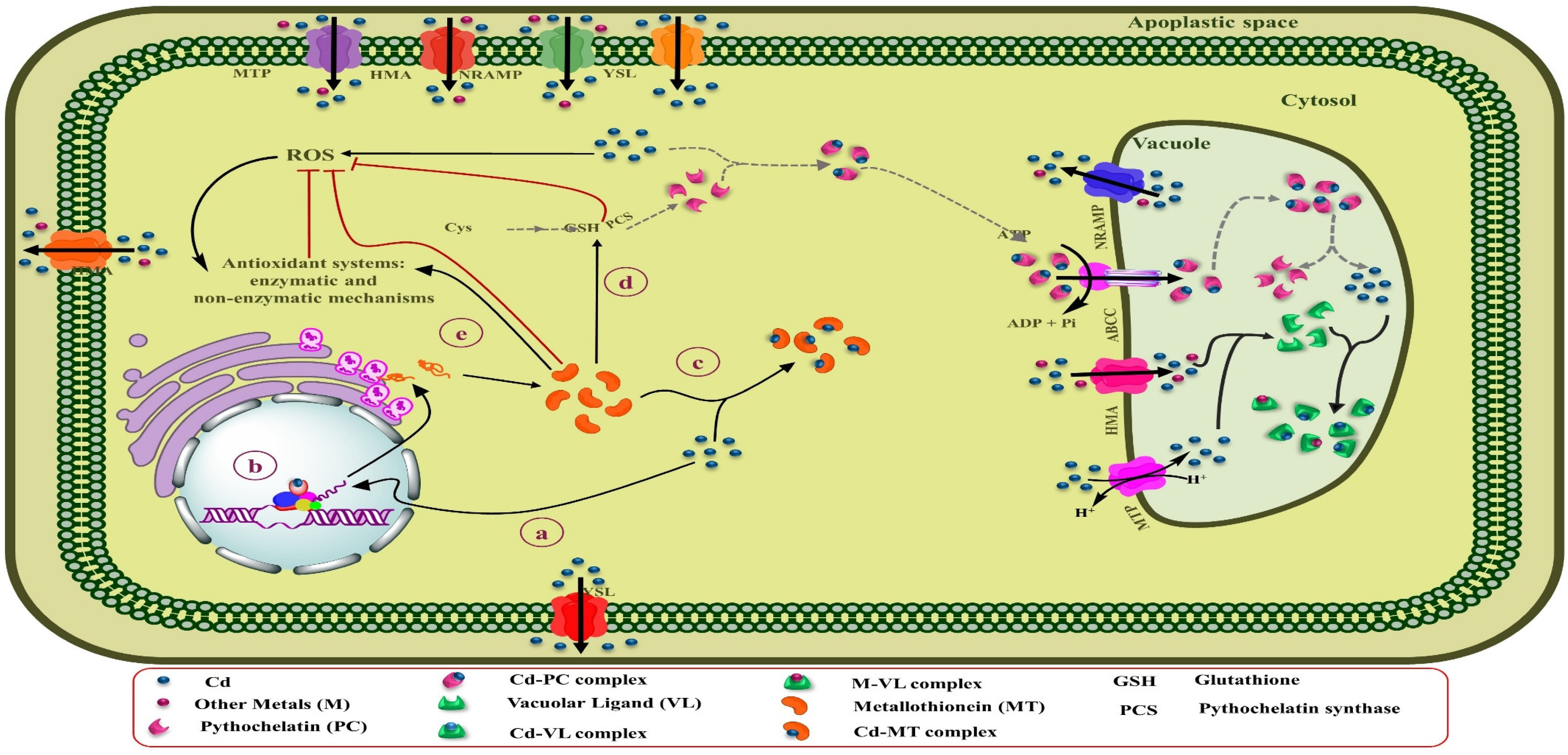{kind=link}
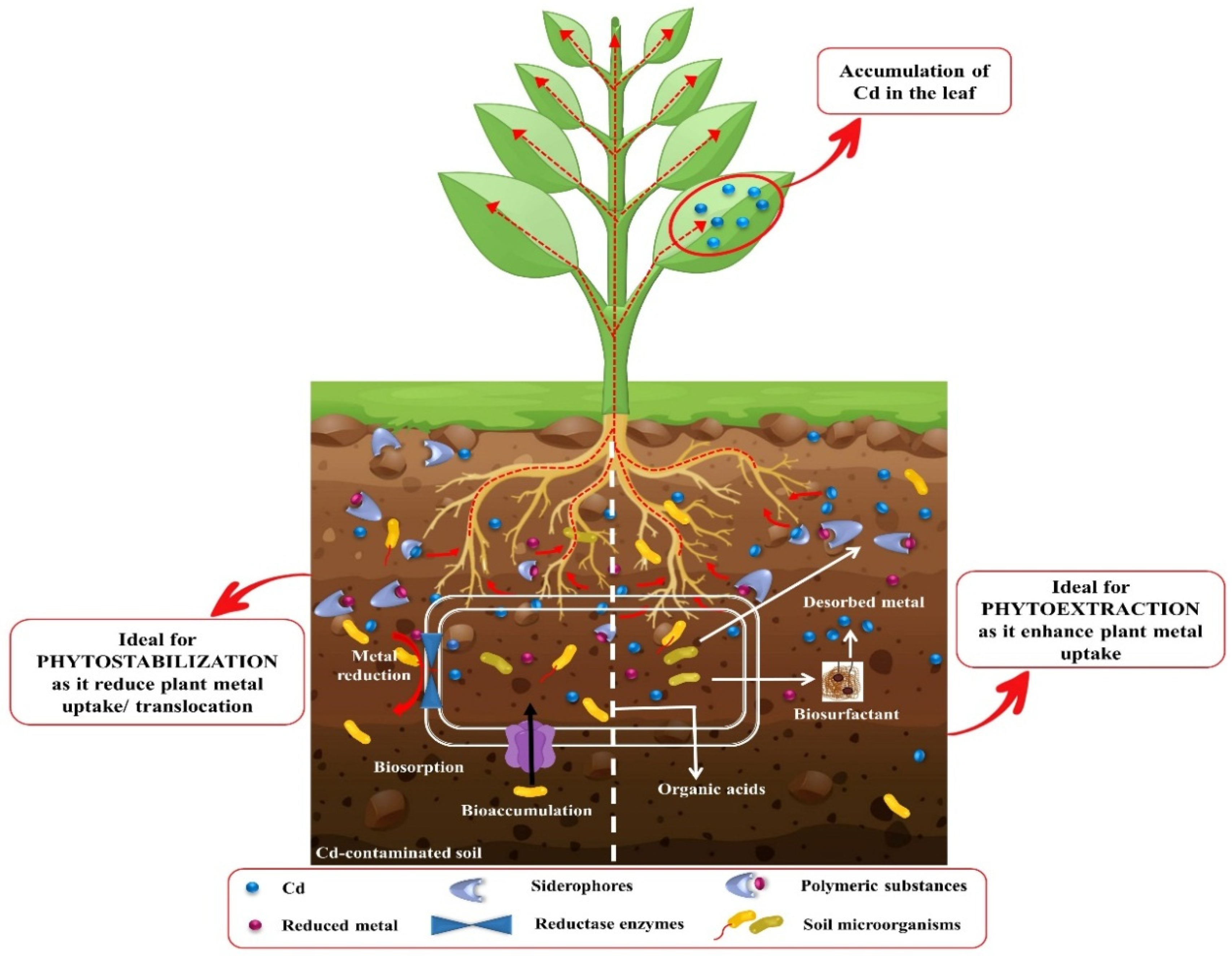{kind=link}
| Types | Process | Mechanism | Plants | References |
|---|---|---|---|---|
| Phytoextraction/ Phytoaccumulation | Bioaugmentation-assisted phytoextraction | Combined with mycorrhiza | Suaeda salsa and Trichoderma asperellum | [102] |
| Chelated-assisted phytoextraction | Chelates like EDTA, SDS, and EGTA | Fagopyrum esculentum | [103] | |
| Phytomining | Phytoextraction for commercial use, like silver (Ag), Ni | Alyssum murale, Odontarrhena chalcidica | [104] | |
| Phyostabilization | Organic fertilizers, biochar | Immobilization of Cd by using biomolecules | Virola surinamensis, Boehmeria nivea | [86,91] |
| Phytofiltration | Biosorption | Metals are absorbed bound in cells, used for phytoremediation | Lythrum salicaria | [105] |
| Rhizofiltration | Metals are absorbed and bound on only roots | Micranthemum umbrosum | [95] | |
| Blastofiltration | Metals are absorbed and bound on only seedlings | Moringa Oliefera, Cucumis melo, Abelmoschus esculentus, Ricinus communis | [106,107] | |
| Caulofiltration | Metals are bond and absorbed on excised plant | Berkheya coddii | [108] | |
| Phytostimulation | Fungi, bacteria | Phytoremediation with the intervention of microorganisms in different terms to remediate soil with organic pollutants | Rumex K-1 (Rumex patientia × R. timschmicus) Viola baoshanesis. Vertiveria zizanioides | [109,110] |
| Transporter | Localization | Function/Substrate | References |
|---|---|---|---|
| Plasma Membrane | |||
| OsNRAMP1 | Roots and shoots | Influx of Cd, Al | [123] |
| AtGLRs | Roots | Cations transporter | [130] |
| OsNRAMP5 | Roots | Uptake of Mn and Cd | [126] |
| AtCNGCs | Roots | Cations transporter | [130] |
| OsLCT1 | Leaves, nodes, phloem parenchyma | Efflux of Cd, Ca, Mg, and Mn | [142] |
| CsMTP9 | Roots endodermal cells | Efflux of Cd and Mn | [134] |
| HvNRAMP5 | Roots | Uptake of Mn and Cd | [129] |
| SnYSL3 | Vascular and epidermal cells of the roots and stems | Transport of nicotianamine complexes containing Fe(II), Cu, Zn, and Cd | [165] |
| OsYSL2 | Vascular bundles, roots | Influx of Mn and Cd | [166] |
| AtNRAMP6 | Young leaves, roots | Mn and Cd influx | [167] |
| OsZIP8 | Roots | Influx of Zn, Cd | [168] |
| OsIRT1/2 | Roots | Uptake of Cd | [169] |
| Tonoplast | |||
| AtABCC1/2 | Roots and shoots | PCs uptake | [170] |
| AtNRAMP4 | Leaves, roots, vascular bundles | Cd and Fe efflux | [171] |
| TgMTP1 | Roots and leaves | Transporter for Zn and Cd | [172] |
| AtMTP1 | Leaves and roots | Zn and Cd transporters | [173] |
| OsNRAMP3 | Vascular bundles, roots, leaves | Uptake of Mn | [136] |
| OsHMA5 | Roots, vascular bundles | Loading of Cu in xylem | [174] |
| OsMTP8.1 | Roots | Sequestration of Mn into vacuoles | [175] |
| AhCAX1 | Shoots | Uptake of Cd | [3] |
| AtABCC3 | Roots and shoots | PCs uptake | [145] |
| SpHMA3 | Roots, shoots | Sequestration of Cd into vacuoles | [152] |
| AtCAX2/4 | Roots | Cd vacuolar transport | [176] |
| OsHMA3 | Roots | Cd sequestration in root vacuoles | [177] |
| Others | |||
| AtNRAMP3 | Mesophyll vacuoles | Cd, Fe, Mn efflux | [171] |
| NcHMA3 | Root pericycle | Cd vascular sequestration, high substrate specification | [150] |
| GmHMA3 | Roots | Divalent transport | [178] |
| SaCAX2 | Leaves | Cd uptake | [144] |
| Plant Specie | Cd Level and Duration | Antioxidant Response to Cd Stress | References |
|---|---|---|---|
| Lolium perenne | 0.2, 0.5 mM Cd; 7 d | ↑POD, CAT, SOD, APX, GPX and GR | [193] |
| Brassica juncea | 25, 50, 100, 150 mg kg−1 CdCl2 soil; 30 days after sowing (DAS) | ↑CAT, APX and GR | [194] |
| Brassica napus | 0.03, 0.3 and 1 mg CdCl2; After three weeks, samples were harvested | ↑CAT, APX, GR, GSH and AsA | [195] |
| Triticum aestivum | 10, 20 kg ha−1 P with 100 µM Cd; 28 d | ↑SOD, POD, CAT, AsA and α-tocopherol levels | [185] |
| Macleaya cordata | 10–60 mg kg−1 Cd soil + 30–180 mg artificially added Cd; 60 d | ↑SOD and MDA ↓CAT activity | [184] |
| Atriplex halimus | 50, 200, 400, 650 and 1350 µM CdCl2; 22 d | ↑SOD, CAT, guaiacol peroxidas | [196] |
| Paspalum scrobiculatu; T. aestivum | 1000 µM CdCl2; 14 d | ↑SOD and POD | [197] |
| Solanum tuberosum | 18 different Cd levels and best selected was 218 µM CdCl2; 20 d | Differential activities of CAT, SOD, GPX and APX ↓Lipid peroxidation, H2O2 | [198] |
| Glycine max | 50, 100, 150 mg L−1; 35 d | ↑GR, CAT, SOD | [189] |
| Petroselinum hortense | 75, 150, 300 µM CdCl2; 15 d | ↑POD increased ↓SOD, CAT and APX activities | [44] |
| Solanum lycopersicum | 35 μM CdCl2; 1, 4 and 7 d | ↑APX, GR and GST ↓GSH content | [4] |
| Abelmoschus manihot | 5, 15, 30, 60 and 100 mg kg−1 CdCl2; 60 d | ↑SOD, CAT, POD | [52] |
| Brassica chinensis | 20 μM Cd; 7 d | ↑SOD, CAT, POD, AsA and GSH | [199] |
| Quercus robur | 10, 50 µM CdCl2; 1 h,3 h, 24 h, 3 and 7 d | ↑APX, SOD and CAT activity | [200] |
| Sedum alfredii | 100 μM CdSO4; 6, 24, 36, 48, 72 h | ↑SOD, GR, GSNOR and γ-ECS ↓POD, CAT and APX | [201] |
| Oryza sativa | 80 μM Cd2SO4 with 1, 3 or 6 mM Glu; 1 or 2 weeks | ↑CAT, POD and GST | [169] |
| Plant Species | Cd Level and Duration | Specific Tissue | Approach | Functional Annotation Methods | Differentially Expressed Genes (DEGs) | Key Findings | References |
|---|---|---|---|---|---|---|---|
| Brassica parachinensis | 50 µM CdCl2; 3 d | Roots | RNA-seq | GO, KOBAS | 3073 DEGs | 10 genes selected for qRT-PCR Cd tolerant genes HMA3, HMA4 and Nramp1 identified. Novel pathways viral carcinogenesis and Parkinson’s disease might be potential in Cd tolerance | [259] |
| Arachis hypogaea | 2 µM CdCl2; 7 d | Roots | RNA-seq | GO, KEGG | 2793 DEGs; 331 genes in Fenghua 1; 1302 genes in Silihong | 16 DEGs expression validated by qRT-PCR. 9 genes related to metal transport identified (PDR1, IRT1, ZIP1, ZIP11, ABCC4, ABCC15, YSL7, MTP4 and DTX43) in low Cd cultivar might be linked to Cd accumulation and transport | [260] |
| Agrostis stolonifera | 2.5 mM (BT2_5); 43mM (BT43) CdCl2; 7 d | Leaves | RNA-seq | NR, NT, Pfam, GO, KEGG, KOG | 279 DEG (BT2_5); 2509 DEGs (BT43) | 9 DEGs selected for qRT-PCR. WRKY, ERF, bZIP and MYB transcription factors linked with Cd stress response | [261] |
| Landoltia punctata | 20 µM CdCl2; 0, 1, 3, 6, 9 d | Plant | RNA-seq | Pfam, Nr NCBI, COG, Swis-Prot, KEGG, GO | 9461 DEGs in CT1; 9847 DEGs in CT3 and 9615 DEGs in CT6. | 15 DEGs selected and validated by qRT-PCR. Sulfur and ROS metabolism genes upregulated along with DNA repair, RNA and protein metabolism and carbohydrate metabolic flux | [264] |
| Lolium multiflorum Lam. | 50 µmol L-1 CdCl2; 6 h | Roots | PacBio ISO-seq + RNA-seq | Nr NCBI, Swiss-Prot, GO, KEGG | 2367 DEGs | 20 DEGs for qRT-PCR construction of UniTransModel. LmAUX1 gene overexpression enhances Cd phytoremediation in Arabidopsis | [262] |
| Siegesbeckia orientalis | 50 µM CdCl2; 1, 3, 5 d | Roots | RNA-seq | Nr NCBI, COG, pfam, Interpro, Swiss-Prot, GO, KEGG | 5896 DEGs C1T1; 3036 C3T3 5149 C5T5 | 6 genes selected for qRT-PCR. DEGs majorly evolve in Cd absorption and transport, ABC signaling, phenylpropanoid biosynthesis and ROS scavenging pathways | [265] |
| Oryza sativa | 15 mg kg−1 CdCl2; 4 months | Panicle nodes, Node1, Stem node (fresh, leafless stem, snap) | RNA-seq, miRNA | miRNA analysis (Rfam, cDNA seq, miRBase), GO, KEGG | 4533 DEGs, 70 miRNA | 14mRNA, 3miRNA selected for qRT-PCR. 5 genes OsIRT1, OsVIT2, OsABCC1, OSNRT1.5A, OSNramps5 and Panicle node and node1 are involved in blocking Cd upward transport | [266] |
| Plant Species | Cd Level and Duration | Specific Tissue | Extraction Protocol | Analytical Approach | Key Findings of Differentially Expressed Proteins (DEPs) | References |
|---|---|---|---|---|---|---|
| Arabidopsis thaliana | 100 mM L−1 CdCl2; 30 d | Leaves | Phenol extraction | LC-MS/MS | 56 DEPs identified Protein linked to disease resistance, defense, energy and metabolism Increase in activity of APX, CAT and GR, MDA activities | [272] |
| Oryza sativa | 10,100 µM CdCl2; 12 d | Leaves | Phenol extraction | LC-MS/MS, iTRAQ, qRT-PCR | 482 DEPs identified Phosphorylated proteins involved in stress tolerance, transcription factor, signaling and neutralizing reactive oxygen specie | [273] |
| Cucumis sativus | 100 µM CdCl2; 10 d | Leaves | TCA/Acetone | iTRAQ, LC-ESI-MS/MS | 1106 DEPs identified in three contrasting groups (377, 297 and 432). Nitic oxide treatment reversed the Cd disruptive pathways as redox homeostasis, photosynthesis, glutathione mediated Cd detoxification, chlorophyll metabolism, Cd transport and localization and Ca2+ signaling transduction | [274] |
| Microsorum pteropus | 100, 250, 500 µM CdCl2; 7 d | Leaves, Roots | TCA/Acetone | 2DE, MALDI-TOF/TOF-MS | 8 DEPs identified in roots with enhanced antioxidants and energy metabolism activities 20 DEPs identified in leaves with enhanced photosynthetic functions and regulate cellular metabolism | [271] |
| Solanum lycopersicum | 35 µM CdCl2; 4 d | Roots, Shoots | Phenol extraction | LC-MS/MS | 4051 non-redundant proteins. 358 DEPs in sensitive genotype. 84 DEPs in the tolerant genotype Proteins identified related to peroxidase, GST, cytoskeleton, S-adenosylmethionine (SAM) biosynthesis, glycolosis, chaperone and oxidoreductase. | [275] |
| Sphagneticola trilobata | 0.5, 5, 25, 35, 50 µM CdCl2; 96 h | Roots, Leaves, Stem | DTPA/TFA soln. for thiol, TFA/acetonitrile for PCs | SDS-PAGE, HPLC | 1306 mg kg−1 Cd content increases in roots, 193 mg kg−1 in stem, 52 mg kg−1 in leaves Plant roots has increase content of L-cystein, glutathione, phytochelatin, and thiols | [276] |
| O. sativa | 100, 200 mM CdCl2; 48 h | Leaves | Phenol extraction | SDS-PAGE, iTRAQ labeling, RP-HPLC, LC-MS/MS | 109 DEPs identified in Bancanggengnuo (japonica), 209 DEPs identified in Yuzhenxiang (indica) Proteins identified linked to carbohydrate metabolism, redox reactions and signal transduction | [277] |
| Species | Stress Condition | Specific Tissue | Analytical Platform | Data Analysis | Key Findings | References |
|---|---|---|---|---|---|---|
| Solanum lycopersicum | 20, 100 μM CdCl2; 90 d | Leaves | NMR analysis, HPLC-PDA | PCA | 28 significantly different metabolites (SDMs) metabolites identified Cd stress cause decrease in ascorbate and proline content Cd stress cause increase in α-tocopherol, tyrosine and asparagine accumulation. | [285] |
| Raphanus sativus | 40 mg L−1 CdCl2; 72 h | Tap roots | GC-MS | PCA, PLS-DA, KEGG | 1,104 different types of metabolites identified Cd stress cause variations in amino acid metabolism, Energy production and Oxidative phosphorylation | [278] |
| Oryza sativa | 10, 50, 100, 1000 μM CdCl2; Upto harvest | Aerial parts, Roots | LC-MS | PLS-DA, MCR-ALS, KEGG | Secondary metabolism pathways and amino acid, carbon, glycerolipid and purine pathways affected 112 SDMs detected 97 SDMs was confirmed under Cd toxicity | [283] |
| Amaranthus hypochondriacus | 100 mg kg−1 CdCl2; 20 d | Leaves | LC-MS/MS HPLC | PCA, KEGG, HMDB, MetaboAnalyst | 41 SDMs involved in 7 metabolic pathways were detected 12 metabolites identified linked with phytochelatin in 3 pathways, i.e., Val, Leu and Ile biosynthesis, Ala, Asp and Glu metabolism, and Arg and Pro metabolism | [282] |
| Amaranthus hypochondriacus | 25, 200 mg kg−1 CdCl2; 20, 40, 60 d | Leaves | LC-MS | PCA, OPLS-DA, KEGG, HMDB, MetaboAnalyst | SDMs produced as: 31 at early vegetative stage, 29 at the intermediate stage, and 30 at late stages of plant growth Purine metabolism is the primary target of Cd absorption | [284] |
| Elodea nuttallii | 280 μg L−1 Cd as (Cd(NO3)2; 24 h | Cytocol, Shoots | GC-MS, LC analysis | PCA, MetaboAnalyst KEGG | Cd stress caused significant variations in aminoacyl-tRNA biosynthesis and branched-chain amino acids pathways In shoot, Cd induce concentration of 11 amino acids, 2 sugars, adonitol and pipecolic acid In cytosol, Cd induce concentration of glycine, ammonium, hydroxyproline and phosphoric acid | [286] |
| Brassica napus | 100 µM CdCl2; 8 d | Leaves | UPLC/MS | PCA, KEGG | SDMs found as: 644 in sensitive genotype ZD622, and 487 in tolerant genotype CB671 genotype (487) Most of the identified SDMs are involved in the Cd-mediated stress tolerance pathways | [287] |
| Plant Species | Genes | Phytoremediation Activity | References |
|---|---|---|---|
| Indian mustard | γ-glutamylcysteine synthetase (γ-ECS) | Higher phytochelatins production directly enhanced the Cd phytoremediation capacity of transgenic plants | [291] |
| Oryza sativa | OsHMA3 | Keep the Cd at roots via sequestrating into root vacuoles | [148] |
| Solanum lycopersicum | FER | FER lack of function mutant indirectly impaired the Cd translocation from root to aerial parts | [292] |
| Arabidopsis | NRT1.1 | Controlled the uptake of Cd in roots | [293] |
| Arabidopsis | AtHMA4 | Better root to shoot translocation of Cd | [294] |
| Arabidopsis | AtBCC3 | Facilitated the Cd phytoremediation in Arabidopsis by enhancing the chelation properties | [145] |
| Oryza sativa | OsHMA3 | Loss-of-function enhanced root-shoot Cd translocation | [295] |
| Arabidopsis | AtFC1 | Increased in the accumulation of Cd was observed | [296] |
| Arabidopsis | AtPDF2.5, AtPDF2.6 | Improved the Cd tolerance accumulation in shoot | [35,290] |
| Arabidopsis | CAL2 | Hastened the Cd accumulation in shoots | [289] |
| Arabidopsis | BTS | BTS lack of function mutant enhanced the Cd accumulative characteristics of Arabidopsis plants | [248] |
| Host Plants | AMF/Fungi Species | Specific Function | References |
|---|---|---|---|
| Salix viminalis | Glomus intraradices | Increased Cd phytoextraction and retained Cd in roots. root and restricted its translocation to shoots | [306] |
| Solanum nigrum | Paecilomyces lilacinus | Improved phytoextraction capacity along with enhanced antioxidant systems | [307] |
| Tagetes erecta | Glomus intraradices, Glomus constrictum and Glomus mosseae | Better phytoextraction of Cd from the soil and also restrict the translocation of Cd from roots to shoots | [308] |
| Lpomoea aquatica | Not specified | Increased the accumulation of Cd along with induced antioxidant enzymes activity and nutrient uptake | [309] |
| Cajanus cajan | Glomus mosseae | Amended the generation of phytochelatins and uptake of Cd by roots without accumulating a higher amount of toxic ions | [310] |
| Helianthus annuus | Rhizophagus irregularis and Funneliformis mosseae | Better phytoextraction and phytostabilization of Cd shown by the sunflower inoculated with AMF | [311] |
| Zea mays | Glomus intraradices, Glomus constrictum and Glomus mosseae | AMF inoculated plants rapped the Cd in roots and shown improved phytostabilization activity than the control plants | [312] |
| Linum usitatissimum | Glomus intraradices | Induced phytoextraction of Cd from the contaminated soil | [313] |
| Lonicera japonica | Glomus versiforme and Rhizophagus intraradices | AMF inoculated plants showed enhanced phytostabilization capacity of Cd | [314] |
| Eichhornia crassipes | Not specified | Enhanced the Cd accumulation capacity of water hyacinth in Cd contaminated soil and water | [303] |
| Solanum nigrum | Not specified | Boosted the phytoavailability of Cd by lowering the soil pH and altered Cd chemical fractions. | [302] |
| Phragmites communis | Simplicillium chinense | Enhanced the Cd phytoremediation by triggering the biosorption process in the host plant | [315] |
| Host Plants | PGRB Species | Specific Functions | References |
|---|---|---|---|
| Solanum nigrum | Serratia nematodiphila LRE07 | Serratia nematodiphila LRE07 inoculated plants induced the phytoaccumulation ability of the host plant resulted in to 70% more uptake of Cd | [323] |
| Sorghum bicolor | Bacillus sp. SLS18 | Augmented phytoextraction and phytostabilization of Cd at root was observed in the inoculated plants resulted in 65% more uptake of Cd from the contaminated soil | [323] |
| Brassica napus | Burkholderia sp. J62 and Pseudomonas thivervalensis Y-1-3-9 | The inoculated plants showed high level of Cd accumulated in their shots than the control plants | [324] |
| Solanum nigrum | Pseudomonas sp. LK9 | Substantial increase in the Cd uptake of upto 46% was recorded in the host plants inoculated with Pseudomonas sp. LK9 | [325] |
| Ocimum ratissimum | Arthrobacter sp. | Cd resistant bacteria along with host plant enhanced the phytoextraction process without altering the quality of grain | [317] |
| Eruca sativa | Pseudomonas putida ATCC 39,213 | Better phytoextraction resulted in increased Cd uptake (29%) in the inoculated plants than the control | [326] |
| Salix dasyclados | Streptomyces sp. | Generation of Siderophores molecules triggered the phytoextraction capacity of the host plant | [327] |
| Sedum plumbizincicolaa | Endophytic bacterium E6S | Production of IAA, ACC and organic acid maintained the pH at low level resulted in better phytoextraction of Cd | [328] |
| Vetiveria zizanioides | Bacillus cereus | Induced production of phosphate and Siderophores which helps in the phytoremediation of Cd | [329] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raza, A.; Habib, M.; Kakavand, S.N.; Zahid, Z.; Zahra, N.; Sharif, R.; Hasanuzzaman, M. Phytoremediation of Cadmium: Physiological, Biochemical, and Molecular Mechanisms. Biology 2020, 9, 177. https://doi.org/10.3390/biology9070177
Raza A, Habib M, Kakavand SN, Zahid Z, Zahra N, Sharif R, Hasanuzzaman M. Phytoremediation of Cadmium: Physiological, Biochemical, and Molecular Mechanisms. Biology. 2020; 9(7):177. https://doi.org/10.3390/biology9070177
Chicago/Turabian StyleRaza, Ali, Madiha Habib, Shiva Najafi Kakavand, Zainab Zahid, Noreen Zahra, Rahat Sharif, and Mirza Hasanuzzaman. 2020. "Phytoremediation of Cadmium: Physiological, Biochemical, and Molecular Mechanisms" Biology 9, no. 7: 177. https://doi.org/10.3390/biology9070177