
Ecology of Saline Watersheds: An Investigation of the Functional Communities and Drivers of Benthic Fauna in Typical Water Bodies of the Irtysh River Basin
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
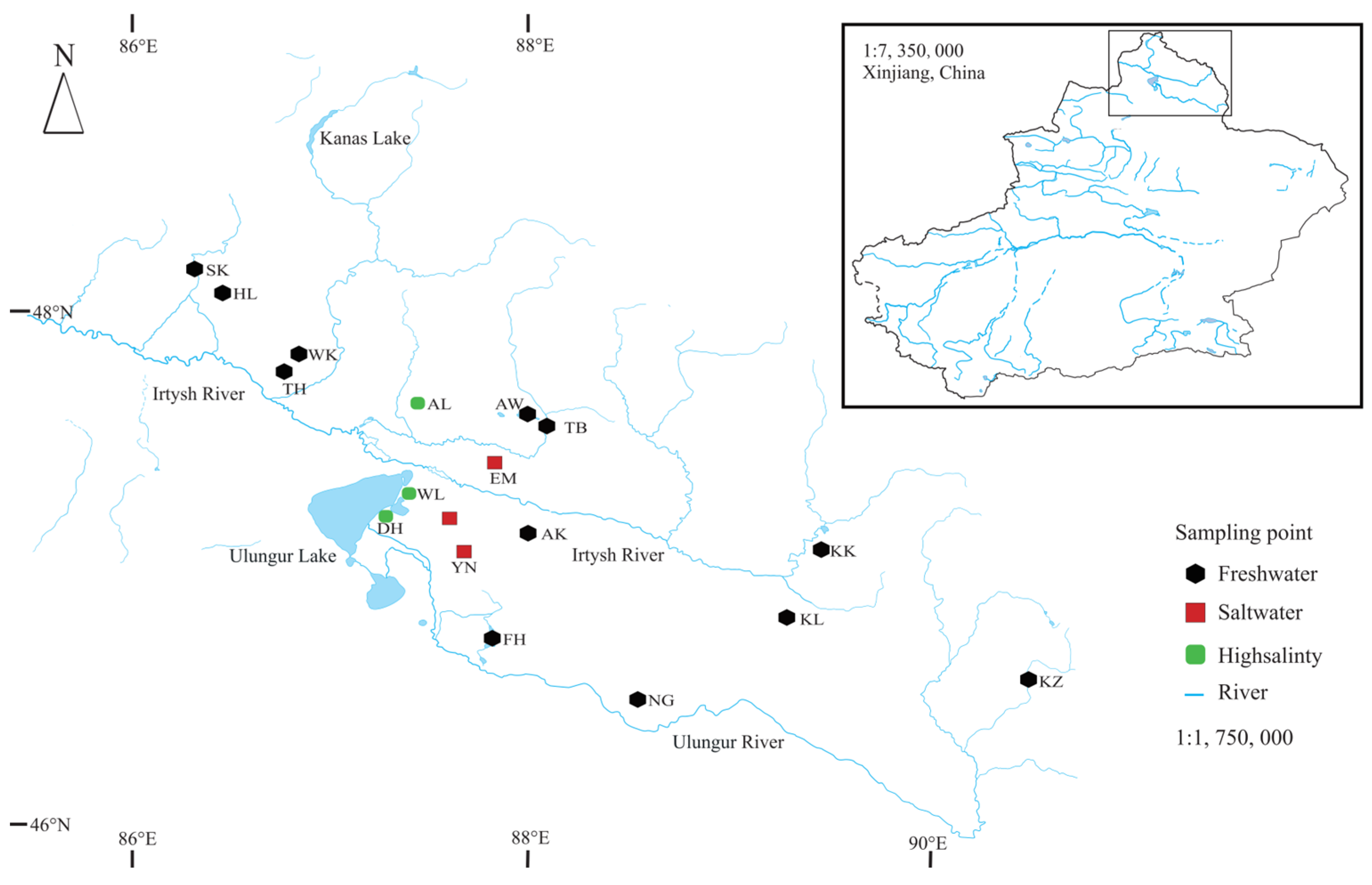
2.1. Field Sampling and Data Acquisition
2.2. Data Analysis
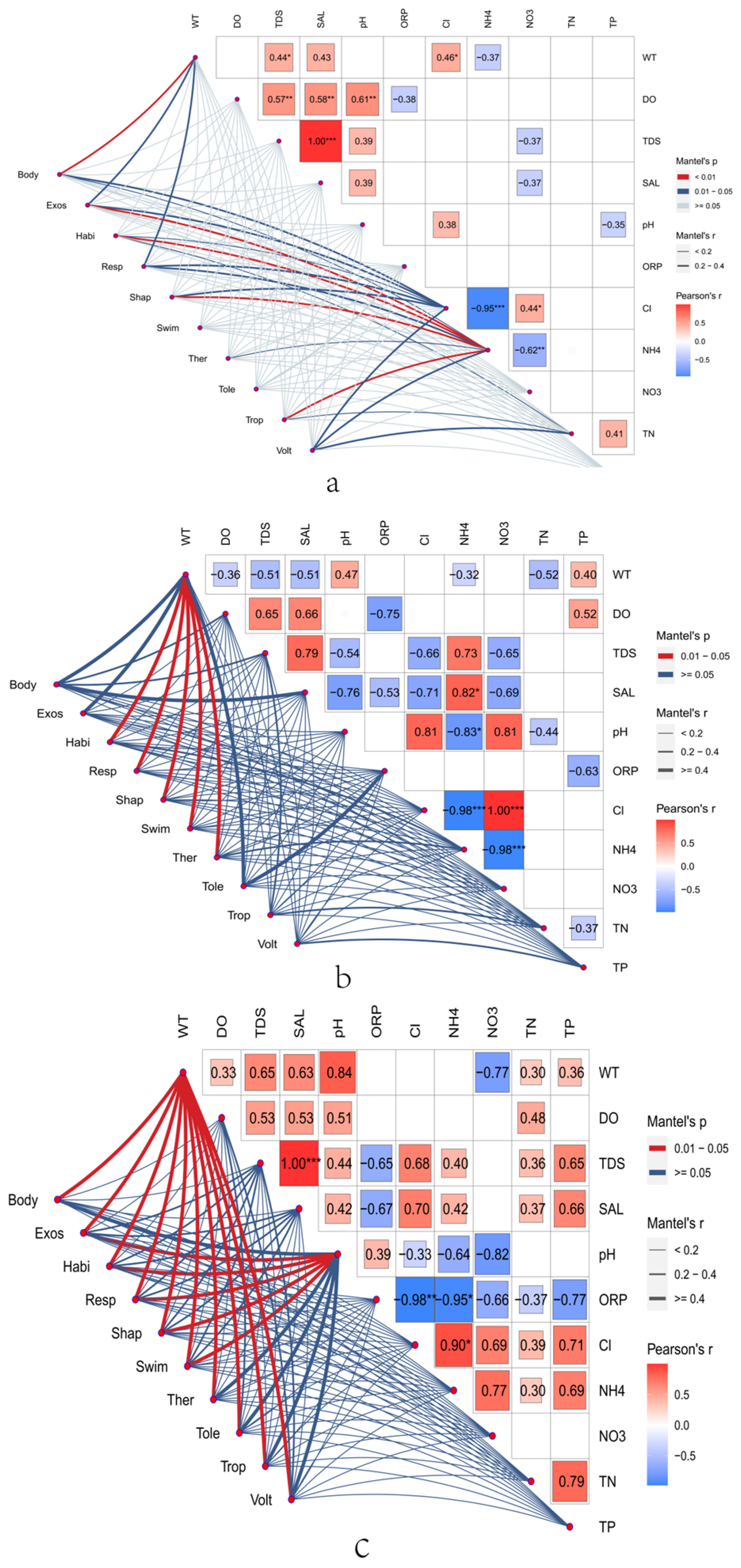
2.2.1. Correlation Analysis between Functional Traits and Environmental Factors of Benthic Organisms
2.2.2. Biodiversity
2.2.3. Functional Diversity
3. Results
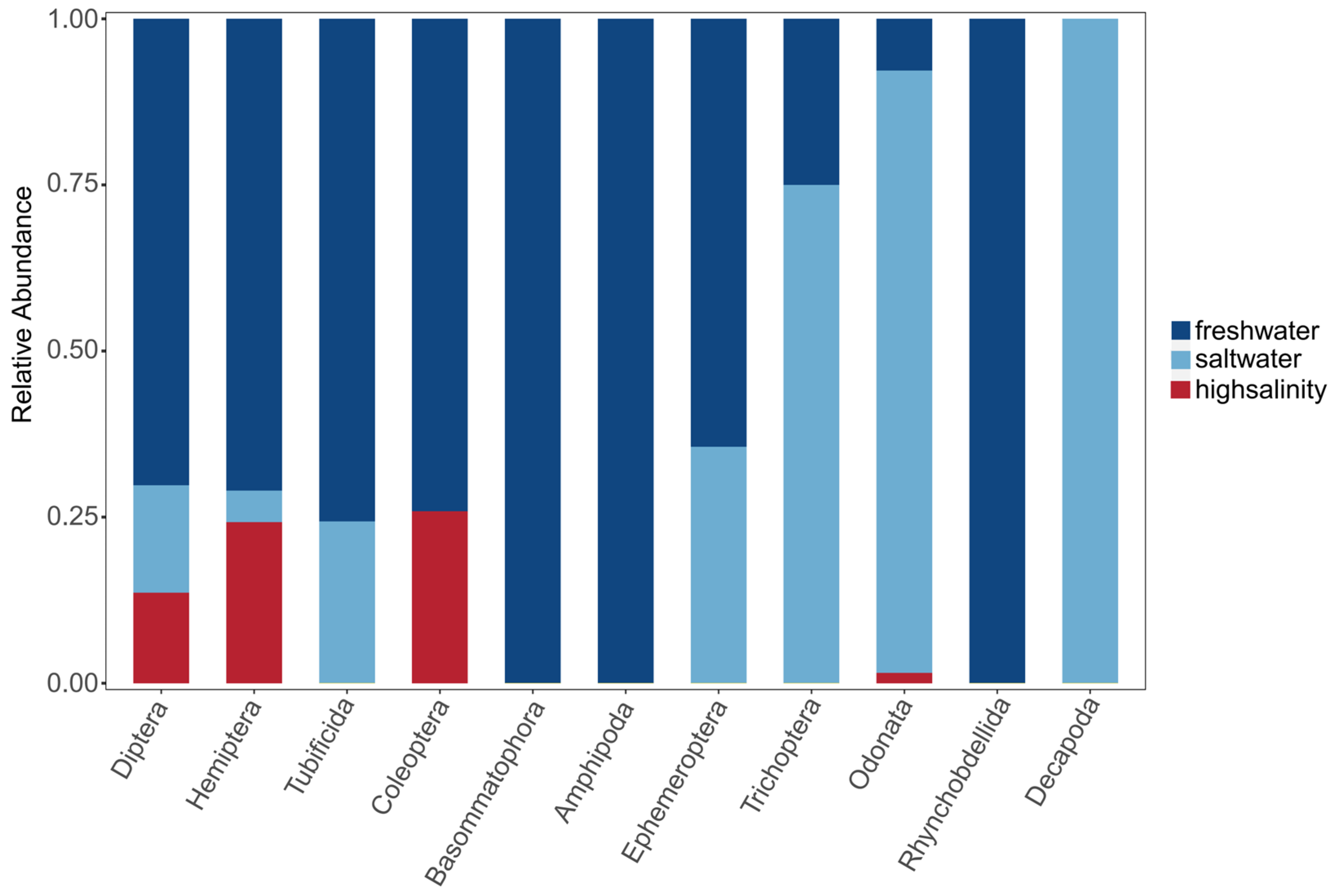
3.1. Characterization and Distribution
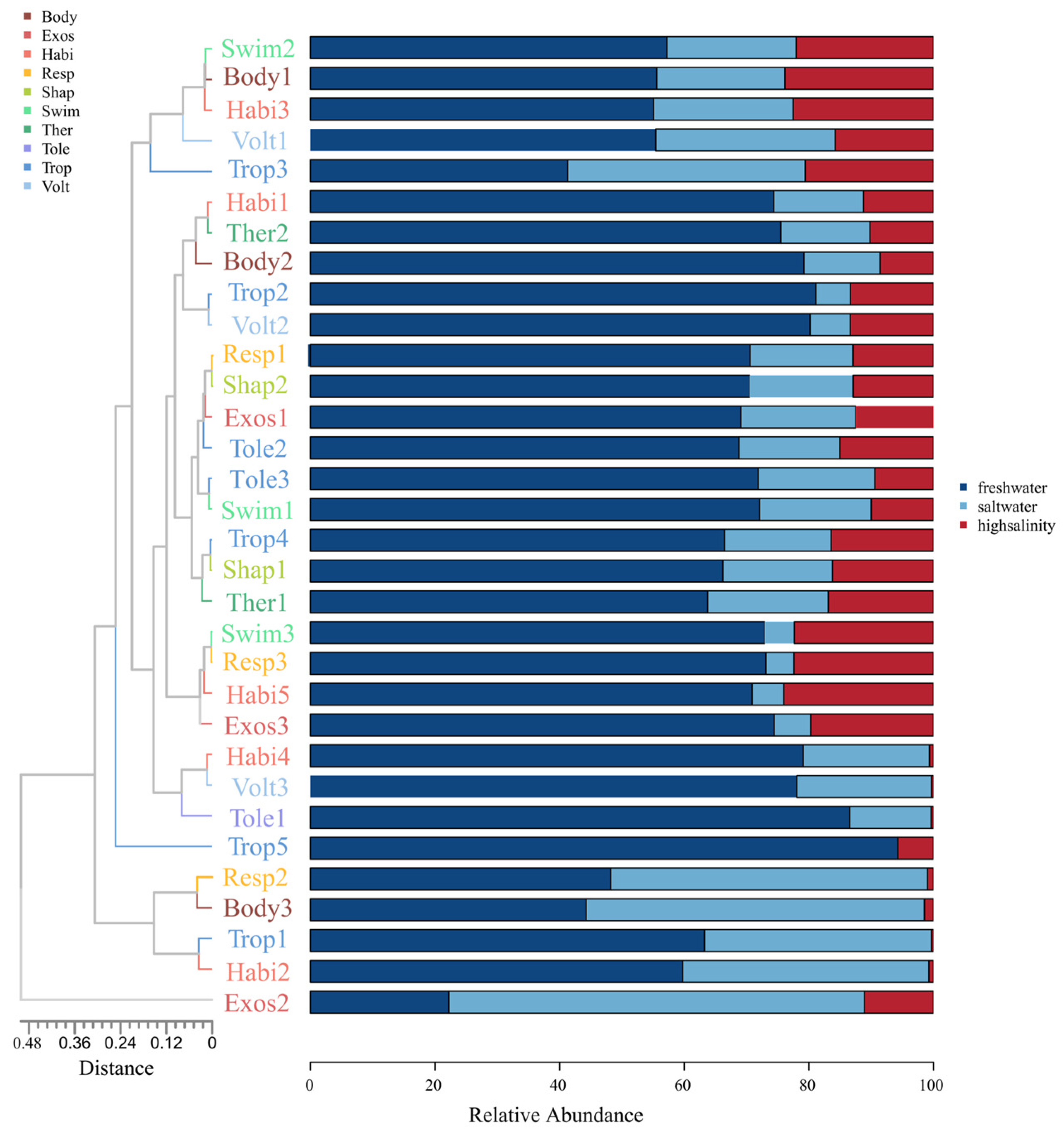
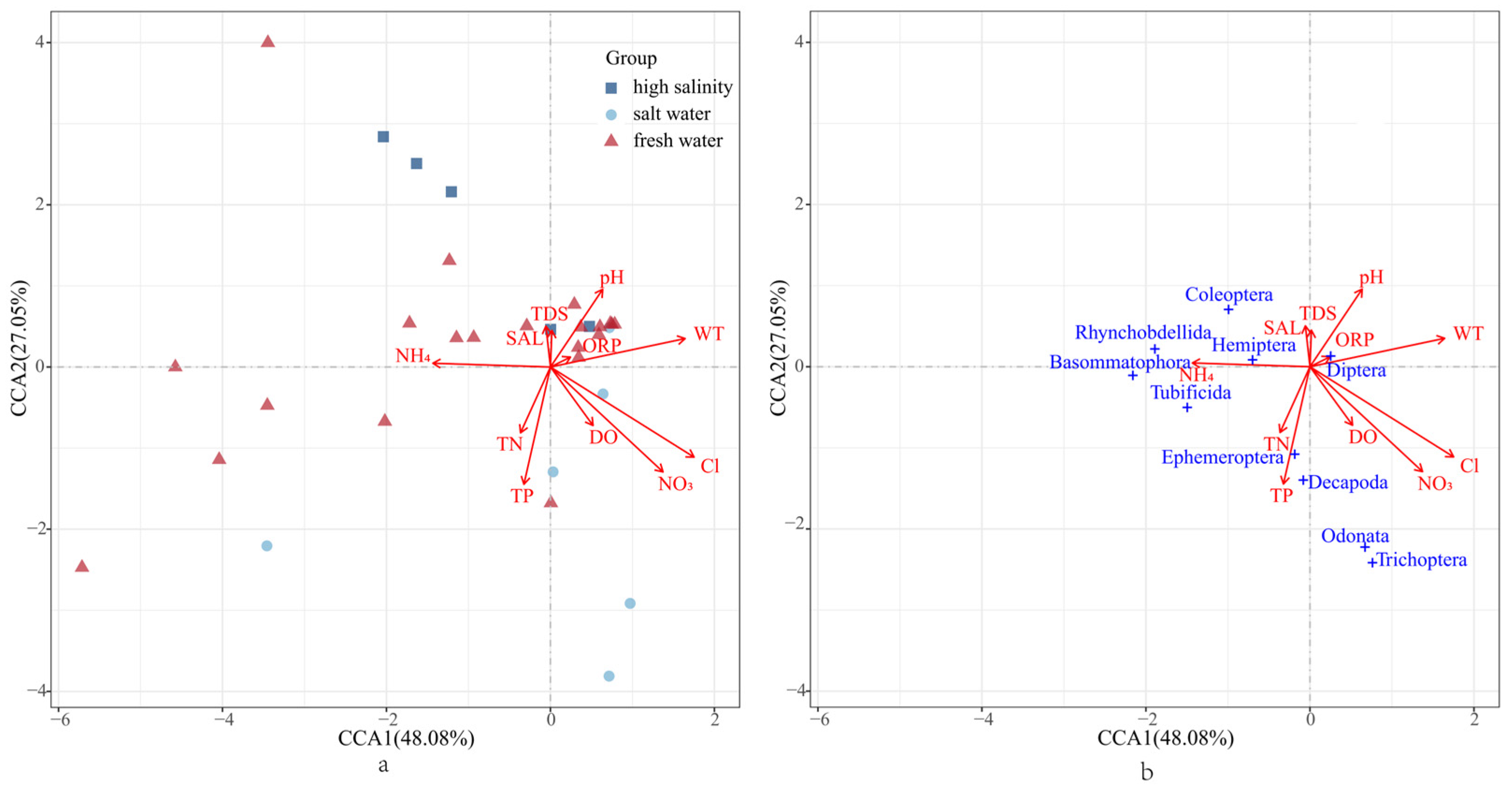
3.2. Environmental Factors and Functional Traits
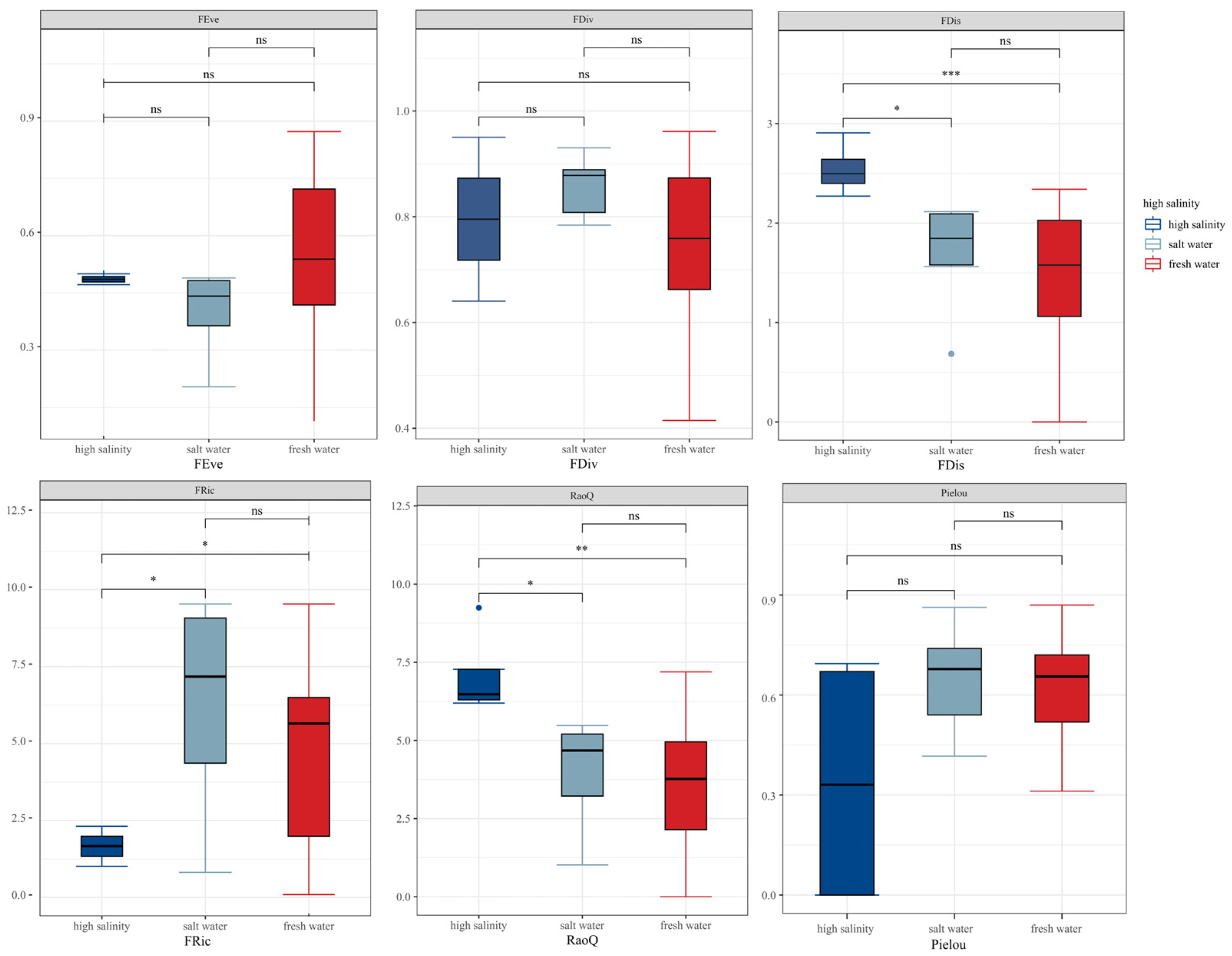
3.3. Correlations and Comparisons of Species and Functional Diversity
4. Discussion
4.1. Mechanisms of the Salinity Stress Response in Benthic Organisms
4.2. Influence of Abiotic Factors on Zonal Distribution Characteristics
4.3. Relationship between Biodiversity and Functional Diversity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aufdenkampe, A.K.; Mayorga, E.; Raymond, P.A.; Melack, J.M.; Doney, S.C.; Alin, S.R.; Aalto, R.E.; Yoo, K. Riverine coupling of biogeochemical cycles between land, oceans, and atmosphere. Front. Ecol. Environ. 2011, 9, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Raymond, P.A.; Hartmann, J.; Lauerwald, R.; Sobek, S.; McDonald, C.; Hoover, M.; Butman, D.; Striegl, R.; Mayorga, E.; Humborg, C.; et al. Global carbon dioxide emissions from inland waters. Nature 2013, 503, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.Y.; Liu, C.Q.; Hu, Y.; Shao, K.Q.; Tang, X.M.; Zhang, L.; Gao, G.; Qin, B.Q. Climate-induced salinization may lead to increased lake nitrogen retention. Water Res. 2023, 228, 119354. [Google Scholar] [CrossRef] [PubMed]
- Chai, H.X.; Cheng, W.M.; Zhou, C.H.; Zhao, S.M.; Liu, H.J. Climate effects on an inland alpine lake in Xinjiang, China over the past 40 years. J. Arid Land 2013, 5, 188–198. [Google Scholar] [CrossRef]
- Bethke, K.; Kropidlowska, K.; Stepnowski, P.; Caban, M. Review of warming and acidification effects to the ecotoxicity of pharmaceuticals on aquatic organisms in the era of climate change. Sci. Total Environ. 2023, 877, 162829. [Google Scholar] [CrossRef]
- Magee, M.R.; Hein, C.L.; Walsh, J.R.; Shannon, P.D.; Vander Zanden, M.J.; Campbell, T.B.; Hansen, G.J.A.; Hauxwell, J.; LaLiberte, G.D.; Parks, T.P.; et al. Scientific advances and adaptation strategies for Wisconsin lakes facing climate change. Lake Reserv. Manag. 2019, 35, 364–381. [Google Scholar] [CrossRef]
- Bonacina, L.; Fasano, F.; Mezzanotte, V.; Fornaroli, R. Effects of water temperature on freshwater macroinvertebrates: A systematic review. Biol. Rev. 2023, 98, 191–221. [Google Scholar] [CrossRef]
- Yin, J.Y.; Xia, J.H.; Xia, Z.C.; Cai, W.W.; Liu, Z.W.; Xu, K.J.; Wang, Y.; Zhang, R.Z.; Dong, X. Temporal Variation and Spatial Distribution in the Water Environment Helps Explain Seasonal Dynamics of Zooplankton in River-Type Reservoir. Sustainability 2022, 14, 13719. [Google Scholar] [CrossRef]
- Rezende, R.S.; Santos, A.M.; Henke-Oliveira, C.; Gonçalves, J.F. Effects of spatial and environmental factors on benthic a macroinvertebrate community. Zoologia 2014, 31, 426–434. [Google Scholar] [CrossRef]
- Jesus, F.; Pereira, J.L.; Campos, I.; Santos, M.; Ré, A.; Keizer, J.; Nogueira, A.; Gonçalves, F.J.M.; Abrantes, N.; Serpa, D. A review on polycyclic aromatic hydrocarbons distribution in freshwater ecosystems and their toxicity to benthic fauna. Sci. Total Environ. 2022, 820, 153282. [Google Scholar] [CrossRef]
- Gao, J.; Zhi, Y.; Huang, Y.Y.; Shi, S.J.; Tan, Q.J.; Wang, C.C.; Han, L.; Yao, J.M. Effects of benthic bioturbation on anammox in nitrogen removal at the sediment-water interface in eutrophic surface waters. Water Res. 2023, 243, 120287. [Google Scholar] [CrossRef] [PubMed]
- Hao, A.M.; Kobayashi, S.; Chen, F.B.; Yan, Z.X.; Torii, T.; Zhao, M.; Iseri, Y. Exploring invertebrate indicators of ecosystem health by focusing on the flow transitional zones in a large, shallow eutrophic lake. Environ. Sci. Pollut. Res. 2023, 30, 82717–82731. [Google Scholar] [CrossRef] [PubMed]
- Li, X.F.; Li, Y.; Gao, D.Z.; Liu, M.; Hou, L.J. Methane Production Linked to Organic Matter Molecule and Methanogenic Community in Estuarine Benthic Sediments. J. Geophys. Res.-Biogeosci. 2022, 127, e2022JG007236. [Google Scholar] [CrossRef]
- Lorrain-Soligon, L.; Perisse, L.; Robin, F.; Jankovic, M.; Brischoux, F. The costs of living on the coast: Reduction in body size and size-specific reproductive output in coastal populations of a widespread amphibian. Funct. Ecol. 2023, 37, 2703–2717. [Google Scholar] [CrossRef]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévéque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef]
- Arnott, S.E.; Fugère, V.; Symons, C.C.; Melles, S.J.; Beisner, B.E.; Cañedo-Argüelles, M.; Hébert, M.P.; Brentrup, J.A.; Downing, A.L.; Gray, D.K.; et al. Widespread variation in salt tolerance within freshwater zooplankton species reduces the predictability of community-level salt tolerance. Limnol. Oceanogr. Lett. 2023, 8, 8–18. [Google Scholar] [CrossRef]
- Timms, B.V. On the influence of season and salinity on the phenology of invertebrates in Australian saline lakes, with special reference to those of the Paroo in the semiarid inland. J. Oceanol. Limnol. 2018, 36, 1907–1916. [Google Scholar] [CrossRef]
- Lawrie, A.D.; Chaplin, J.; Pinder, A. Biology and conservation of the unique and diverse halophilic macroinvertebrates of Australian salt lakes. Mar. Freshw. Res. 2021, 72, 1553–1576. [Google Scholar] [CrossRef]
- Timms, B.V. Drivers restricting biodiversity in Australian saline lakes: A review. Mar. Freshw. Res. 2021, 72, 462–468. [Google Scholar] [CrossRef]
- Remane, A.; Schlieper, C. Biology of Brackish Water; Wiley & Sons: Hoboken, NJ, USA, 1972. [Google Scholar]
- Tang, Q.H.; Liu, X.C.; Zhou, Y.Y.; Wang, P.Y.; Li, Z.Q.; Hao, Z.X.; Liu, S.X.; Zhao, G.; Zhu, B.Q.; He, X.L.; et al. Climate change and water security in the northern slope of the Tianshan Mountains. Geogr. Sustain. 2022, 3, 246–257. [Google Scholar] [CrossRef]
- Yue, L.W.; Zan, F.Q.; Liu, X.G.; Yuan, Q.Q.; Shen, H.F. The Spatio-Temporal Reconstruction of Lake Water Levels Using Deep Learning Models: A Case Study on Altai Mountains. IEEE J. Sel. Top. Appl. Earth Observ. Remote Sens. 2022, 15, 4919–4940. [Google Scholar] [CrossRef]
- Liu, Y.T.; Yang, J.; Chen, Y.N.; Fang, G.H.; Li, W.H. The Temporal and Spatial Variations in Lake Surface Areas in Xinjiang, China. Water 2018, 10, 431. [Google Scholar] [CrossRef]
- Gao, J.Q.; Yu, Y.; Wang, D.H.; Liu, L.J.; Wang, W.; Dai, H.Z.; Cen, K. Hydrogeochemical characterization and water quality assessment in Altay, Xinjiang, northwest China. Environ. Monit. Assess. 2022, 194, 832. [Google Scholar] [CrossRef] [PubMed]
- Ran, M.; Zhang, C.J.; Feng, Z.D. Climatic and hydrological variations during the past 8000 years in northern Xinjiang of China and the associated mechanisms. Quat. Int. 2015, 358, 21–34. [Google Scholar] [CrossRef]
- Liu, H.Y.; Yin, Y.; Piao, S.L.; Zhao, F.J.; Engels, M.; Ciais, P. Disappearing Lakes in Semiarid Northern China: Drivers and Environmental Impact. Environ. Sci. Technol. 2013, 47, 12107–12114. [Google Scholar] [CrossRef]
- Li, Y.; Huang, X.Z.; Xiang, L.X.; Zhang, J.; Xu, Y.; Tian, L.H.; Jin, Y.X.; Jin, X.; Zhao, Y.T.; Ren, L.L.; et al. Holocene hydroclimatic change in the Altai Mountains and its impact on human migration. Catena 2022, 219, 106617. [Google Scholar] [CrossRef]
- Yang, X.K.; Lu, X.X. Drastic change in China’s lakes and reservoirs over the past decades. Sci. Rep. 2014, 4, 6041. [Google Scholar] [CrossRef] [PubMed]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef]
- Zhang, X.; Hao, Z.C.; Singh, V.P.; Zhang, Y.; Feng, S.F.; Xu, Y.; Hao, F.H. Drought propagation under global warming: Characteristics, approaches, processes, and controlling factors. Sci. Total Environ. 2022, 838, 156021. [Google Scholar] [CrossRef]
- Rivers, W.; Stribling, J.B.; Barbour, M.T.; Faulkner, C.; Gerritsen, J. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish; US Environmental Protection Agency, Office of Water: Washington, DC, USA, 1999.
- Bonada, N.; Dolédec, S. Does the Tachet trait database report voltinism variability of aquatic insects between Mediterranean and Scandinavian regions? Aquat. Sci. 2017, 80, 7. [Google Scholar] [CrossRef]
- Li, X.X.; Yang, W.; Li, S.Z.; Sun, T.; Bai, J.H.; Pei, J.; Xie, T.; Cui, B.S. Asymmetric responses of spatial variation of different communities to a salinity gradient in coastal wetlands. Mar. Environ. Res. 2020, 158, 105008. [Google Scholar] [CrossRef] [PubMed]
- Josefson, A.B. Species Sorting of Benthic Invertebrates in a Salinity Gradient—Importance of Dispersal Limitation. PLoS ONE 2016, 11, 21. [Google Scholar] [CrossRef] [PubMed]
- Krug, P.J.; Shimer, E.; Rodriguez, V.A. Differential Tolerance and Seasonal Adaptation to Temperature and Salinity Stress at a Dynamic Range Boundary Between Estuarine Gastropods. Biol. Bull. 2021, 241, 105–122. [Google Scholar] [CrossRef] [PubMed]
- Josefson, A.B.; Göke, C. Disentangling the effects of dispersal and salinity on beta diversity in estuarine benthic invertebrate assemblages. J. Biogeogr. 2013, 40, 1000–1009. [Google Scholar] [CrossRef]
- Obolewski, K.; Glinska-Lewczuk, K.; Szymanska, M.; Mrozinska, N.; Bakowska, M.; Astel, A.; Lew, S.; Paturej, E. Patterns of salinity regime in coastal lakes based on structure of benthic invertebrates. PLoS ONE 2018, 13, e0207825. [Google Scholar] [CrossRef] [PubMed]
- Rodil, I.F.; Lucena-Moya, P.; Jokinen, H.; Ollus, V.; Wennhage, H.; Villnäs, A.; Norkko, A. The role of dispersal mode and habitat specialization for metacommunity structure of shallow beach invertebrates. PLoS ONE 2017, 12, e0172160. [Google Scholar] [CrossRef]
- Heino, J. Does dispersal ability affect the relative importance of environmental control and spatial structuring of littoral macroinvertebrate communities? Oecologia 2013, 171, 971–980. [Google Scholar] [CrossRef]
- De Bie, T.; De Meester, L.; Brendonck, L.; Martens, K.; Goddeeris, B.; Ercken, D.; Hampel, H.; Denys, L.; Vanhecke, L.; Van der Gucht, K.; et al. Body size and dispersal mode as key traits determining metacommunity structure of aquatic organisms. Ecol. Lett. 2012, 15, 740–747. [Google Scholar] [CrossRef]
- Soininen, J.; McDonald, R.; Hillebrand, H. The distance decay of similarity in ecological communities. Ecography 2007, 30, 3–12. [Google Scholar] [CrossRef]
- Feng, J.X.; Huang, Q.; Chen, H.; Guo, J.M.; Lin, G.H. Restoration of native mangrove wetlands can reverse diet shifts of benthic macrofauna caused by invasive cordgrass. J. Appl. Ecol. 2018, 55, 905–916. [Google Scholar] [CrossRef]
- Ford, H.; Garbutt, A.; Jones, L.; Jones, D.L. Grazing management in saltmarsh ecosystems drives invertebrate diversity, abundance and functional group structure. Insect. Conserv. Divers. 2013, 6, 189–200. [Google Scholar] [CrossRef]
- Pétillon, J.; Lambeets, K.; Ract-Madoux, B.; Vernon, P.; Renault, D. Saline stress tolerance partly matches with habitat preference in ground-living wolf spiders. Physiol. Entomol. 2011, 36, 165–172. [Google Scholar] [CrossRef]
- Mahaney, W.C.; Hart, K.M.; O’Reilly, S.S.; Allen, C.C.R.; Dohm, J.M.; Hancock, R.G.V.; Kelleher, B.P.; Milner, M.W. Coleoptera and microbe biomass in Antarctic Dry Valley paleosols adjacent to the Inland Ice: Implications for Mars. Planet Space Sci. 2012, 60, 386–398. [Google Scholar] [CrossRef]
- Amri, C.; Ouchtati, N. Family Carabidae (Coleoptera) in an Algerian salt wetland: Taxonomic diversity, functional traits and effect of environmental drivers on their structure and composition. Orient. Insects 2023, 57, 1061–1083. [Google Scholar] [CrossRef]
- Ersoy, Z.; Abril, M.; Cañedo-Argüelles, M.; Espinosa, C.; Vendrell-Puigmitja, L.; Proia, L. Experimental assessment of salinization effects on freshwater zooplankton communities and their trophic interactions under eutrophic conditions. Environ. Pollut. 2022, 313, 120127. [Google Scholar] [CrossRef] [PubMed]
- Lorrain-Soligon, L.; Robin, F.; Lelong, V.; Jankovic, M.; Barbraud, C.; Brischoux, F. Distance to coastline modulates morphology and population structure in a coastal amphibian. Biol. J. Linn. Soc. 2022, 135, 478–489. [Google Scholar] [CrossRef]
- Nebra, A.; Alcaraz, C.; Caiola, N.; Muñoz-Camarillo, G.; Ibáñez, C. Benthic macrofaunal dynamics and environmental stress across a salt wedge Mediterranean estuary. Mar. Environ. Res. 2016, 117, 21–31. [Google Scholar] [CrossRef]
- Lin, M.W.; Kuan, P.L.; Sung, P.H.; Tseng, Y.C. Metabolic trade-offs associated with homeostatic adjustments in pelagic and benthic cephalopods: Comparative evaluations of NH4+/H+ transport machinery in gills. Front. Mar. Sci. 2022, 9, 971764. [Google Scholar] [CrossRef]
- Fiore, C.L.; Baker, D.M.; Lesser, M.P. Nitrogen Biogeochemistry in the Caribbean Sponge, Xestospongia muta: A Source or Sink of Dissolved Inorganic Nitrogen? PLoS ONE 2013, 8, e0072961. [Google Scholar] [CrossRef]
- Wollheim, W.M.; Vöösmarty, C.J.; Peterson, B.J.; Seitzinger, S.P.; Hopkinson, C.S. Relationship between river size and nutrient removal. Geophys. Res. Lett. 2006, 33, 6410-1–6410-4. [Google Scholar] [CrossRef]
- Wang, S.Y.; Zhang, P.; Zhang, D.T.; Chang, J.B. Evaluation and comparison of the benthic and microbial indices of biotic integrity for urban lakes based on environmental DNA and its management implications. J. Environ. Manag. 2023, 341, 118026. [Google Scholar] [CrossRef] [PubMed]
- Zilius, M.; Daunys, D.; Bartoli, M.; Marzocchi, U.; Bonaglia, S.; Cardini, U.; Castaldelli, G. Partitioning benthic nitrogen cycle processes among three common macrofauna holobionts. Biogeochemistry 2022, 157, 193–213. [Google Scholar] [CrossRef]
- Politi, T.; Bariseviciute, R.; Bartoli, M.; Bonaglia, S.; Cardini, U.; Castaldelli, G.; Kancauskaite, A.; Marzocchi, U.; Petkuviene, J.; Samuiloviene, A.; et al. A bioturbator, a holobiont, and a vector: The multifaceted role of Chironomus plumosus in shaping N-cycling. Freshw. Biol. 2021, 66, 1036–1048. [Google Scholar] [CrossRef]
- Telesh, I.V.; Khlebovich, V.V. Principal processes within the estuarine salinity gradient: A review. Mar. Pollut. Bull. 2010, 61, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Paz, L.E.; Rodriguez, M.; Gullo, B.; Capítulo, A.R. Impacts of urban and industrial pollution on functional traits of benthic macroinvertebrates: Are some traits advantageous for survival? Sci. Total Environ. 2022, 807, 150650. [Google Scholar] [CrossRef]
- Reizopoulou, S.; Nicolaidou, A. Benthic diversity of coastal brackish-water lagoons in western Greece. Aquat. Conserv. Mar. Freshw. Ecosyst. 2004, 14, S93–S102. [Google Scholar] [CrossRef]
- Suurkuukka, H.; Meissner, K.K.; Muotka, T. Species turnover in lake littorals: Spatial and temporal variation of benthic macroinvertebrate diversity and community composition. Divers. Distrib. 2012, 18, 931–941. [Google Scholar] [CrossRef]
- Fontana, G.; Ugland, K.I.; Gray, J.S.; Willis, T.J.; Abbiati, M. Influence of rare species on beta diversity estimates in marine benthic assemblages. J. Exp. Mar. Biol. Ecol. 2008, 366, 104–108. [Google Scholar] [CrossRef]
- Maciolek, N.J.; Smith, W.K. Benthic species diversity along a depth gradient: Boston Harbor to Lydonia Canyon. Deep-SEA Res. Part II Top. Stud. Oceanogr. 2009, 56, 1763–1774. [Google Scholar] [CrossRef]
- Josefson, A.B. Additive partitioning of estuarine benthic macroinvertebrate diversity across multiple spatial scales. Mar. Ecol. Prog. Ser. 2009, 396, 283–292. [Google Scholar] [CrossRef]
- Bian, L.; Che, X.Q. Analysis of the correlation between benthic species community and ecosystem diversity factors in the south china sea. Fresenius Environ. Bull. 2022, 31, 3727–3734. [Google Scholar]
- Marin, I.N.; Matsyura, A.V. Taxonomic diversity of benthic invertebrates of the Moskva River in the Moscow area: Diversity and distribution. Ukr. J. Ecol. 2017, 7, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lu, X.; Liu, X.S. Patterns of species and functional diversity of macrofaunal assemblages and the bioassessment of benthic ecological quality status in the southern Yellow Sea. Mar. Pollut. Bull. 2021, 171, 112784. [Google Scholar] [CrossRef] [PubMed]
- Koutsoubas, D.; Dounas, C.; Arvanitidis, C.; Kornilios, S.; Petihakis, G.; Triantafyllou, G.; Eleftheriou, A. Macrobenthic community structure and disturbance assessment in Gialova Lagoon, Ionian Sea. ICES J. Mar. Sci. 2000, 57, 1472–1480. [Google Scholar] [CrossRef]
- Bray, L.; Faulwetter, S.; Kaberi, H.; Kastanidi, E.; Karageorgis, A.P.; Katsiaras, N.; Pavlidou, A.; Providakis, N.; Sigala, K.; Voutsinas, E.; et al. Assessing pressure drivers on the benthic ecosystem in the coastal zone of Western Messinia, Greece. Estuar. Coast. Shelf Sci. 2022, 274, 107935. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Trait State | Code | Score |
|---|---|---|---|
| Voltinism | Semivoltine | Volt1 | 1 |
| Univoltine | Volt2 | 2 | |
| Bior multivoltine | Volt3 | 3 | |
| Body size | Small (<9 mm) | Body1 | 1 |
| Medium (9–16 mm) | Body2 | 2 | |
| Large (>16 mm) | Body3 | 3 | |
| Exoskeleton or external protection | Soft-bodied forms | Exos1 | 1 |
| Lightly sclerotized | Exos2 | 2 | |
| Heavily sclerotized | Exos3 | 3 | |
| Shape | Streamlined | Shap1 | 1 |
| Not streamlined | Shap2 | 2 | |
| Respiration | Tegument | Resp1 | 1 |
| Gills | Resp2 | 2 | |
| Air (spiracles, tracheae, plastrons) | Resp3 | 3 | |
| Swimming ability | None | Swim1 | 1 |
| Weak | Swim2 | 2 | |
| Strong | Swim3 | 3 | |
| Thermal preference | Cool eurythermal | Ther1 | 1 |
| Cool/warm eurythermal | Ther2 | 2 | |
| Habit | Burrower | Habi1 | 1 |
| Climber | Habi2 | 2 | |
| Sprawler | Habi3 | 3 | |
| Clinger | Habi4 | 4 | |
| Swimmer | Habi5 | 5 | |
| Trophic habit | Collector-gatherer | Trop1 | 1 |
| Collector-filterer | Trop2 | 2 | |
| Scraper | Trop3 | 3 | |
| Predator | Trop4 | 4 | |
| Shredder | Trop5 | 5 | |
| Pollution resistance value | Weak | Tole1 | 1 |
| Moderate | Tole2 | 2 | |
| Strong | Tole3 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zi, F.; Wang, B.; Yang, L.; Huo, Q.; Wang, Z.; Ren, D.; Huo, B.; Song, Y.; Chen, S. Ecology of Saline Watersheds: An Investigation of the Functional Communities and Drivers of Benthic Fauna in Typical Water Bodies of the Irtysh River Basin. Biology 2024, 13, 27. https://doi.org/10.3390/biology13010027
Zi F, Wang B, Yang L, Huo Q, Wang Z, Ren D, Huo B, Song Y, Chen S. Ecology of Saline Watersheds: An Investigation of the Functional Communities and Drivers of Benthic Fauna in Typical Water Bodies of the Irtysh River Basin. Biology. 2024; 13(1):27. https://doi.org/10.3390/biology13010027
Chicago/Turabian StyleZi, Fangze, Baoqiang Wang, Liting Yang, Qiang Huo, Zhichao Wang, Daoquan Ren, Bin Huo, Yong Song, and Shengao Chen. 2024. "Ecology of Saline Watersheds: An Investigation of the Functional Communities and Drivers of Benthic Fauna in Typical Water Bodies of the Irtysh River Basin" Biology 13, no. 1: 27. https://doi.org/10.3390/biology13010027